Guarana (Paullinia cupana) Stimulates Mitochondrial Biogenesis in Mice Fed High-Fat Diet



Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
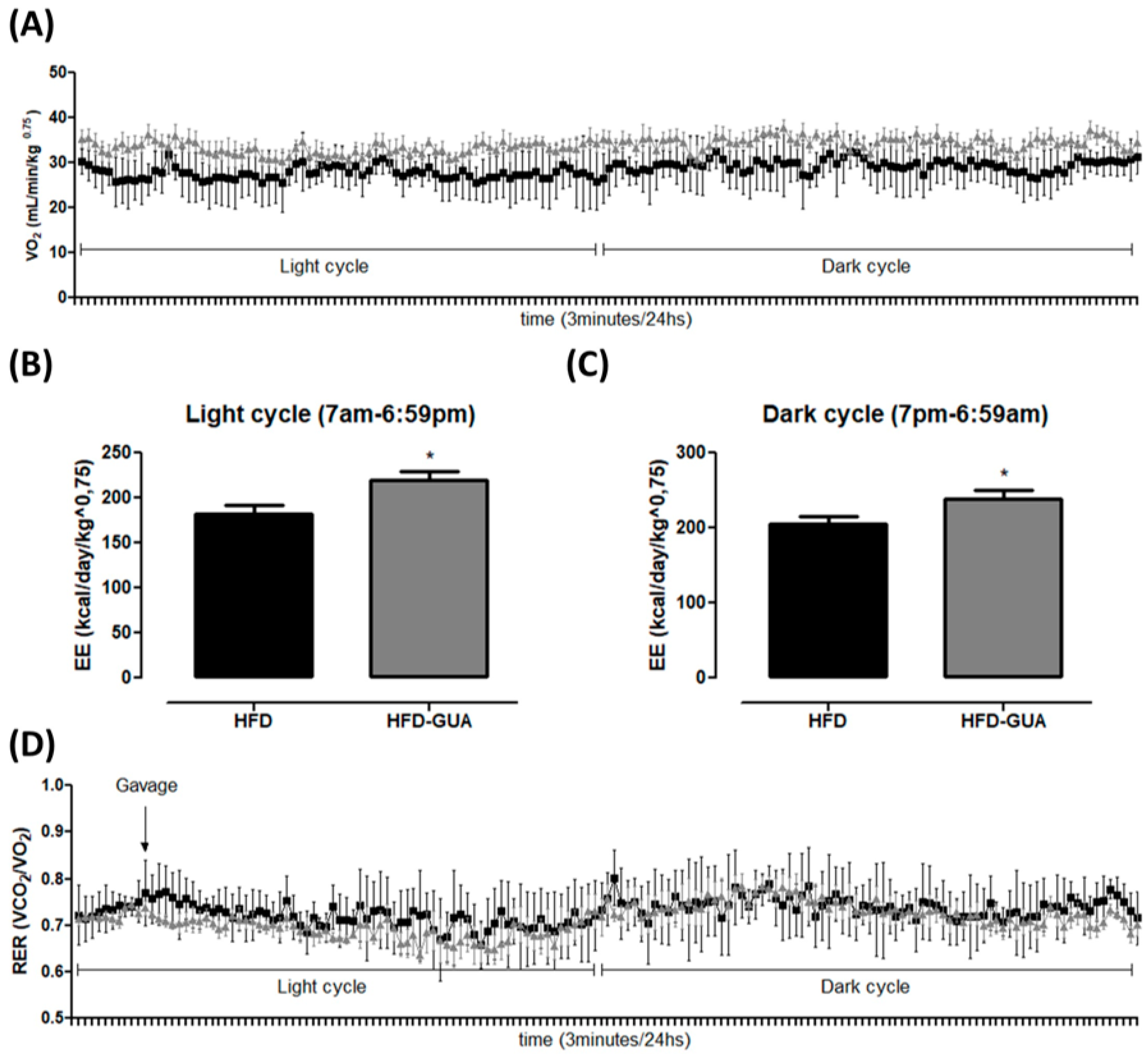
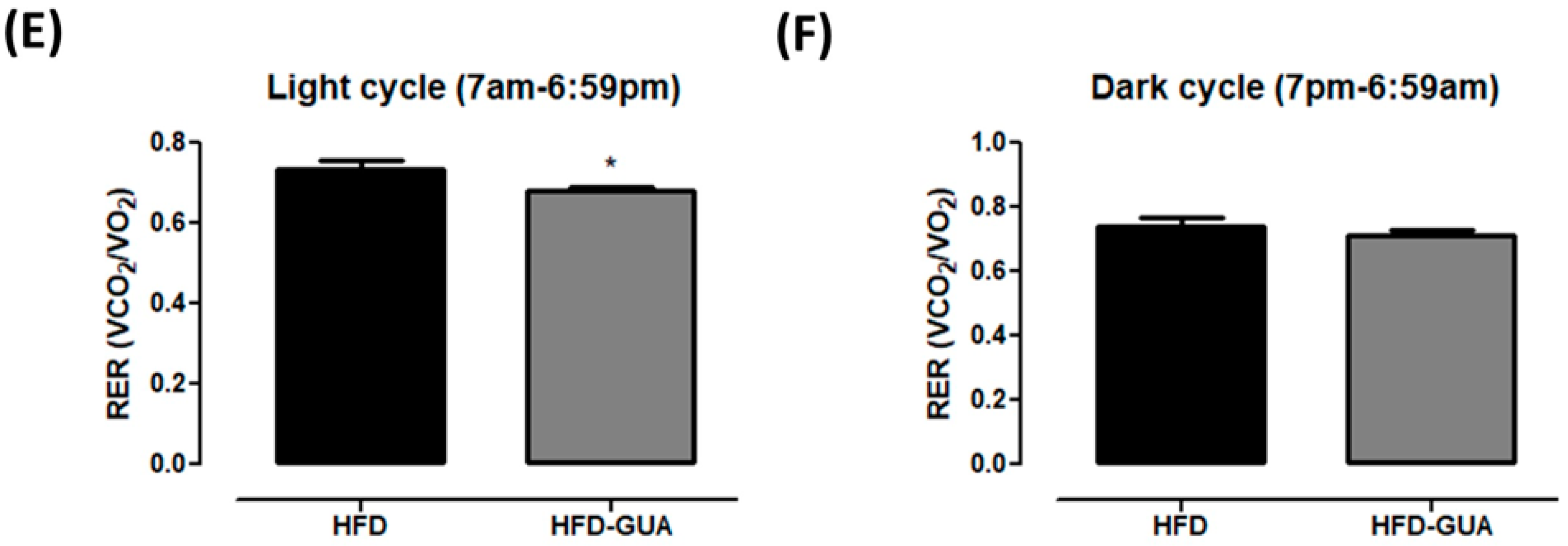
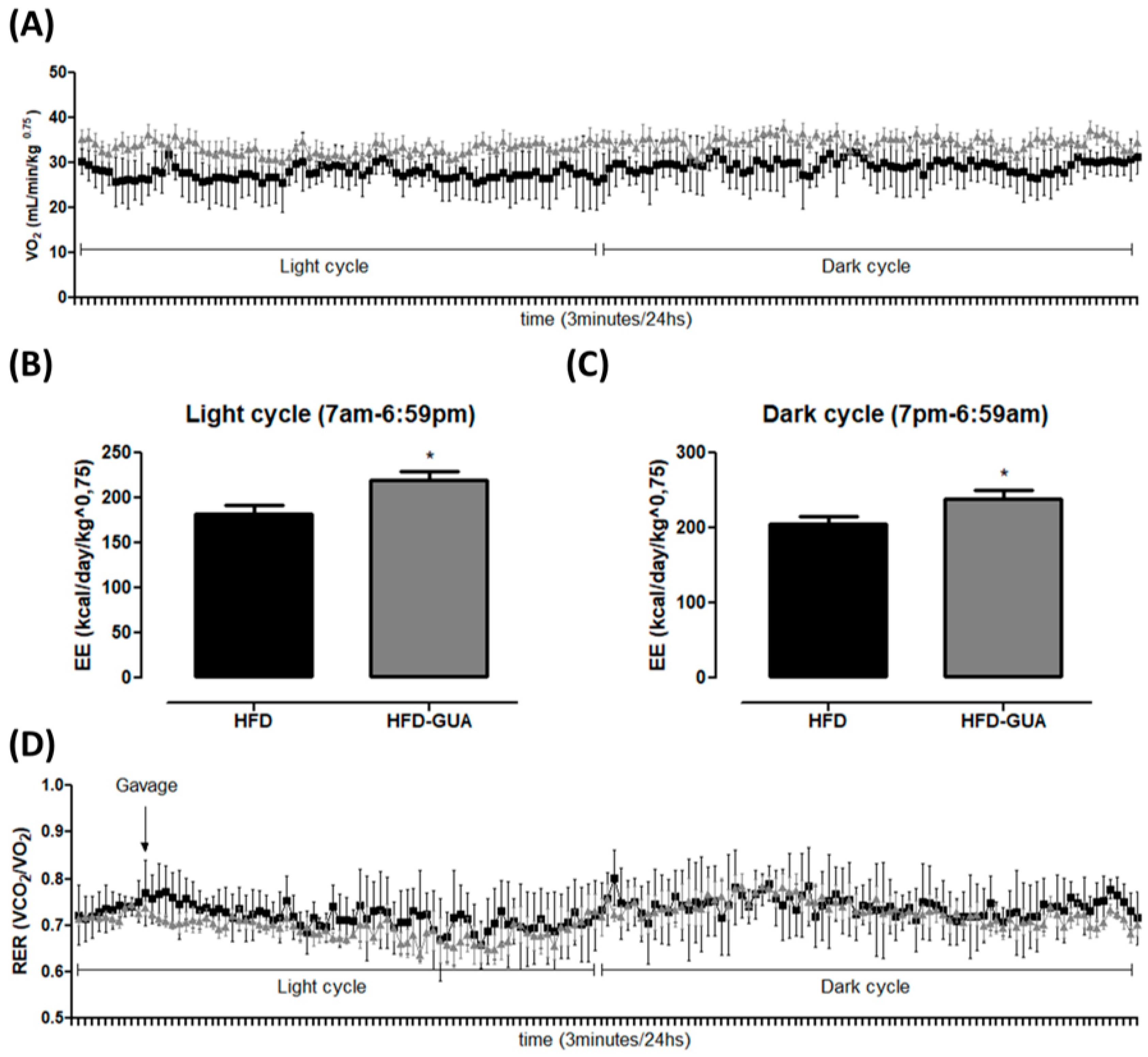
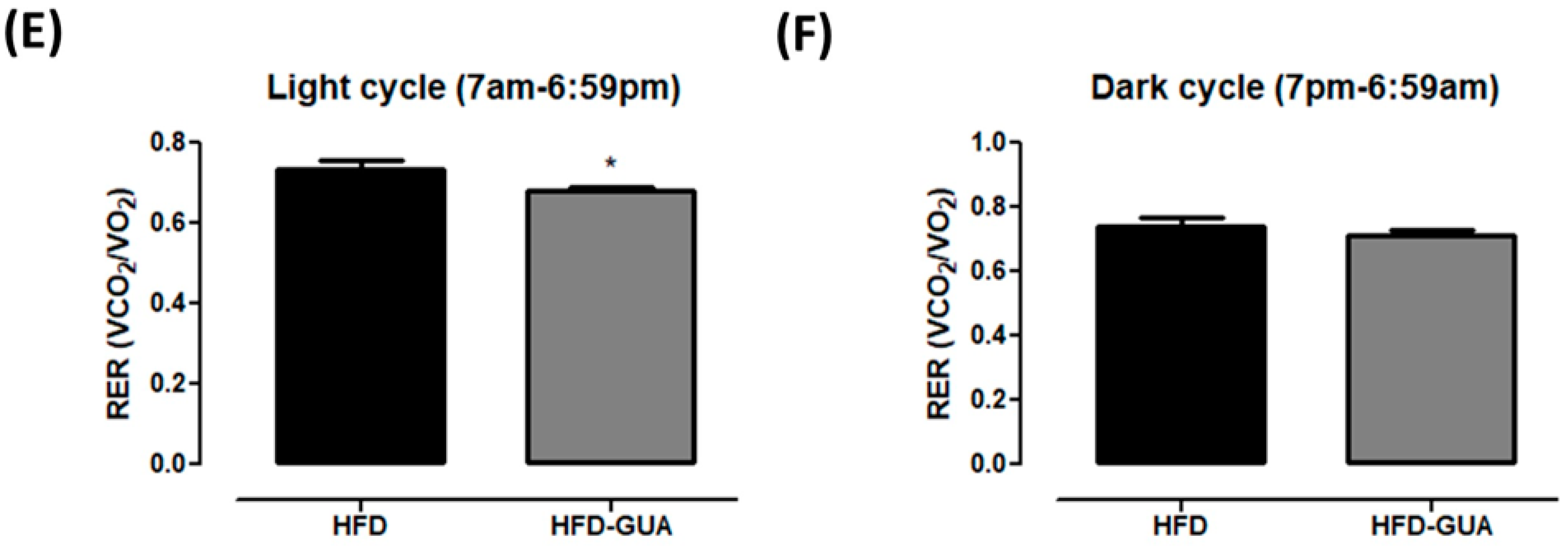
2.2. Indirect Calorimetry
2.3. Animal Procedure and Tissue Dissection
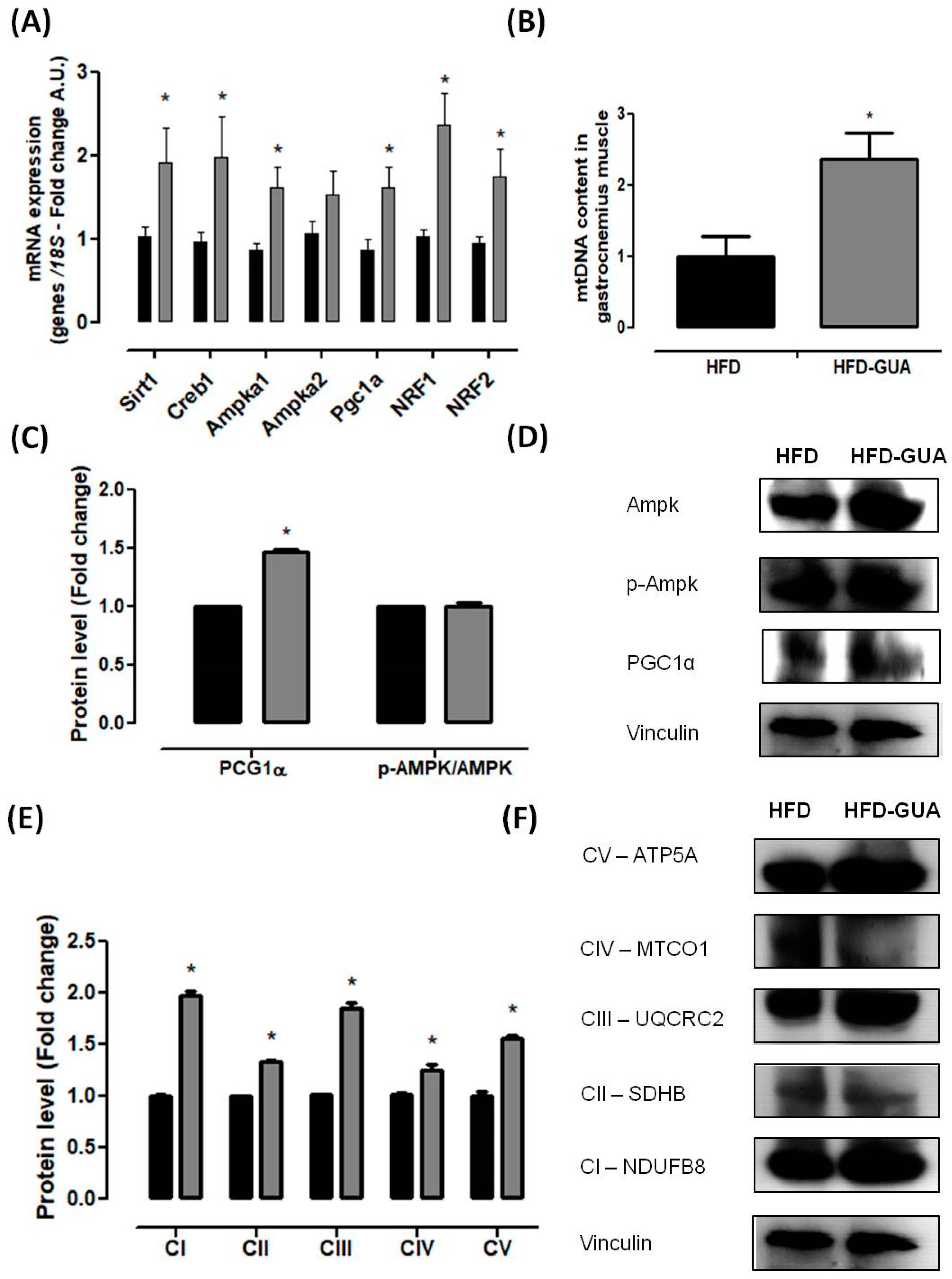
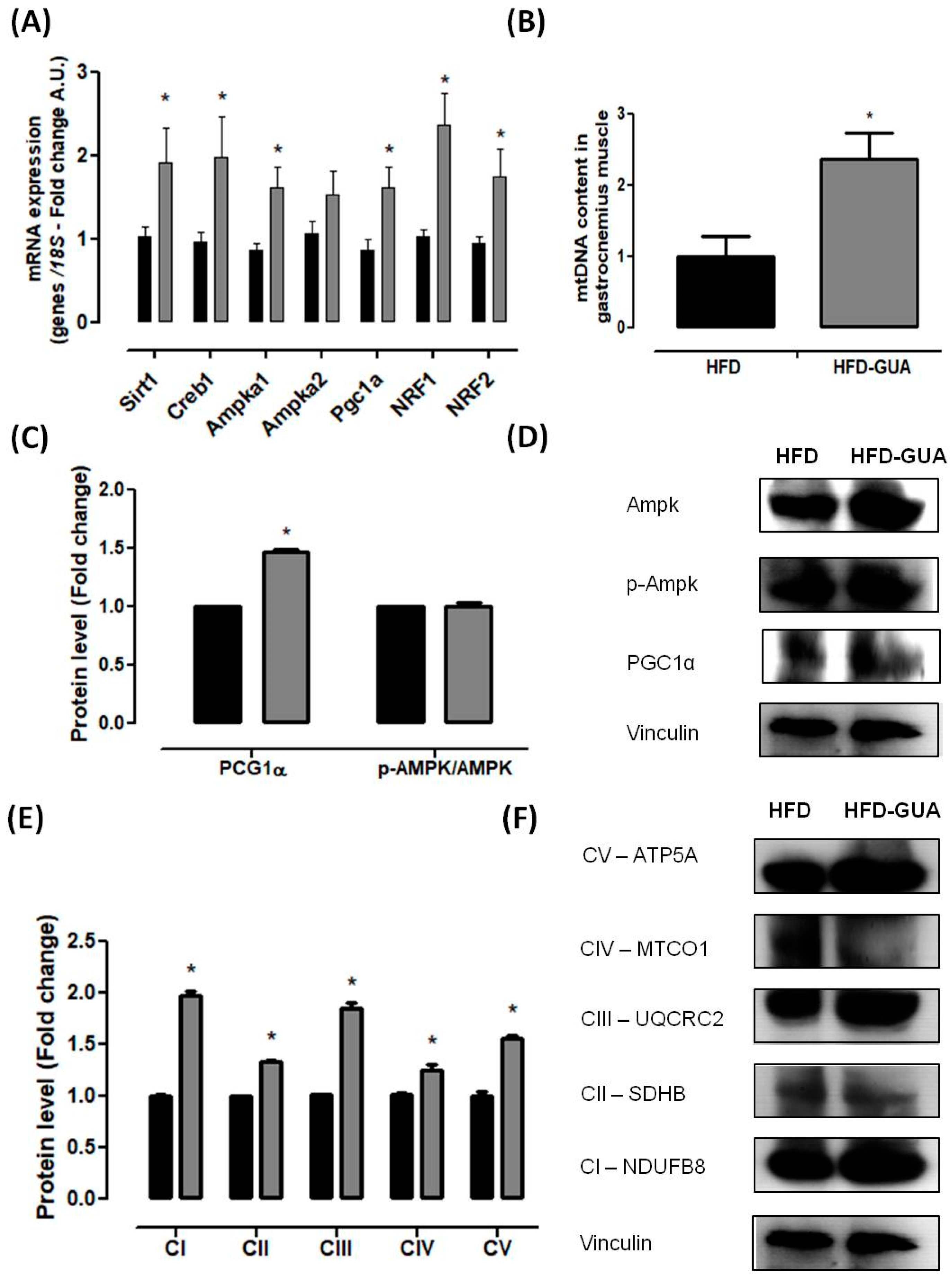
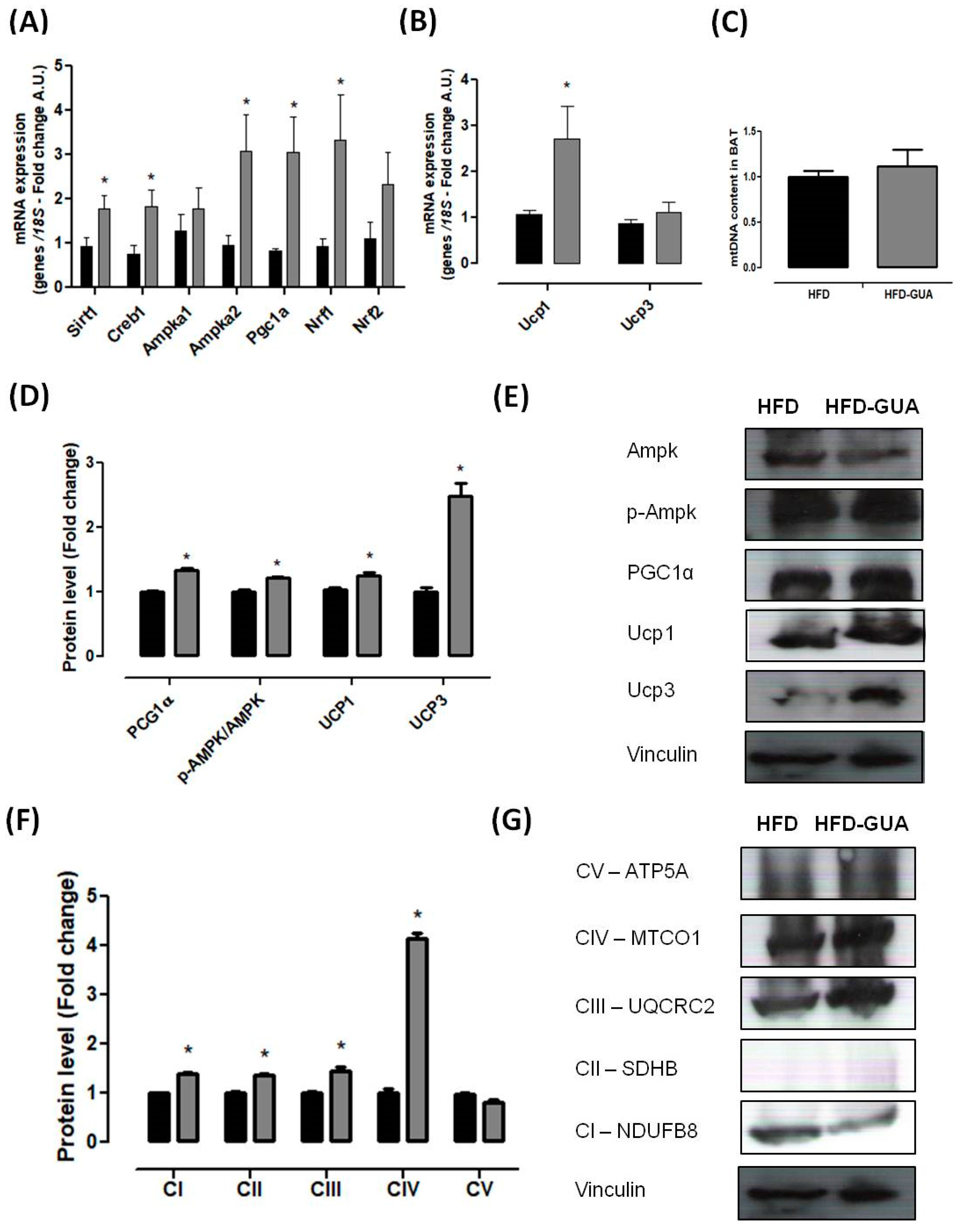
2.4. mRNA (Messenger Robonucleic Acid) Expression Analysis
2.5. Mitochondrial DNA Quantification (Mtdna)
2.6. Western Blotting Analysis
2.7. Statistical Analysis
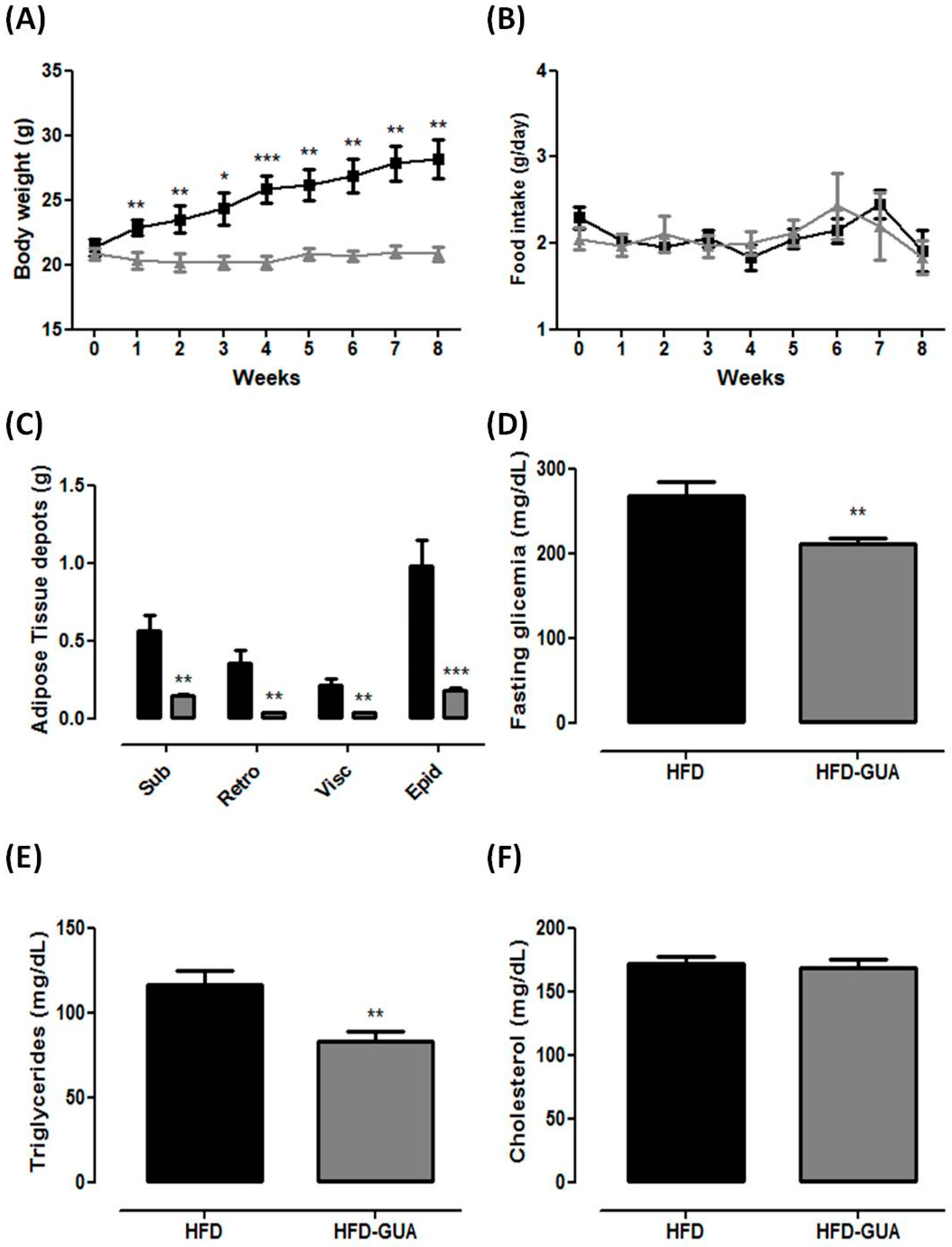
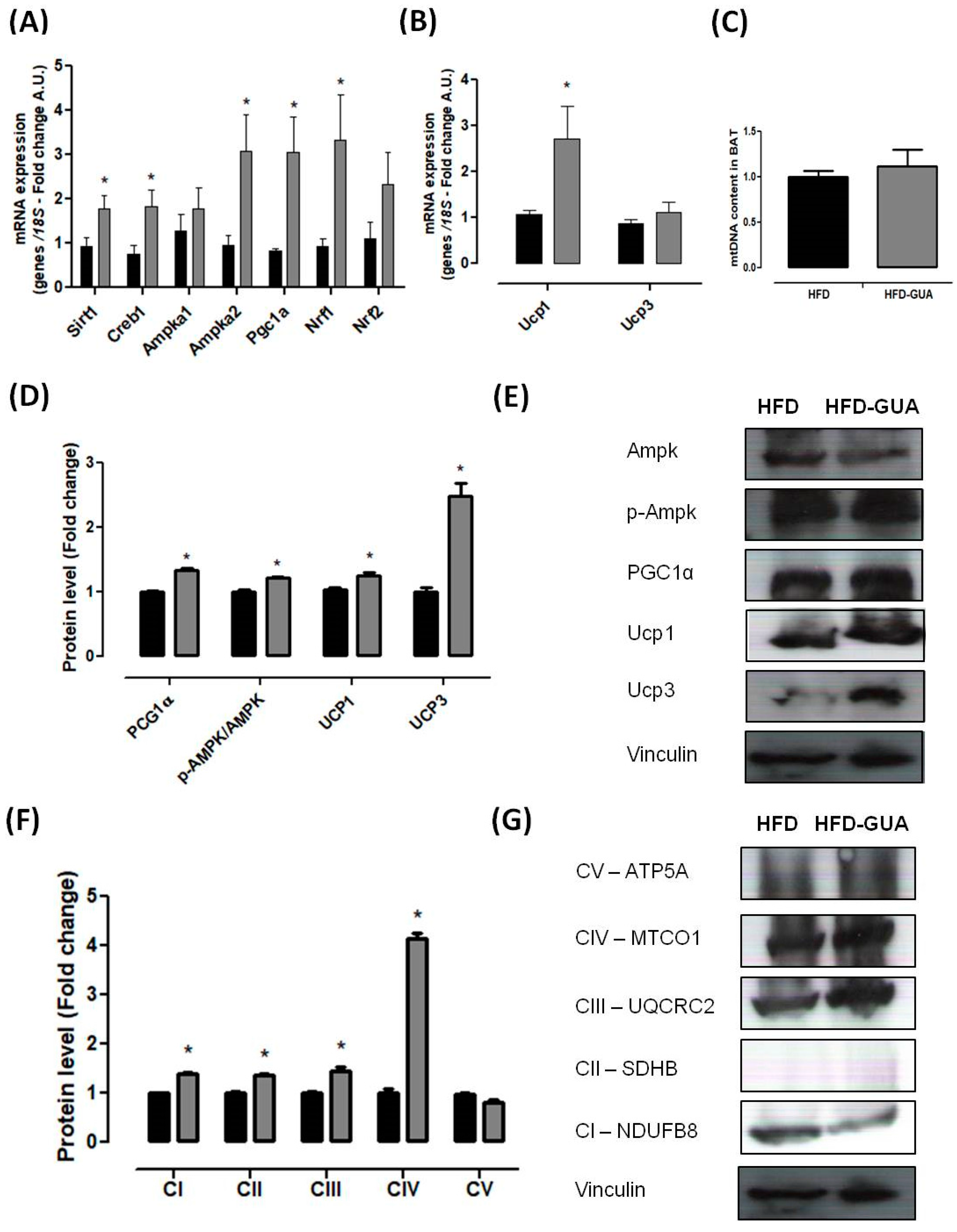
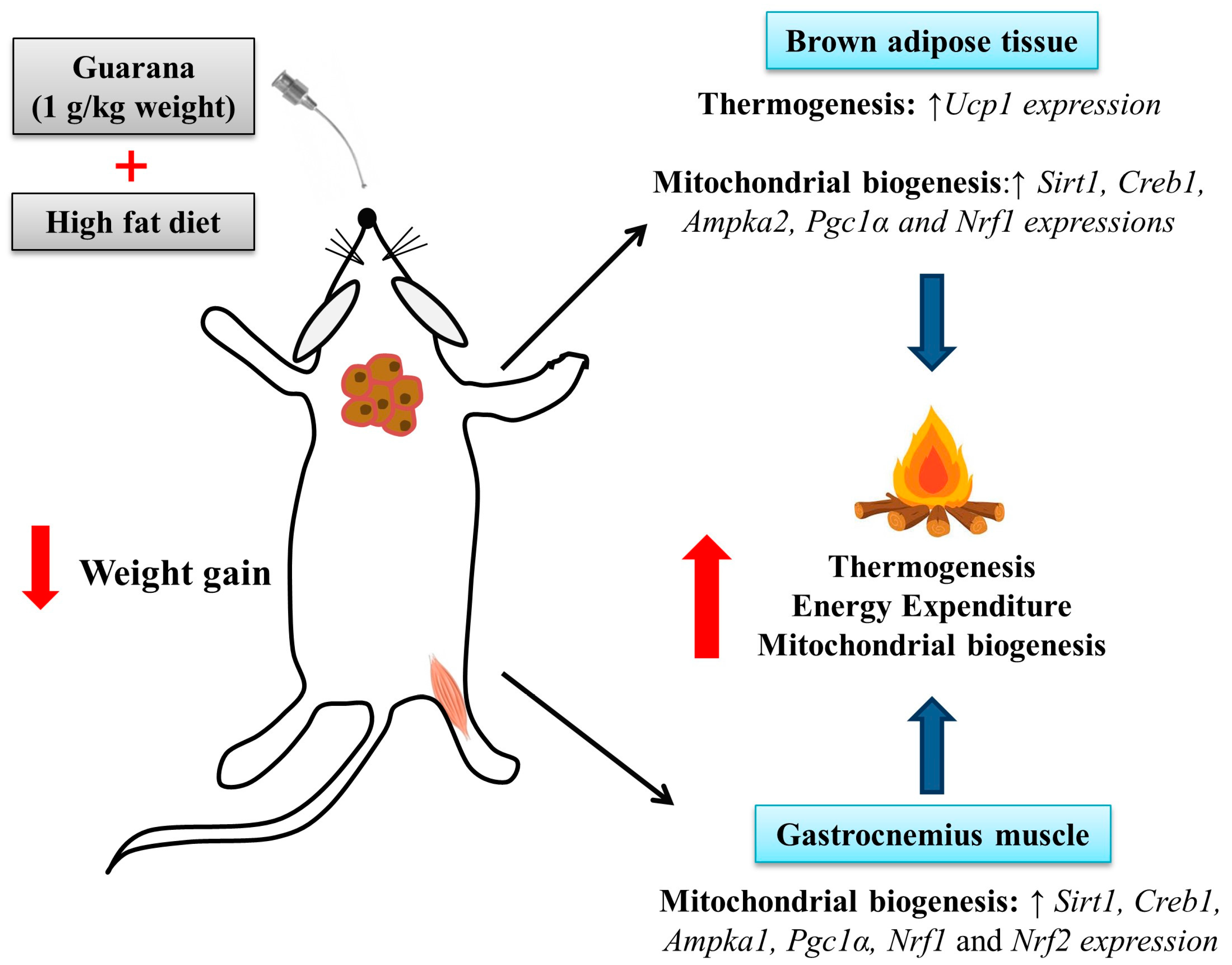
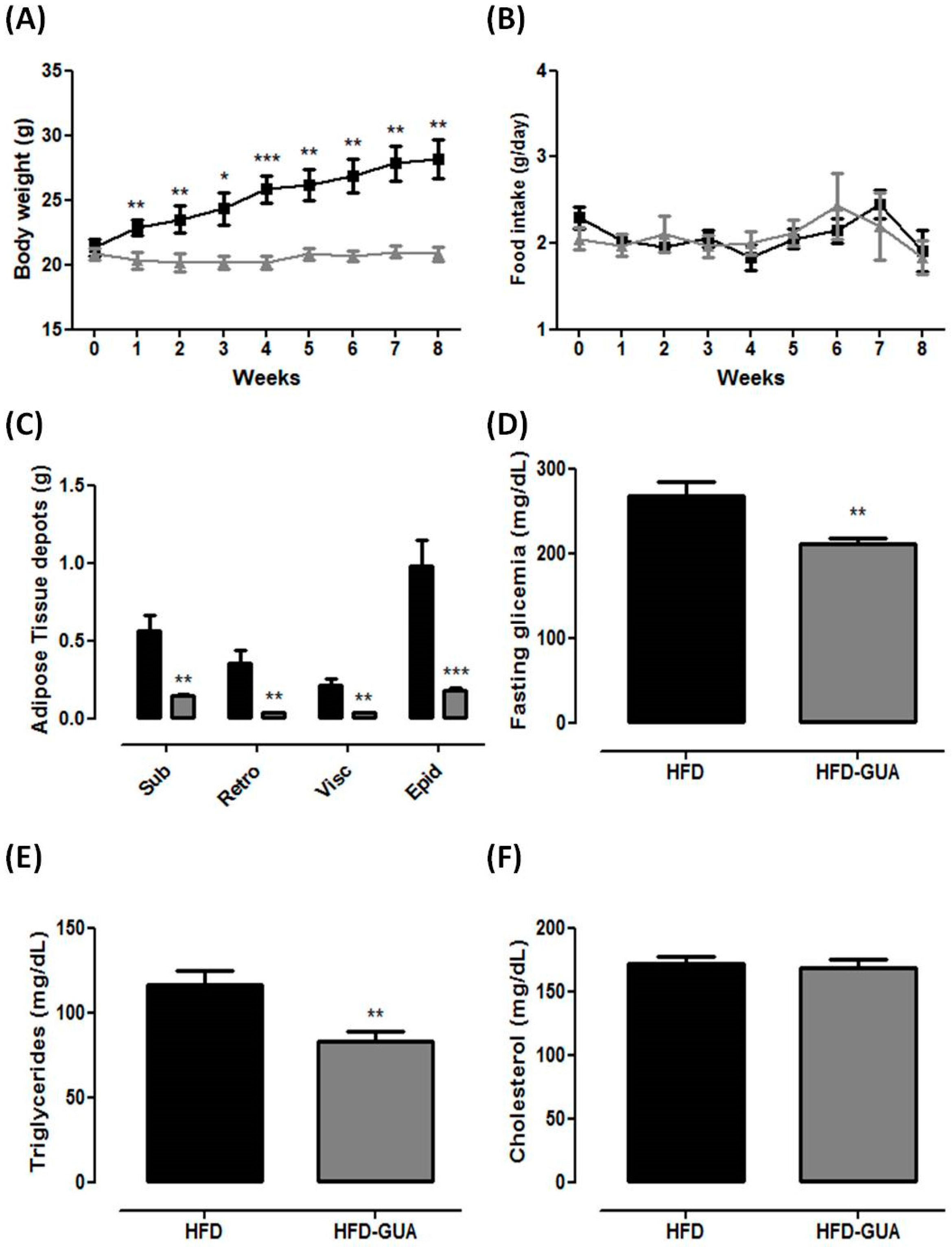
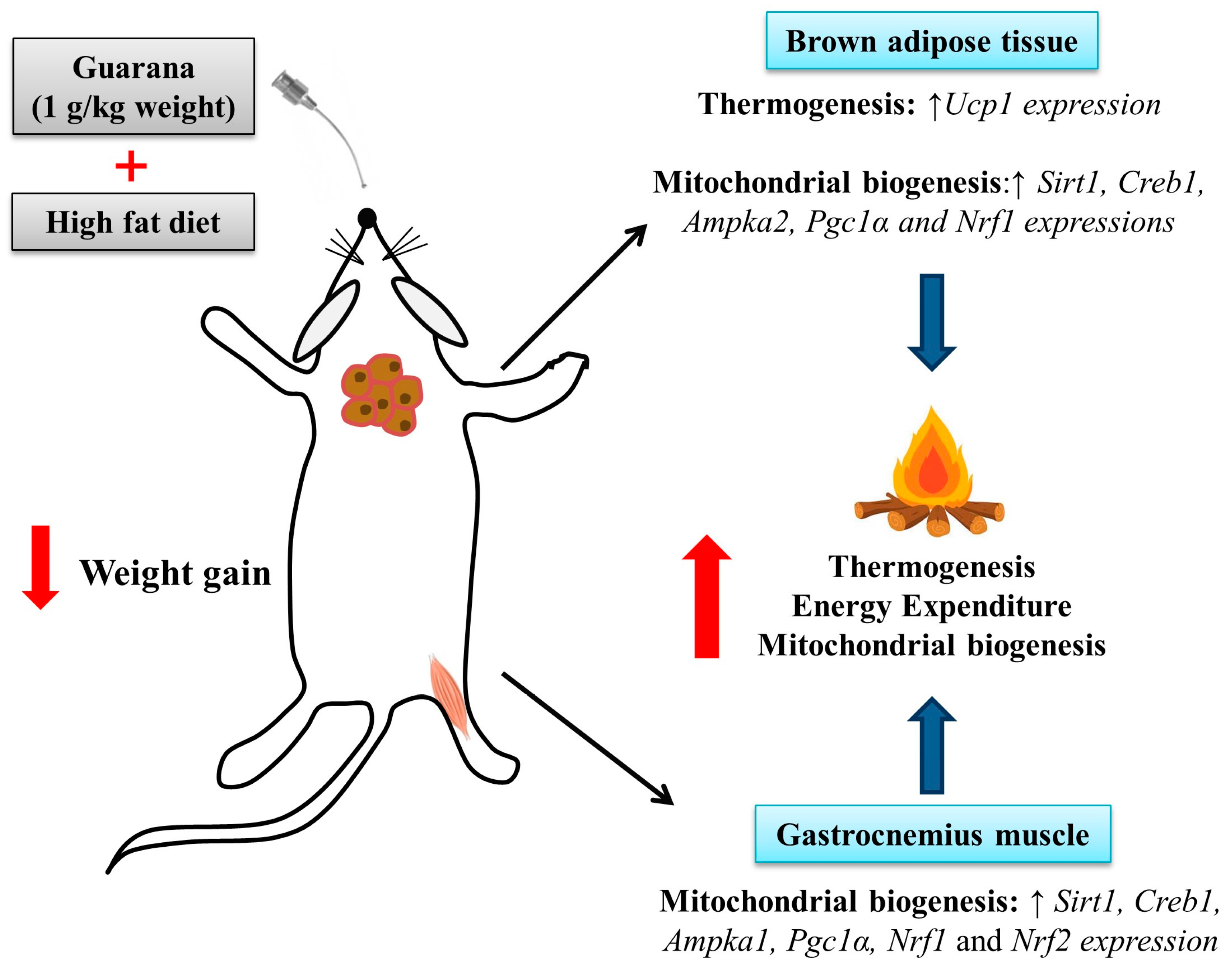
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Pearce, M.; Bray, I.; Horswell, M. Weight gain in mid-childhood and its relationship with the fast food environment. J. Public Health (Oxf.) 2017, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Krssak, M.; Falk Petersen, K.; Dresner, A.; DiPietro, L.; Vogel, S.M.; Rothman, D.L.; Roden, M.; Shulman, G.I. Intramyocellular lipid concentrations are correlated with insulin sensitivity in humans: A 1H-NMR spectroscopy