Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents

 ,
,  , , ,
, , , 
Abstract
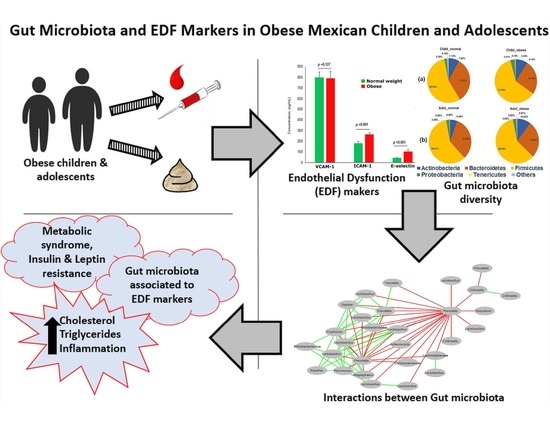
1. Introduction
2. Materials and Methods
2.1. Selection of Study Subjects
2.2. Anthropometrical Evaluation
2.3. Dietary Profile
2.4. Biochemical Test
2.5. Measurement of EDF Markers
2.6. Collection of Fecal Samples
2.7. DNA Extraction and High Throughput Sequencing
2.8. Measurement of SCFAs
2.9. Gut Microbial Diversity
2.10. Gut Microbial Abundance
2.11. Multivariate Analysis
2.12. Co-Occurrence Analysis
2.13. Statistical Analysis
3. Results
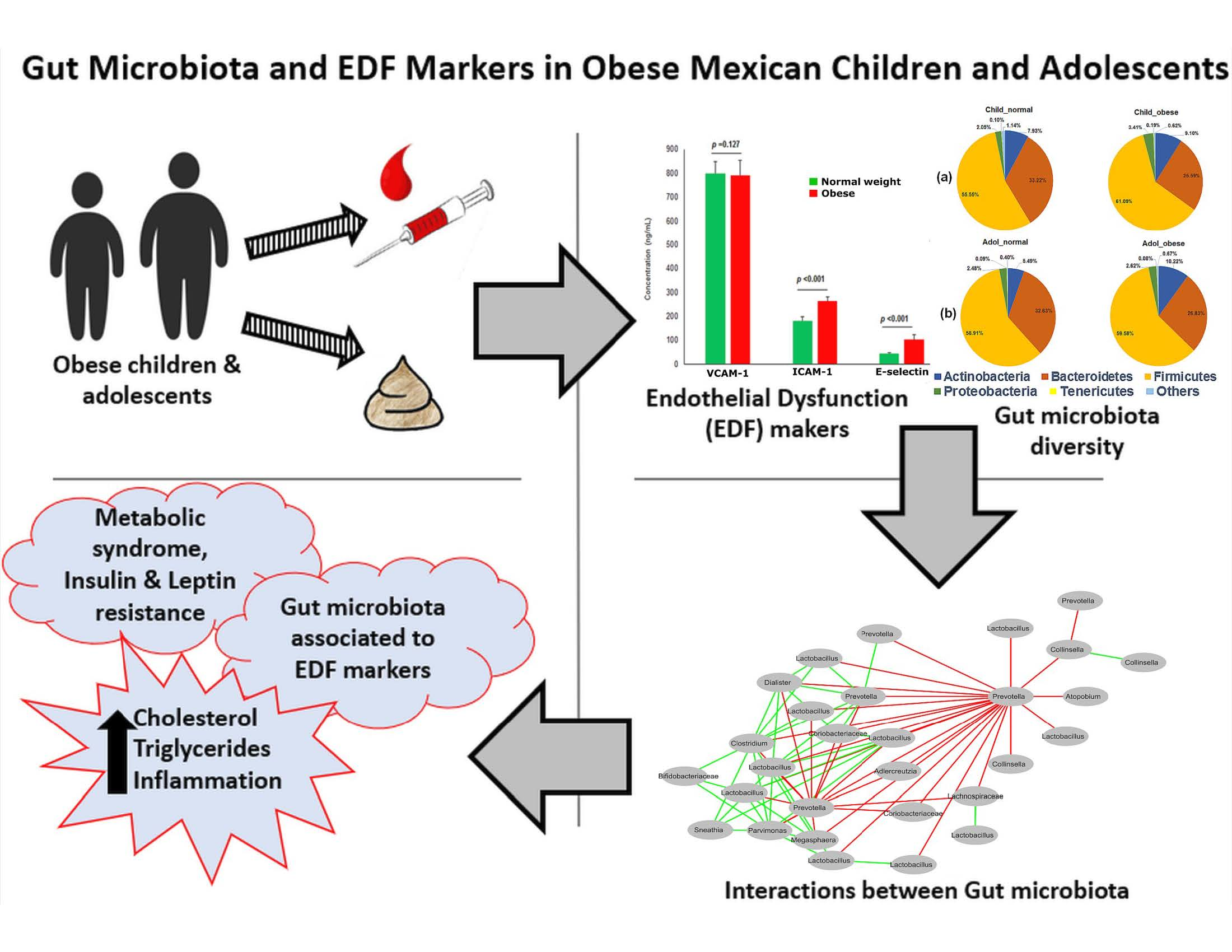
3.1. Obese Children and Adolescents Have Dyslipidemia, Insulin Resistance, and Metabolic Syndrome
