Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.2.1. Chronic Experiment
2.2.2. Acute Experiment
2.3. Analytical Methods
2.3.1. Muscle Homogenization
2.3.2. Mitochondrial Enzyme Activity
2.3.3. Western Blotting
2.4. Statistical Analysis
3. Results
3.1. Chronic Experiment
3.1.1. Food Consumption and Body Weight
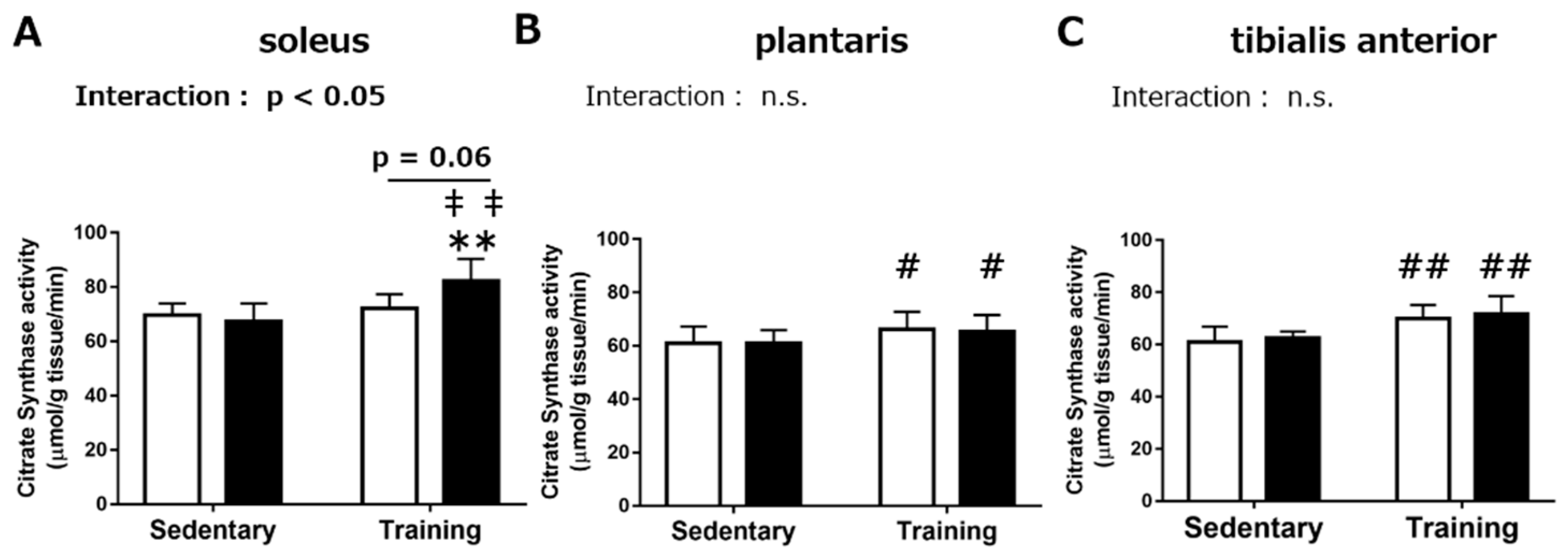
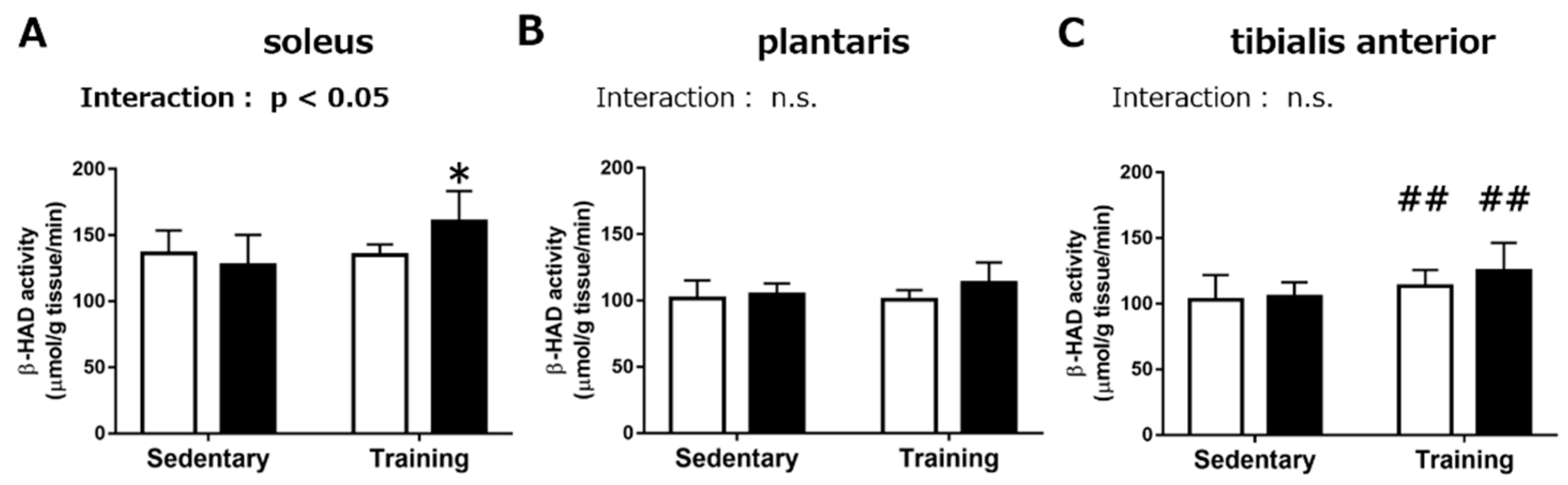
3.1.2. Maximal Activities of Mitochondrial Enzymes
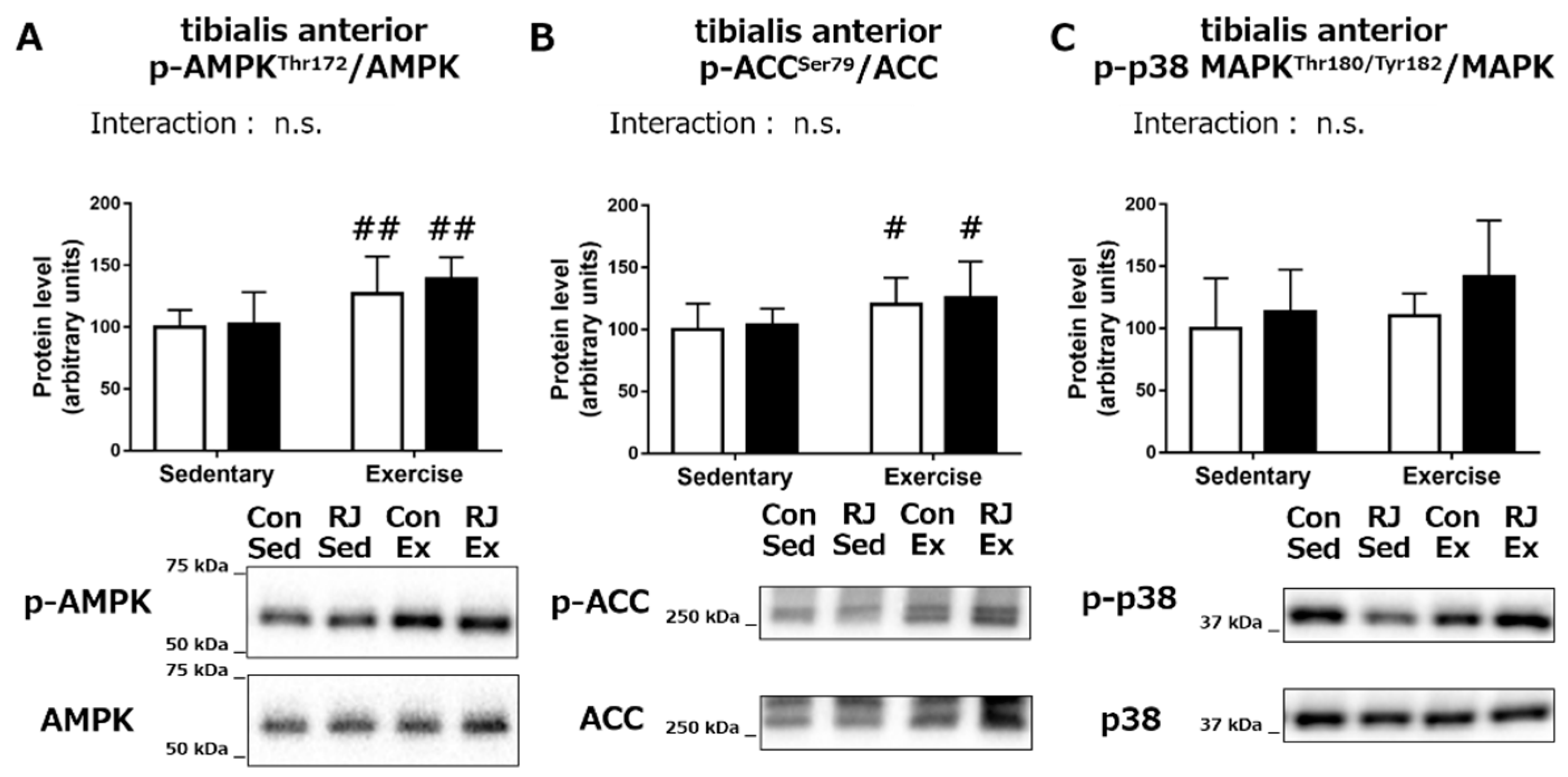
3.2. Acute Experiment
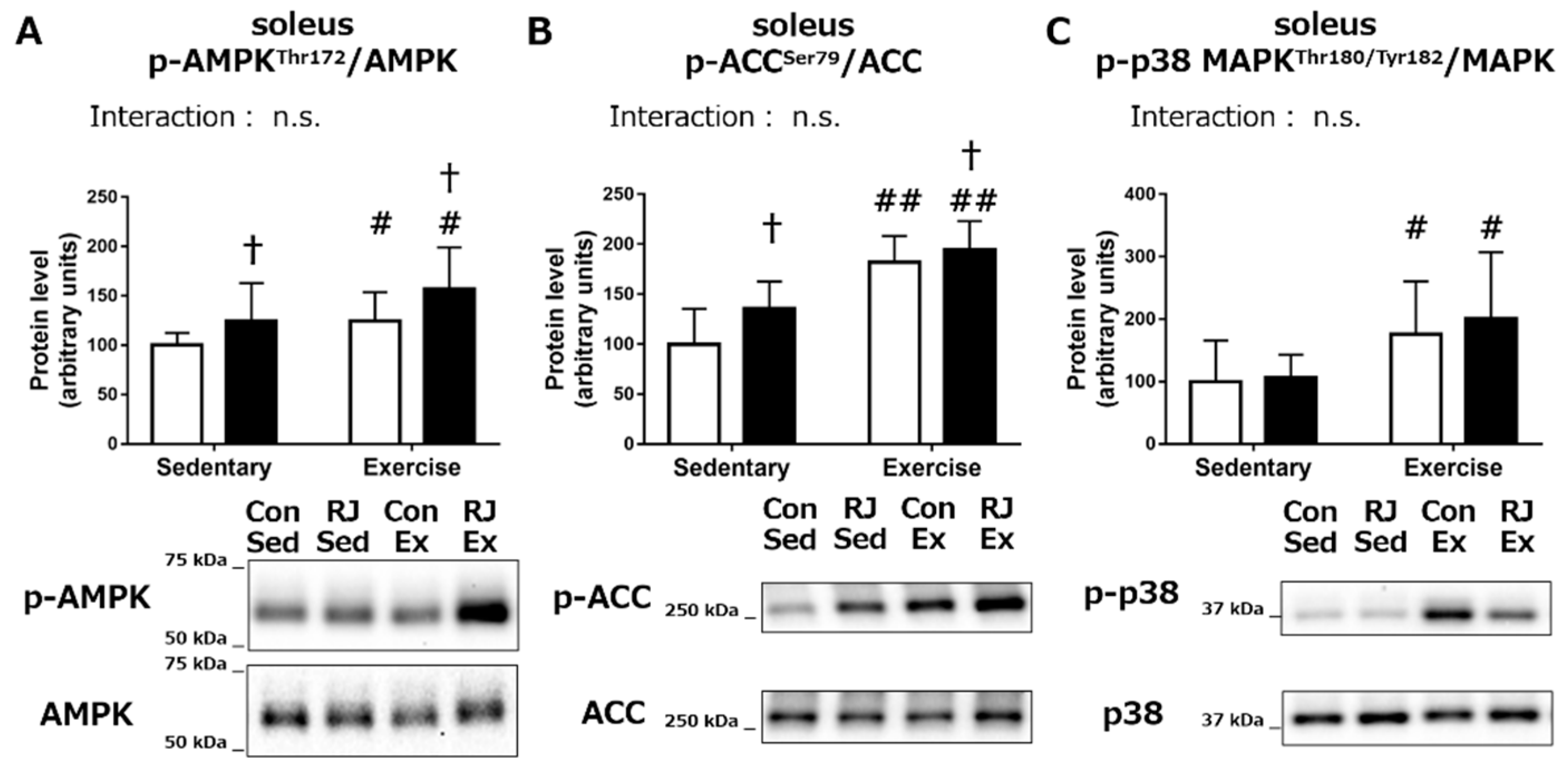
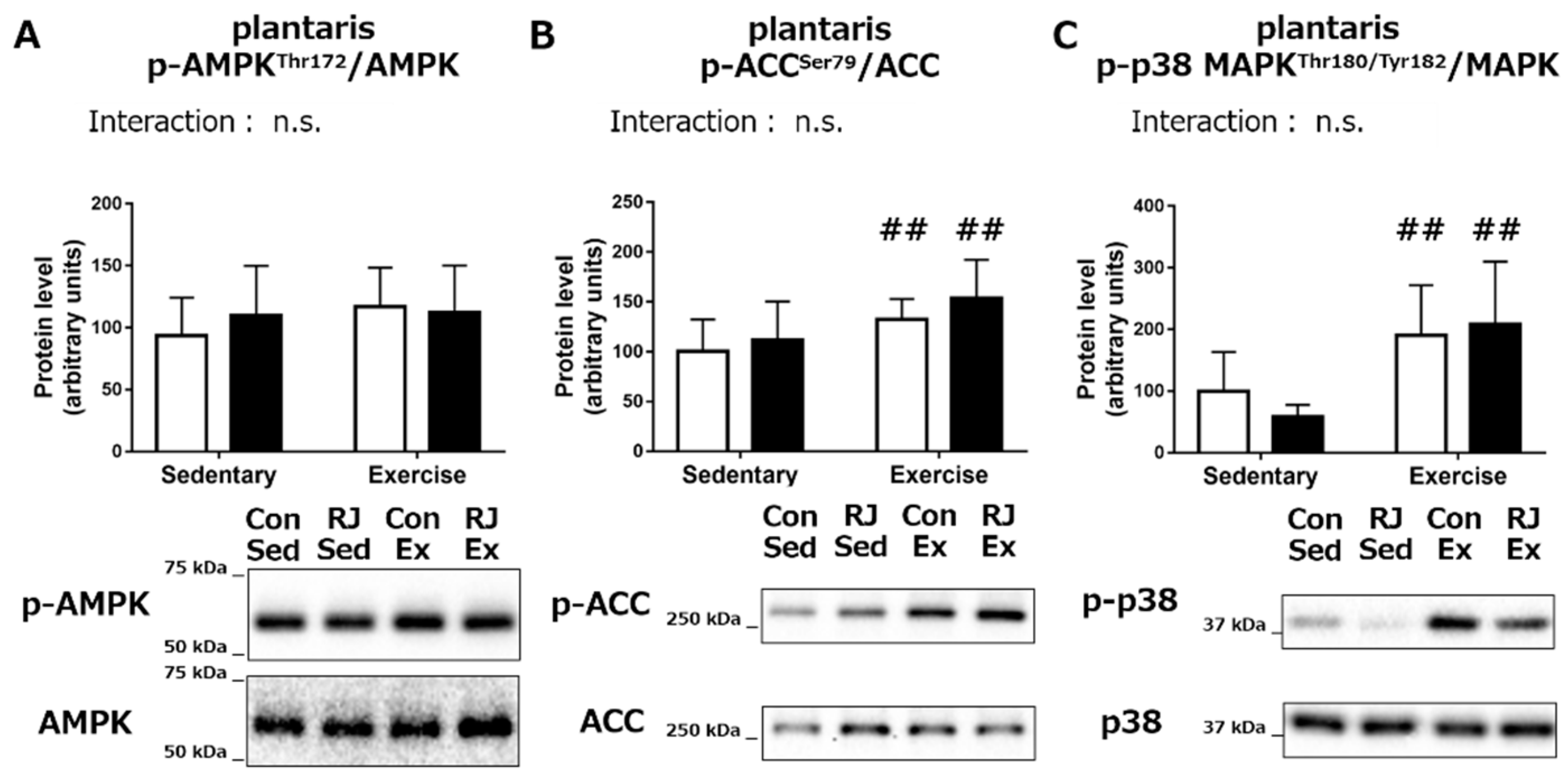
Phosphorylation Status of the Proteins Involved in Mitochondrial Biogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Holloszy, J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967, 242, 2278–2282. [Google Scholar] [PubMed]
- Fitts, R.; Booth, F.; Winder, W.; Holloszy, J. Skeletal muscle respiratory capacity, endurance, and glycogen utilization. Am. J. Physiol. 1975, 228, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Ibebunjo, C.; Chick, J.M.; Kendall, T.; Eash, J.K.; Li, C.; Zhang, Y.; Vickers, C.; Wu, Z.; Clarke, B.A.; Shi, J.; et al. Genomic and Proteomic Profiling Reveals Reduced Mitochondrial Function and Disruption of the Neuromuscular Junction Driving Rat Sarcopenia. Mol. Cell Biol. 2013, 33, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Turner, N. Mitochondrial dysfunction and insulin resistance: An update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef] [PubMed]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-Chain Amino Acid Supplementation Promotes Survival and Supports