DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl Bark Extracts, Improves Hyperuricemia through Inhibition of Xanthine Oxidase Activity and Increasing Urine Excretion



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. DKB114 Preparation
2.2. Animals
2.3. Sample Treatment in Normal Rats
2.4. Hyperuricemia Induction and Sample Treatment
2.5. Collection of Serum, Urine, and Tissues
2.6. Analysis of Uric Acid Levels in Serum, Urine, and Liver Tissues
2.7. In Vitro and In Vivo XOD Inhibition Assay
2.8. Western Blotting Analysis of Kidney Samples
2.9. Cell Culture
2.10. Establishment of hURAT1-Expressing Oocytes and HEK293 Cells
2.11. Uptake Experiment in hURAT1-Expressing Oocytes and HEK293 Cells
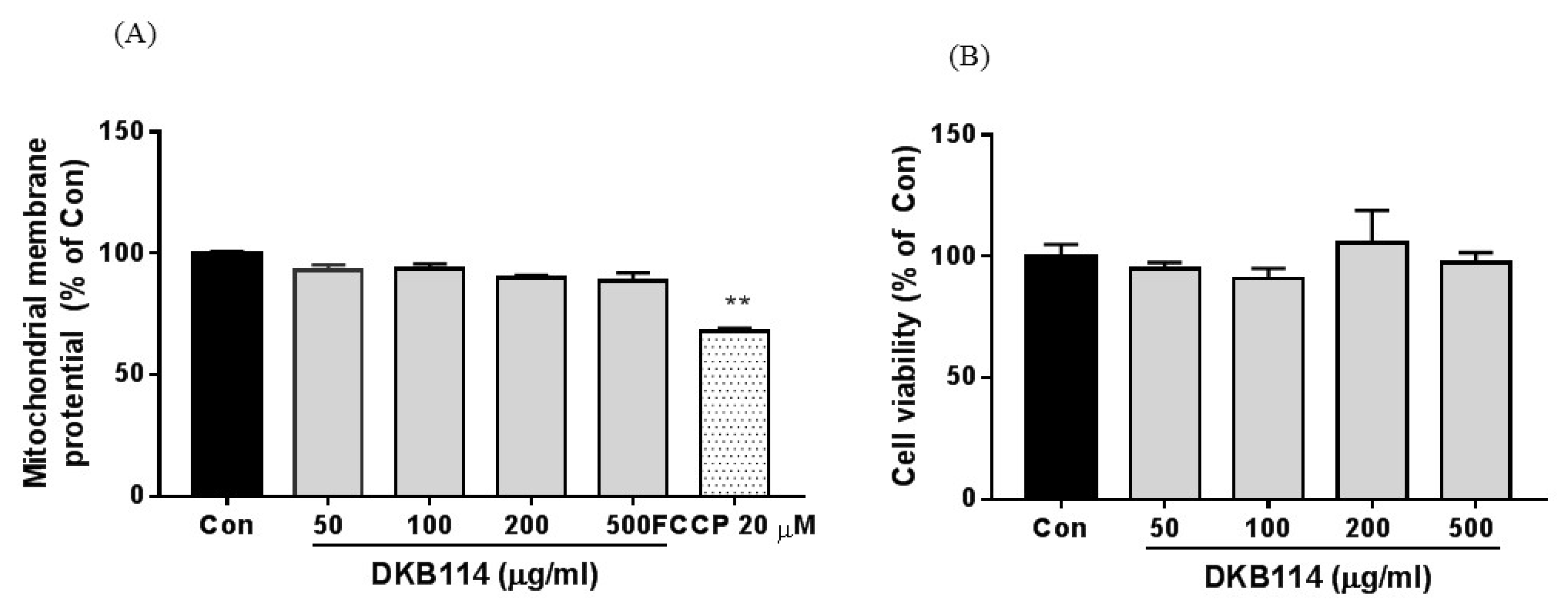
2.12. Mitochondrial Toxicity Study in HepG2 Cells
2.13. Statistical Analysis
3. Results
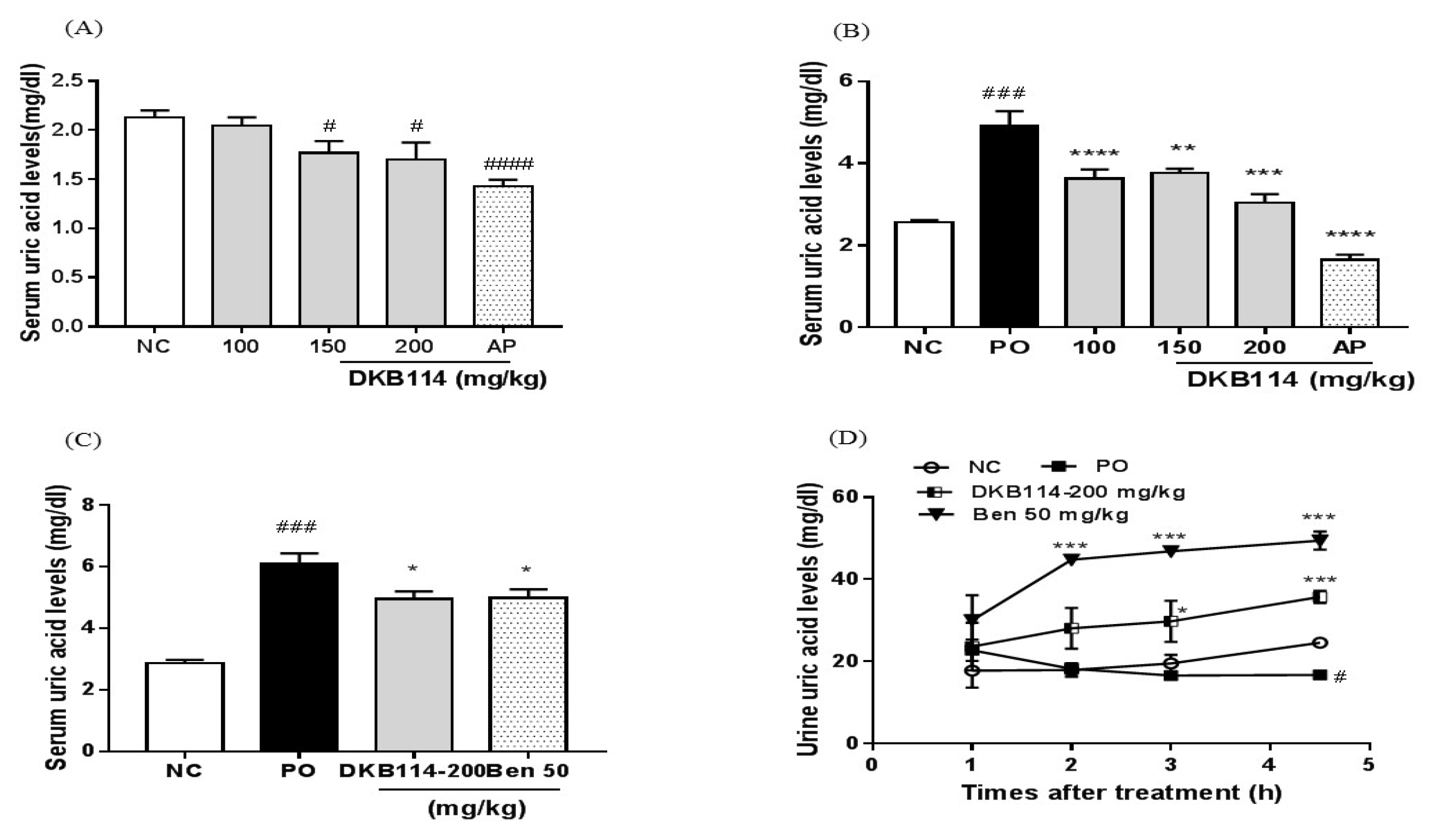
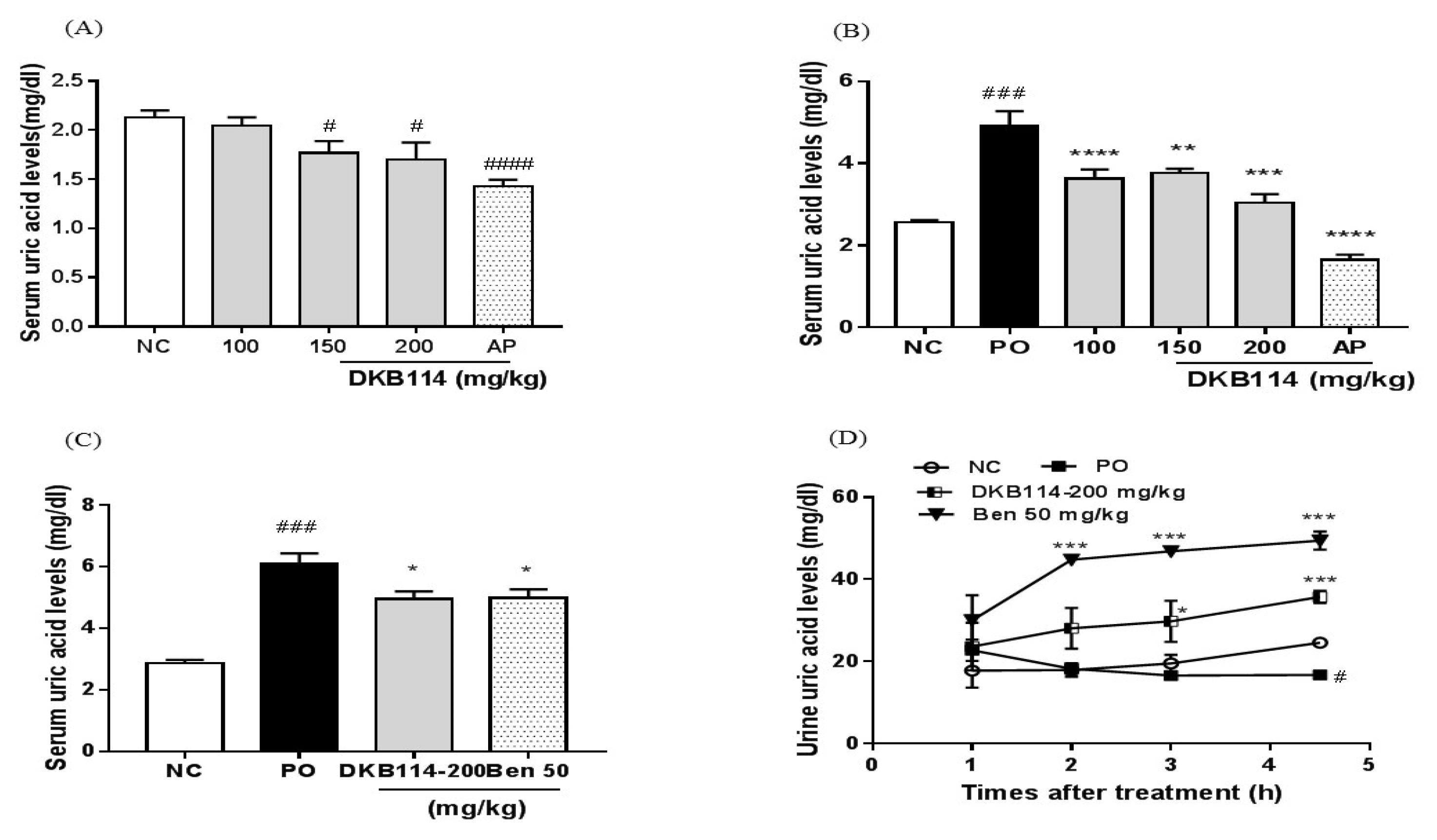
3.1. Effects of DKB114 on Serum and Urinary Uric Acid Levels in Normal Rats and Rats with PO-Induced Hyperuricemia
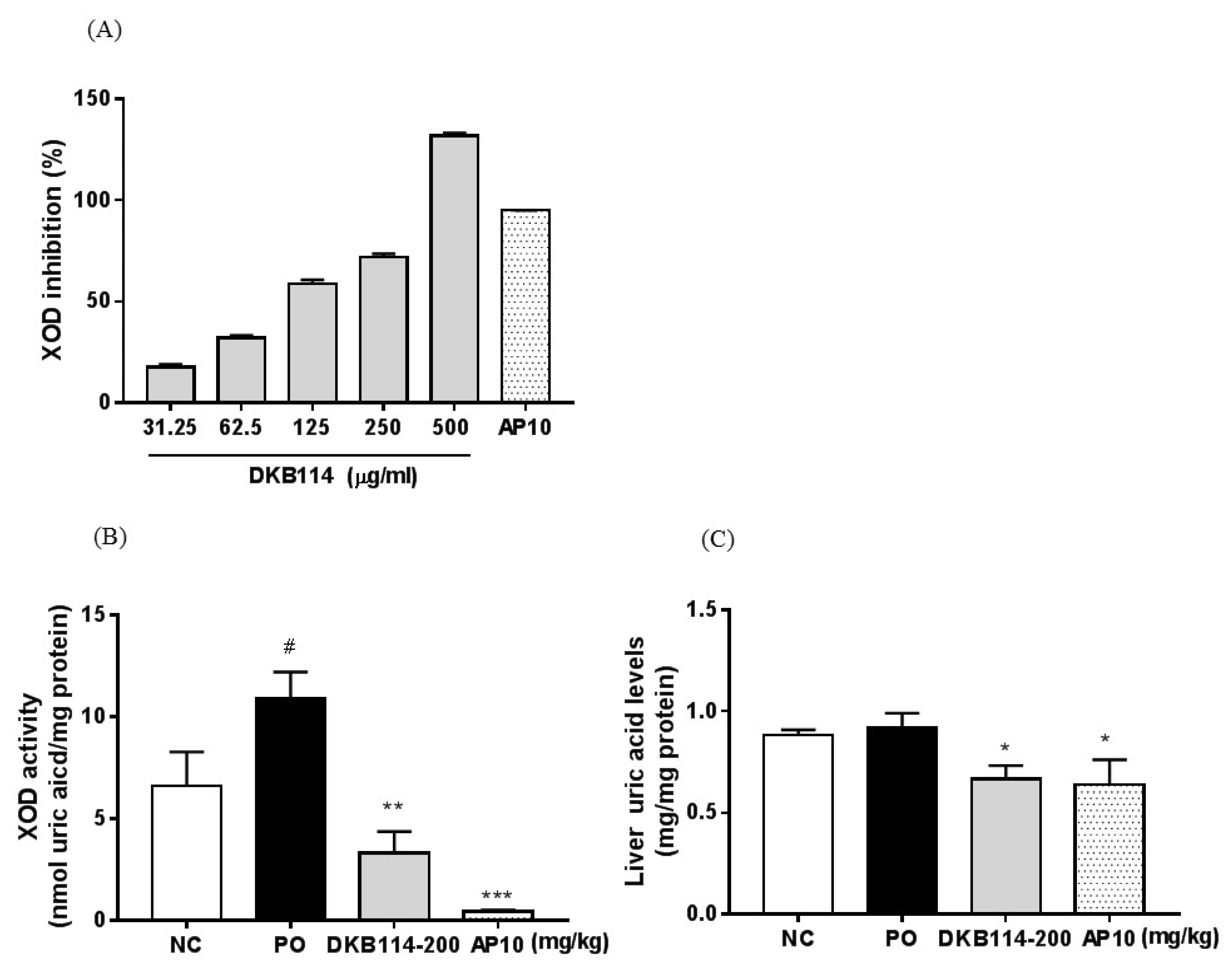
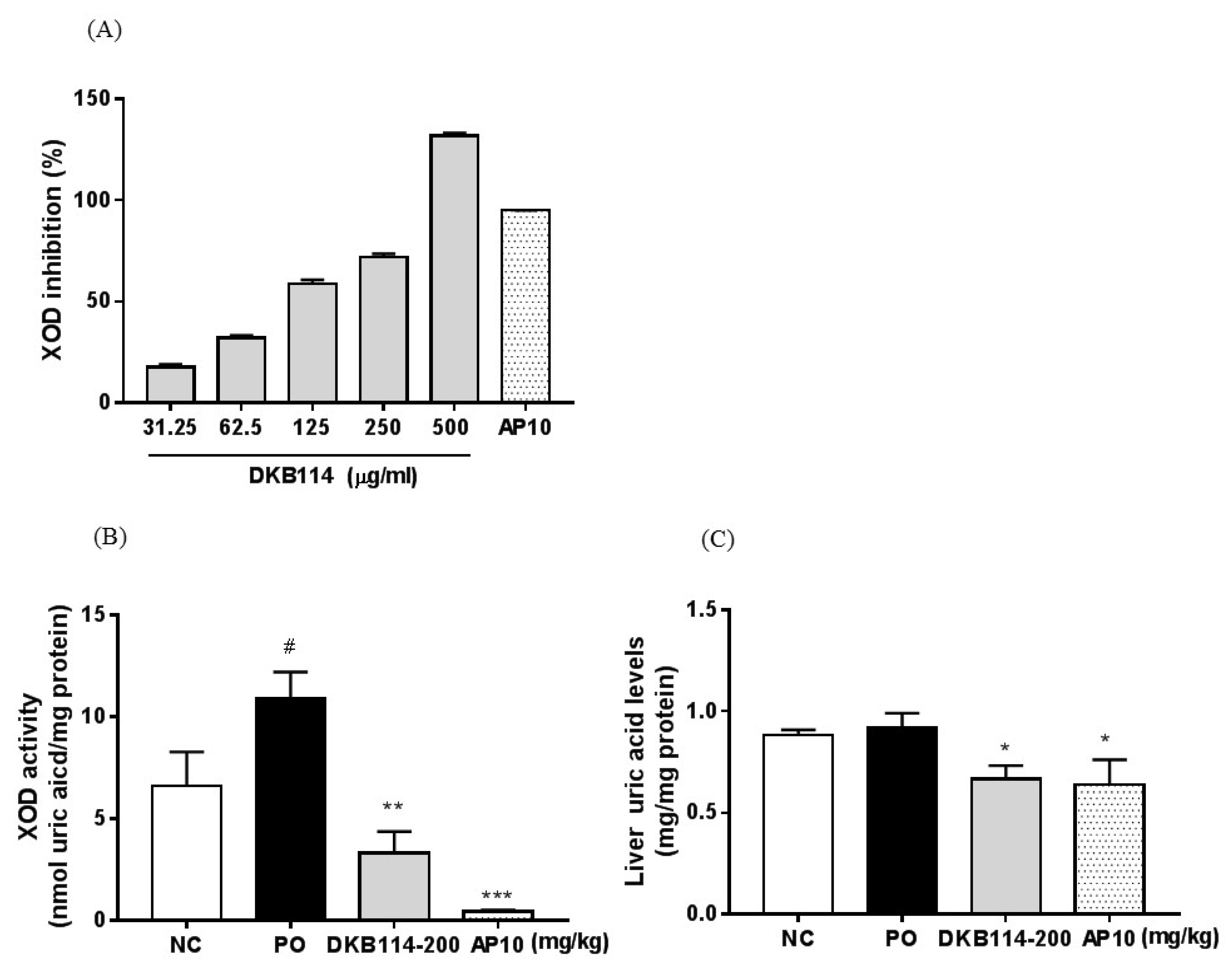
3.2. Effects of DKB114 on XOD Inhibition Activity in Vitro/Vivo and Liver Uric Acid Levels
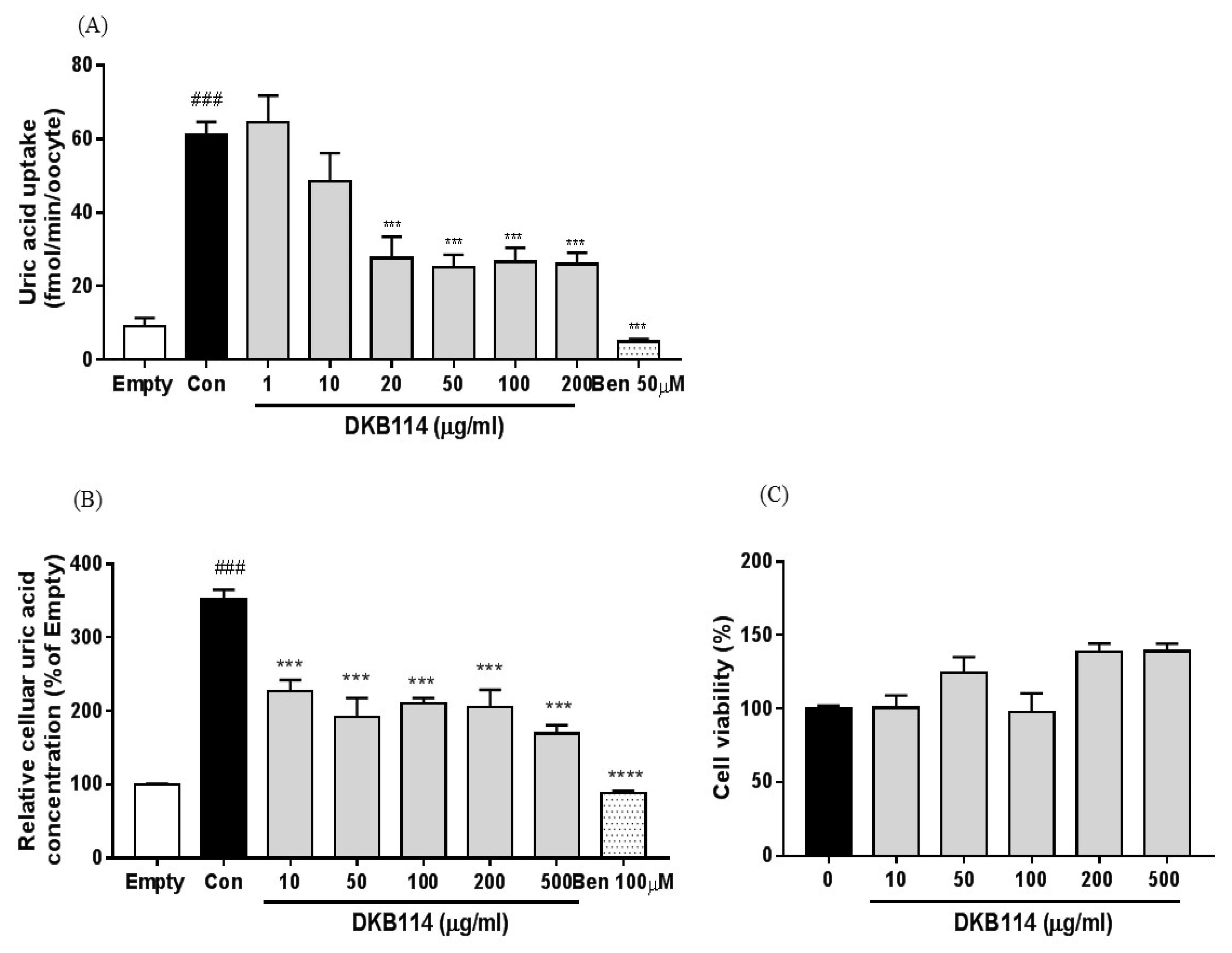
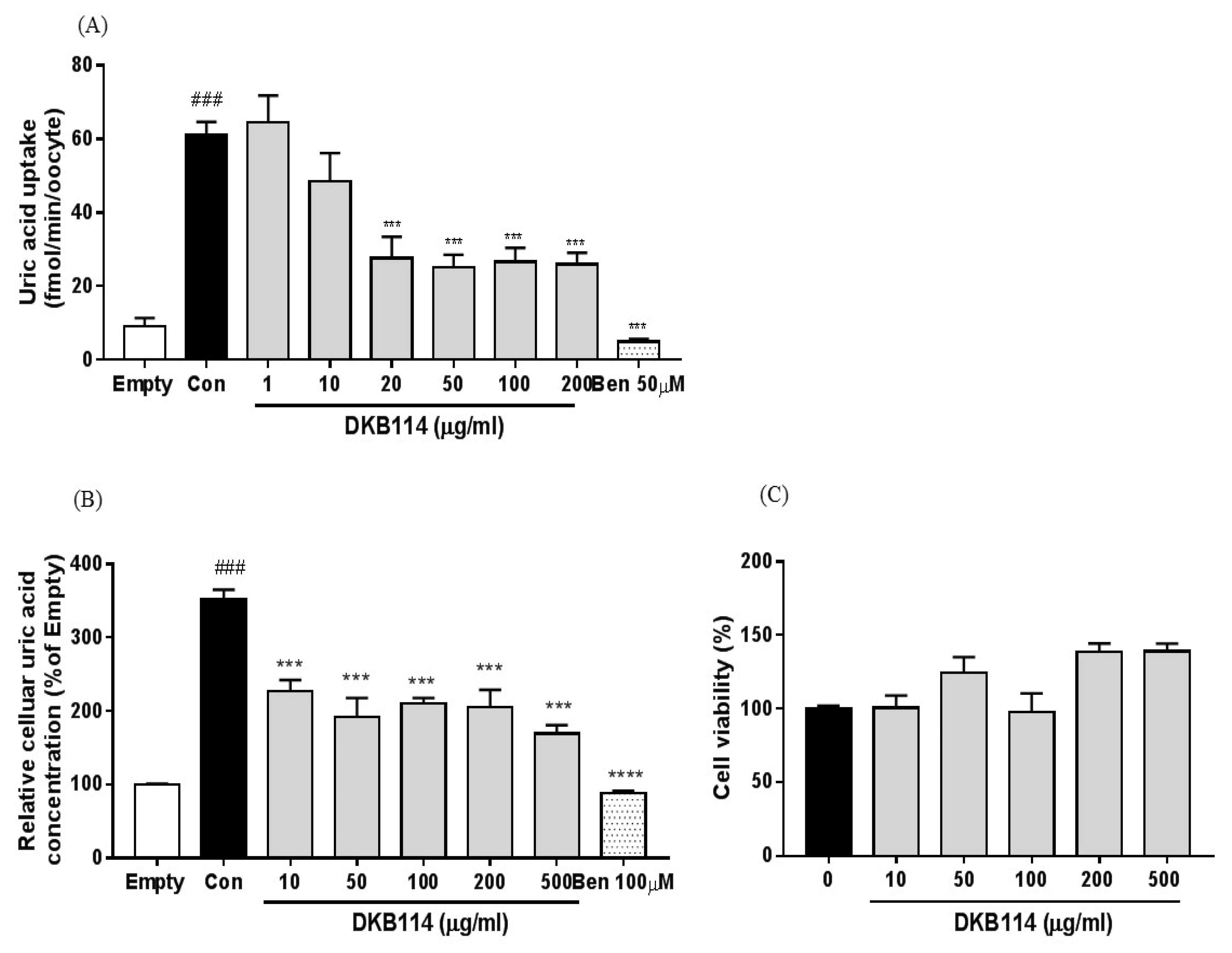
3.3. Effects of DKB114 on Uric Acid Uptake in hURAT1-Expressing Oocytes and HEK293 Cells
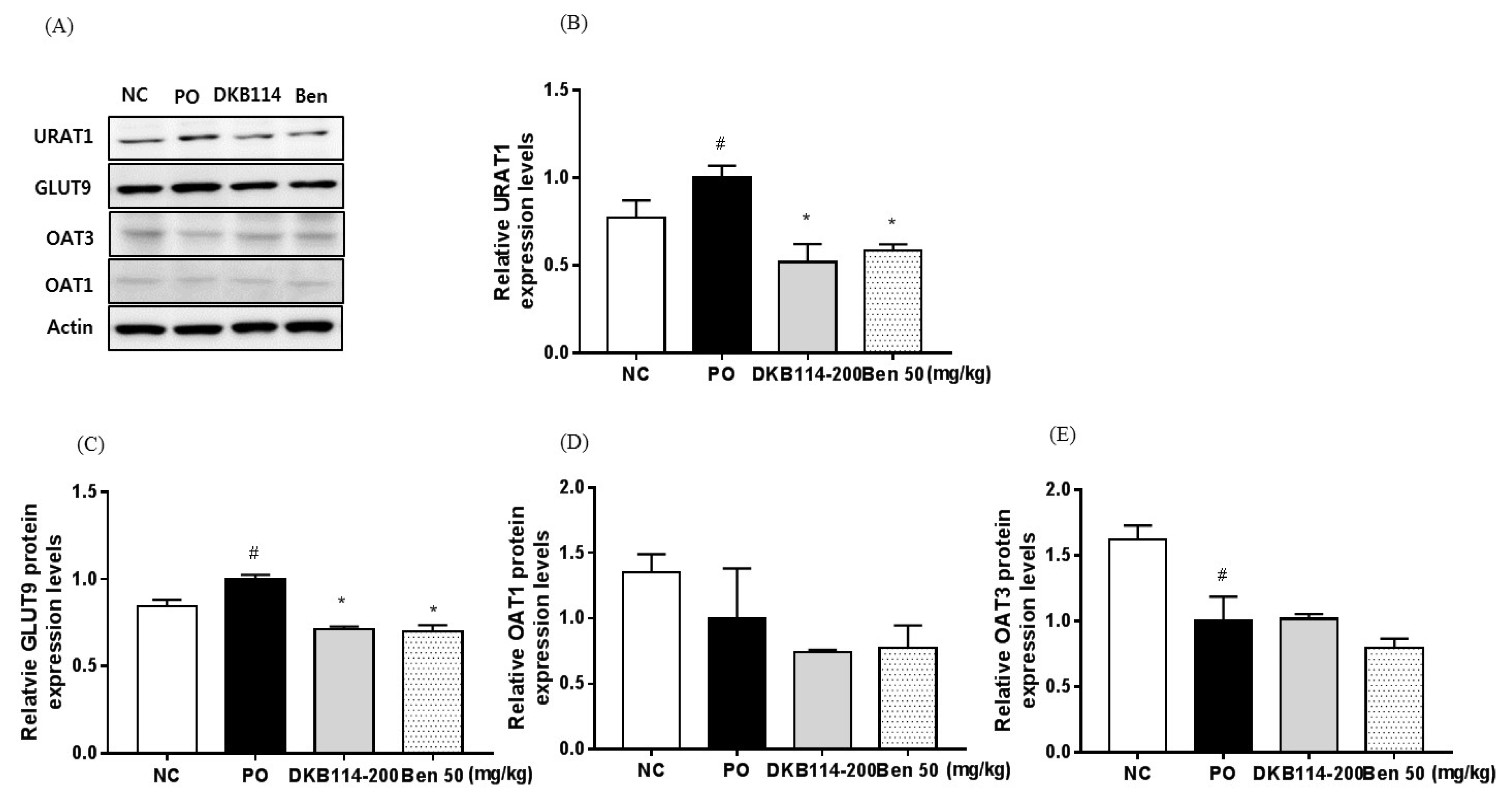