study. Diabetologia 1999, 42, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Greco, A.V.; Mingrone, G.; Giancaterini, A.; Manco, M.; Morroni, M.; Cinti, S.; Granzotto, M.; Vettor, R.; Camastra, S.; Ferrannini, E. Insulin resistance in morbid obesity reversal with intramyocellular fat depletion. Diabetes 2002, 51, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.A. New insights into PGC-1 coactivators: Redefining their role in the regulation of mitochondrial function and beyond. FEBS J. 2015, 282, 647–672. [Google Scholar] [CrossRef] [PubMed]
- Viscomi, C.; Bottani, E.; Civiletto, G.; Cerutti, R.; Moggio, M.; Fagiolari, G.; Schon, E.A.; Lamperti, C.; Zeviani, M. In vivo correction of COX deficiency by activation of the AMPK/PGC-1α axis. Cell Metab. 2011, 14, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Ichinose, T.; Jinde, M.; Kawashima, Y.; Tachiyashiki, K.; Imaizumi, K. Tea catechins enhance the mRNA expression of uncoupling protein 1 in rat brown adipose tissue. J. Nutr. Biochem. 2008, 19, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; Ye, Z.; Xu, C.; Zhang, M.; Ruan, B.; Wei, M.; Jiang, Y.; Zhang, Y.; Wang, L.; et al. Curcumin promotes browning of white adipose tissue in a norepinephrine-dependent way. Biochem. Biophys. Res. Commun. 