Cardiac and Skeletal Muscle Mitochondrial Biogenesis in Middle-Aged Mice. Cell Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, Y.; Tamura, Y.; Takahashi, Y.; Masuda, H.; Hoshino, D.; Kitaoka, Y.; Saito, N.; Nakamura, H.; Takeda, Y.; Hatta, H. Pre-exercise casein peptide supplementation enhances endurance training-induced mitochondrial enzyme activity in slow twitch muscle, but not fast twitch muscle of high fat diet-fed mice. J. Phys. Fit. Sport. Med. 2015, 4, 377–384. [Google Scholar] [CrossRef]
- Liang, C.; Curry, B.J.; Brown, P.L.; Zemel, M.B. Leucine Modulates Mitochondrial Biogenesis and SIRT1-AMPK Signaling in C2C12 Myotubes. J. Nutr. Metab. 2014, 2014, 239750. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, Y.; Sakata, Y.; Yago, T.; Nakamura, H.; Shimizu, T.; Takeda, Y. Effects of Glucose with Casein Peptide Supplementation on Post-Exercise Muscle Glycogen Resynthesis in C57BL/6J Mice. Nutrients 2018, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, M.; Kumagai, A.; Hirata, H.; Soga, M.; Yamashita, Y.; Ueda, M.; Ashida, H.; Tsuda, T. 10-Hydroxy-2-decenoic acid, a unique medium-chain fatty acid, activates 5’-AMP-activated protein kinase in L6 myotubes and mice. Mol. Nutr. Food Res. 2013, 57, 1794–1802. [Google Scholar] [CrossRef] [PubMed]
- Watadani, R.; Kotoh, J.; Sasaki, D.; Someya, A.; Matsumoto, K.; Maeda, A. 10-Hydroxy-2-decenoic acid, a natural product, improves hyperglycemia and insulin resistance in obese/diabetic KK-Ay mice, but does not prevent obesity. J. Vet. Med. Sci. 2017, 79, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Hayashi, K.; Watadani, R.; Okano, Y.; Tanimura, K.; Kotoh, J.; Sasaki, D.; Matsumoto, K.; Maeda, A. Royal