The Effects of Carbohydrates, in Isolation and Combined with Caffeine, on Cognitive Performance and Mood—Current Evidence and Future Directions


Abstract
1. Introduction
2. Carbohydrates and Cognitive Function
2.1. Glucose
2.2. Factors Moderating the Effect of Glucose
2.2.1. The Effect of Dose
2.2.2. The Effect of Age
2.2.3. The Effect of Task Demand
2.2.4. The Effect of Glucoregulatory Control
2.2.5. Emotional Valence
2.2.6. Expectancy Effects
2.3. Glucose and Subjective Mood
2.4. Other Carbohydrates and Cognitive Function
2.4.1. Fructose
2.4.2. Sucrose
2.4.3. Isomaltulose
2.5. Summary and Unanswered Questions
3. Glycemic Response and Cognitive Performance
3.1. Manipulating Glycaemic Response
3.2. Moderation of Glycaemic Response by Vehicle
3.3. Summary and Unanswered Questions
4. Caffeine, Carbohydrates, and Cognitive Function
4.1. Caffeine
4.2. Combined Effects of Caffeine and CHO
4.2.1. Cognitive Performance Outcomes
4.2.2. Subjective Outcomes
4.3. Interactive Effects of Caffeine and CHO
4.3.1. Cognitive Performance Outcomes
4.3.2. Subjective Outcomes
4.4. Summary of Interactive Effects
4.5. Dose Effects
4.5.1. Cognitive Performance Outcomes
4.5.2. Subjective Outcomes
4.6. Mechanisms of Action
4.7. Unanswered Questions and Recommendations for Future Research
4.7.1. Interactive Effects
4.7.2. Timescale of Effect
4.7.3. Vehicle of Administration
4.7.4. Effects on Subjective States
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hafermann, G. Schulmüdigkeit und Blutzuckerverhalten. Öffentl. Gesundh. 1955, 17, 11–17. [Google Scholar]
- Bellisle, F.; Blundell, J.E.; Dye, L.; Fantino, M.; Fern, E.; Fletcher, R.J.; Lambert, J.; Roberfoid, M.; Specter, S.; Westenhofer, J.; et al. Functional food science and behaviour and psychological functions. Br. J. Nutr. 1998, 80, S173–S193. [Google Scholar] [CrossRef] [PubMed]
- Dye, L.; Blundell, J. Functional foods: Psychological and behavioural functions. Br. J. Nutr. 2002, 88, S187–S211. [Google Scholar] [CrossRef] [PubMed]
- Sünram-Lea, S.-I.; Owen, L.; Robertson, B. Glycaemic control and cognition: Evidence across the lifespan. In Nutrition for Brain Health and Cognitive Performance; Dye, T.B.L., Ed.; CRC Press-Taylor & Francis Group: Boca Raton, FL, USA, 2015; pp. 125–150. [Google Scholar]
- Sieber, F.E.; Traystman, R.J. Special issues—Glucose and the brain. Crit. Care Med. 1992, 20, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Foster, J.K. The impact of a high versus a low glycaemic index breakfast cereal meal on verbal episodic memory in healthy adolescents. Nutr. Neurosci. 2008, 11, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.K.; Lidder, P.G.; Sunram, S.I. Glucose and memory: Fractionation of enhancement effects? Psychopharmacology (Berlin) 1998, 137, 259–270. [Google Scholar] [CrossRef]
- Riby, L.M.; Law, A.S.; McLaughlin, J.; Murray, J. Preliminary evidence that glucose ingestion facilitates prospective memory performance. Nutr. Res. (N. Y.) 2011, 31, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Riby, L.M.; Meikle, A.; Glover, C. The effects of age, glucose ingestion and gluco-regulatory control on episodic memory. Age Ageing 2004, 33, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Riby, L.M.; Marriott, A.; Bullock, R.; Hancock, J.; Smallwood, J.; McLaughlin, J. The effects of glucose ingestion and glucose regulation on memory performance in older adults with mild cognitive impairment. Eur. J. Clin. Nutr. 2009, 63, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Manning, C.A.; Ragozzino, M.E.; Gold, P.E. Glucose enhancement of memory in patients with probable senile dementia of the alzheimers type. Neurobiol. Aging 1993, 14, 523–528. [Google Scholar] [CrossRef]
- Brandt, K.R. Effects of glucose administration on category exclusion recognition. J. Psychopharmacol. 2015, 29, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Owen, L.; Scholey, A.; Finnegan, Y.; Suenram-Lea, S.I. Response variability to glucose facilitation of cognitive enhancement. Br. J. Nutr. 2013, 110, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Sunram-Lea, S.; Brandt, K. Effect of glucose ingestion on recognition memory for emotionally arousing stimuli. Int. J. Psychol. 2008, 43, 224. [Google Scholar]
- Sunram-Lea, S.I.; Foster, J.K.; Durlach, P.; Perez, C. Investigation into the significance of task difficulty and divided allocation of resources on the glucose memory facilitation effect. Psychopharmacology (Berlin) 2002, 160, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Sunram-Lea, S.I.; Owen, L.; Finnegan, Y.; Hu, H.L. Dose-response investigation into glucose facilitation of memory performance and mood in healthy young adults. J. Psychopharmacol. 2011, 25, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Sunram-Lea, S.I.; Foster, J.K.; Durlach, P.; Perez, C. Glucose facilitation of cognitive performance in healthy young adults: Examination of the influence of fast-duration, time of day and pre-consumption plasma glucose levels. Psychopharmacology (Berlin) 2001, 157, 46–54. [Google Scholar] [CrossRef]
- Sunram-Lea, S.I.; Foster, J.K.; Durlach, P.; Perez, C. The effect of retrograde and anterograde glucose administration on memory performance in healthy young adults. Behav. Brain Res. 134, 505–516. [CrossRef]
- Mohanty, A.; Flint, R.W., Jr. Differential effects of glucose on modulation of emotional and nonemotional spatial memory tasks. Cogn. Affect. Behav. Neurosci. 2001, 1, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.B.; Fowles, K.A. Retrograde enhancement of kinesthetic memory by alcohol and by glucose. Neurobiol. Learn. Mem. 2002, 78, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.S.; Benton, D. The impact of raising blood-glucose on reaction-times. Neuropsychobiology 1994, 30, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L.; Gonderfrederick, L.A.; Chewning, W.W.; Silveira, J.; Gold, P.E. Glucose enhancement of performance on memory tests in young and aged humans. Neuropsychologia 1989, 27, 1129–1138. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Scholey, A.B. Glucose administration, heart rate and cognitive performance: Effects of increasing mental effort. Psychopharmacology (Berlin) 2000, 149, 63–71. [Google Scholar] [CrossRef]
- Miller, H.C.; Bourrasseau, C.; Blampain, J. Can you enhance executive control without glucose? The effects of fructose on problem solving. J. Psychopharmacol. 2013, 27, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.K.; Sunram-Lea, S.I.; Wesnes, K.A. Acute ingestion of different macronutrients differentially enhances aspects of memory and attention in healthy young adults. Biol. Psychol. 2012, 89, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Meikle, A.; Riby, L.M.; Stollery, B. The impact of glucose ingestion and gluco-regulatory control on cognitive performance: A comparison of younger and middle aged adults. Hum. Psychopharmacol.-Clin. Exp. 2004, 19, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Reay, J.L.; Kennedy, D.O.; Scholey, A.B. Effects of panax ginseng, consumed with and without glucose, on blood glucose levels and cognitive performance during sustained ‘mentally demanding’ tasks. J. Psychopharmacol. 2006, 20, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Stollery, B.; Christian, L. Glucose and memory: The influence of drink, expectancy, and beliefs. Psychopharmacology (Berlin) 2013, 228, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Azari, N.P. Effects of glucose on memory processes in young-adults. Psychopharmacology (Berlin) 1991, 105, 521–524. [Google Scholar] [CrossRef]
- Benton, D.; Owens, D.S. Blood-glucose and human-memory. Psychopharmacology (Berlin) 1993, 113, 83–88. [Google Scholar] [CrossRef]
- Manning, C.A.; Parsons, M.W.; Cotter, E.M.; Gold, P.E. Glucose effects on declarative and nondeclarative memory in healthy elderly and young adults. Psychobiology 1997, 25, 103–108. [Google Scholar]
- Winder, R.; Borrill, J. Fuels for memory: The role of oxygen and glucose in memory enhancement. Psychopharmacology (Berlin) 1998, 136, 349–356. [Google Scholar] [CrossRef]
- Benton, D. The impact of increasing blood-glucose on psychological functioning. Biol. Psychol. 1990, 30, 13–19. [Google Scholar] [CrossRef]
- Craft, S.; Murphy, C.; Wemstrom, J. Glucose effects on complex memory and nonmemory tasks—The influence of age, sex, and glucoregulatory response. Psychobiology 1994, 22, 95–105. [Google Scholar]
- Benton, D.; Owens, D.S.; Parker, P.Y. Blood-glucose influences memory and attention in young-adults. Neuropsychologia 1994, 32, 595–607. [Google Scholar] [CrossRef]
- Parker, P.Y.; Benton, D. Blood-glucose levels selectively influence memory for word lists dichotically presented to the right ear. Neuropsychologia 1995, 33, 843–854. [Google Scholar] [CrossRef]
- Messier, C.; Pierre, J.; Desrochers, A.; Gravel, M. Dose-dependent action of glucose on memory processes in women: Effect on serial position and recall priority. Cogn. Brain Res. 1998, 7, 221–233. [Google Scholar] [CrossRef]
- Messier, C.; Desrochers, A.; Gagnon, M. Effect of glucose, glucose regulation, and word imagery value on human memory. Behav. Neurosci. 1999, 113, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, R.T.; Benton, D.C. Cognitive functioning is susceptible to the level of blood glucose. Psychopharmacology 1999, 145, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.M. Glucose enhancement of a facial recognition task in young adults. Physiol. Behav. 2000, 68, 549–553. [Google Scholar] [CrossRef]
- Green, M.W.; Taylor, M.A.; Elliman, N.A.; Rhodes, O. Placebo expectancy effects in the relationship between glucose and cognition. Br. J. Nutr. 2001, 86, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Morris, N.; Sarll, P. Drinking glucose improves listening span in students who miss breakfast. Educ. Res. 2001, 43, 201–207. [Google Scholar] [CrossRef]
- Scholey, A.B.; Harper, S.; Kennedy, D.O. Cognitive demand and blood glucose. Physiol. Behav. 2001, 73, 585–592. [Google Scholar] [CrossRef]
