Vitamin D-Binding Protein Polymorphisms, 25-Hydroxyvitamin D, Sunshine and Multiple Sclerosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.1.1. Case Identification
2.1.2. Control Selection
2.2. Data Collection
2.3. Genotyping
2.4. Statistical Analysis
2.4.1. Sun Exposure
2.4.2. 25OHD and Genotype
2.4.3. 25OHD, UVR, Vitamin D-Binding Protein Genotype and MS
3. Results
3.1. Characteristics of Participants
3.2. Polymorphisms in the Vitamin D-Binding Protein Gene and 25OHD
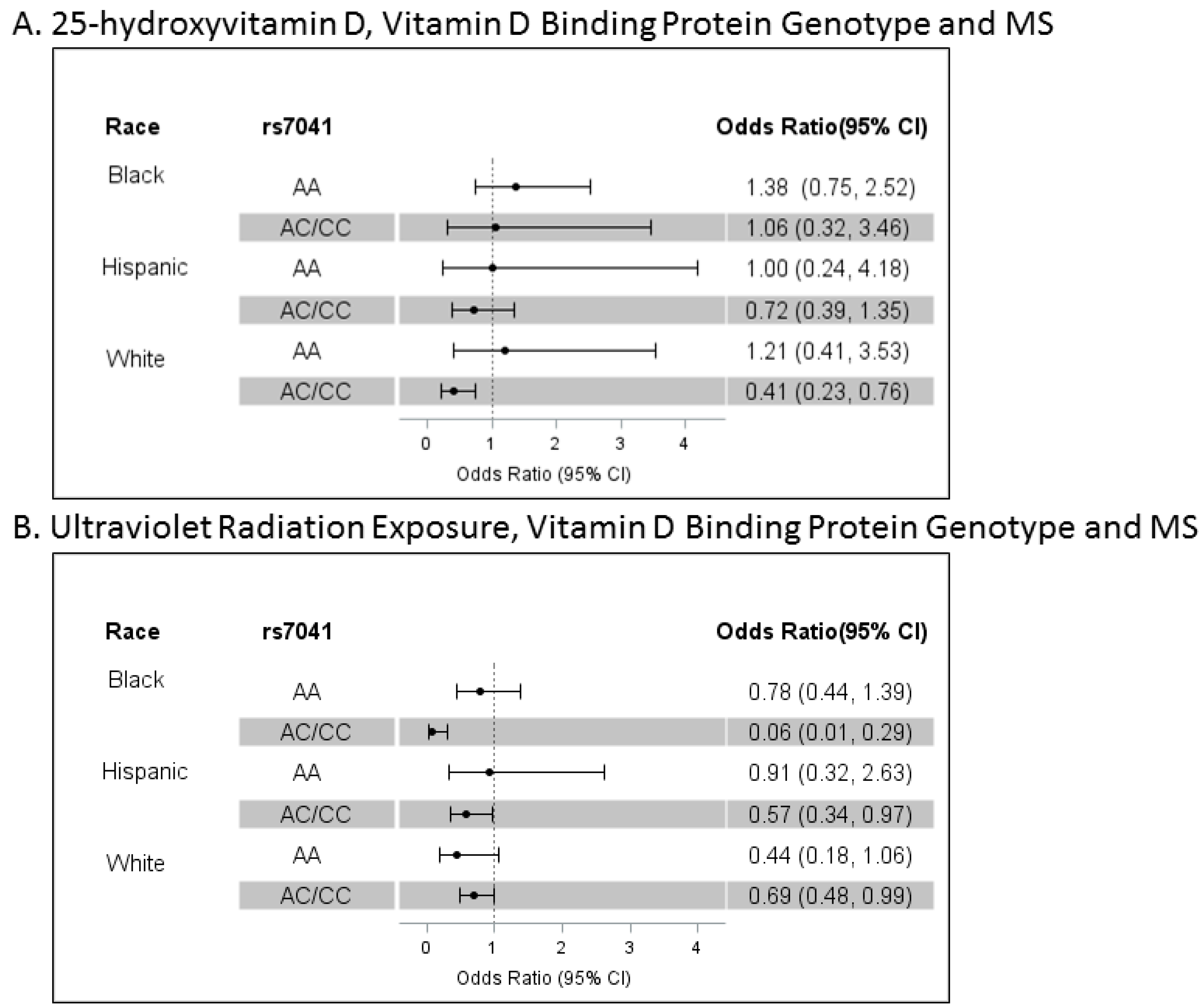
3.3. Polymorphisms in the Vitamin D-Binding Protein Gene, 25OHD, Cumulative Lifetime UVR and MS
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
Appendix A.1. Supplementary Methods
Appendix A.1.1. Study Population, Case and Control Identification and Recruitment
Appendix A.1.2. Genotyping
Appendix A.2. Ancestry Structure
Appendix A.3. Deseasonalized 25OHD Values for Other Analyses
Appendix A.4. Cumulative Lifetime UVR
Appendix A.5. Supplementary Results

{kind=link}
| Race/Ethnicity | SNP | n | Reference Allele | Variant Allele | Variant Allele Frequency | Β Estimate * | Change in Total 25OHD Level Per Variant Allele Copy (95% CI), nmol/L | p |
|---|---|---|---|---|---|---|---|---|
| Black | rs7041 | 217 | C | A | 0.825 | −0.1322 | −6.3 (−13.1 to −0.4) | 0.0358 |
| Hispanic | rs7041 | 347 | C | A | 0.488 | −0.0490 | −2.8 (−5.9 to 0.3) | 0.0733 |
| White | rs7041 | 433 | C | A | 0.441 | −0.0675 | −4.9 (−8.4 to −1.5) | 0.0044 |
| Black | rs4588 | 217 | G | T | 0.124 | −0.2290 | −9.2 (−13.7 to −4.1) | 0.0009 |
| Hispanic | rs4588 | 347 | G | T | 0.232 | −0.0933 | −4.9 (−8.1 to −1.5) | 0.0052 |
| White | rs4588 | 433 | G | T | 0.281 | −0.1191 | −7.9 (−10.9 to −4.6) | <0.0001 |
References
- Lucas, R.M.; Ponsonby, A.L.; Dear, K.; Valery, P.C.; Pender, M.P.; Taylor, B.V.; Kilpatrick, T.J.; Dwyer, T.; Coulthard, A.; Chapman, C.; et al. Sun exposure and vitamin d are independent risk factors for cns demyelination. Neurology 2011, 76, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Van der Mei, I.A.; Ponsonby, A.L.; Dwyer, T.; Blizzard, L.; Simmons, R.; Taylor, B.V.; Butzkueven, H.; Kilpatrick, T. Past exposure to sun, skin phenotype, and risk of multiple sclerosis: Case-control study. BMJ 2003, 327, 316. [Google Scholar] [CrossRef] [PubMed]