jelly improves hyperglycemia in obese/diabetic KK-Ay mice. J. Vet. Med. Sci. 2017, 79, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Ljubicic, V.; Hood, D.A. Specific attenuation of protein kinase phosphorylation in muscle with a high mitochondrial content. Am. J. Physiol. Metab. 2009, 297, E749–E758. [Google Scholar] [CrossRef] [PubMed]
- Augusto, V.; Padovani, C.R.; Rocha Campos, G. Skeletal muscle fiber types in C57Bl6J mice. Braz. J. Morphol. Sci. 2004, 27, 89–94. [Google Scholar]
- Denies, M.S.; Johnson, J.; Maliphol, A.B.; Bruno, M.; Kim, A.; Rizvi, A.; Rustici, K.; Medler, S. Diet-induced obesity alters skeletal muscle fiber types of male but not female mice. Physiol. Rep. 2014, 2, e00204. [Google Scholar] [CrossRef] [PubMed]
- Kammoun, M.; Cassar-Malek, I.; Meunier, B.; Picard, B. A simplified immunohistochemical classification of skeletal muscle fibres in mouse. Eur. J. Histochem. 2014, 58, 2254. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Guo, H.; Guo, Y.; Ebihara, S.; Asada, M.; Ohrui, T.; Furukawa, K.; Ichinose, M.; Yanai, K.; Kudo, Y.; et al. Royal Jelly Prevents the Progression of Sarcopenia in Aged Mice In Vivo and In Vitro. J. Gerontol. Biol. Sci. Med. Sci. 2013, 68, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, Y.; Takeda, K.; Tamura, Y.; Fujimaki, S.; Takemasa, T.; Hatta, H. Nrf2 deficiency does not affect denervation-induced alterations in mitochondrial fission and fusion proteins in skeletal muscle. Physiol. Rep. 2016, 4, e13064. [Google Scholar] [CrossRef] [PubMed]
