
Spectral Reflectance of Palauan Reef-Building Coral with Different Symbionts in Response to Elevated Temperature

 ,
, 
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
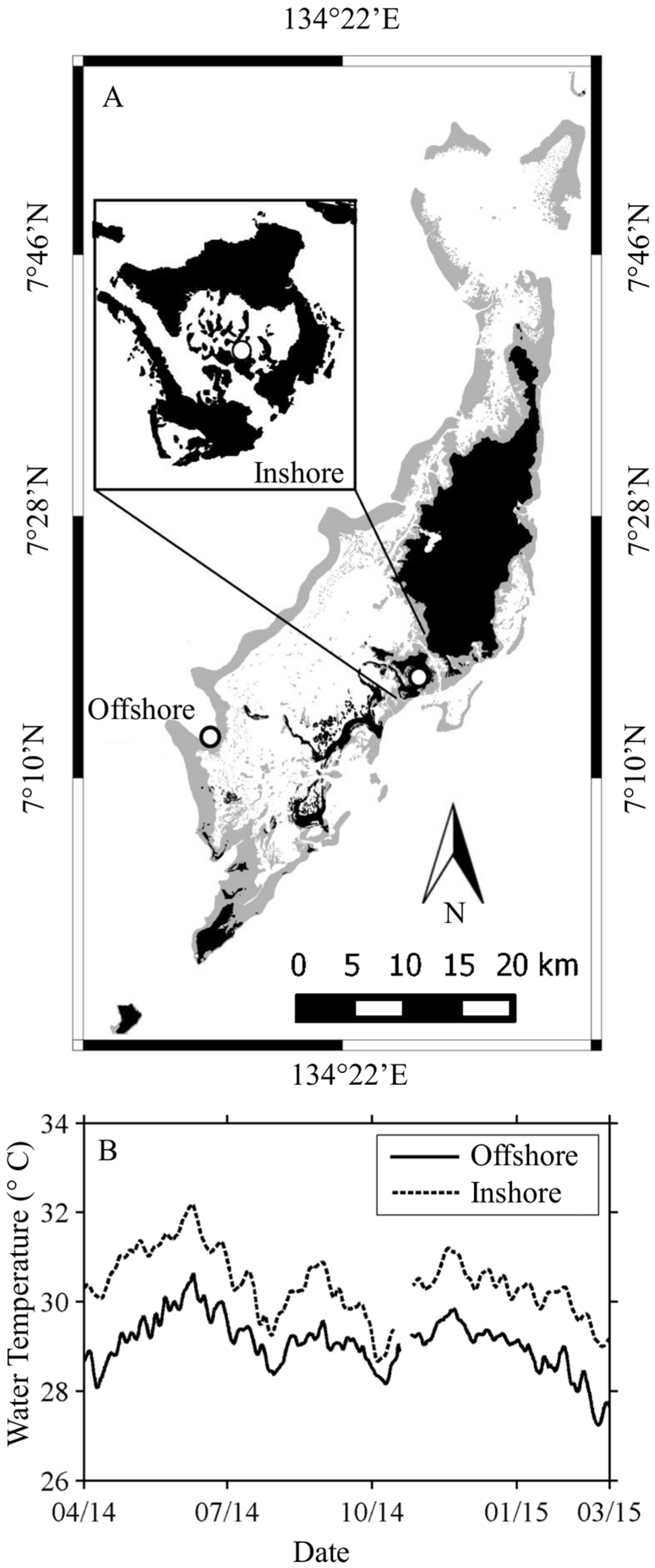
2.1. Study Sites
2.1.1. Nikko Bay—Inshore
2.1.2. Rebotel Reef—Offshore
2.2. Coral Collection and Maintenance
2.3. Experimental Setup
2.4. Symbiont Species and Density