3.2. Obese Children and Adolescents Have a Trend in Higher Abundance of Firmicutes and Lower Abundance of Bacteroidetes
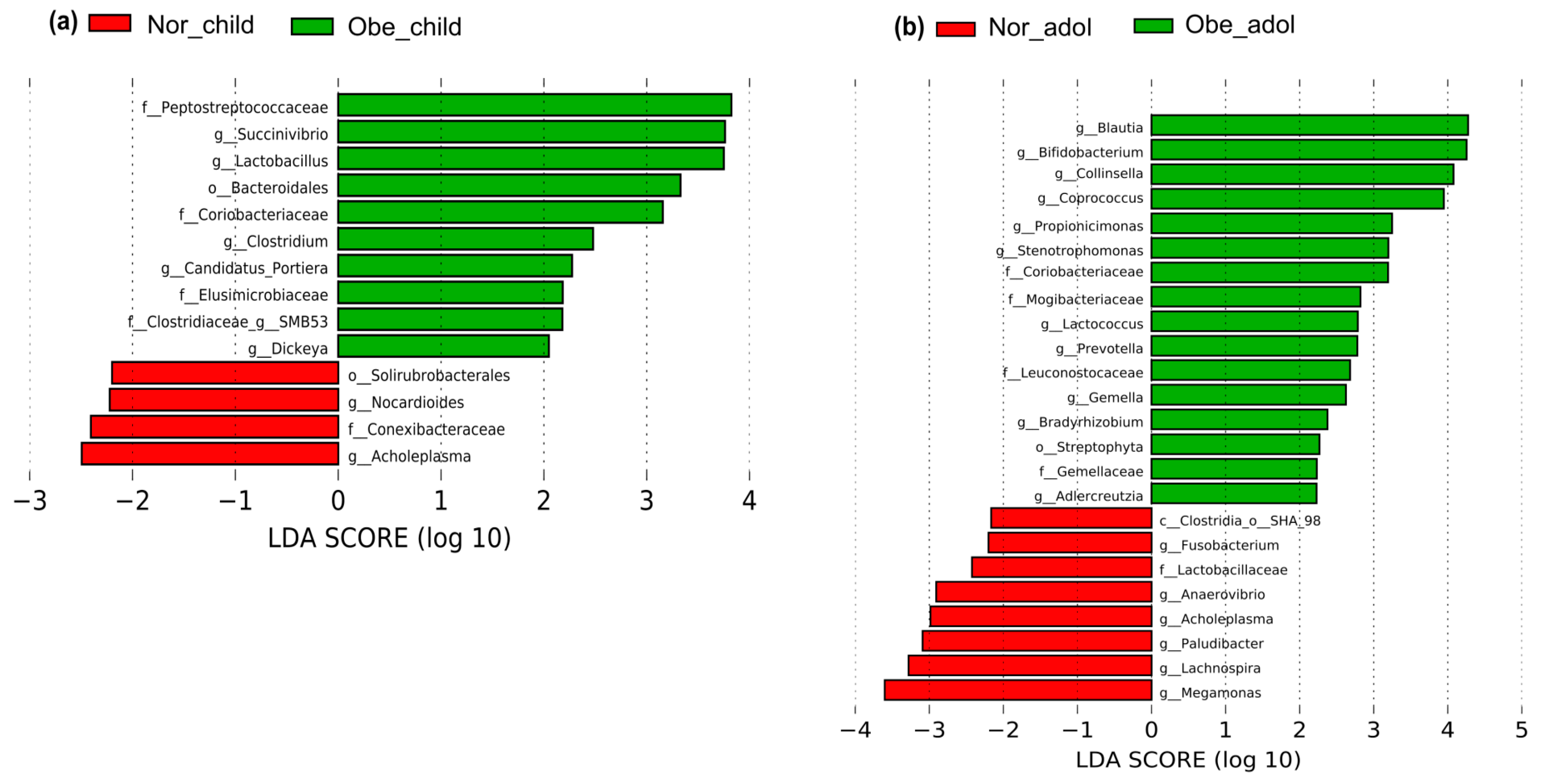
3.3. Differences in Abundance of Gut Bacteria between Normal Weight and Obese Children and Adolescents
3.4. Significant Association between Gut Microbiota Members and EDF and Dyslipidemia Markers in Obesity
3.5. Interactions between Gut Microbiota in Obese Children and Adolescents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Obesity and Overweight. Available online: http://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 25 September 2018).