- Srere, P.A. [1] Citrate synthase: [EC 4.1.3.7. Citrate oxaloacetate-lyase (CoA-acetylating)]. Methods Enzymol. 1969, 13, 3–11. [Google Scholar] [CrossRef]
- Bass, A.; Brdiczka, D.; Eyer, P.; Hofer, S.; Pette, D. Metabolic differentiation of distinct muscle types at the level of enzymatic organization. Eur. J. Biochem. 1969, 10, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Matsunaga, Y.; Tamura, Y.; Terada, S.; Hatta, H. Pre-Exercise High-Fat Diet for 3 Days Affects Post-Exercise Skeletal Muscle Glycogen Repletion. J. Nutr. Sci. Vitaminol. 2017, 63, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Daa Schroder, H.; Boushel, R.; Wulff Helge, J.; Dela, F.; et al. Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. Authors. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef] [PubMed]
- Silici, S.; Ekmekcioglu, O.; Eraslan, G.; Deniz, K. Antioxidative effect of royal jelly in cisplatin-induced testes damage. Urology. 2009, 74, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Silici, S.; Ekmekcioglu, O.; Kanbur, M.; Deniz, K. The protective effect of royal jelly against cisplatin-induced renal oxidative stress in rats. World. J. Urol. 2011, 29, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 7074209. [Google Scholar] [CrossRef] [PubMed]
- Irrcher, I.; Ljubicic, V.; Hood, D.A. Interactions between ROS and AMP kinase activity in the regulation of PGC-1alpha transcription in skeletal muscle cells. Am. J. Physiol. Cell. Physiol. 2009, 296, C116–C123. [Google Scholar] [CrossRef] [PubMed]