- Awad, N.; Gagnon, M.; Desrochers, A.; Tsiakas, M.; Messier, C. Impact of peripheral glucoregulation on memory. Behav. Neurosci. 2002, 116, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Ford, C.E.; Scholey, A.B.; Ayre, G.; Wesnes, K. The effect of glucose administration and the emotional content of words on heart rate and memory. J. Psychopharmacol. 2002, 16, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Flint, R.W.; Turek, C. Glucose effects on a continuous performance test of attention in adults. Behav. Brain Res. 2003, 142, 217–228. [Google Scholar] [CrossRef]
- Meikle, A.; Riby, L.M.; Stollery, B. Memory processing and the glucose facilitation effect: The effects of stimulus difficulty and memory load. Nutr. Neurosci. 2005, 8, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Riby, L.M.; McMurtrie, H.; Smallwood, J.; Ballantyne, C.; Meikle, A.; Smith, E. The facilitative effects of glucose ingestion on memory retrieval in younger and older adults: Is task difficulty or task domain critical? Br. J. Nutr. 2006, 95, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.R.; Sunram-Lea, S.I.; Qualtrough, K. The effect of glucose administration on the emotional enhancement effect in recognition memory. Biol. Psychol. 2006, 73, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Gailliot, M.T.; Baumeister, R.F. The physiology of willpower: Linking blood glucose to self-control. Pers. Soc. Psychol. Rev. 2007, 11, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Masicampo, E.J.; Baumeister, R.F. Toward a physiology of dual-process reasoning and judgment—Lemonade, willpower, and expensive rule-based analysis. Psychol. Sci. 2008, 19, 255–260. [Google Scholar] [CrossRef] [PubMed]
- DeWall, C.N.; Baumeister, R.F.; Gailliot, M.T.; Maner, J.K. Depletion makes the heart grow less helpful: Helping as a function of self-regulatory energy and genetic relatedness. Personal. Soc. Psychol. Bull. 2008, 34, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Morris, N. Elevating blood glucose level increases the retention of information from a public safety video. Biol. Psychol. 2008, 78, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Riby, L.M.; McLaughlin, J.; Riby, D.M. Lifestyle, glucose regulation and the cognitive effects of glucose load in middle-aged adults. Br. J. Nutr. 2008, 100, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.B.; Sunram-Lea, S.I.; Greer, J.; Elliott, J.; Kennedy, D.O. Glucose administration prior to a divided attention task improves tracking performance but not word recognition: Evidence against differential memory enhancement? Psychopharmacology (Berlin) 2009, 202, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.B.; Sunram-Lea, S.I.; Greer, J.; Elliott, J.; Kennedy, D.O. Glucose enhancement of memory depends on initial thirst. Appetite 2009, 53, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Owen, L.; Finnegan, Y.; Hu, H.; Scholey, A.B.; Suenram-Lea, S.I. Glucose effects on long-term memory performance: Duration and domain specificity. Psychopharmacology (Berlin) 2010, 211, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.R.; Sunram-Lea, S.I.; Jenkinson, P.M.; Jones, E. The effects of glucose dose and dual-task performance on memory for emotional material. Behav. Brain Res. 2010, 211, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Parent, M.B.; Krebs-Kraft, D.L.; Ryan, J.P.; Wilson, J.S.; Harenski, C.; Hamann, S. Glucose administration enhances fmri brain activation and connectivity related to episodic memory encoding for neutral and emotional stimuli. Neuropsychologia 2011, 49, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.; Hay, P.; Campbell, L.; Trollor, J.N. A review of the association between obesity and cognitive function across the lifespan: Implications for novel approaches to prevention and treatment. Obes. Rev. 2011, 12, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.R.; Gibson, E.L.; Rackie, J.M. Differential facilitative effects of glucose administration on stroop task conditions. Behav. Neurosci. 2013, 127, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.; Macpherson, H.; Suenram-Lea, S.; Elliott, J.; Stough, C.; Kennedy, D. Glucose enhancement of recognition memory: Differential effects on effortful processing but not aspects of ‘remember-know’ responses. Neuropharmacology 2013, 64, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.A.; Riby, L.M. Glucose enhancement of event-related potentials associated with episodic memory and attention. Food Funct. 2013, 4, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Lange, F.; Eggert, F. Sweet delusion. Glucose drinks fail to counteract ego depletion. Appetite 2014, 75, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Stollery, B.; Christian, L. Glucose, relational memory, and the hippocampus. Psychopharmacology (Berlin) 2015, 232, 2113–2125. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, H.; Roberstson, B.; Suenram-Lea, S.; Stough, C.; Kennedy, D.; Scholey, A. Glucose administration and cognitive function: Differential effects of age and effort during a dual task paradigm in younger and older adults. Psychopharmacology (Berlin) 2015, 232, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.E. Modulation of memory processing: Enhancement ofmemory in rodents and humans. In The Neuropsychology of Memory, 2nd ed.; Squire, L.R., Butters, N., Eds.; Guilford: New York, NY, USA, 1992. [Google Scholar]
- Gold, P.E.; Vogt, J.; Hall, J.L. Glucose effects on memory—Behavioral and pharmacological characteristics. Behav. Neural Biol. 1986, 46, 145–155. [Google Scholar] [CrossRef]
- Rodriguez, W.A.; Vanausdle, L.R.; Dhanens, K.; Mondragon, A.N. Glucose modulates recently reactivated memories. Psychobiology 1993, 21, 93–100. [Google Scholar]
- Stone, W.S.; Walser, B.; Gold, S.D.; Gold, P.E. Scopolamine-induced and morphine-induced impairments of spontaneous-alternation performance in mice—Reversal with glucose and with cholinergic and adrenergic agonists. Behav. Neurosci. 1991, 105, 264–271. [Google Scholar] [CrossRef] [PubMed]
- White, N.M. Peripheral and Central Memory—Enhancing Actions of Glucose; American Psychological Association: Washington, DC, USA, 1991; Volume 6, pp. 421–441. [Google Scholar]
- Lamport, D.J.; Lawton, C.L.; Mansfield, M.W.; Moulin, C.A.J.; Dye, L. Type 2 diabetes and impaired glucose tolerance are associated with word memory source monitoring recollection deficits but not simple recognition familiarity deficits following water, low glycaemic load, and high glycaemic load breakfasts. Physiol. Behav. 2014, 124, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.W.; Gold, P.E. Glucose enhancement of memory in elderly humans—An inverted-u dose-response curve. Neurobiol. Aging 1992, 13, 401–404. [Google Scholar] [CrossRef]
- Hoyland, A.; Lawton, C.L.; Dye, L. Acute effects of macronutrient manipulations on cognitive test performance in healthy young adults: A systematic research review. Neurosci. Biobehav. Rev. 2008, 32, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Messier, C. Glucose improvement of memory: A review. Eur. J. Pharmacol. 2004, 490, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Gold, S.M.; Dziobek, I.; Rogers, K.; Bayoumy, A.; McHugh, P.F.; Convit, A. Hypertension and hypothalamo-pituitary-adrenal axis hyperactivity affect frontal lobe integrity. J. Clin. Endocrinol. Metab. 2005, 90, 3262–3267. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Lindenberger, U.; Hommel, B.; Aschersleben, G.; Prinz, W.; Baltes, P.B. Transformations in the couplings among intellectual abilities and constituent cognitive processes across the life span. Psychol. Sci. 2004, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.; Parker, P.Y.; Donohue, R.T. The supply of glucose to the brain and cognitive functioning. J. Biosoc. Sci. 1997, 28, 463–479. [Google Scholar]
- McNay, E.C.; Fries, T.M.; Gold, P.E. Decreases in rat extracellular hippocampal glucose concentration associated with cognitive demand during a spatial task. Proc. Natl. Acad. Sci. USA 2000, 97, 2881–2885. [Google Scholar] [CrossRef] [PubMed]
- Fairclough, S.H.; Houston, K. A metabolic measure of mental effort. Biol. Psychol. 2004, 66, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Gailliot, M.T.; Baumeister, R.F.; DeWall, C.N.; Maner, J.K.; Plant, E.A.; Tice, D.M.; Brewer, L.E.; Schmeichel, B.J. Self-control relies on glucose as a limited energy source: Willpower is more than a metaphor. J. Personal. Soc. Psychol. 2007, 92, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Inzlicht, M.; Schmeichel, B.J. What is ego depletion? Toward a mechanistic revision of the resource model of self-control. Perspect. Psychol. Sci. 2012, 7, 450–463. [Google Scholar] [CrossRef] [PubMed]
- Muraven, M.; Baumeister, R. Self-regulation and depletion of limited resources: Does self-control resemble a muscle? Psychol. Bull. 2000, 126, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Hagger, M.S.; Chatzisarantis, N.L.D. The sweet taste of success: The presence of glucose in the oral cavity moderates the depletion of self-control resources. Personal. Soc. Psychol. Bull. 2013, 39, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Kurzban, R. Does the brain consume additional glucose during self-control tasks? Evol. Psychol. 2010, 8, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.; Shirk, S.; Burgin, C.; Martin, L. The gargle effect rinsing the mouth with glucose enhances self-control. Psychol. Sci. 2012, 23, 1470–1472. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Jeukendrup, A.E.; Jones, D.A. The effect of carbohydrate mouth rinse on 1-h cycle time trial performance. Med. Sci. Sports Exerc. 2004, 36, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.; Bridge, M.; Jones, D. Carbohydrate sensing in the human mouth: Effects on exercise performance and brain activity. J. Physiol. 2009, 587, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Pottier, A.; Bouckaert, J.; Gilis, W.; Roels, T.; Derave, W. Mouth rinse but not ingestion of a carbohydrate solution improves 1-h cycle time trial performance. Scand. J. Med. Sci. Sports 2010, 20, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kringelbach, M.L. Food for thought: Hedonic experience beyond homeostasis in the human brain. Neuroscience 2004, 126, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Molden, D.C.; Hui, C.M.; Scholer, A.A.; Meier, B.P.; Noreen, E.E.; D’Agostino, P.R.; Martin, V. Motivational versus metabolic effects of carbohydrates on self-control. Psychol. Sci. 2012, 23, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Boyle, N.; Lawton, C.L.; Newens, K.; Allen, A.; Croden, F.; Smith, K.; Dye, L. The effect of ingesting and rinsing sucrose and sucralose on depleted self-control performance. Physiol. Behav. 2015, in press. [Google Scholar]