- Obesity Update 2017. Available online: https://www.oecd.org/els/health-systems/Obesity-Update-2017.pdf (accessed on 25 September 2018).
- Narro, J.; Secretario, R.; Salud, D.E. Directorio Secretaría De Salud. Available online: https://www.gob.mx/cms/uploads/attachment/file/209093/ENSANUT.pdf (accessed on 5 October 2018).
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, S.; Ulloa-Martínez, M.; Martínez-Rojano, H.; Galván-Rodríguez, F.M.; Miranda-Brito, C.; Romano, M.C.; Piña-Escobedo, A.; Pizano-Zárate, M.L.; Hoyo-Vadillo, C.; García-Mena, J. Study of the diversity and short-chain fatty acids production by the bacterial community in overweight and obese Mexican children. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Murugesan, S.; Nirmalkar, K.; Hoyo-Vadillo, C.; García-Espitia, M.; Ramírez-Sánchez, D.; García-Mena, J. Gut microbiome production of short-chain fatty acids and obesity in children. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Hadi, H.A.R.; Carr, C.S.; Al Suwaidi, J. Endothelial dysfunction: Cardiovascular risk factors, therapy, and outcome. Vasc. Health Risk Manag. 2005, 1, 183–198. [Google Scholar]
- Gomes, F.; Telo, D.F.; Souza, H.P.; Nicolau, J.C.; Halpern, A.; Serrano, C.V. Obesity and coronary artery disease: Role of vascular inflammation. Arq. Bras. Cardiol. 2010, 94, 273–279. [Google Scholar] [CrossRef]
- Ludmer, P.L.; Selwyn, A.P.; Shook, T.L.; Wayne, R.R.; Mudge, G.H.; Alexander, R.W.; Ganz, P. Paradoxical vasoconstriction induced by acetylcholine in atherosclerotic coronary arteries. N. Engl. J. Med. 1986, 315, 1046–1051. [Google Scholar] [CrossRef]
- Korkmaz, H.; Onalan, O. Evaluation of endothelial dysfunction: Flow-mediated dilation. Endothelium 2008, 15, 157–163. [Google Scholar] [CrossRef]
- Lopez-Garcia, E.; Schulze, M.B.; Fung, T.T.; Meigs, J.B.; Rifai, N.; Manson, J.E.; Hu, F.B. Major dietary patterns are related to plasma concentrations of markers of inflammation and endothelial dysfunction. Am. J. Clin. Nutr. 2004, 80, 1029–1035. [Google Scholar] [CrossRef]
- Eikemo, H.; Sellevold, O.F.M.; Videm, V. Markers for endothelial activation during open heart surgery. Ann. Thorac. Surg. 2004, 77, 214–219. [Google Scholar] [CrossRef]
- Madrigal, J.C.; Correa, S.M.L.; Gómez, V.G.; Ramírez, T.L.; Gamiño, G.B.; Pérez, Y.C. Valores de función endotelial en niños mexicanos. Med. Interna Mex. 2011, 27, 429–438. [Google Scholar]
- Davignon, J.; Ganz, P. Role of endothelial dysfunction in atherosclerosis. Circulation 2004, 109, III-27–III-32. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Saldaña, J.; Rodriguez-Flores, M.; Cantú-Brito, C.; Aguirre-Garcia, J. A pathological study of the epidemiology of atherosclerosis in Mexico city. Cardiol. Res. Pract. 2014, 2014, 264205. [Google Scholar] [CrossRef] [PubMed]
- Meyers, M.R.; Gokce, N. Endothelial dysfunction in obesity: Etiological role in atherosclerosis. Curr. Opin. Endocrinol. Diabetes Obes. 2007, 14, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Schächinger, V.; Zeiher, A.M. Atherosclerosis-associated endothelial dysfunction. Ärztez. Kardiol. 2000, 89, IX70–IX74. [Google Scholar] [CrossRef]
- Koren, O.; Spor, A.; Felin, J.; Fak, F.; Stombaugh, J.; Tremaroli, V.; Behre, C.J.; Knight, R.; Fagerberg, B.; Ley, R.E.; et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc. Natl. Acad. Sci. USA 2011, 108, 4592–4598. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Fåk, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Bäckhed, F.; Nielsen, J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef]