- Morales-Alamo, D.; Ponce-González, J.G.; Guadalupe-Grau, A.; Rodríguez-García, L.; Santana, A.; Cusso, R.; Guerrero, M.; Dorado, C.; Guerra, B.; Calbet, J.A. Critical role for free radicals on sprint exercise-induced CaMKII and AMPKα phosphorylation in human skeletal muscle. J. Appl. Physiol. 2013, 114, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Umanskaya, A.; Santulli, G.; Xie, W.; Andersson, D.C.; Reiken, S.R.; Marks, A.R. Genetically enhancing mitochondrial antioxidant activity improves muscle function in aging. Proc. Natl. Acad. Sci. USA 2014, 111, 15250–15255. [Google Scholar] [CrossRef] [PubMed]
- Mungai, P.T.; Waypa, G.B.; Jairaman, A.; Prakriya, M.; Dokic, D.; Ball, M.K.; Schumacker, P.T. Hypoxia triggers AMPK activation through reactive oxygen species-mediated activation of calcium release-activated calcium channels. Mol. Cell. Biol. 2011, 31, 3531–3545. [Google Scholar] [CrossRef] [PubMed]
- Onan, M.C.; Fisher, J.S.; Ju, J.-S.; Fuchs, B.C.; Bode, B.P. Type I diabetes affects skeletal muscle glutamine uptake in a fiber-specific manner. Exp. Biol. Med. 2005, 230, 606–611. [Google Scholar] [CrossRef]
- Hodson, N.; Brown, T.; Joanisse, S.; Aguirre, N.; West, D.; Moore, D.; Baar, K.; Breen, L.; Philp, A. Characterisation of L-Type Amino Acid Transporter 1 (LAT1) Expression in Human Skeletal Muscle by Immunofluorescent Microscopy. Nutrients 2017, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Staron, R.S.; Hagerman, F.C.; Hikida, R.S.; Murray, T.F.; Hostler, D.P.; Crill, M.T.; Ragg, K.E.; Toma, K. Fiber type composition of the vastus lateralis muscle of young men and women. J. Histochem. Cytochem. 