2015, 466, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Q.X.; Wang, X.; Zhang, L.; Qu, W.; Bao, B.; Liu, C.A.; Liu, J. Dietary luteolin activates browning and thermogenesis in mice through an AMPK/PGC1α pathway-mediated mechanism. Int. J. Obes. (Lond.) 2016, 40, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Krewer Cda, C.; Ribeiro, E.E.; Ribeiro, E.A.; Moresco, R.N.; da Rocha, M.I.; Montagner, G.F.; Machado, M.M.; Viegas, K.; Brito, E.; da Cruz, I.B. Habitual intake of guarana and metabolic morbidities: An epidemiological study of an elderly Amazonian population. Phytother. Res. 2011, 25, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Harrold, J.A.; Hughes, G.M.; O’Shiel, K.; Quinn, E.; Boyland, E.J.; Williams, N.J.; Halford, J.C. Acute effects of a herb extract formulation and inulin fibre on appetite, energy intake and food choice. Appetite 2013, 62, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Lima, N.S.; Numata, E.P.; Mesquita, L.M.S.; Dias, P.H.; Vilegas, W.; Gambero, A.; Ribeiro, M.L. Modulatory effects of guarana (Paullinia cupana) on adipogenesis. Nutrients 2017, 20, 635. [Google Scholar] [CrossRef]
- Bérubé-Parent, S.; Pelletier, C.; Doré, J.; Tremblay, A. Effects of encapsulated green tea and Guarana extracts containing a mixture of epigallocatechin-3-gallate and caffeine on 24 h energy expenditure and fat oxidation in men. Br. J. Nutr. 2005, 94, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Gotardo, E.M.; Piraee, M.; Gambero, A.; Ribeiro, M.L. Effects of yerba maté, a plant extract formulation (“YGD”) and resveratrol in 3T3-L1 adipogenesis. Molecules 2014, 19, 16909–16924. [Google Scholar] [CrossRef] [PubMed]
- Celestino, M.M.; Gomes, A.C.; Botelho, P.B.; Gambero, A.; de Souza Mesquita, L.M.; Vilegas, W.; Ribeiro, M.L.; Mota, J.F. South American herbal extracts reduce food intake through modulation of gastrointestinal hormones in overweight and obese women. J. Funct. Foods 2017. [Google Scholar] [CrossRef]
- Alkhatib, A.; Seijo, M.; Larumbe, E.; Naclerio, F. Acute effectiveness of a “fat-loss” product on substrate utilization, perception of hunger, mood state and rate of perceived exertion at rest and during exercise. J. Int. Soc. Sports Nutr. 2015, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Kalinovich, A.V.; Mattsson, C.L.; Youssef, M.R.; Petrovic, N.; Ost, M.; Skulachev, V.P.; Shabalina, I.G. Mitochondria-targeted dodecyltriphenylphosphonium (C12TPP) combats high-fat-diet-induced obesity in mice. Int. J. Obes. (Lond.) 2016, 40, 1864–1874. [Google Scholar] [CrossRef] [PubMed]