- Du, H.; Feskens, E. Dietary determinants of obesity. Acta Cardiol. 2010, 65, 377–386. [Google Scholar] [CrossRef]
- Freedman, D.S.; Mei, Z.; Srinivasan, S.R.; Berenson, G.S.; Dietz, W.H. Cardiovascular risk factors and excess adiposity among overweight children and adolescents: The Bogalusa heart study. J. Pediatr. 2007, 150, 12–17. [Google Scholar] [CrossRef]
- Nirmalkar, K.; Murugesan, S.; Pizano-Zárate, M.L.; Romero-Figueroa, S.; Hoyo-Vadillo, C.; Garcia-Mena, J. Endothelial dysfunction in Mexican obese children, is there a role of the gut microbiota ? Obes. Control Ther. 2017. [Google Scholar] [CrossRef]
- de Onis, M.; Onyango, A.W.; Borghi, E.; Siyam, A.; Nishida, C.; Siekmann, J. Development of a WHO growth reference for school-aged children and adolescents. Bull. World Health Organ. 2007, 85, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimationof the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Perichart-Perera, O.; Muñoz-Manrique, C.; Reyes-López, A.; Tolentino-Dolores, M.; Espino, Y.; Sosa, S.; Ramírez-González, M.C. Metabolic markers during pregnancy and their association with maternal and newborn weight status. PLoS ONE 2017, 12, e0180874. [Google Scholar] [CrossRef]
- Aradillas-García, C.; Rodríguez-Morán, M.; Garay-Sevilla, M.E.; Malacara, J.M.; Rascon-Pacheco, R.A.; Guerrero-Romero, F. Distribution of the homeostasis model assessment of insulin resistance in Mexican children and adolescents. Eur. J. Endocrinol. 2012, 166, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Burguete García, A.I.; Váldes Villalpando, Y.N.; Cruz, M. Definiciones para el diagnóstico de síndrome metabólico en población infantil. Gac. Médica México 2014, 150, 79–87. [Google Scholar]
- American Academy of Pediatrics. National Cholesterol Education Program: Report of the Expert Panel on Blood Cholesterol Levels in Children and Adolescents; NIH Publication: Bethesda, MD, USA, 1992; p. 2732. [Google Scholar]
- Weiss, R.; Dziura, J.; Burgert, T.S.; Tamborlane, W.V.; Taksali, S.E.; Yeckel, C.W.; Allen, K.; Lopes, M.; Savoye, M.; Morrison, J.; et al. Obesity and the Metabolic Syndrome in Children and Adolescents. N. Engl. J. Med. 2004, 350, 2362–2374. [Google Scholar] [CrossRef]
- Magge, S.N.; Goodman, E.; Armstrong, S.C. The metabolic syndrome in children and adolescents: Shifting the focus to cardiometabolic risk factor clustering. Pediatrics 2017, 140, e20171603. [Google Scholar] [CrossRef]
- De Baere, S.; Eeckhaut, V.; Steppe, M.; De Maesschalck, C.; De Backer, P.; Van Immerseel, F.; Croubels, S. Development of a HPLC–UV method for the quantitative determination of four short-chain fatty acids and lactic acid produced by intestinal bacteria during in vitro fermentation. J. Pharm. Biomed. Anal. 2013, 80, 107–115. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple. J. R. Statist. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Faust, K.; Raes, J. CoNet app: Inference of biological association networks using Cytoscape. F1000Res. 2016, 5, 1519. [Google Scholar] [CrossRef] [PubMed]