2000, 48, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Simoneau, J.A.; Lortie, G.; Boulay, M.R.; Marcotte, M.; Thibault, M.C.; Bouchard, C. Human skeletal muscle fiber type alteration with high-intensity intermittent training. Eur. J. Appl. Physiol. Occup. Physiol. 1985, 54, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Wang, H.; Pei, Y.; Li, Y.; Wu, H.; Song, Y.; Guo, Q.; Guo, H.; Fukushima, S.; Tatefuji, T.; et al. Effects of protease-treated royal jelly on muscle strength in elderly nursing home residents: A randomized, double-blind, placebo-controlled, dose-response study. Sci. Rep. 2017, 7, 11416. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macronutrients | Royal Jelly Specific Fatty Acids | ||
| Water | 2.2 g | (E)-10-hydroxy-2-decenoic acid | 4.3% |
| Protein | 40.9 g | 10-hydroxydecanoic acid | 0.94% |
| Fat | 5.5 g | ||
| Minerals | 2.8 g | ||
| Ash | 2.8 g | ||
| Carbohydrate | 48.6 g | ||
| Calories | 408 kcal | ||
| Amino Acids | Free Amino Acids | ||
| Arginine | 2.01 g | Free Arginine | 0.07 g |
| Lysine | 2.83 g | Free Lysine | 0.67 g |
| Histidine | 1.07 g | Free Histidine | 0.02 g |
| Phenylalanine | 1.70 g | Free Phenylalanine | 0.004 g |
| Tyrosine | 1.65 g | Free Tyrosine | 0.006 g |
| Leucine | 2.88 g | Free Leucine | 0.005 g |
| Isoleucine | 1.83 g | Free Isoleucine | 0.005 g |
| Methionine | 1.03 g | Free Methionine | not detected |
| Valine | 2.09 g | Free Valine | 0.01 g |
| Alanine | 1.20 g | Free Alanine | 0.01 g |
| Glycine | 1.29 g | Free Glycine | 0.01 g |
| Proline | 1. 69 g | Free Proline | 0.67 g |
| Glutamic acid | 3.84 g | Free Glutamic acid | 0.14 g |
| Serine | 2.31 g | Free Serine | 0.004 g |
| Threonine | 1.72 g | Free Threonine | 0.002 g |