- Convit, A. Links between cognitive impairment in insulin resistance: An explanatory model. Neurobiol. Aging 2005, 26, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Gruetter, R.; Novotny, E.J.; Boulware, S.D.; Rothman, D.L.; Shulman, R.G. 1 h NMR studies of glucose transport in the human brain. J. Cereb. Blood Flow Metab. 1996, 16, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Triguero, D.; Farrell, C.R. Downregulation of blood-brain barrier glucose transporter in experimental diabetes. Diabetes 1990, 39, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Haier, R.J.; Siegel, B.V.; Maclachlan, A.; Soderling, E.; Lottenberg, S.; Buchsbaum, M.S. Regional glucose metabolic changes after learning a complex visuospatial motor task—A positron emission tomographic study. Brain Res. 1992, 570, 134–143. [Google Scholar] [CrossRef]
- Brands, A.M.A.; Biessels, G.J.; De Haan, E.H.F.; Kappelle, L.J.; Kessels, R.P.C. The effects of type 1 diabetes on cognitive performance—A meta-analysis. Diabetes Care 2005, 28, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.E. Role of glucose in regulating the brain and cognition. Am. J. Clin. Nutr. 1995, 61, S987–S995. [Google Scholar] [CrossRef]
- Greenwood, C.E.; Kaplan, R.J.; Hebblethwaite, S.; Jenkins, D.J.A. Carbohydrate-induced memory impairment in adults with type 2 diabetes. Diabetes Care 2003, 26, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.V.; Frier, B.M.; Strachan, M.W.J. The relationship between type 2 diabetes and cognitive dysfunction: Longitudinal studies and their methodological limitations. Eur. J. Pharmacol. 2004, 490, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Lamport, D.J.; Lawton, C.L.; Mansfield, M.W.; Dye, L. Impairments in glucose tolerance can have a negative impact on cognitive function: A systematic research review. Neurosci. Biobehav. Rev. 2009, 33, 394–413. [Google Scholar] [CrossRef] [PubMed]
- Strachan, M.W.J. Insulin and cognitive function in humans: Experimental data and therapeutic considerations. Biochem. Soc. Trans. 2005, 33, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Hamann, S. Cognitive and neural mechanisms of emotional memory. Trends Cogn. Sci. 2001, 5, 394–400. [Google Scholar] [CrossRef]
- Blake, T.M.; Varnhagen, C.K.; Parent, M.B. Emotionally arousing pictures increase blood glucose levels and enhance recall. Neurobiol. Learn. Mem. 2001, 75, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Parent, M.B.; Varnhagen, C.; Gold, P.E. A memory-enhancing emotionally arousing narrative increases blood glucose levels in human subjects. Psychobiology 1999, 27, 386–396. [Google Scholar]
- Fillmore, M.; Vogelsprott, M. Expected effect of caffeine on motor-performance predicts the type of response to placebo. Psychopharmacology (Berlin) 1992, 106, 209–214. [Google Scholar] [CrossRef]
- Gorby, H.E.; Brownawell, A.M.; Falk, M.C. Do specific dietary constituents and supplements affect mental energy? Review of the evidence. Nutr. Rev. 2010, 68, 697–718. [Google Scholar] [CrossRef] [PubMed]
- Gailliot, M.T.; Peruche, B.M.; Plant, E.A.; Baumeister, R.F. Stereotypes and prejudice in the blood: Sucrose drinks reduce prejudice and stereotyping. J. Exp. Soc. Psychol. 2009, 45, 288–290. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; de Araujo, I.E.T.; Rolls, E.T. Taste-related activity in the human dorsolateral prefrontal cortex. Neuroimage 2004, 21, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Purnell, J.Q.; Klopfenstein, B.A.; Stevens, A.A.; Havel, P.J.; Adams, S.H.; Dunn, T.N.; Krisky, C.; Rooney, W.D. Brain functional magnetic resonance imaging response to glucose and fructose infusions in humans. Diabetes Obes. Metab. 2011, 13, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Thayer, J.F.; Hansen, A.L.; Saus-Rose, E.; Johnsen, B.H. Heart rate variability, prefrontal neural function, and cognitive performance: The neurovisceral integration perspective on self-regulation, adaptation, and health. Ann. Behav. Med. 2009, 37, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, H.B.; Stoddart, C.; Simeon, J.G. Double-blind challenge studies of behavioral and cognitive effects of sucrose-aspartame ingestion in normal-children. Nutr. Rev. 1986, 44, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wolraich, M.L.; Lindgren, S.D.; Stumbo, P.J.; Stegink, L.D.; Appelbaum, M.I.; Kiritsy, M.C. Effects of diets high in sucrose or aspartame on the behavior and cognitive performance of children. N. Engl. J. Med. 1994, 330, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwaluw, N.L.; van de Rest, O.; Kessels, R.P.C.; de Groot, L. Short-term effects of glucose and sucrose on cognitive performance and mood in elderly people. J. Clin. Exp. Neuropsychol. 2014, 36, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Harte, C.B.; Kanarek, R.B. The effects of nicotine and sucrose on spatial memory and attention. Nutr. Neurosci. 2004, 7, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Dye, L.; Gilsenan, M.B.; Quadt, F.; Martens, V.E.G.; Bot, A.; Lasikiewiczi, N.; Camidge, D.; Croden, F.; Lawton, C. Manipulation of glycemic response with isomaltulose in a milk-based drink does not affect cognitive performance in healthy adults. Mol. Nutr. Food Res. 2010, 54, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Kashimura, J.; Nagai, Y.; Ebashi, T. The effect of palatinose on mental concentration in humans. J. Nutr. Sci. Vitaminol. (Tokyo) 2003, 49, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Lina, B.A.R.; Jonker, D.; Kozianowski, G. Isomaltulose (palatinose®): A review of biological and toxicological studies. Food Chem. Toxicol. 2002, 40, 1375–1381. [Google Scholar] [CrossRef]
- Holub, I.; Gostner, A.; Theis, S.; Nosek, L.; Kudlich, T.; Melcher, R.; Scheppach, W. Novel findings on the metabolic effects of the low glycaemic carbohydrate isomaltulose (palatinose™). Br. J. Nutr. 2010, 103, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Okuda, Y.; Yamashita, K. Changes in blood glucose and insulin after an oral palatinose administration in normal subjects. Endocrinol. Jpn. 1985, 32, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Mizuno, A.; Sakuma, M.; Fukaya, M.; Matsuo, K.; Muto, K.; Sasaki, H.; Matsuura, M.; Okumura, H.; Yamamoto, H.; et al. Effects of a palatinose-based liquid diet (inslow) on glycemic control and the second-meal effect in healthy men. Metabolism 2007, 56, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Sekartini, R.; Wiguna, T.; Bardosono, S.; Novita, D.; Arsianti, T.; Calame, W.; Schaafsma, A. The effect of lactose-isomaltulose-containing growing-up milks on cognitive performance of Indonesian children: A cross-over study. Br. J. Nutr. 2013, 110, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Taib, M.N.M.; Shariff, Z.M.; Wesnes, K.A.; Abu Saad, H.; Sariman, S. The effect of high lactose-isomaltulose on cognitive performance of young children. A double blind cross-over design study. Appetite 2012, 58, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Young, H.; Benton, D. The effect of using isomaltulose (palatinose) to modulate the glycaemic properties of breakfast on the cognitive performance of children. Eur. J. Nutr. 2015, 54, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Brouns, F.; Bjorck, I.; Frayn, K.N.; Gibbs, A.L.; Lang, V.; Slama, G.; Wolever, T.M.S. Glycaemic index methodology. Nutr. Res. Rev. 2005, 18, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Gilsenan, M.B.; de Bruin, E.A.; Dye, L. The influence of carbohydrate on cognitive performance: A critical evaluation from the perspective of glycaemic load. Br. J. Nutr. 2009, 101, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Brand-Miller, J.C.; Thomas, M.; Swan, V.; Ahmad, Z.I.; Petocz, P.; Colagiuri, S. Physiological validation of the concept of glycemic load in lean young adults. J. Nutr. 2003, 133, 2728–2732. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E.E.; Antoine, J.M.; Benton, D.; Bjorck, I.; Bozzetto, L.; Brouns, F.; Diamant, M.; Dye, L.; Hulshof, T.; Holst, J.J.; et al. Impact of postprandial glycaemia on health and prevention of disease. Obes. Rev. 2012, 13, 923–984. [Google Scholar] [CrossRef] [PubMed]
- Amiel, S.A. Cognitive function testing in studies of acute hypoglycaemia: Rights and wrongs? Diabetologia 1998, 41, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Colombani, P.C.; Langhans, W.; Wenk, C. Cognitive performance and its relationship with postprandial metabolic changes after ingestion of different macronutrients in the morning. Br. J. Nutr. 2001, 85, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Colombani, P.C.; Langhans, W.; Wenk, C. Carbohydrate to protein ratio in food and cognitive performance in the morning. Physiol. Behav. 2002, 75, 411–423. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Ostman, E.M.; Granfeldt, Y.; Bjorck, I.M.E. Effect of cereal test breakfasts differing in glycemic index and content of indigestible carbohydrates on daylong glucose tolerance in healthy subjects. Am. J. Clin. Nutr. 2008, 87, 645–654. [Google Scholar] [PubMed]