- Rojas, E.; Rodríguez-Molina, D.; Bolli, P.; Israili, Z.H.; Faría, J.; Fidilio, E.; Bermúdez, V.; Velasco, M. The role of adiponectin in endothelial dysfunction and hypertension. Curr. Hypertens. Rep. 2014, 16, 463. [Google Scholar] [CrossRef]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta 2013, 417, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Dron, J.S.; Hegele, R.A. Genetics of triglycerides and the risk of atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 31. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef]
- Clavel, T.; Desmarchelier, C.; Haller, D.; Gérard, P.; Rohn, S.; Lepage, P.; Daniel, H. Intestinal microbiota in metabolic diseases. Gut Microbes 2014, 5, 544–551. [Google Scholar] [CrossRef]
- Wurdemann, D.; Tindall, B.J.; Pukall, R.; Lunsdorf, H.; Strompl, C.; Namuth, T.; Nahrstedt, H.; Wos-Oxley, M.; Ott, S.; Schreiber, S.; et al. Gordonibacter pamelaeae gen. nov., sp. nov., a new member of the Coriobacteriaceae isolated from a patient with Crohn’s disease, and reclassification of Eggerthella hongkongensis Lau et al. 2006 as Paraeggerthella hongkongensis gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1405–1415. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- de La Serre, C.B.; Ellis, C.L.; Lee, J.; Hartman, A.L.; Rutledge, J.C.; Raybould, H.E. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am. J. Physiol. Liver Physiol. 2010, 299, G440–G448. [Google Scholar] [CrossRef] [PubMed]
- Zitomersky, N.L.; Atkinson, B.J.; Franklin, S.W.; Mitchell, P.D.; Snapper, S.B.; Comstock, L.E.; Bousvaros, A. Characterization of adherent bacteroidales from intestinal biopsies of children and young adults with inflammatory bowel disease. PLoS ONE 2013, 8, e63686. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Angelakis, E.; Paul, M.; Armougom, F.; Leibovici, L.; Raoult, D. Comparative meta-analysis of the effect of Lactobacillus species on weight gain in humans and animals. Microb. Pathog. 2012, 53, 100–108. [Google Scholar] [CrossRef]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed]
- Kant, R.; Blom, J.; Palva, A.; Siezen, R.J.; de Vos, W.M. Comparative genomics of Lactobacillus. Microb. Biotechnol. 2011, 4, 323–332. [Google Scholar] [CrossRef]
- Khalyfa, A.; Kheirandish-Gozal, L.; Bhattacharjee, R.; Khalyfa, A.A.; Gozal, D. Circulating microRNAs as potential biomarkers of endothelial dysfunction in obese children. Chest 2016, 149, 786–800. [Google Scholar] [CrossRef]
- Karolina, D.S. MicroRNAs and endothelial dysfunction in relation to obesity and type 2 diabetes. J. Mol. Genet. Med. 2014, S1. [Google Scholar] [CrossRef]
- Vikram, A.; Kim, Y.R.; Kumar, S.; Li, Q.; Kassan, M.; Jacobs, J.S.; Irani, K. Vascular microRNA-204 is remotely governed by the microbiome and impairs endothelium-dependent vasorelaxation by downregulating Sirtuin1. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Widlansky, M.E.; Gokce, N.; Keaney, J.F.; Vita, J.A. The clinical implications of endothelial dysfunction. J. Am. Coll. Cardiol. 2003, 42, 1149–1160. [Google Scholar] [CrossRef]
- Catry, E.; Bindels, L.B.; Tailleux, A.; Lestavel, S.; Neyrinck, A.M.; Goossens, J.F.; Lobysheva, I.; Plovier, H.; Essaghir, A.; Demoulin, J.B.; et al. Targeting the gut microbiota with inulin-type fructans: Preclinical demonstration of a novel approach in the management of endothelial dysfunction. Gut 2018, 67, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Flammer, A.J.; Anderson, T.; Celermajer, D.S.; Creager, M.A.; Deanfield, J.; Ganz, P.; Hamburg, N.M.; Lüscher, T.F.; Shechter, M.; Taddei, S.; et al. The assessment of endothelial function. Circulation 2012, 126, 753–767. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Normal Weight | Obesity | p-Value |