- Kampman, M.T.; Wilsgaard, T.; Mellgren, S.I. Outdoor activities and diet in childhood and adolescence relate to ms risk above the arctic circle. J. Neurol. 2007, 254, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Baarnhielm, M.; Hedstrom, A.K.; Kockum, I.; Sundqvist, E.; Gustafsson, S.A.; Hillert, J.; Olsson, T.; Alfredsson, L. Sunlight is associated with decreased multiple sclerosis risk: No interaction with human leukocyte antigen-drb1*15. Eur. J. Neurol. 2012, 19, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Munger, K.L.; Levin, L.I.; Hollis, B.W.; Howard, N.S.; Ascherio, A. Serum 25-hydroxyvitamin d levels and risk of multiple sclerosis. JAMA 2006, 296, 2832–2838. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.; Hallmans, G.; Nystrom, M.; Stenlund, H.; Wadell, G.; Sundstrom, P. Vitamin d as a protective factor in multiple sclerosis. Neurology 2012, 79, 2140–2145. [Google Scholar] [CrossRef] [PubMed]
- Langer-Gould, A.M.; Lucas, R.M.; Xiang, A.H.; Chen, L.H.; Wu, J.; Gonzales, E.; Haraszti, S.; Smith, J.B.; Quach, H.; Barcellos, L.F. Sun exposure but not vitamin d is associated with multiple sclerosis risk in blacks and hispanics. Nutrients 2017, in press. [Google Scholar]
- Powe, C.E.; Evans, M.K.; Wenger, J.; Zonderman, A.B.; Berg, A.H.; Nalls, M.; Tamez, H.; Zhang, D.; Bhan, I.; Karumanchi, S.A.; et al. Vitamin d-binding protein and vitamin d status of black americans and white americans. N. Engl. J. Med. 2013, 369, 1991–2000. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.H.; Powe, C.E.; Evans, M.K.; Wenger, J.; Ortiz, G.; Zonderman, A.B.; Suntharalingam, P.; Lucchesi, K.; Powe, N.R.; Karumanchi, S.A.; et al. 24,25-dihydroxyvitamin d3 and vitamin d status of community-dwelling black and white americans. Clin. Chem. 2015, 61, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, M.M.; Speeckaert, R.; van Geel, N.; Delanghe, J.R. Vitamin d binding protein: A multifunctional protein of clinical importance. Adv. Clin. Chem. 2014, 63, 1–57. [Google Scholar] [PubMed]
- Koebnick, C.; Langer-Gould, A.M.; Gould, M.K.; Chao, C.R.; Iyer, R.L.; Smith, N.; Chen, W.; Jacobsen, S.J. Do the sociodemographic characteristics of members of a large, integrated health care system represent the population of interest? Perm. J. 2012, 16, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Langer-Gould, A.; Zhang, J.L.; Chung, J.; Yeung, Y.; Waubant, E.; Yao, J. Incidence of acquired cns demyelinating syndromes in a multiethnic cohort of children. Neurology 2011, 77, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Langer-Gould, A.; Brara, S.M.; Beaber, B.E.; Zhang, J.L. Incidence of multiple sclerosis in multiple racial and ethnic groups. Neurology 2013, 80, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the mcdonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Barnes, G.; Benjamin, S.; Bowen, J.D.; Cutter, N.; De Lateur, B.J.; Dietrich, W.D.; Dowling, M.M.; Griffin, J.W.; Hummers, L.; Irani, D.; et al. Proposed diagnostic criteria and nosology of acute transverse myelitis. Neurology 2002, 59, 499–505. [Google Scholar]