3.4. Effects of DKB114 on the Expression of Uric Acid Transporters in Rats with PO-Induced Hyperuricemia
3.5. Effects of DKB114 on Mitochondrial Toxicity in HepG2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.; Zhao, M.; Xin, Y.; Liu, J.; Wang, M.; Zhao, C. 1H NMR and MS based metabolomics study of the therapeutic effect of Cortex Fraxini on hyperuricemic rats. J. Ethnopharmacol. 2016, 185, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Huang, P.; Zhu, C.M.; Yuan, X.; Yu, C.H. Anti-hyperuricemic and nephroprotective effects of Modified Simiao Decoction in hyperuricemic mice. J. Ethnopharmacol. 2012, 142, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Lemos Lima Rde, C.; Ferrari, F.C.; de Souza, M.R.; de Sá Pereira, B.M.; de Paula, C.A.; Saúde-Guimarães, D.A. Effects of extracts of leaves from Sparattosperma leucanthum on hyperuricemia and gouty arthritis. J. Ethnopharmacol. 2015, 161, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Zhang, M.; Chen, D.; Shuai, O.; Chen, S.; Su, J.; Jiao, C.; Feng, D.; Xie, Y. Actions of water extract from Cordyceps militaris in hyperuricemic mice induced by potassium oxonate combined with hypoxanthine. J. Ethnopharmacol. 2016, 194, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Mount, D.B.; Reginato, A.M. Pathogenesis of gout. Ann. Intern. Med. 2005, 143, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Rashidi, M.R.; Keshavarz, S.A.; Mahboob, S.A.; Eshraghian, M.R.; Shahi, M.M. Effects