|---|---|---|---|
| Number (F/M) | 49 (30/19) | 62 (27/35) | Nd |
| Age (years) | 9.14 ± 0.22 | 9.50 ± 0.18 | 0.146 µ |
| Age Range (years) | 7–11 | 6–11 | Nd |
| Anthropometric | |||
| Weight (kg) | 31.49 ± 1.02 | 48.54 ± 1.37 | <0.001 µ |
| Height (m) | 1.35 ± 0.01 | 1.39 ± 0.01 | 0.094 Ω |
| BMI (kg/m2) | 16.89 ± 0.21 | 24.90 ± 0.42 | <0.001 µ |
| BMI pc | 57.44 ± 2.71 | 98.78 ± 0.18 | <0.001 µ |
| BMI pc Scale | <85 | >95 | Nd |
| WC (cm) | 59.53 ± 0.68 | 80.55 ± 1.29 | <0.001 µ |
| WC pc | 59.46 ± 3.78 | 97.47 ± 0.72 | <0.001 µ |
| Blood Pressure | |||
| SBP (mm Hg) | 88.42 ± 3.64 | 100.61 ± 1.78 | 0.010 µ |
| SBP pc | 33.20 ± 3.69 | 38.96 ± 3.53 | 0.312 µ |
| DBP (mm Hg) | 59.44 ± 2.66 | 65.56 ± 1.42 | 0.048 µ |
| DBP pc | 56.56 ± 3.87 | 64.54 ± 2.86 | 0.123 µ |
| Metabolic Factors | |||
| Fasting Glucose (mg/dL) | 89.04 ± 0.99 | 91.32 ± 1.33 | 0.223 µ |
| Triglycerides (mg/dL) | 84.64 ± 5.30 | 119.73 ± 7.74 | <0.001 µ |
| Total Cholesterol (mg/dL) | 162.88 ± 3.45 | 166.62 ± 2.70 | 0.651 µ |
| HDL (mg/dL) | 53.25 ± 1.57 | 46.34 ± 2.16 | <0.001 µ |
| LDL (mg/dL) | 92.70 ± 2.82 | 96.34 ± 2.67 | 0.394 µ |
| Insulin (µIU/mL) | 6.49 ± 0.61 ∞ | 13.53 ± 1.42 | <0.001 µ |
| HOMA-IR | 1.44 ± 0.14 | 3.12 ± 0.33 | <0.001 µ |
| HOMA-β (%) | 91.89 ± 8.86 | 148.49 ± 34.61 | <0.001 µ |
| Adipokines & Inflammatory Markers | |||
| CRP (mg/L) | 1.03 ± 0.21 * | 4.05 ± 0.87 ** | <0.001 µ |
| IL-1β (pg/mL) | 2.40 ± 0.10 α | 2.58 ± 1.55 αα | 0.879 Ω |
| IL-6 (pg/mL) | 2.83 ± 0.36 β | 3.03 ± 0.23 ββ | 0.192 Ω |
| TNF-α (pg/mL) | 12.45 ± 0.73 € | 16.32 ± 1.70 €€ | 0.006 Ω |
| Adiponectin (µg/mL) | 13.86 ± 0.81 £ | 11.24 ± 0.61 | 0.019 Ω |
| Leptin (ng/mL) | 6.71 ± 0.69 ¥ | 25.11 ± 1.98 | <0.001 µ |
| Endothelial Dysfunction Markers | |||
| VCAM-1 (ng/mL) | 761.73 ± 31.60 | 789.37 ± 34.93 | 0.771 Ω |
| ICAM-1 (ng/mL) | 205.36 ± 8.62 | 246.81 ± 19.16 | 0.137 Ω |
| E-selectin (ng/mL) | 58.09 ± 4.53 | 175.88 ± 71.52 | <0.001 Ω |
| Fecal SCFAs | |||
| Acetic Acid (mM/100 mg) | 237.84 ± 20.19 | 221.80 ± 17.14 | 0.489 Ω |
| Propionic Acid (mM/100 mg) | 14.57 ± 1.91 | 11.68 ± 2.07 | 0.027 Ω |
| Butyric Acid (mM/100 mg) | 13.81 ± 1.44 | 12.18 ± 1.08 | 0.289 Ω |
| Characteristics | Normal Weight | Obesity | p-Value |
|---|---|---|---|
| Number (F/M) | 27 (12/15) | 34 (18/16) | Nd |
| Age (years) | 13.00 ± 0.28 | 13.61 ± 0.28 | 0.254 µ |
| Age Range (years) | 12–16 | 12–18 | Nd |
| Anthropometric | |||
| Weight (kg) | 44.83 ± 1.54 | 69.43 ± 2.46 | <0.001 Ω |
| Height (m) | 1.52 ± 0.01 | 1.55 ± 0.01 | 0.224 Ω |
| BMI (kg/m2) | 19.25 ± 0.42 | 28.78 ± 0.83 | <0.001 Ω |
| BMI pc | 52.60 ± 4.34 | 98.01 ± 0.36 | <0.001 µ |
| BMI pc Scale | <85 | >95 | Nd |
| WC | 68.91 ± 1.27 | 88.68 ± 1.57 | <0.001 µ |
| WC pc | 65.59 ± 5.53 | 97.93 ± 0.76 | <0.001 µ |
| Blood Pressure | |||
| SBP (mm Hg) | 93.84 ± 2.22 | 104.48 ± 1.73 | <0.001 µ |
| SBP pc | 20.49 ± 3.90 | 36.51 ± 4.73 | 0.021 µ |
| DBP (mm Hg) | 63.12 ± 1.41 | 68.30 ± 1.77 | 0.048 µ |
| DBP pc | 51.20 ± 3.89 | 62.52 ± 4.50 | 0.074 µ |
| Metabolic Factors | |||
| Fasting Glucose (mg/dL) | 91.45 ± 2.07 | 92.89 ± 1.68 | 0.345 µ |
| Triglycerides (mg/dL) | 91.14 ± 5.67 | 136.94 ± 8.33 | <0.001 µ |
| Total Cholesterol (mg/dL) | 161.98 ± 2.64 | 176.93 ± 5.20 | 0.041 µ |