- Moro, C.; Harant, I.; Badin, P.M.; Patarca, F.X.; Guilland, J.C.; Bourlier, V.; Langin, D.; De Glisezinski, I. Influence of lipolysis and fatty acid availability on fuel selection during exercise. J. Physiol. Biochem. 2014, 70, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Cadoná, F.C.; Machado, A.K.; Azzolin, V.F.; Barbisan, F.; Dornelles, E.B.; Glanzner, W.; Gonçalves, P.B.; Assmann, C.E.; Ribeiro, E.E.; da Cruz, I.B. Guaraná a richest caffeine food increase oxaliplatin sensitivity of colorectal HT-29 cells by apoptosis pathway modulation. Anticancer Agents Med. Chem. 2015, 16, 26673759. [Google Scholar]
- Hursel, R.; Viechtbauer, W.; Dulloo, A.G.; Tremblay, A.; Tappy, L.; Rumpler, W.; Westerterp-Plantenga, M.S. The effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: A meta-analysis. Obes. Rev. 2011, 12, e573–e581. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Trujillo, K.A.; Conn, C.A. Effects of caffeine on metabolism and mitochondria biogenesis in rhabdomyosarcoma cells compared with 2,4-dinitrophenol. Nutr. Metab. Insights 2012, 5, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Meng, J.; Shen, Q.; Zhang, Y.; Pan, S.; Chen, Z.; Zhu, L.Q.; Lu, Y.; Huang, Y.; Zhang, G. Caffeine inhibits hypothalamic A1R to excite oxytocin neuron and ameliorate dietary obesity in mice. Nat. Commun. 2017, 27, 15904. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Sayama, K.; Okubo, T.; Juneja, L.R.; Oguni, I. Anti-obesity effects of three major components of green tea, catechins, caffeine and theanine, in mice. In Vivo 2004, 18, 55–62. [Google Scholar] [PubMed]