| Aspartic acid | 6.82 g | Free Aspartic acid | 0.006 g |
| Tryptophan | 0.49 g | Free Tryptophan | not detected |
| Cysteine | 0.40 g | Free Cysteine | not detected |
| Title | Con + Sed Group | RJ + Sed Group | Con + Tr Group | RJ + Tr Group |
|---|---|---|---|---|
| Initial body weight (g) | 38.6 ± 0.7 | 38.1 ± 0.9 | 37.7 ± 0.8 | 38.4 ± 0.5 |
| Final body weight (g) | 40.5 ± 0.8 | 40.2 ± 0.7 | 39.1 ± 0.9 | 39.3 ± 0.6 |
| Δ body weight (g) | 2.0 ± 0.3 | 2.0 ± 0.5 | 1.4 ± 0.6 | 0.9 ± 0.7 |
| Total food consumption (kcal/g body weight) | 9.1 ± 0.2 | 9.5 ± 0.4 | 9.1 ± 0.5 | 9.1 ± 0.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, Y.; Hijikata, K.; Seike, K.; Nakano, S.; Banjo, M.; Sato, Y.; Takahashi, K.; Hatta, H. Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle. Nutrients 2018, 10, 1735. https://doi.org/10.3390/nu10111735
Takahashi Y, Hijikata K, Seike K, Nakano S, Banjo M, Sato Y, Takahashi K, Hatta H. Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle. Nutrients. 2018; 10(11):1735. https://doi.org/10.3390/nu10111735
Chicago/Turabian StyleTakahashi, Yumiko, Kamiyu Hijikata, Kohei Seike, Suguru Nakano, Mai Banjo, Yosuke Sato, Kenya Takahashi, and Hideo Hatta. 2018. "Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle" Nutrients 10, no. 11: 1735. https://doi.org/10.3390/nu10111735
APA StyleTakahashi, Y., Hijikata, K., Seike, K., Nakano, S., Banjo, M., Sato, Y., Takahashi, K., & Hatta, H. (2018). Effects of Royal Jelly Administration on Endurance Training-Induced Mitochondrial Adaptations in Skeletal Muscle. Nutrients, 10(11), 1735. https://doi.org/10.3390/nu10111735