| HDL (mg/dL) | 48.61 ± 1.84 | 43.86 ± 1.70 | 0.069 Ω |
| LDL (mg/dL) | 95.13 ± 2.35 | 105.68 ± 4.30 | 0.089 µ |
| Insulin (µIU/mL) | 8.64 ± 0.67 | 17.69 ± 1.51 | <0.001 µ |
| HOMA-IR | 1.93 ± 0.14 | 4.07 ± 0.36 | <0.001 µ |
| HOMA-β (%) | 126.69 ± 14.67 | 256.49 ± 38.70 | <0.001 µ |
| Adipokines & Inflammatory Markers | |||
| CRP (mg/L) | 2.27 ± 0.76 * | 3.15 ± 0.80 | 0.022 µ |
| IL-1β (pg/mL) | 2.40 ± 0.20 ∞ | 2.83 ± 0.44 ∞∞ | 0.926 µ |
| IL-6 (pg/mL) | 2.74 ± 0.25 α | 3.26 ± 0.51 αα | 0.824 µ |
| TNF-α (pg/mL) | 13.50 ± 1.23 β | 11.12 ± 0.86 ββ | 0.092 µ |
| Adiponectin (µg/mL) | 12.50 ± 1.08 | 10.49 ± 0.86 | 0.105 µ |
| Leptin (ng/mL) | 14.73 ± 4.19 | 34.13 ± 4.61 | <0.001 µ |
| Endothelial Dysfunction Markers | |||
| VCAM-1 (ng/mL) | 799.69 ± 47.67 | 792.23 ± 60.90 | 0.127 µ |
| ICAM-1 (ng/mL) | 182.10 ± 17.05 | 263.11 ± 17.40 | <0.001 µ |
| E-selectin (ng/mL) | 44.33 ± 3.35 | 103.12 ± 19.53 | <0.001 µ |
| Fecal SCFAs | |||
| Acetic Acid (mM/100mg) | 245.10 ± 28.42 | 238.76 ± 17.76 | 0.576 µ |
| Propionic Acid (mM/100mg) | 14.79 ± 3.25 | 8.50 ± 1.56 | 0.379 µ |
| Butyric Acid (mM/100mg) | 13.60 ± 1.82 | 16.65 ± 3.71 | 0.596 µ |
| Obese Children (Figure 6a) | Obese Adolescents (Figure 6b) | ||||
|---|---|---|---|---|---|
| No. | OTUs ID | Bacteria | No. | OTUs ID | Bacteria |
| 1 | 335 | Phascolarctobacterium | 1 | 1184 | Collinsella |
| 2 | 4007 | Peptococcus | 2 | 7367 | Prevotella |
| 3 | 6529 | Lactobacillus | 3 | 9022 | Lactobacillus |
| 4 | 7389 | Haemophilus | 4 | 9328 | Collinsella |
| 5 | 9453 | Prevotella | 5 | 10278 | Lactobacillus |
| 6 | 11321 | Phascolarctobacterium | 6 | 10560 | Collinsella |
| 7 | 12345 | Phascolarctobacterium | 7 | 12479 | Lactobacillus |
| 8 | 13188 | Prevotella | 8 | 12723 | Lachnospiraceae |
| 9 | 19296 | Bacteroides | 9 | 12974 | Lactobacillus |
| 10 | 19314 | Parabacteroides | 10 | 29566 | Sneathia |
| 11 | 21736 | Phascolarctobacterium | 11 | 128300 | Lactobacillus |
| 12 | 22231 | Haemophilus | 12 | 130468 | Lactobacillus |
| 13 | 24722 | Bacteroides | 13 | 130864 | Lactobacillus |
| 14 | 41229 | Sutterella | 14 | 133372 | Parvimonas |
| 15 | 157424 | Phascolarctobacterium | 15 | 137183 | Bifidobacteriaceae |
| 16 | 179261 | Sutterella | 16 | 225846 | Dialister |
| 17 | 183603 | Bacteroides | 17 | 236308 | Lactobacillus |
| 18 | 196604 | Catenibacterium | 18 | 272516 | Adlercreutzia |
| 19 | 215331 | Peptococcus | 19 | 292921 | Prevotella |
| 20 | 235591 | Lactobacillus | 20 | 354905 | Lactobacillus |
| 21 | 269937 | Prevotella | 21 | 383885 | Lactobacillus |
| 22 | 293883 | Phascolarctobacterium | 22 | 469663 | Atopobium |
| 23 | 309133 | Enterococcaceae | 23 | 566154 | Coriobacteriaceae |
| 24 | 339685 | Peptococcus | 24 | 568118 | Prevotella |
| 25 | 365496 | Bacteroides | 25 | 663885 | Prevotella |
| 26 | 370086 | Ruminococcaceae | 26 | 840914 | Prevotella |
| 27 | 403701 | Dialister | 27 | 851726 | Megasphaera |
| 28 | 524371 | Prevotella | 28 | 858535 | Coriobacteriaceae |
| 29 | 524884 | Eubacterium | 29 | 986513 | Clostridium |
| 30 | 583746 | Dialister | |||
| 31 | 587753 | Coriobacteriaceae | |||
| 32 | 639310 | Bifidobacterium | |||
| 33 | 716286 | Lactobacillus | |||
| 34 | 850218 | Phascolarctobacterium | |||
| 35 | 4226929 | Bacteroides | |||
| Taxa | This Work | Other Reports | Reference |
|---|---|---|---|
| Phylum Actinobacteria | |||