- Murase, T.; Nagasawa, A.; Suzuki, J.; Hase, T.; Tokimitsu, I. Beneficial effects of tea catechins on dietinduced obesity: Stimulation of lipid catabolism in the liver. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Dangol, M.; Kim, S.; Li, C.G.; Fakhraei Lahiji, S.; Jang, M.; Ma, Y.; Huh, I.; Jung, H. Anti-obesity effect of a novel caffeine-loaded dissolving microneedle patch in high-fat diet-induced obese C57BL/6J mice. J. Control Release 2017, 265, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.I.; Zito, G.; Colquhoun, R.; Martinez, N.; Kendall, K.; Buchanan, L.; Lehn, M.; Johnson, M.; St Louis, C.; Smith, Y.; et al. The effects of a single-dose thermogenic supplement on resting metabolic rate and hemodynamic variables in healthy females—A randomized, double-blind, placebo-controlled, cross-over trial. J. Int. Soc. Sports Nutr. 2016, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Cedikova, M.; Kripnerová, M.; Dvorakova, J.; Pitule, P.; Grundmanova, M.; Babuska, V.; Mullerova, D.; Kuncova, J. Mitochondria in White, Brown, and Beige Adipocytes. Stem Cells Int. 2016, 2016, 6067349. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, J.; Ma, Y.; Wu, G.; Tian, Y.; Shi, Y.; Le, G. Resveratrol preserves mitochondrial function, stimulates mitochondrial biogenesis, and attenuates oxidative stress in regulatory T cells of mice fed a high-fat diet. J. Food Sci. 2014, 79, H1823–H1831. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.C.; Geiger, P.C.; Han, D.H.; Jones, T.E.; Holloszy, J.O. Calcium induces increases in peroxisome proliferator-activated receptor gamma coactivator-1alpha and mitochondrial biogenesis by a pathway leading to p38 mitogen-activated protein kinase activation. J. Biol. Chem. 2007, 282, 18793–18799. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | High-Fat Diet | |
|---|---|---|
| g kg−1 | kcal kg−1 | |
| Cornstarch | 115.5 | 462 |
| Casein | 200 | 800 |
| Sucrose | 100 | 400 |
| Dextrinated starch | 132 | 528 |
| Lard | 312 | 2808 |
| Soybean oil | 40 | 360 |
| Cellulose | 50 | - |
| Mineral mix | 35 | - |
| Vitamin mix | 10 | - |
| l-cystine | 3 | - |
| Choline | 2.5 | - |
| TOTAL | 1000 | 5358 |
| Gene | Primer | Sequence (5′→3′) |
|---|---|---|
| Sirt1 | Sense | AGTGGCACATGCCAGAGTC |
| Antisense | TCCAGATCCTCCAGCACAAT | |
| Creb1 | Sense | TTTGTCCTTGCTTTCCGAAT |
| Antisense | CACTTTGGCTGGACATCTTG | |
| Ampka1 | Sense | TGAGAACGTCCTGCTTGAATG |
| Antisense | ATCATTGGCTGAGCCACAGC | |
| Ampka2 | Sense | ACAGGCCATAAAGTGGCAGT |
| Antisense | GTCGGAGTGCTGATCACGTG | |
| Pgc1α | Sense | CCGAGAATTCATGGAGCAAT |
| Antisense | TTTCTGTGGGTTTGGTGTGA | |