of onion on serum uric acid levels and hepatic xanthine dehydrogenase/xanthine oxidase activiti2001es in hyperuricemic rats. Pak. J. Biol. Sci. 2008, 11, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Fukunari, A.; Okamoto, K.; Nishino, T.B.; Eger, T.; Pai, E.F.; Kamezawa, M.; Yamada, I.; Kato, N. Y-700 [1-[3-Cyano-4-(2,2-dimethylpropoxy)phenyl]-1H-pyrazole-4-carboxylic acid]: A potent xanthine oxidoreductase inhibitor with hepatic excretion. J. Pharmacol. Exp. Ther. 2004, 311, 519–528. [Google Scholar] [CrossRef] [PubMed]
- So, A.; Thorens, B. Uric acid transport and disease. J. Clin. Investig. 2010, 120, 1791–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibuchi, S.; Morimoto, H.; Oe, T.; Ikebe, T.; Inoue, H.; Fukunari, A.; Kamezawa, M.; Yamada, I.; Naka, Y. Synthesis and structure-activity relationships of 1-phenylpyrazoles as xanthine oxidase inhibitors. Bioorg. Med. Chem. Lett. 2001, 11, 879–882. [Google Scholar] [CrossRef]
- Azevedo, V.F.; Buiar, P.G.; Giovanella, L.H.; Severo, C.R.; Carvalho, M. Allopurinol, benzbromarone, or a combination in treating patients with gout: Analysis of a series of outpatients. Int. J. Rheumatol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Bardin, T. Current management of gout in patients unresponsive or allergic to allopurinol. Jt. Bone Spine 2004, 71, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Nivorozhkin, A.; Szabo, C. Therapeutic effects of xanthine oxidase inhibitors: Renaissance half a century after the discovery of allopurinol. Pharmacol. Rev. 2006, 58, 87–114. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Zhang, C.; Song, H. Natural Products Improving Hyperuricemia with Hepatorenal Dual Effects. Evid. Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Li, J.; You, T.; Hu, C. Anti-inflammatory and immunomodulatory activities of the extracts from the inflorescence of Chrysanthemum indicum Linne. J. Ethnopharmacol. 2005, 101, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Toguchida, I.; Harima, S.; Yoshikawa, M. Medicinal flowers. VI. Absolute stereostructures of two new flavanone glycosides and a phenylbutanoid glycoside from the flowers of Chrysanthemum indicum L.: Their inhibitory activities for rat lens aldose reductase. Chem. Pharm. Bull. 2002, 50, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Safdar, M.; Ali Khan, M.M.; Khattak, K.N.; Anderson, R.A. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 2003, 26, 3215–3218. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Trikha, P.; Athar, M.; Raisuddin, S. Inhibition of benzo[a]pyrene-and cyclophoshamide-induced mutagenicity by Cinnamomum cassia. Mutat. Res. 2001, 480, 179–188. [Google Scholar] [CrossRef]
- Zhao, X.; Zhu, J.X.; Mo, S.F.; Pan, Y.; Kong, L.D. Effects of cassia oil on serum and hepatic uric acid levels in oxonate-induced mice and xanthine dehydrogenase and xanthine oxidase activities in mouse liver. J. Ethnopharmacol. 2006, 103, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Kawamoto, S.; Tanaka, H.; Kishida, H.; Kitagawa, M.; Nakai, Y.; Abe, K.; Hirata, D. Administered chrysanthemum flower oil attenuates hyperuricemia: Mechanism of action as revealed by DNA microarray analysis. Biosci. Biotechnol. Biochem. 2014, 78, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Yang, C.W.; Liao, J.W.; Zhen, W.W.; Chu, F.H.; Chang, S.T. Essential oil from leaves of Cinnamomum osmophloeum acts as a xanthine oxidase inhibitor and reduces the serum uric acid levels in oxonate-induced mice. Phytomedicine 2008, 15, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Son, S.E.; Kim, S.H.; Lee, Y.M.; Kim, O.S.; Kim, D.S. Synergistic Uric Acid-Lowering Effects of the Combination of Chrysanthemum indicum Linne Flower and Cinnamomum cassia (L.) J. Persl Bark Extracts. Evid. Based Complement. Altern. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, C.P.; Hu, Q.H.; Lv, Y.Z.; Zhang, X.; Ouyang, Z.; Kong, L.D. The dual actions of Sanmiao wan as a hypouricemic agent: Down-regulation of hepatic XOD and renal mURAT1 in hyperuricemic mice. J. Ethnopharmacol. 