- Ingwersen, J.; Defeyter, M.A.; Kennedy, D.O.; Wesnes, K.A.; Scholey, A.B. A low glycaemic index breakfast cereal preferentially prevents children’s cognitive performance from declining throughout the morning. Appetite 2007, 49, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Wesnes, K.A.; Pincock, C.; Richardson, D.; Helm, G.; Hails, S. Breakfast reduces declines in attention and memory over the morning in schoolchildren. Appetite 2003, 41, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Micha, R.; Rogers, P.J.; Nelson, M. The glycaemic potency of breakfast and cognitive function in school children. Eur. J. Clin. Nutr. 2010, 64, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.; Nabb, S. Breakfasts that release glucose at different speeds interact with previous alcohol intake to influence cognition and mood before and after lunch. Behav. Neurosci. 2004, 118, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.; Ruffin, M.P.; Lassel, T.; Nabb, S.; Messaoudi, M.; Vinoy, S.; Desor, D.; Lang, V. The delivery rate of dietary carbohydrates affects cognitive performance in both rats and humans. Psychopharmacology (Berlin) 2003, 166, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Nabb, S.; Benton, D. The influence on cognition of the interaction between the macro-nutrient content of breakfast and glucose tolerance. Physiol. Behav. 2006a, 87, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Nabb, S.L.; Benton, D. The effect of the interaction between glucose tolerance and breakfasts varying in carbohydrate and fibre on mood and cognition. Nutr. Neurosci. 2006b, 9, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.S. Dietary fibre, glycaemic response, and diabetes. Mol. Nutr. Food Res. 2005, 49, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.; Prabhasankar, P. Health based pasta: Redefining the concept of the next generation convenience food. Crit. Rev. Food Sci. Nutr. 2011, 52, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Dammann, K.W.; Bell, M.; Kanter, M.; Berger, A. Effects of consumption of sucromalt, a slowly digestible carbohydrate, on mental and physical energy questionnaire responses. Nutr. Neurosci. 2013, 16, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Granfeldt, Y.; Nyberg, L.; Bjorck, I. Muesli with 4 g oat [beta]-glucans lowers glucose and insulin responses after a bread meal in healthy subjects. Eur. J. Clin. Nutr. 2007, 62, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Allgood, L.D.; Turner, J.; Oeltgen, P.R.; Daggy, B.P. Effects of psyllium on glucose and serum lipid responses in men with type 2 diabetes and hypercholesterolemia. Am. J. Clin. Nutr. 1999, 70, 466–473. [Google Scholar] [PubMed]
- Nilsson, M.; Holst, J.J.; Björck, I.M. Metabolic effects of amino acid mixtures and whey protein in healthy subjects: Studies using glucose-equivalent drinks. Am. J. Clin. Nutr. 2007, 85, 996–1004. [Google Scholar] [PubMed]
- Nilsson, M.; Stenberg, M.; Frid, A.; Holst, J.; Bjorck, I. Glycemia and insulinemia in healthy subjects after lactose-equivalent meals of milk and other food proteins: The role of plasma amino acids and incretins. Am. J. Clin. Nutr. 2004, 80, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.; Ward, L.; Bastian, E.; Jenkins, A.; Campbell, J.; Vuksan, V. A whey protein supplement decreases post-prandial glycemia. Nutr. J. 2009, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Gunnerud, U.J.; Ostman, E.M.; Bjorck, I.M.E. Effects of whey proteins on glycaemia and insulinaemia to an oral glucose load in healthy adults; a dose-response study. Eur. J. Clin. Nutr. 2013, 67, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Sunram-Lea, S.I.; Foster, J.K.; Durlach, P.; Perez, C. The influence of fat co-administration on the glucose memory facilitation effect. Nutr. Neurosci. 2004, 7, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A.; Daval, J.L.; Debry, G. Caffeine and the central-nervous-system—Mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res. Rev. 1992, 17, 139–169. [Google Scholar] [CrossRef]
- Kamimori, G.H.; Johnson, D.; Thorne, D.; Belenky, G. Multiple caffeine doses maintain vigilance during early morning operations. Aviat. Space Environ. Med. 2005, 76, 1046–1050. [Google Scholar] [PubMed]
- Spaeth, A.M.; Goel, N.; Dinges, D.F. Cumulative neurobehavioral and physiological effects of chronic caffeine intake: Individual differences and implications for the use of caffeinated energy products. Nutr. Rev. 2014, 72, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Scharff, J.L.; Broida, J.P.; Conway, K.; Yue, A. The interaction of parental alcoholism, adaptation role, and familial dysfunction. Addict. Behav. 2004, 29, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, N.L. Clinical-pharmacology of caffeine. Annu. Rev. Med. 1990, 41, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Glade, M.J. Caffeine-not just a stimulant. Nutrition 2010, 26, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.J.; Dernoncourt, C. Regular caffeine consumption: A balance of adverse and beneficial effects for mood and psychomotor performance. Pharmacol. Biochem. Behav. 1998, 59, 1039–1045. [Google Scholar] [CrossRef]
- Smith, A. Effects of caffeine on human behavior. Food Chem. Toxicol. 2002, 40, 1243–1255. [Google Scholar] [CrossRef]
- Warburton, D.M. Effects of caffeine on cognition and mood without caffeine abstinence. Psychopharmacology (Berlin) 1995, 119, 66–70. [Google Scholar] [CrossRef]
- Loke, W.H. Effects of caffeine on mood and memory. Physiol. Behav. 1988, 44, 367–372. [Google Scholar] [CrossRef]
- Mumford, G.K.; Evan, S.M.; Kaminski, B.J.; Preston, K.L.; Sannerud, C.A.; Silverman, K.; Griffiths, R.R. Discriminative stimulus and subjective effects of theobromine and caffeine in humans. Psychopharmacology (Berlin) 1994, 115, 1–8. [Google Scholar] [CrossRef]
- Ruijter, J.; De Ruiter, M.B.; Snel, J. The effects of caffeine on visual selective attention to color: An ERP study. Psychophysiology 2000, 37, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.; de Ruiter, M.B.; Snel, J.; Lorist, M.M. The influence of caffeine on spatial-selective attention: An event-related potential study. Clin. Neurophysiol. 2000, 111, 2223–2233. [Google Scholar] [CrossRef]
- Hewlett, P.; Smith, A. Effects of repeated doses of caffeine on performance and alertness: New data and secondary analyses. Hum. Psychopharmacol.-Clin. Exp. 2007, 22, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R.; Wurtman, R.J.; Emde, G.G.; Roberts, C.; Coviella, I.L.G. The effects of low-doses of caffeine on human-performance and mood. Psychopharmacology (Berlin) 1987, 92, 308–312. [Google Scholar] [CrossRef]
- Peeling, P.; Dawson, B. Influence of caffeine ingestion on perceived mood states, concentration, and arousal levels during a 75-min university lecture. Adv. Physiol. Educ. 2007, 31, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Foreman, N.; Barraclough, S.; Moore, C.; Mehta, A.; Madon, M. High doses of caffeine impair performance of a numerical version of the stroop task in men. Pharmacol. Biochem. Behav. 1989, 32, 399–403. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion on the substantiation of health claims related to caffeine and increased fat oxidation leading to a reduction in body fat mass (ID 735, 1484), increased energy expenditure leading to a reduction in body weight (ID 1487), increased alertness (ID 736, 1101, 1187, 1485, 1491, 2063, 2103) and increased attention (ID 736, 1485, 1491, 2375) pursuant to article 13(1) of regulation (EC) No. 1924/2006. EFSA J. 2011, 9, 2054. [Google Scholar]
- European Food Safety Authority (EFSA). Scientific opinion on the substantiation of a health claim related to caffeine and increased alertness pursuant to article 13(5) of regulation (EC) No. 1924/2006. EFSA J. 2014, 12, 3574. [Google Scholar]
- Seifert, S.M.; Schaechter, J.L.; Hershorin, E.R.; Lipshultz, S.E. Health effects of energy drinks on children, adolescents, and young adults. Pediatrics 2011, 127, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Van den Eynde, F.; van Baelen, P.C.; Portzky, M.; Audenaert, K. The effects of energy drinks on cognitive performance. Tijdschr. Psychiatr. 2008, 50, 273–281. [Google Scholar] [PubMed]
- Kennedy, D.O.; Scholey, A.B. A glucose-caffeine ‘energy drink’ ameliorates subjective and performance deficits during prolonged cognitive demand. Appetite 2004, 42, 331–333. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Hu, H.L.; Nobre, A.C. The effects of combined caffeine and glucose drinks on attention in the human brain. Nutr. Neurosci. 2005, 8, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Warburton, D.M.; Bersellini, E.; Sweeney, E. An evaluation of a caffeinated taurine drink on mood, memory and information processing in healthy volunteers without caffeine abstinence. Psychopharmacology (Berlin) 2001, 158, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Aniţei, M.; Schuhfried, G.; Chraif, M. The influence of energy drinks and caffeine on time reaction and cognitive processes in young Romanian students. Procedia Soc. Behav. Sci. 2011, 30, 662–670. [Google Scholar] [CrossRef]
- Gendle, M.H.; Smucker, D.M.; Stafstrom, J.A.; Helterbran, M.C.; Glazer, K.S. Attention and reaction time in university students following the consumption of red bull®. Open Nutr. J. 2009, 9, 8–10. [Google Scholar] [CrossRef]
- Howard, M.A.; Marczinski, C.A. Acute effects of a glucose energy drink on behavioral control. Exp. Clin. Psychopharmacol. 2010, 18, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Childs, E.; de Wit, H. Enhanced mood and psychomotor performance by a caffeine-containing energy capsule in fatigued individuals. Exp. Clin. Psychopharmacol. 2008, 16, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Smit, H.J.; Cotton, J.R.; Hughes, S.C.; Rogers, P.J. Mood and cognitive performance effects of “energy” drink constituents: Caffeine, glucose and carbonation. Nutr. Neurosci. 2004, 7, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.A.; Reyner, L.A. Beneficial effects of an “energy drink” given to sleepy drivers. Amino Acids 2001, 20, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.; Savage, K.; O’Neill, B.V.; Owen, L.; Stough, C.; Priestley, C.; Wetherell, M. Effects of two doses of glucose and a caffeine-glucose combination on cognitive performance and mood during multi-tasking. Hum. Psychopharmacol.-Clin. Exp. 2014, 29, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Horne, J.A. A high sugar content, low caffeine drink does not alleviate sleepiness but may worsen it. Hum. Psychopharmacol.-Clin. Exp. 2006, 21, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Reyner, L.A.; Horne, J.A. Efficacy of a ‘functional energy drink’ in counteracting driver sleepiness. Physiol. Behav. 2002, 75, 331–335. [Google Scholar] [CrossRef]