- Jacob, A.; Weinshenker, B.G. An approach to the diagnosis of acute transverse myelitis. Semin. Neurol. 2008, 28, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- King, L.; Xiang, F.; Swaminathan, A.; Lucas, R.M. Measuring sun exposure in epidemiological studies: Matching the method to the research question. J. Photochem. Photobiol. B 2015, 153, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Rito, Y.; Flores, J.; Fernandez Aguilar, A.; Escalante Membrillo, C.; Gutierrez Lanz, E.; Barboza, M.A.; Rivas Alonso, V.; Trevino Frenk, I.; Corona Vazquez, T. Vitamin d in multiple sclerosis patients: Not the same risk for everybody. Mult. Scler. 2016, 22, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Amezcua, L.; Chung, R.H.; Conti, D.V.; Langer-Gould, A.M. Vitamin d levels in hispanics with multiple sclerosis. J. Neurol. 2012, 259, 2565–2570. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.E.; Byrne, S.N. The immunologic revolution: Photoimmunology. J. Investig. Dermatol. 2012, 132, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Marling, S.J.; Beaver, E.F.; Severson, K.S.; Deluca, H.F. Uv light selectively inhibits spinal cord inflammation and demyelination in experimental autoimmune encephalomyelitis. Arch. Biochem. Biophys. 2015, 567, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.H.; Lucas, R.M.; Booth, D.R.; Carroll, W.M.; Nolan, D.; Cole, J.M.; Jones, A.P.; Kermode, A.G. Narrowband UVB phototherapy for clinically isolated syndrome: A trial to deliver the benefits of vitamin d and other UVB-induced molecules. Front. Immunol. 2017, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Becklund, B.R.; Severson, K.S.; Vang, S.V.; DeLuca, H.F. Uv radiation suppresses experimental autoimmune encephalomyelitis independent of vitamin d production. Proc. Natl. Acad. Sci. USA 2010, 107, 6418–6423. [Google Scholar] [CrossRef] [PubMed]
- Autier, P.; Boniol, M.; Pizot, C.; Mullie, P. Vitamin d status and ill health—Author’s reply. Lancet Diabetes Endocrinol. 2014, 2, 275–276. [Google Scholar] [CrossRef]
- Bischoff-Ferrari, H.A.; Dawson-Hughes, B.; Orav, E.J.; Staehelin, H.B.; Meyer, O.W.; Theiler, R.; Dick, W.; Willett, W.C.; Egli, A. Monthly high-dose Vitamin D treatment for the prevention of functional decline: A randomized clinical trial. JAMA Intern. Med. 2016, 176, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Litonjua, A.A.; Carey, V.J.; Laranjo, N.; Harshfield, B.J.; McElrath, T.F.; O’Connor, G.T.; Sandel, M.; Iverson, R.E., Jr.; Lee-Paritz, A.; Strunk, R.C.; et