2010, 128, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Ramallo, I.A.; Zacchino, S.A.; Furlan, R.L. A rapid TLC autographic method for the detection of xanthine oxidase inhibitors and superoxide scavenger. Phytochem. Anal. 2006, 17, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Unno, T.; Sugimoto, A.; Kakuda, T. Xanthine oxidase inhibitors from the leaves of Lagerstroemia speciosa (L.) Pers. J. Ethnopharmacol. 2004, 93, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Ichida, K.; Matsuo, H.; Takada, T.; Nakayama, A.; Murakami, K.; Shimizu, T.; Yamanashi, Y.; Kasuga, H.; Nakashima, H.; Nakamura, T.; et al. Decreased extra-renal urate excretion is a common cause of hyperuricemia. Nat. Commun. 2012, 3, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, A.; Endou, H. Roles of organic anion transporters (OATs) and a urate transporter (URAT1) in the pathophysiology of human disease. Clin. Exp. Nephrol. 2005, 9, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Preitner, F.; Bonny, O.; Laverriere, A.; Rotman, S.; Firsov, D.; Da Costa, A.; Metref, S.; Thorens, B. Glut9 is a major regulator of urate homeostasis and its genetic inactivation induces hyperuricosuria and urate nephropathy. Proc. Natl. Acad. Sci. USA 2009, 106, 15501–15506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Zhou, M.; Zhu, H.; Lu, G.; Zheng, D.; Li, H.; Hao, K. (E)-2-(4-bromophenyl)-1-(2,4-dihydroxyphenyl)ethanone oxime is a potential therapeutic agent for treatment of hyperuricemia through its dual inhibitory effects on XOD and URAT1. Biomed. Pharmacother. 