- Mets, M.A.; Ketzer, S.; Blom, C.; van Gerven, M.H.; van Willigenburg, G.M.; Olivier, B.; Verster, J.C. Positive effects of Red Bull® energy drink on driving performance during prolonged driving. Psychopharmacology (Berlin) 2011, 214, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Smit, H.J.; Grady, M.L.; Finnegan, Y.E.; Hughes, S.A.C.; Cotton, J.R.; Rogers, P.J. Role of familiarity on effects of caffeine- and glucose-containing soft drinks. Physiol. Behav. 2006, 87, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Sunram-Lea, S.I.; Owen-Lynch, J.; Robinson, S.; Jones, E.; Hu, H.L. The effect of energy drinks on cortisol levels, cognition and mood during a fire-fighting exercise. Psychopharmacology (Berlin) 2012, 219, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Bono, E.; Rohleder, N.; Hellhammer, D.H.; Salvador, A.; Kirschbaum, C. Glucose but not protein or fat load amplifies the cortisol response to psychosocial stress. Horm. Behav. 2002, 41, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, C.; Bono, E.G.; Rohleder, N.; Gessner, C.; Pirke, K.M.; Salvador, A.; Hellhammer, D.H. Effects of fasting and glucose load on free cortisol responses to stress and nicotine. J. Clin. Endocrinol. Metab. 1997, 82, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T.E. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Sünram-Lea, S.I.; Leach, J.; Owen-Lynch, P.J. Cognitive and biochemical changes in individuals exposed to an acute stressor. Proc. Br. Psychol. Soc. 2004, 13, 24. [Google Scholar]
- Scholey, A.B.; Kennedy, D.O. Cognitive and physiological effects of an “energy drink”: An evaluation of the whole drink and of glucose, caffeine and herbal flavouring fractions. Psychopharmacology (Berlin) 2004, 176, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Adan, A.; Serra-Grabulosa, J.M. Effects of caffeine and glucose, alone and combined, on cognitive performance. Hum. Psychopharmacol.-Clin. Exp. 2010, 25, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Serra-Grabulosa, J.M.; Adan, A.; Falcon, C.; Bargallo, N. Glucose and caffeine effects on sustained attention: An exploratory fmri study. Hum. Psychopharmacol.-Clin. Exp. 2010, 25, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.E.; Mahoney, C.R.; Brunye, T.T.; Gardony, A.L.; Taylor, H.A.; Kanarek, R.B. Differential cognitive effects of energy drink ingredients: Caffeine, taurine, and glucose. Pharmacol. Biochem. Behav. 2012, 102, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Young, H.A.; Benton, D. Caffeine can decrease subjective energy depending on the vehicle with which it is consumed and when it is measured. Psychopharmacology (Berlin) 2013, 228, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Maridakis, V.; O’Connor, P.J.; Tomporowski, P.D. Sensitivity to change in cognitive performance and mood measures of energy and fatigue in response to morning caffeine alone or in combination with carbohydrate. Int. J. Neurosci. 2009, 119, 1239–1258. [Google Scholar] [CrossRef] [PubMed]
- James, J.E. Does caffeine enhance or merely restore degraded psychomotor performance. Neuropsychobiology 1994, 30, 124–125. [Google Scholar] [CrossRef] [PubMed]
- James, J.E.; Rogers, P.J. Effects of caffeine on performance and mood: Withdrawal reversal is the most plausible explanation. Psychopharmacology (Berlin) 2005, 182, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A.; Boyet, S. Dose-response study of caffeine effects on cerebral functional activity with a specific focus on dependence. Brain Res. 2000, 858, 71–77. [Google Scholar] [CrossRef]
- Graham, T.E.; Sathasivam, P.; Rowland, M.; Marko, N.; Greer, F.; Battram, D. Caffeine ingestion elevates plasma insulin response in humans during an oral glucose tolerance test. Can. J. Physiol. Pharmacol. 2001, 79, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Pizziol, A.; Tikhonoff, V.; Paleari, C.D.; Russo, E.; Mazza, A.; Ginocchio, G.; Onesto, C.; Pavan, L.; Casiglia, E.; Pessina, A.C. Effects of caffeine on glucose tolerance: A placebo-controlled study. Eur. J. Clin. Nutr. 1998, 52, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Blokland, A. Acetylcholine: A neurotransmitter for learning and memory? Brain Res. Brain Res. Rev. 1995, 21, 285–300. [Google Scholar] [CrossRef]
- Rusted, J.M.; Eatonwilliams, P.; Warburton, D.M. A comparison of the effects of scopolamine and diazepam on working memory. Psychopharmacology (Berlin) 1991, 105, 442–445. [Google Scholar] [CrossRef]
- Rusted, J.M.; Warburton, D.M. The effects of scopolamine on working memory in healthy-young volunteers. Psychopharmacology (Berlin) 1988, 96, 145–152. [Google Scholar] [CrossRef]
- Biaggioni, I.; Paul, S.; Puckett, A.; Arzubiaga, C. Caffeine and theophylline as adenosine receptor antagonists in humans. J. Pharmacol. Exp. Ther. 1991, 258, 588–593. [Google Scholar] [PubMed]
- Wenk, G.L. An hypothesis on the role of glucose in the mechanism of action of cognitive enhancers. Psychopharmacology (Berlin) 1989, 99, 431–438. [Google Scholar] [CrossRef]
- Smith, A.; Brice, C.; Nash, J.; Rich, N.; Nutt, D.J. Caffeine and central noradrenaline: Effects on mood, cognitive performance, eye movements and cardiovascular function. J. Psychopharmacol. 2003, 17, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.P.; Canzler, E.; Giers, H.; Walther, H. Time course of impaired elimination of caffeine by the oral depot contraceptive deposiston. Zentralbl. Gynakol. 1991, 113, 297–302. [Google Scholar] [PubMed]
- Van Soeren, M.H.; Graham, T.E. Effect of caffeine on metabolism, exercise endurance, and catecholamine responses after withdrawal. J. Appl. Physiol. 1998, 85, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.; Wade, D.; Workman, R.; Woosley, R.L.; Oates, J.A. Tolerance to the humoral and hemodynamic-effects of caffeine in man. J. Clin. Investig. 1981, 67, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W.; Fredholm, B.B. Caffeine—An atypical drug of dependence. Drug Alcohol Depend. 1998, 51, 199–206. [Google Scholar] [CrossRef]
- Ploutz-Snyder, L.; Foley, J.; Ploutz-Snyder, R.; Kanaley, J.; Sagendorf, K.; Meyer, R. Gastric gas and fluid emptying assessed by magnetic resonance imaging. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 79, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.E. Caffeine and exercise—Metabolism, endurance and performance. Sports Med. 2001, 31, 785–807. [Google Scholar] [CrossRef] [PubMed]
- Keijzers, G.B.; De Galan, B.E.; Tack, C.J.; Smits, P. Caffeine can decrease insulin sensitivity in humans. Diabetes Care 2002, 25, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A. A circumplex model of affect. J. Pers. Soc. Psychol. 1980, 39, 1161–1178. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A. Negative affectivity—The disposition to experience aversive emotional states. Psychol. Bull. 1984, 96, 465–490. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Dye, L.; Siobhan Mitchell, E.; Layé, S.; Saunders, C.; Boyle, N.; Schuermans, J.; Sijben, J. Examining techniques for measuring the effects of nutrients on mental performance and mood state. Eur. J. Nutr. 2016, 55, 1991–2000. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cognitive Domains | Subcomponents | Cognitive Test Examples | Related Factors |
|---|---|---|---|
| Episodic Memory: Memory of autobiographical events (times, places, associated emotions, and other contextual who, what, when, where, why knowledge) that can be explicitly stated | |||
| Immediate Recall: (Verbal or Visual/spatial). Learning/encoding and recall of new information | Logical or Paragraph memory, List Learning tasks (e.g., California Verbal Learning), Paired Associate Verbal Learning Test; Pattern Recall | Primacy/Recency effects: Stimuli shown at the beginning (primacy) and the end (recency) of a presentation are more likely to be recalled Emotional valence: The intrinsic attractiveness (positive valence) or aversiveness (negative valence) of an event, stimuli, or situation | |
| Delayed Recall: (Verbal or Visual/spatial) Recall of previously learned information | As above | ||
| Recognition: (Verbal or Visual/spatial/faces). Ability to accurately recognize learned information (in the case of source monitoring, identifying the context in which the information was learned) | As above | ||
| Semantic Memory: General knowledge (facts, ideas, meaning and concepts) accumulated throughout life that can be retrieved without reference to the circumstances in which it was originally acquired | Tests of general knowledge | ||
| Implicit Memory: The use of previous experiences to aid the performance of a task without conscious awareness of these previous experiences | |||
| Procedural memory: Memory for performance of particular types of action. Procedural memory guides the processes we perform (e.g., driving) and most frequently resides below the level of conscious awareness | Pursuit Rotor Task; Serial Reaction Time Task; divided attention tasks | ||
| Priming: Exposure to a stimulus influences the responses to a subsequent stimulus | Word-stem Completion Task; Lexical Decision Task; word association tests | ||
| Attention: The behavioral and cognitive process of selectively concentrating on a discrete aspect of information, whether deemed subjective or objective, while ignoring other perceivable information. Attention can also be considered the allocation of limited processing resources | |||
| Attentional Capacity: Accuracy of attention span (e.g., repeating digit sequence) | Digit Span (especially Digits Forward); Digit Symbol Substitution (DSST) | Divided attention/multi-tasking: the performance of multiple tasks concurrently to apply extra demand/load on attentional resources | |
| Vigilance/Focus: Sustaining attention over time to detect target stimuli, often with a demand to ignore distractors | Repeated Digits Vigilance, Continuous Performance, Bakan/Rapid Visual Information Processing (RVIP); Digit/Letter Cancellation | ||
| Processing/Perceptual Speed: Ability to process information and execute relevant operations within the allotted time | Trail-making Test (Part A and B); Simple/Choice Reaction time | ||
| Executive Functions: An umbrella term for the management (regulation, control) of cognitive processes, including working memory, reasoning, task flexibility, and problem solving as well as planning and execution | |||