| Nrf1 | Sense | CAACAGGGAAGAAACGGAAA |
| Antisense | CACTCGCGTCGTGTACTCAT | |
| Nrf2 | Sense | AGGACATGGAGCAAGTTTGG |
| Antisense | TCTGTCAGTGTGGCTTCTGG | |
| Ucp1 | Sense | TCAGGGCTGAGTCCTTTTGT |
| Antisense | CTGAAACTCCGGCTGAGAAG | |
| Ucp3 | Sense | CTCACTTTTCCCCTGGACAC |
| Antisense | GTCAGGATGGTACCCAGCAC | |
| 18S | Sense | AAACGGCTACCACATCCAAG |
| Antisense | CAATTACAGGGCCTCGAAAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, N.D.S.; Teixeira, L.; Gambero, A.; Ribeiro, M.L. Guarana (Paullinia cupana) Stimulates Mitochondrial Biogenesis in Mice Fed High-Fat Diet. Nutrients 2018, 10, 165. https://doi.org/10.3390/nu10020165
Lima NDS, Teixeira L, Gambero A, Ribeiro ML. Guarana (Paullinia cupana) Stimulates Mitochondrial Biogenesis in Mice Fed High-Fat Diet. Nutrients. 2018; 10(2):165. https://doi.org/10.3390/nu10020165
Chicago/Turabian StyleLima, Natália Da Silva, Lucimara Teixeira, Alessandra Gambero, and Marcelo Lima Ribeiro. 2018. "Guarana (Paullinia cupana) Stimulates Mitochondrial Biogenesis in Mice Fed High-Fat Diet" Nutrients 10, no. 2: 165. https://doi.org/10.3390/nu10020165
APA StyleLima, N. D. S., Teixeira, L., Gambero, A., & Ribeiro, M. L. (2018). Guarana (Paullinia cupana) Stimulates Mitochondrial Biogenesis in Mice Fed High-Fat Diet. Nutrients, 10(2), 165. https://doi.org/10.3390/nu10020165