2017, 86, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Gao, L.; Niu, Y.; Zhou, Y.; Lin, H.; Jiang, J.; Kong, X.; Liu, X.; Li, L. Mangiferin inhibits renal urate reabsorption by modulating urate transporters in experimental hyperuricemia. Biol. Pharm. Bull. 2015, 38, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jin, L.; Liu, J.; Wang, W.; Yu, H.; Li, J.; Chen, Q.; Wang, T. Effect and mechanism of dioscin from Dioscorea spongiosa on uric acid excretion in animal model of hyperuricemia. J. Ethnopharmacol. 2018, 214, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.W.; Lin, S.Y.; Kuo, C.C.; Huang, C.C. Serum uric acid and progression of kidney disease: A longitudinal analysis and mini-review. PLoS ONE 2017, 12, e0170393. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ruiz, F.; Hernandez-Baldizon, S.; Herrero-Beites, A.M.; Gonzalez-Gay, M.A.; Perez-Ruiz, F.; Hernandez-Baldizon, S.; Herrero-Beites, A.M.; Gonzalez-Gay, M.A. Risk factors associated with renal lithiasis during uricosuric treatment of hyperuricemia in patients with gout. Arthritis Care Res. 2010, 62, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Graham, G.G.; Williams, K.M.; Day, R.O. A benefit-risk assessment of benzbromarone in the treatment of gout. Was its withdrawal from the market in the best interest of patients? Drug Saf. 2008, 31, 643–665. [Google Scholar] [CrossRef] [PubMed]