| Reasoning/Planning: Thinking with conscious intent to reach a conclusion (planning involves induction, reasoning is more deductive) | Graduate and Managerial Assessment Test of Abstract Reasoning; Tower of Hanoi | ||
| Inhibitory Control/Self-control: Effortful inhibition of predominant responses, emotions, thoughts, and impulses, permitting behavior to vary adaptively moment to moment | Attention-switching tests; Go/No-Go; Stroop Color and Word Test | ||
| Working Memory: Allows information maintained in temporary storage to be manipulated for complex cognitive operations | Paced Auditory Serial Addition Task; Serial 3s, Serial 7s; Brown Peterson Trigrams; Corsi Block Tapping | ||
| Problem-solving: Using generic or ad hoc methods, in an orderly manner, for finding solutions to problems | Anagram Tasks; Mathematical Problem Solving | ||
| Language: Ability to speak or perform in an acquired language | |||
| Verbal Fluency: Oral production of words fitting a specified category (e.g., animals) or beginning with a specified letter | Category Fluency; Phonemic fluency | ||
| Verbal Reasoning: Ability to read and think about information presented and apply logic to determine whether specific conclusions can be drawn from the information | Verbal Reading-Comprehension Test | ||
| Motor Performance: Movements and motions carried out by co-ordination of the brain, nervous system, and muscles | |||
| Gross motor speed: Speeded gross manual dexterity | Simple tapping task | Driving: Measures of driving performance require fine, gross and psychomotor skills | |
| Fine motor speed: Speeded fine manual dexterity | Grooved Pegboard | ||
| Psychomotor skill: The physical encoding of information, with movement and/or with activities where the gross and fine muscles are used for expressing or interpreting information or concepts | Psychomotor Vigilance Task (PVT); throwing; manipulation of objects | ||
| Reaction Time (RT): Speed of a response (in seconds or milliseconds) to a cue, stimulus or event | |||
| Simple RT: Speed of response to a target (e.g., pressing a button when a cross appears) | Simple Reaction Time Test | Note: RT can be used as an index of performance on other domains of cognitive function (e.g., speed of recalling words, speed of working memory performance) | |
| Choice RT: Analogous to simple RT except that stimulus and response uncertainty are introduced by having multiple possible stimuli and responses | 2-choice Reaction Time Test | ||
| VisuoSpatial Function: The ability to comprehend and conceptualize visual representations and spatial relationships in learning and performing a task | Judgment of Line Orientation Test; Clock Test; Hooper Visual Organization Task |
| Authors | Sample Size (Age) | Dose (Glucose) | Design (Within or Between Subjects) | Cognitive Outcomes | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Episodic Memory | Working Memory | Attention | Recognition Memory | Visuospatial Memory | Semantic Memory | Face Recognition | Verbal Fluency | Visuospatial Functioning | Executive Functioning | Problem Solving | Implicit Memory | Self-Control | Processing Speed/RT | Mood Effects | ||||
| Hall et al. 1989 [22] | 12 (M = 20) 11 (M = 67.4) | 50 g 50 g | Within (overnight fast) Within (overnight fast) | − O | O − | − − | ||||||||||||
| Benton, 1990 [33] | 20 + 40 (M = 20.4 & 21.05) | 25 g | Between (4 h fast) | O | ||||||||||||||
| Azari, 1991 [29] | 18 (M = 21) | 30 g | Within (10 h fast) | − | − | |||||||||||||
| Azari, 1991 [29] | 18 (M = 21) | 100 g | Within (10 h fast) | − | − | |||||||||||||
| Benton & Owens, 1993 [30] | 100 (M = 21.7) 53 (M = 21.5) | 50 g 50 g (+25 g at +45 & +75 min) | Between (4 h fast) Between (4 h fast) | − − | − − | |||||||||||||
| Owens & Benton, 1994 [21] | 96 (M = 21.2) | 50 g | Between (No dietary restriction) | O 3 | ||||||||||||||
| Craft et al. 1994 [34] | 27 (M = 20.8) 32 (M = 68.5) | 50 g 50 g | Within (overnight fast) Within (overnight fast) | O 3,1 O 3,1 | − − | − − | − − | − − | ||||||||||
| Benton et al., 1994 [35] | 70 + 50 (M = 21.5 & 21.7) | 50 (+25 g at +30 min) | Between (No dietary restriction) | O 3 | − | − | ||||||||||||
| Parker & Benton, 1995 [36] | 100 (M = 20.15) | 50 (+25 g at +30 min) | Between (No dietary restriction) | O 2 | − | |||||||||||||
| Manning et al., 1997 [31] | 24 (M = 18.6) 23 (M = 67) | 50 g 50 g | Within (8 h fast) Within (8 h fast) | − O | − − | − − | ||||||||||||
| Foster et al., 1998 [7] | 30 (M = 19.5) | 25 g | Between (12 h fast) | O a | − | − | − | |||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 10 mg/kg | Between (No dietary restriction) | − | ||||||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 100 mg/kg | Between (No dietary restriction) | − | ||||||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 300 mg/kg | Between (No dietary restriction) | O 4 | ||||||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 500 mg/kg | Between (No dietary restriction) | − | ||||||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 800 mg/kg | Between (No dietary restriction) | O 4 | ||||||||||||||
| Messier et al. 1998 [37] | 100 (M = 21.3) | 1000 mg/kg | Between (No dietary restriction) | − | ||||||||||||||
| Winder & Borrill, 1998 [32] | 104 (M = 29.2) | 50 g | Between (No dietary restriction) | − | − | |||||||||||||
| Messier et al. 1999 [38] | 31 (M = 21.3) | 50 g | Within (overnight fast) | O 3 | ||||||||||||||
| Donohoe & Benton, 1999 [39] | 67 + 69 (M = 21.8 & 20.2) | 50 g | Between (No dietary restriction) | O | − | − | ||||||||||||
| Metzger, 2000 [40] | 34 (M = 21.1) | 50 g | Between (9 h fast) | O | ||||||||||||||
| Kennedy & Scholey, 2000 [23] | 20 (M = 20.4) | 25 g | Within (overnight fast) | O b | − ᵇ | |||||||||||||
| Green et al. 2001 [41] | 26 (18-40) | 50 g | Between (8 h fast) | − | O 5 | O | Vigilance 5 | |||||||||||
| Morris & Sarll, 2001 [42] | 80 (M = 21.2) | 50 g | Between (overnight fast) | O c | ||||||||||||||
| Scholey et al. 2001 [43] | 20 (M = 22.7) | 25 g | Between (overnight fast) | − ᵇ | O ᵇ | − ᵇ | ||||||||||||
| Mohanty & Flint, 2001 [19] | 77 (M = 20.6) | 50 g | Between (overnight fast) | X 6 | ||||||||||||||
| Mohanty & Flint, 2001 [19] | 78 (M = 20.6) | 100 mg/kg | Between (overnight fast) | O X 6 | ||||||||||||||
| Sunram-Lea et al. 2001 [17] | 60 (18–28) | 25 g | Between (overnight fast vs. breakfast vs. lunch) | O a | − | O a | O a | |||||||||||
| Awad et al, 2002 [44] | 74 (M = 21) | 75 g | Between (overnight fast) | O a,ᵇ | ||||||||||||||
| Scholey & Fowles, 2002 [20] | 35 (M = 23.6) | 25 g | Between (No dietary restriction) | O | − | |||||||||||||
| Sunram-Lea et al. 2011 [16] | 60 (M = 21) | 25 g | Between (2 h fast) | O | − | O | O | − | ||||||||||
| Sunram-Lea et al. 2002a [18] | 80 (M = 20) | 25 g | Between (2 h fast) | O a | O a | O a | O a | − | ||||||||||
| Ford et al. 2002 [45] | 20 (20–23) | 25 g | Within (overnight fast) | − 6 | − 6 | |||||||||||||
| Flint & Turek, 2003 [46] | 67 (M = 19.49) | 10 mg/kg | Between (8 h fast) | − | ||||||||||||||
| Flint & Turek, 2003 [46] | 67 (M = 19.49) | 100 mg/kg | Between (8 h fast) | X | ||||||||||||||
| Flint & Turek, 2003 [46] | 67 (M = 19.49) | 500 mg/kg | Between (8 h fast) | − | ||||||||||||||
| Flint & Turek, 2003 [46] | 67 (M = 19.49) | 50 g | Between (8 h fast) | − | ||||||||||||||
| Meikle et al. 2004 3 [26] | 14 (M = 21.8) | 25 g | Within (overnight fast) | O | − | − | − | − | − | |||||||||
| Meikle et al. 2004 3 [26] | 14 (M = 21.8) | 50 g | Within (overnight fast) | O | − | − | − | − | − | |||||||||
| Meikle et al. 2004 3 [26] | 11 (M = 38.4) | 25 g | Within (overnight fast) | O ᵇ | O | O | − | − | − | |||||||||
| Meikle et al. 2004 3 [26] | 11 (M = 38.4) | 50 g | Within (overnight fast) | O ᵇ | O | O | − | − | − | |||||||||
| Meikle et al. 2005 [47] | 37 + 24 (M = 28.5 & 18.9) | 25 g | Between (overnight fast) | O ᵇ | ||||||||||||||
| Reay et al. 2006 [27] | 27 (M = 21.9) | 25 g | Within (overnight fast) | O ᵇ | O ᵇ | Mental Fatigue | ||||||||||||
| Riby et al. 2006 [48] | 14 (M = 30.1) | 25 g | Within (overnight fast) | O a | O a | − a | ||||||||||||
| Brandt et al. 2006 [49] | 40 (M = 22) | 25 g | Between (2 h fast) | − 6 | ||||||||||||||
| Gailliot et al. 2007 [50] | 62 + 73 + 18 | Not stated | Between | O | ||||||||||||||
| Masicampo & Baumeister, 2008 [51] | 121 | Not stated | Between | O | ||||||||||||||
| DeWall et al. 2008 [52] | 37 | Not stated | Between | O | ||||||||||||||
| Morris, 2008 [53] | 72 (M = 22.4) | 50 g | Between (No dietary restriction) | O | − | |||||||||||||
| Riby et al. 2008 [54] | 33 (35–55) | 25 g | Within (2 h fast) | − | − | |||||||||||||
| Riby et al. 2008 [54] | 33 (35–55) | 50 g | Within (2 h fast) | O | − | − | ||||||||||||
| Sunram-Lea et al. 2008 [14] | 56 (M = 20) | 25 g | Between (2 h fast) | O | ||||||||||||||
| Scholey & Kennedy, 2009 [55] | 120 (M = 21.6) | 25 g | Between (overnight fast) | − a | O a | |||||||||||||
| Scholey et al. 2009 [56] | 120 (M = 21.6) | 25 g | Within (overnight fast) | O 7 | O a | − a | ||||||||||||
| Owen et al. 2010 [57] | 90 (M = 21) | 25 g | Between (12 h fast) | − | − | − | − | |||||||||||
| Owen et al. 2010 [57] | 90 (M = 21) | 60 g | Between (12 h fast) | O | O | − | O | |||||||||||
| Brandt et al, 2010 [58] | 40 (M = 19.1) | 15 g | Between (2 h fast) | − 6 | ||||||||||||||
| Brandt et al, 2010 [58] | 40 (M = 21) | 25 g | Between (2 h fast) | − 6,ᵇ | ||||||||||||||
| Parent et al. 2011 [59] | 14 (M = 21.4) | 50 g | Within | O 8 | ||||||||||||||
| Smith et al. 2011 [60] | 40 (M = 15.5) | 25 g | Between (overnight fast) | O 9,ᵇ | − | |||||||||||||
| Sunram-Lea et al. 2011 [16] | 30 (M = 20) | 15 g | Between (12 h fast) | − | − | − | ||||||||||||
| Sunram-Lea et al. 2011 [16] | 30 (M = 20) | 25 g | Between (12 h fast) | O | −O 10 | O | ||||||||||||
| Sunram-Lea et al. 2011 [16] | 30 (M = 20) | 50 g | Between (12 h fast) | − | − | − | ||||||||||||
| Sunram-Lea et al. 2011 [16] | 30 (M = 20) | 60 g | Between (12 h fast) | − | − | − | ||||||||||||
| Jones et al. 2012 11 [25] | 18 (M = 19) | 25 g | Between (12 h fast) | X | X | O | Alertness | |||||||||||