al. Effect of prenatal supplementation with vitamin d on asthma or recurrent wheezing in offspring by age 3 years: The vdaart randomized clinical trial. JAMA 2016, 315, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, J.M.; Cree, B.A.; McElroy, J.; Oksenberg, J.; Green, R.; Mowry, E.M.; Miller, J.W.; Hauser, S.L.; Green, A.J. Vitamin d in african americans with multiple sclerosis. Neurology 2011, 76, 1824–1830. [Google Scholar] [CrossRef] [PubMed]
- Kamboh, M.I.; Ferrell, R.E. Ethnic variation in vitamin d-binding protein (GC): A review of isoelectric focusing studies in human populations. Hum. Genet. 1986, 72, 281–293. [Google Scholar] [CrossRef] [PubMed]
| Black (n = 247) | Hispanic (n = 380) | White (n = 514) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Case (n = 116) | Control (n = 131) | p * | Case (n = 183) | Control (n = 197) | p * | Case (n = 247) | Control (n = 267) | p * | |
| Age, mean (SD), y | 38.4 (12.8) | 38.5 (13.0) | 0.93 | 32.5 (10.7) | 32.6 (11.1) | 0.95 | 39.7 (12.0) | 39.9 (12.2) | 0.87 |
| Female, n (%) | 92 (79.3) | 103 (78.6) | 0.90 | 132 (72.1) | 145 (73.6) | 0.75 | 164 (66.4) | 174 (65.2) | 0.77 |
| Smoking, n (%) | 27 (23.3) | 33 (25.2) | 0.73 | 44 (24.0) | 38 (19.3) | 0.26 | 115 (46.6) | 90 (33.7) | 0.003 |
| BMI, mean (SD) | 30.5 (7.4) | 31.6 (8.1) | 0.26 | 29.5 (6.8) | 30.1 (7.0) | 0.42 | 28.5 (6.7) | 28.5 (6.9) | 0.95 |
| VitD supplement users *, n (%) | 28 (24.1) | 2 (1.5) | <0.001 | 30 (16.4) | 3 (1.5) | <0.001 | 62 (25.1) | 19 (7.1) | <0.001 |
| Season **, n (%) | 56 (48.3) | 65 (49.6) | 0.83 | 85 (46.5) | 104 (52.8) | 0.22 | 116 (47.0) | 142 (53.2) | 0.16 |
| cumulative UV dose ***, 1000 KJ/m2, mean (SD) | 1.43 (0.77) | 1.63 (0.79) | 0.05 | 1.14 (0.61) | 1.22 (0.68) | 0.22 | 1.53 (0.79) | 1.64 (0.83) | 0.15 |
| serum 25OHD, nmol/L, median (IQR) | 43.7 (32.4,74.9) | 47.4 (30.0,64.9) | 0.53 | 54.9 (42.4,69.9) | 57.4 (44.9,69.9) | 0.35 | 67.4 (49.9,87.4) | 72.4 (59.9,92.4) | 0.003 |
| Vitamin D-Binding Protein | |||||||||
| rs7041, n (%) | 0.53 | 0.64 | 0.37 | ||||||
| AA | 79 (68.1) | 94 (71.7) | 40 ( 21.9) | 47 (23.8) | 56 (22.7) | 52 (19.4) | |||
| AC | 32 (27.6) | 31 (23.7) | 93 (50.8) | 100 (50.8) | 116 (47.0) | 123 (46.1) | |||
| CC | 5 (4.3) | 6 (4.6) | 50 (27.3) | 50 (25.4) | 75 (30.3) | 92 (34.5) | |||
| rs4588, n (%) | 0.13 | 0.52 | 0.44 | ||||||
| GG | 96 (82.8) | 98 (74.8) | 110 (60.1) | 112 (56.8) | 122 (49.4) | 141 (52.8) | |||
| GT | 18 (15.5) | 29 (22.1) | 66 (36.1) | 75 (38.1) | 105 (42.5) | 99 (37.1) | |||
| TT | 2 (1.7) | 4 (3.1) | 7 (3.8) | 10 (5.1) | 20 (8.1) | 27 (10.1) | |||
| HLA-DRB1*1501, n (%) | 0.20 | 0.0002 | <0.0001 | ||||||