- Felser, A.; Lindinger, P.W.; Schnell, D.; Kratschmar, D.V.; Odermatt, A.; Mies, S.; Jenö, P.; Krähenbühl, S. Hepatocellular toxicity of benzbromarone: Effects on mitochondrial function and structure. Toxicology 2008, 324, 136–146. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-S.; Kim, S.-H.; Yuk, H.J.; Kim, D.-S. DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl Bark Extracts, Improves Hyperuricemia through Inhibition of Xanthine Oxidase Activity and Increasing Urine Excretion. Nutrients 2018, 10, 1381. https://doi.org/10.3390/nu10101381
Lee Y-S, Kim S-H, Yuk HJ, Kim D-S. DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl Bark Extracts, Improves Hyperuricemia through Inhibition of Xanthine Oxidase Activity and Increasing Urine Excretion. Nutrients. 2018; 10(10):1381. https://doi.org/10.3390/nu10101381
Chicago/Turabian StyleLee, Young-Sil, Seung-Hyung Kim, Heung Joo Yuk, and Dong-Seon Kim. 2018. "DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl Bark Extracts, Improves Hyperuricemia through Inhibition of Xanthine Oxidase Activity and Increasing Urine Excretion" Nutrients 10, no. 10: 1381. https://doi.org/10.3390/nu10101381
APA StyleLee, Y.-S., Kim, S.-H., Yuk, H. J., & Kim, D.-S. (2018). DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl Bark Extracts, Improves Hyperuricemia through Inhibition of Xanthine Oxidase Activity and Increasing Urine Excretion. Nutrients, 10(10), 1381. https://doi.org/10.3390/nu10101381