| Brandt, 2013 [61] | 60 (M = 19.7) | 25g | Between (overnight fast) | O ᵇ | ||||||||||||||
| Scholey et al. 2013 [62] | 20 (18–35) | 25 g | Between (12 h fast) | O a | ||||||||||||||
| Owen et al. 2013 [13] | 24 (M = 20) | 25 g | Mixed (12 h fast) | O 3 | O 12 | O | − | |||||||||||
| Owen et al. 2013 [13] | 24 (M = 20) | 60 g | Mixed (12 h fast) | − | O 12 | O 13 | − | |||||||||||
| Brown & Riby, 2013 [63] | 35 (M = 22.19) | 25 g | Between (2 h fast) | O ᵇ | − | |||||||||||||
| Stollery & Christian, 2013 [28] | 93 (M = 20.7) | 50 g | Between | O 5 | O | − | ||||||||||||
| Miller et al. 2013 [24] | 36 (M = 23.25) | 25 g | Between (3 h fast) | O | ||||||||||||||
| Lange & Eggert, 2014 [64] | 70 + 115 (M = 21.80) | Not-stated | Between | − | ||||||||||||||
| Stollery & Christian, 2015 [65] | 80 (M = 22.4) | 25 g | Between | O 14 | − | |||||||||||||
| Brandt, 2015 [12] | 40 (M = 19.47) | 25 g | Between (overnight fast) | O a | ||||||||||||||
| Macpherson, 2015 [66] | 24 (M = 20.6) | 25 g | Within (overnight fast) | − a | ||||||||||||||
| CHO Source | Authors | Sample Size (Age) | Drink (Volume/Vehicle) | Design (Within or Between Subjects) | Cognitive Outcomes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Verbal Episodic Memory | Working Memory | Attention | Recognition Memory | Problem Solving | Semantic Memory | Face Recognition | Verbal Fluency | Visuospatial Functioning | Executive Functioning | Psychomotor Function | Self-control | |||||
| Fructose | Miller et al. 2013 [24] | 36 (M = 23.25) | (300 mL) 25 g glucose vs. 25 g fructose vs. sucralose placebo | Between (3 h fast) | O | |||||||||||
| Sucrose | Kashimura et al. 2003 [117] | 14 (M = 40.2) | (200 mL) 40 g sucrose vs. 40 g Palatinose | Between (12 h fast) | O | |||||||||||
| Harte & Kanarek, 2004 [115] | 14 (18–20) | (227.3 mL) Lemonade (17 g sucrose) vs. aspartame placebo | Within (2 h fast) | O ¹ | O | |||||||||||
| Gailliot et al. 2009 [108] | 56 | (397.7 mL) Sucrose vs. sucralose | Between | O ² | ||||||||||||
| Dye et al. 2010 [116] | 24 (18–32) | (429 mL) Milk-based drink containing isomaltulose vs. sucrose vs. water | Within (overnight fast) | − | − | − | ||||||||||
| Isomaltulose | Kashimura et al. 2003 [117] | 14 (M = 40.2) | (200 mL) 40 g sucrose vs. 40 g Palatinose | Between (12 h fast) | O | |||||||||||
| Kashimura et al. 2003 [117] | 14 (M = 32.8) | (185 g) 5 g Palatinose vs. (180 g) 10 g Palatinose | Between (12 h fast) | O | ||||||||||||
| Dye et al. 2010 [116] | 24 (18–32) | (429 mL) Milk-based drink containing isomaltulose vs. sucrose vs. water | Within (overnight fast) | − | − | − | ||||||||||
| Authors | Sample Size (Age) | Intervention | Design (Within or Between Subjects) | Cognitive Outcomes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Verbal Episodic Memory | Executive Function | Working Memory | Attention | Processing Speed/RT | Problem Solving | Moderating Effect of Postprandial Glycemic Response | ||||
| Benton et al. 2003 [137] | 71 (M = 21) | High-SAG biscuit, 50 g: 34 g CHO (8 g SAG + 20 g RAG, GI = 42) vs. Low-SAG cereal bar, 50 g: 31 g CHO (0.05 g SAG + 21 g RAG, GI = 66) | Between (overnight fast) | O | Enhanced after low GI at 150 & 210 min | |||||
| Benton & Nabb 2004 [136] | 323 (M = 21) | No breakfast vs. High-SAG biscuit, 50 g: 34 g CHO (7.9 g SAG, 18.8 g RAG, GI = 42) vs. Low-SAG cereal bar, 49 g: 34 g CHO (0.4 g SAG + 21.6 g RAG, GI = 66) or (0.05 g SAG + 21.10 g RAG) | Between (overnight fast) | O | − | Enhanced after low GI at 210 min | ||||
| Nabb & Benton, 2006b [139] | 189 (M = 20) | 8 breakfast conditions differing in energy (114–407 kcal), & contained either low or high levels of CHO (24 or 59 g), fat (1 or 16 g) or proteins (2 or 10 g) | Between (overnight fast) | O | O | O | Episodic: better glucose tolerance, low caloric intake & lower levels of blood glucose = enhanced performance. RT & vigilance: better glucose tolerance, higher levels of blood glucose = faster RT and better vigilance | |||
| Nabb & Benton, 2006a [138] | 168 (M = 20) | 8 breakfast conditions differing in contents of available CHO and dietary fiber: Low carb (15 g) with low or medium DF [100 mL milk vs. Medium CHO (30 g) with low, medium or high DF [200 mL milk] vs. High CHO (50 g) with low, medium or high DF [200 mL milk] | Between | O | Episodic: high carb meal + better glucose tolerance = forgot less words vs. poor glucose tolerance ppts. Poor glucose tolerance + low carb meal = forgot less words vs. high carb meal & poorer word recall after low vs. high fiber. Attention: better glucose tolerance + medium and high carb meals = faster RT (90 min) | |||||
| Smith & Foster, 2008 [6] | 36 (M = 15.6) | 30 g All-Bran (GI = 30) vs. 30 g Cornflakes (GI = 77). Served with 125 mL of milk | Between (overnight fast) | − a O a | Episodic: no effect on verbal learning. High GI = fewer items forgotten in long delay recall vs. short delay (vs. low GI) | |||||
| Micha et al. 2010 [135] | 60 (M = 13) | Classification of habitual breakfast intake into 4 groups: HIGH GL:low or high GI and LOW GL:low or high GI | Between (overnight fast) | O a,1 − b | − | O 2 | O 2 | O 3 | Fractionation of effects on specific cognitive tests by GL and GI breakfast forms. Enhancing effects in High GL forms which were associated with higher BG levels ~120 min post ingestion | |
| Author | Sample Size (Age) | Design (Within or Between Subjects) | Performance Measured (Relative to Drink Intake) | Drink (Volume/Vehicle) | Outcome Measures | Outcomes |
|---|---|---|---|---|---|---|
| Horne & Reyner, 2001 [179] | 11 (M = 24) | Within (restricted sleep (5 h); overnight caffeine fast) | 30 min drive–30 min break (drink)–2 h driving | (500 mL) caffeine 160 mg + 28.25 g CHO (11.3 g/100 mL) vs. placebo energy drink | Driving simulator (lane drifting and RT) | Caffeine + CHO significantly improved both lane drifting and RT. Effect strongest in 1st h |
| Warburton et al. 2001 [173] | Study 1: 20; Study 2: 22 (18–24) | Within (1 h caffeine abstinence) | +45 min | (250 mL) (Red Bull) 80 mg caffeine + 21 g sucrose + 5 g glucose +1 g taurine vs. Study 1: sugar-free water; Study 2: water + ~6 g glucose | RVIP; verbal reasoning; verbal and non-verbal memory test; Bond-Lader mood VAS | Energy drink improved attention, and verbal reasoning RT vs. glucose and non-glucose placebo, and reduced variability in RT performance. No difference between glucose and non-glucose drinks. No memory effects |
| Reyner & Horne, 2002 [182] | 12 (M = 24) | Within (overnight caffeine fast; restricted sleep (5 h)) | 30 min drive–30 min break (drink)–2 h driving | (250 mL) (Red Bull) 80 mg caffeine + 21 g sucrose + 5 g glucose vs. placebo version | Driving simulator (lane drifting and RT); EEG; Karolinska Sleepiness Scale | Caffeine + CHO = reduced sleep-related driving incidents and subjective sleepiness during the afternoon. Effect strongest in 1st 90 min |
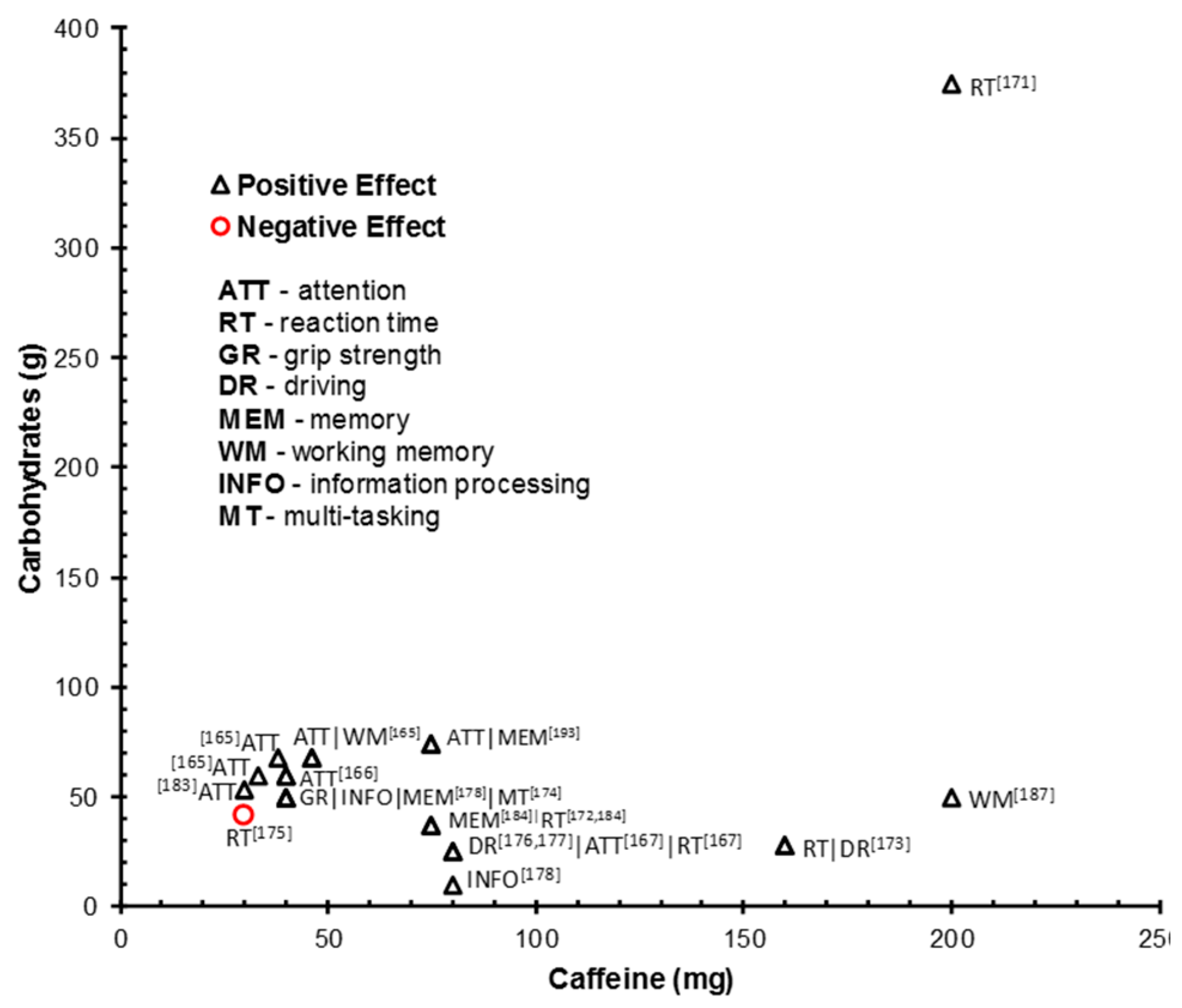
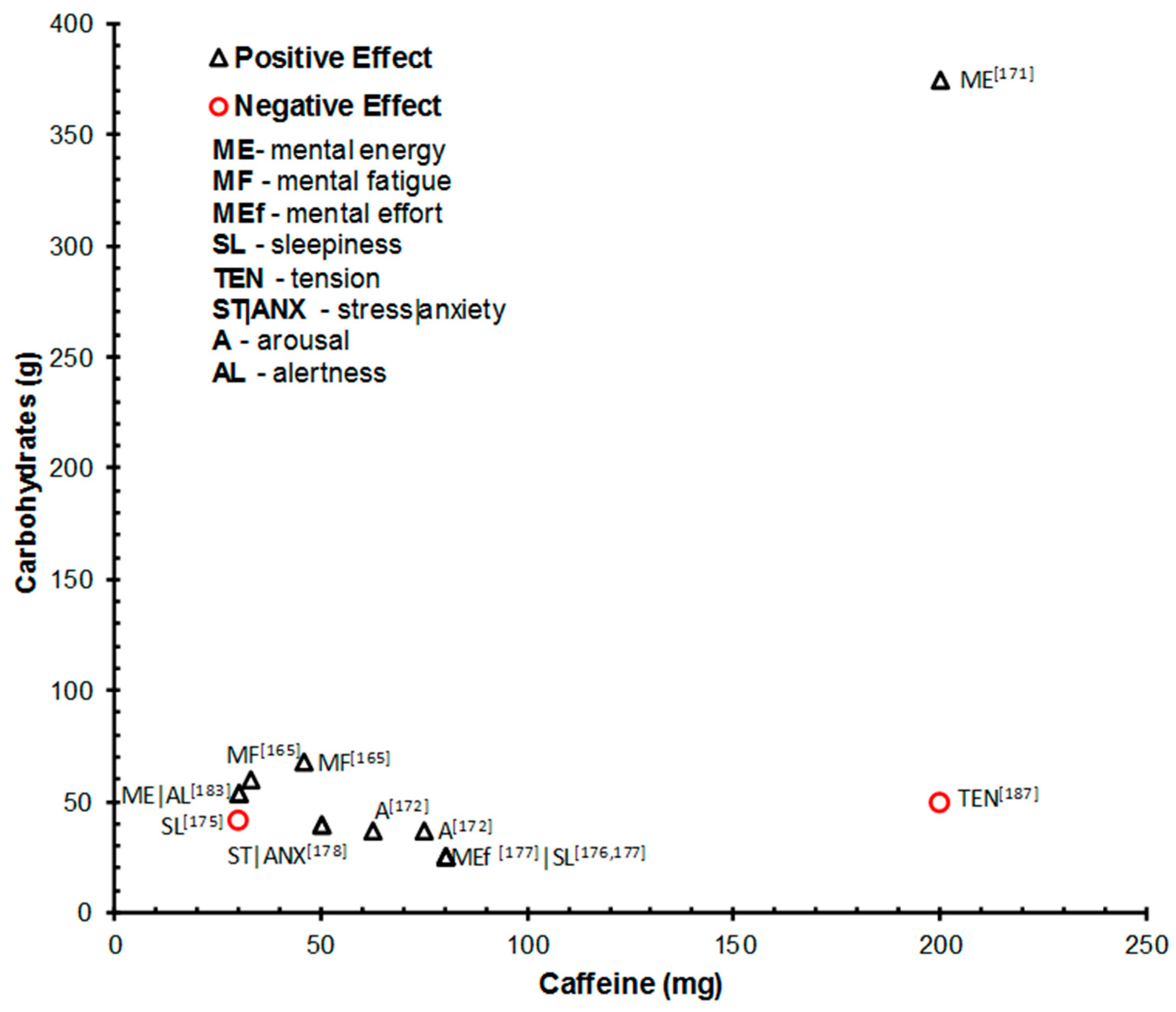
| Kennedy & Scholey, 2004 [171] | Study 1: 30 (18–25); Study 2: 26 (18–24) | Double-blind, placebo-controlled, cross-over design (24 h; overnight fast and caffeine abstinence) | +10 min | Study 1: (380-mL) 38 mg caffeine + 68 g glucose vs. 46 mg caffeine + 68 g of glucose, vs. vehicle placebo; Study 2: (330-mL) 33 mg caffeine + 60 g glucose vs. just the vehicle. | 10 min cognitive test battery × 6 times (=60 min cog. demand): Serial 3s and 7s; RVIP; mental fatigue VAS | Both studies: improved accuracy of RVIP performance with all 3 active treatments. Effects emerged + 35 (38g and 46g caffeine) and +45 (33g caffeine) min after drink intake. 46 mg caffeine drink improve WM in initial 2 blocks. Higher dose of caffeine (46 mg) and caffeine drink (33 mg) reduced self-assessed mental fatigue during the extended period of cognitive performance (no effect of 38 g = baseline effect?) |
| Smit et al. 2004 [178] | Study 1: 28 (18–49); Study 3: 97 (18–55) | Study 1: Within (overnight caffeine abstinence); Study 3: Between (CHO (breakfast) deprived) | +5–+90 min | (250 mL) Study 1: 75 mg caffeine + 37.5 g glucose vs. placebo vs. water; Study 3: 62.5 mg caffeine + 37.5 g glucose vs. 62.5 mg caffeine vs. 62.5 mg caffeine + 37.5 g glucose non-carbonated | Simple RT; RVIP; immediate and delayed word recall; letter search task; mood VAS | Caffeine + glucose drinks improved and/or maintained mood (arousal) and RT performance during fatiguing and cognitively demanding tasks relative to placebo |
| Rao et al. 2005 [172] | 40 (18–30) | Between (no fasting; caffeine abstinence on test day) | Not known | (330 mL) 40 mg caffeine + 60 g glucose syrup vs. sweetness/flavor matched placebo | BP; HR; EEG; ERP; sustained selective attention | Glucose + caffeine drink = improved accuracy and RT on sustained selective-attention task vs. placebo. Glucose + caffeine = improved stimulus processing at several stages of information processing (ERP) |
| Anderson & Horne, 2006 [181] | 10 (=22.4) | Double blind, crossover design (1 week; restricted sleep (5 h); taken with soup lunch; ~14 h caffeine abstinence) | +10 min | (250 mL) 30 mg caffeine + 42 g sugars (glucose, fructose, sucrose) vs. sugar- caffeine-free orange flavored drink | Psychomotor Vigilance Test; Karolinska Sleepiness Scale | Energy drink did not counteract sleepiness and = slower RTs and more lapses 80 min post-intake |
| Smit et al. 2006 [184] | 76 (18–40) | Between (overnight food and caffeine fast) | +7–+120 min | (330 mL) Familiar drink: 30 mg caffeine + 54 g glucose vs. familiar drink placebo vs. Novel drink: 30 mg caffeine + 54 g glucose vs. novel drink placebo | Simple RT; RVIP; serial 7’s; letter search task; mood VAS | First exposure: familiar drink and its placebo improved alertness, mental energy and mental performance vs. baseline and novel placebo drink. Repeated exposure/increased familiarity with the novel drinks: both caffeine + CHO containing drinks = sustained beneficial effects vs. placebo drinks and baseline measures |