| Family Coriobacteriaceae | 3-fold more abundant in obese children and adolescents than normal weight | Higher abundance in human and mouse gut. Involved in bile acid metabolism and linked to gut barrier and metabolic dysfunctions | [44] |
| Isolated from the gut of Crohn’s disease suffering adult patient from Germany | [45] | ||
| Collinsella | 4-fold more abundant in obese adolescents than normal weight | More than 3-fold enriched in Swedish adult patients with symptomatic atherosclerosis | [21] |
| Abundant in American rheumatoid arthritis patients, strongly correlated with production of pro-inflammatory molecules and alters the gut permeability | [46] | ||
| Phylum Bacteroidetes | |||
| Order Bacteroidales | 3-fold more abundant in obese children than normal weight | Higher abundance in high-fat diet (HFD) fed Sprague-Dawley rats compared with low fat diet (LFD) fed rats (LFD vs. HFD, p < 0.01) | [47] |
| Reported in intestinal biopsies of American children and adults with Inflammatory Bowel Disease | [48] | ||
| Prevotella | 2-fold more abundant in obese adolescents than normal weight | Higher abundance of Prevotella in American adult rheumatoid arthritis patients. Induces inflammatory reactions by stimulating epithelial cells and production of interleukins | [49] |
| Phylum Firmicutes | |||
| Lactobacillus | 3-fold more abundant in obese children than normal weight | Different Lactobacillus species are associated with weight gain in farm animals | [50] |
| Higher abundance in obese French adults than normal weight | [51] | ||
| Blautia | 4-fold more abundant in obese adolescents than normal weight | Found higher abundance of Blautia in overweight and obese Mexican children | [5] |
| Coprococcus | 3-fold more abundant in obese adolescents than normal weight | Reported higher abundance of Coprococcus in overweight and obese Mexican children | [5] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nirmalkar, K.; Murugesan, S.; Pizano-Zárate, M.L.; Villalobos-Flores, L.E.; García-González, C.; Morales-Hernández, R.M.; Nuñez-Hernández, J.A.; Hernández-Quiroz, F.; Romero-Figueroa, M.d.S.; Hernández-Guerrero, C.; et al. Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients 2018, 10, 2009. https://doi.org/10.3390/nu10122009
Nirmalkar K, Murugesan S, Pizano-Zárate ML, Villalobos-Flores LE, García-González C, Morales-Hernández RM, Nuñez-Hernández JA, Hernández-Quiroz F, Romero-Figueroa MdS, Hernández-Guerrero C, et al. Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients. 2018; 10(12):2009. https://doi.org/10.3390/nu10122009
Chicago/Turabian StyleNirmalkar, Khemlal, Selvasankar Murugesan, María Luisa Pizano-Zárate, Loan Edel Villalobos-Flores, Cristina García-González, Rosa María Morales-Hernández, Jorge Arturo Nuñez-Hernández, Fernando Hernández-Quiroz, María del Socorro Romero-Figueroa, César Hernández-Guerrero, and et al. 2018. "Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents" Nutrients 10, no. 12: 2009. https://doi.org/10.3390/nu10122009
APA StyleNirmalkar, K., Murugesan, S., Pizano-Zárate, M. L., Villalobos-Flores, L. E., García-González, C., Morales-Hernández, R. M., Nuñez-Hernández, J. A., Hernández-Quiroz, F., Romero-Figueroa, M. d. S., Hernández-Guerrero, C., Hoyo-Vadillo, C., & García-Mena, J. (2018). Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients, 10(12), 2009. https://doi.org/10.3390/nu10122009