| GG | 95 (81.9) | 115 (87.8) | 134 (73.2) | 174 (88.3) | 136 (55.1) | 214 (80.1) | |||
| AG | 21 (18.1) | 15 (11.5) | 47 (25.7) | 19 (9.7) | 92 (37.2) | 52 (19.5) | |||
| AA | 0 (0.0) | 1 (0.8) | 2 (1.1) | 4 (2.0) | 19 (7.7) | 1 (0.4) | |||
| Ancestry % median (IQR) | |||||||||
| European | 21.2 (14.6, 28.0) | 22.8 (16.3, 31.9) | 58.6 (50.1, 66.0) | 56.6 (48.5, 64.3) | 99.0 (97.6, 99.7) | 99.1 (97.6, 99.8) | |||
| African | 77.6 (69.8, 84.4) | 75.4 (66.2, 83.0) | 5.1 (3.8, 6.9) | 4.9 (3.8, 6.4) | 0.1 (0.0, 0.9) | 0.0 (0.0, 0.8) | |||
| Amerindian | 0.3 (0.1, 1.4) | 0.7 (0.2, 1.9) | 35.7 (27.0,42.8) | 38.0 (31.0, 46.1) | 0.5 (0.2, 1.1) | 0.4 (0.2, 1.2) | |||
| Race/Ethnicity | SNP | n | Reference Allele | Variant Allele | Variant Allele Frequency | β Estimate * | Change in Total 25OHD Level Per Variant Allele Copy (95% CI), nmol/L | p |
|---|---|---|---|---|---|---|---|---|
| Black | rs7041 | 247 | C | A | 0.828 | −0.1136 | −5.5 (−11.9 to 0.2) | 0.0573 |
| Hispanic | rs7041 | 380 | C | A | 0.483 | −0.0492 | −2.7 (−5.9 to 0.3) | 0.0757 |
| White | rs7041 | 514 | C | A | 0.443 | −0.0813 | −5.9 (−9.3 to −2.7) | 0.0002 |
| Black | rs4588 | 247 | G | T | 0.119 | −0.2148 | −8.9 (−13.5 to −3.7) | 0.0014 |
| Hispanic | rs4588 | 380 | G | T | 0.230 | −0.0836 | −4.4 (−7.6 to −0.9) | 0.0135 |
| White | rs4588 | 514 | G | T | 0.290 | −0.1160 | −7.7 (−10.5 to −4.7) | <0.0001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langer-Gould, A.; Lucas, R.M.; Xiang, A.H.; Wu, J.; Chen, L.H.; Gonzales, E.; Haraszti, S.; Smith, J.B.; Quach, H.; Barcellos, L.F. Vitamin D-Binding Protein Polymorphisms, 25-Hydroxyvitamin D, Sunshine and Multiple Sclerosis. Nutrients 2018, 10, 184. https://doi.org/10.3390/nu10020184
Langer-Gould A, Lucas RM, Xiang AH, Wu J, Chen LH, Gonzales E, Haraszti S, Smith JB, Quach H, Barcellos LF. Vitamin D-Binding Protein Polymorphisms, 25-Hydroxyvitamin D, Sunshine and Multiple Sclerosis. Nutrients. 2018; 10(2):184. https://doi.org/10.3390/nu10020184
Chicago/Turabian StyleLanger-Gould, Annette, Robyn M. Lucas, Anny H. Xiang, Jun Wu, Lie H. Chen, Edlin Gonzales, Samantha Haraszti, Jessica B. Smith, Hong Quach, and Lisa F. Barcellos. 2018. "Vitamin D-Binding Protein Polymorphisms, 25-Hydroxyvitamin D, Sunshine and Multiple Sclerosis" Nutrients 10, no. 2: 184. https://doi.org/10.3390/nu10020184
APA StyleLanger-Gould, A., Lucas, R. M., Xiang, A. H., Wu, J., Chen, L. H., Gonzales, E., Haraszti, S., Smith, J. B., Quach, H., & Barcellos, L. F. (2018). Vitamin D-Binding Protein Polymorphisms, 25-Hydroxyvitamin D, Sunshine and Multiple Sclerosis. Nutrients, 10(2), 184. https://doi.org/10.3390/nu10020184