| Childs & de Wit, 2008 [177] | 35 (18–35) | Within (caffeine abstinence on test day) | Remained awake 5 p.m.–5 a.m. Energy capsule or placebo 3:30 a.m. Cog. testing +30 min | (Capsule) 200 mg caffeine + 50 mg white willow bark + 30 mg magnesium oxide + 10 mg taurine + 375 g dextrose vs. 375 g dextrose placebo | BP; physical activity meter; Simple and choice RT task; POMS and mood VAS | Caffeine = improved mood and mental energy and counteracted increases in simple and choice RT vs. placebo |
| Gendle et al. 2009 [175] | 36 (18–21) | Within (4 h fast and caffeine abstinence) | +30 min | (250 mL) 80 mg caffeine + 1000 mg taurine + 27 g glucose/sucrose vs. sugar and caffeine free version | Visual attention and RT (Conner’s Continuous Performance Test II) | No effects |
| Howard & Marczinski, 2010 [176] | 80 (M = 20.1) | Between (2 h fast; 8 h caffeine abstinence) | +30 min | Energy drink doses calculated by body weight. Caffeine content for average 78 kg ppt given in (): 1.8 mL/kg energy drink (45.6 mg) vs. 3.6 mL/kg energy drink (91.2 mg/30.8 g CHO) vs. 5.4 mL/kg energy drink (136.7 mg) vs. 3.6 mL/kg placebo drink (29.3 g CHO) vs. no drink) | Cued go/no-go task; mental fatigue VAS | Energy drink = increased stimulation, decreased mental fatigue, and decrease behavioral control RT. No effect on response inhibition. Lowest caffeine dose = greater RT and subjective measure improvement. Improvements diminished as the dose increased |
| Mets et al. 2011 [183] | 24 (M = 21–35) | Within | Drive 2 h–drink intake–drive 2h | (250 mL) (Red Bull) 80 mg caffeine + 21 g sucrose + 5 g glucose + 1 g taurine + vs. placebo (Red bull) drink | STISIM Drive™ driving simulator (standard deviation of lateral position (SDLP); standard deviation of speed); subjective driving quality and mental effort; Karolinska Sleepiness Scale | Energy drink significantly improved driving relative to placebo: SDLP reduced in 3rd and 4th h. Reduced standard deviation of speed, improved subjective driving quality, and reduced mental effort during 3rd hr. Subjective sleepiness was significantly decreased in 3rd and 4th h of driving |
| Aniţei et al. 2011 [174] | 153 (18–21) | Between | +40 min | 275 mg caffeine coffee vs. energy drink (1000 mg taurine + 80 mg caffeine + sucrose/glucose (not stated) vs. 275 mg caffeine + energy drink vs. no drink | Perceptual speed; visual and auditory attention RT; visual orientation performance; vigilance test | Caffeine alone and combined with CHO in energy drink increased motor reactivity, short-term attention (under 30 min) and visual attention RT. Effects less consistent/smaller when caffeine and energy drink combined (365 mg caffeine) |
| Sünram-Lea et al. 2012 [185] | 81 (M = 26) | Between (overnight fast + standardized breakfast; caffeine abstinence from waking) | +10 (pre-stressor) and +60 min (post-stressor) | (330-mL) 40 mg caffeine + 50 g glucose vs. 80 mg caffeine + 10.25 g fructose (41%)/glucose (59%) vs. placebo drink | Salivary cortisol; CBG; immediate and delayed free word call; letter cancellation task; grammatical reasoning task; letter digit substitution task; hand grip strength | 50 g glucose +40 mg caffeine =increased grip strength and improved memory performance. Both active drinks = improved information processing (letter-digit substitution task) performance vs. placebo. 50 g glucose/40 mg caffeine = reduced anxiety and subjective stress. No effects on reasoning and attention or subjective alertness |
| Scholey et al. 2014 [180] | 150 (18–55) | Between (12 h fast and caffeine abstinence) | +30 min | (330 mL) 40 mg caffeine + 60 g glucose vs. 25 g glucose vs. 60 g glucose | CBG; salivary caffeine level; multi-tasking framework (4 simultaneous tasks: mathematical processing task; stroop; memory search; target tracker task); Bond–Lader mood VAS; stress and fatigue VAS | Co-administration of glucose and caffeine = greater multi-tasking performance than placebo or glucose alone |
| Author | Sample Size (Age) | Design (Within or Between Subjects) | Performance Measured (Relative to Drink Intake) | Drink [Volume/Vehicle] | Outcome Measures | Outcomes | Interactive Effect of Caffeine and Glucose |
|---|---|---|---|---|---|---|---|
| Smit et al. 2004 [178] | Study 2: 146 (18–54) | Between (overnight caffeine abstinence) | +5–+90 min | 75 mg caffeine + 37.5 g glucose vs. 37.5 g glucose vs. 75 mg caffeine vs. 75 mg caffeine + 37.5 g glucose non-carbonated | Simple RT; RVIP; immediate and delayed word recall; letter search task; mood VAS | Main treatment effect suggesting caffeine = main component associated with improved simple RT and increased arousal; comparatively minor, weak effects of CHO demonstrated | x |
| Scholey & Kennedy, 2004 [190] | 20 (18–32) | Within (overnight fast; morning coffee abstinence) | +30 min | (250 mL) Placebo (artificially flavored and sweetened water vehicle) vs. vehicle + 75 mg caffeine vs. vehicle + 37.5 g glucose vs. vehicle + flavoring levels of herbs (12.5 mg ginseng extract and 2.004 mg ginkgo biloba extract) vs. complete energy drink (75 mg caffeine, 37.5 g glucose + flavoring levels of herbs) | CBG; HR; Digit Symbol Substitution Task; CDR (immediate and delayed word + picture recall and recognition; Simple and choice RT; digit vigilance; spatial and numeric WM. Factor analyzed for global “quality of memory” outcomes; Bond-Lader mood VAS; POMS | No effect of glucose/caffeine/herbs in isolation. Whole drink = improved “Secondary memory” (combined % accuracy scores delayed word recognition, delayed picture recognition, immediate word recall and delayed word recall) and “speed of attention” performance vs. placebo (only) | √ |
| Maridakis et al. 2009 [195] | 17 (M = 23.8) | Within (8 h fast) | ~+30 min | (Capsule) 200 mg caffeine + 50 g CHO (white bread) vs. 200 mg caffeine vs. 50 g CHO vs. placebo capsule vs. 50 g CHO + placebo pill | CPT; BAKAN; POMS; Activation-Deactivation Checklist; State-Trait Energy and Fatigue scales | Caffeine improved attention. No additional performance benefit of adding CHO. Caffeine increase energy, lowered fatigue. No additional benefit of adding CHO. CHO in isolation = less effects on mood | x |
| Adan & Serra-Grabulosa, 2010 [191] | 72 (18–25) | Between (8 h fast; 18 h caffeine abstinence) | +30 min | (150 mL) water vs. water + 75 mg caffeine vs. water + 75 g glucose vs. water + 75 mg caffeine/75 g glucose | CBG; salivary caffeine level; RAVLT; Purdue-Pegboard; Benton Judgement of Line Orientation Test (visuo-spatial function); CCAP (attention, RT and visual scanning speed); digit span; mood VAS | Caffeine + glucose = beneficial effects on attention (sequential RT tasks) and verbal memory learning and consolidation (not shown by ingredients in isolation). Caffeine alone = improved simple RT. Glucose alone = improved simple and sequential RT tasks and manual dexterity assembly task. | √ |
| Serra-Grabulosa et al. 2010 [192] | 40 (18–25) | Between (8 h fast; 12 h caffeine abstinence) | +30 min | (150 mL) Water + 75 g glucose vs. water + 75 mg caffeine vs. water + 75 g glucose/75 mg caffeine | CBG; salivary caffeine level; CPT (sustained attention); fMRI | No effect of drink on cognitive performance. Glucose + caffeine = decreased activation in the bilateral parietal and left prefrontal cortex (areas associated with sustained attention and WM processes). Interpreted as increased efficiency of the attentional system | √ |
| Giles et al. 2012 [193] | 48 (M = 20.08) | Mixed (standardized meal +2 h fast; 24 h caffeine abstinence) | +30 min (WM); +60 min (RT) | (Capsule) Within (all P’s): 200 mg caffeine/0 mg taurine vs. 0 mg caffeine/2000 mg taurine vs. 200 mg caffeine/2000 mg taurine vs. 0 mg caffeine/0 mg taurine; [500 mL] Between (50:50 sample split) 50 g glucose vs. 50 g stevia | HR; Attention network test (alerting, orienting, executive control); N-back task; simple and choice RT; salivary cortisol; POMS | Caffeine = most consistent effects on cognitive performance. Glucose slowed RT. Glucose + caffeine enhanced object WM. Glucose + taurine, enhanced orienting attention. Taurine = selective effects (+ive at high load). Caffeine reduced headache symptoms and tiredness and increased alertness. Caffeine reduced fatigue and increased feelings of tension and vigor. Glucose potentiated caffeine-induced feelings of tension. Taurine intake opposed caffeine effects on mood | x |
| Young & Benton 2013 [194] | 345 (M = 21.78) | Between (2 h fast) | +30; +90; +150 min | (250 mL) yoghurt (GL = 3.6) + no caffeine vs. yoghurt (GL = 3.6) + 80 mg caffeine vs. 39 g glucose (GL = 30) + no caffeine vs. 39 g glucose (GL = 30) + 80 mg caffeine vs. flavored water + no caffeine vs. flavored water + 80 mg caffeine | CBG and CGMS (subsample n = 38); immediate and delayed word recall; choice RT; serial sevens; arrow flankers (selective attention); vigilance/sustained attention; POMS | Caffeine, irrespective of vehicle, = better memory, faster RT (choice reaction time test and WM) and increased vigilance. Greater subjective energy reported 30 min after consuming caffeine and water, vs. water alone; after 90 and 150 min caffeine administered in water increased tiredness, hostility and confusion. Combining caffeine with a yoghurt-based drink increased energy, agreeableness and clear-headedness later in the morning. No effects of caffeine + glucose on mood | x |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyle, N.B.; Lawton, C.L.; Dye, L. The Effects of Carbohydrates, in Isolation and Combined with Caffeine, on Cognitive Performance and Mood—Current Evidence and Future Directions. Nutrients 2018, 10, 192. https://doi.org/10.3390/nu10020192
Boyle NB, Lawton CL, Dye L. The Effects of Carbohydrates, in Isolation and Combined with Caffeine, on Cognitive Performance and Mood—Current Evidence and Future Directions. Nutrients. 2018; 10(2):192. https://doi.org/10.3390/nu10020192
Chicago/Turabian StyleBoyle, Neil Bernard, Clare Louise Lawton, and Louise Dye. 2018. "The Effects of Carbohydrates, in Isolation and Combined with Caffeine, on Cognitive Performance and Mood—Current Evidence and Future Directions" Nutrients 10, no. 2: 192. https://doi.org/10.3390/nu10020192
APA StyleBoyle, N. B., Lawton, C. L., & Dye, L. (2018). The Effects of Carbohydrates, in Isolation and Combined with Caffeine, on Cognitive Performance and Mood—Current Evidence and Future Directions. Nutrients, 10(2), 192. https://doi.org/10.3390/nu10020192