2.5. Imaging and Reflectance Processing
2.6. Reflectance and Symbiont Density Comparison
2.7. Reflectance Analysis
3. Results
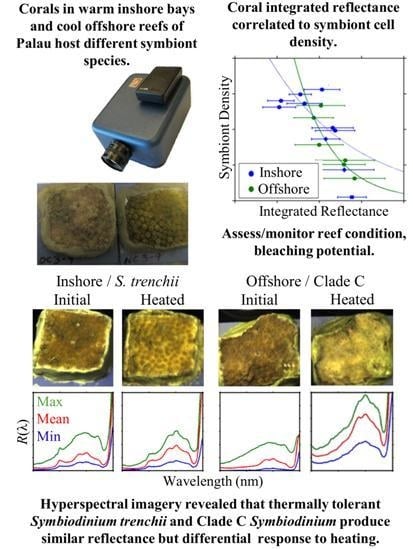
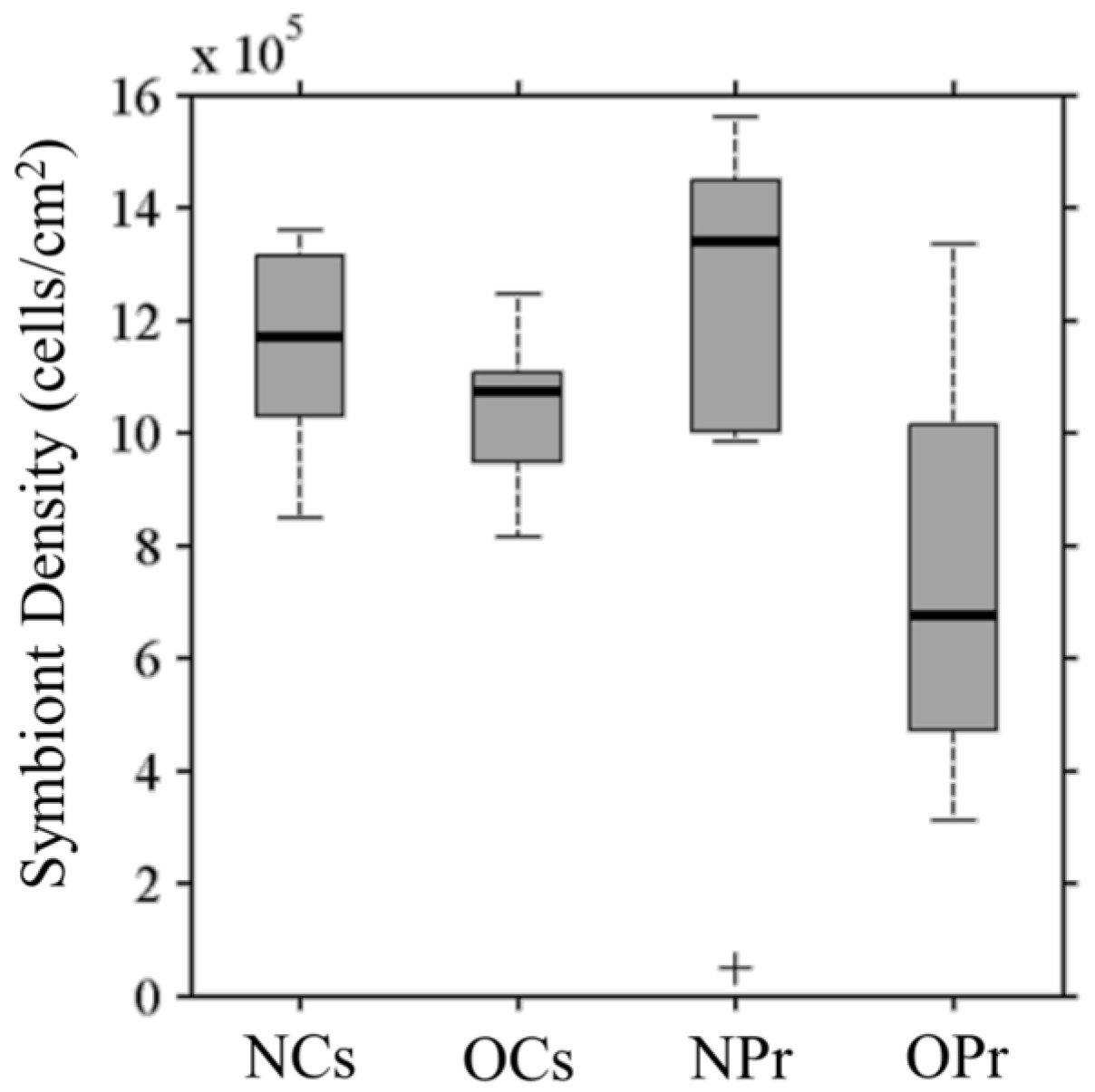
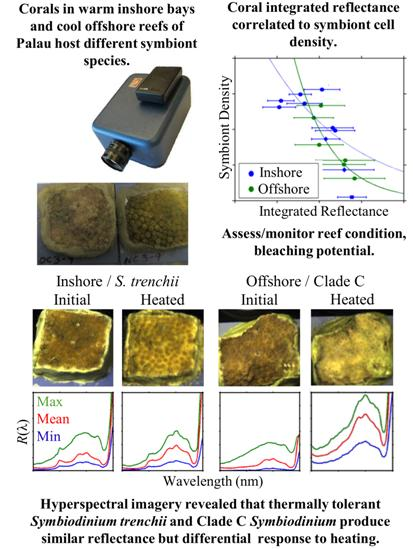
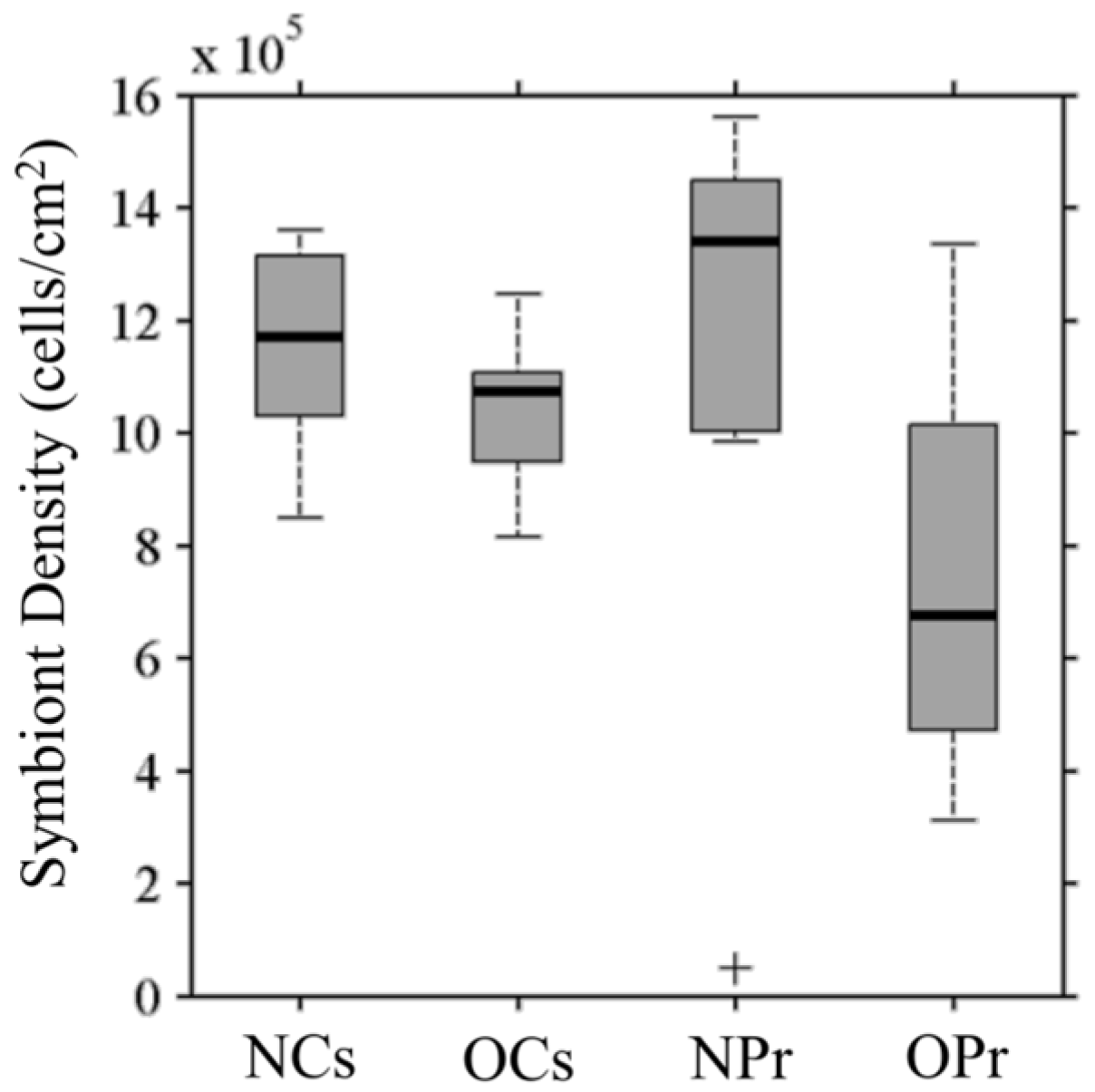
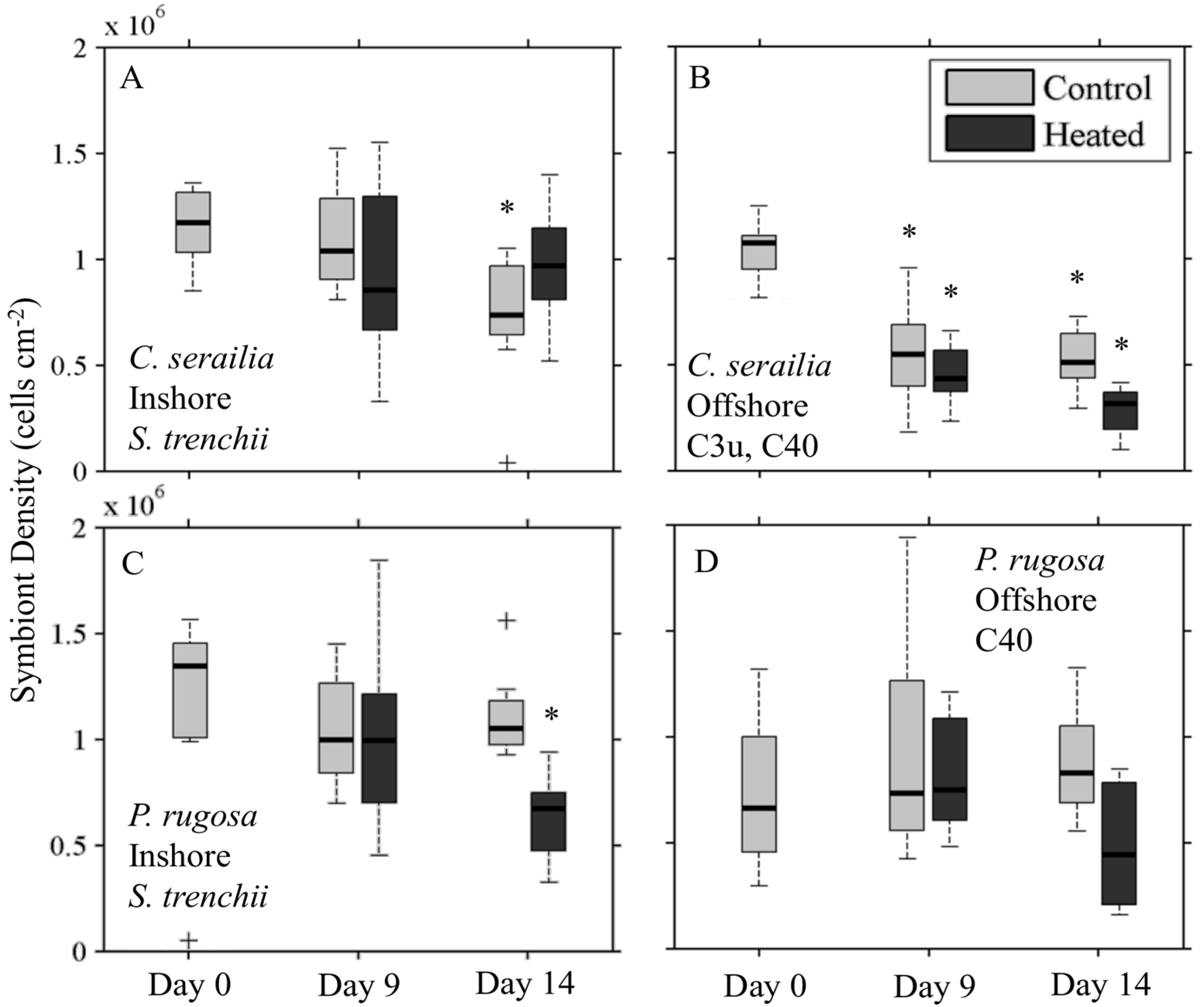
3.1. Symbiont Species and Density
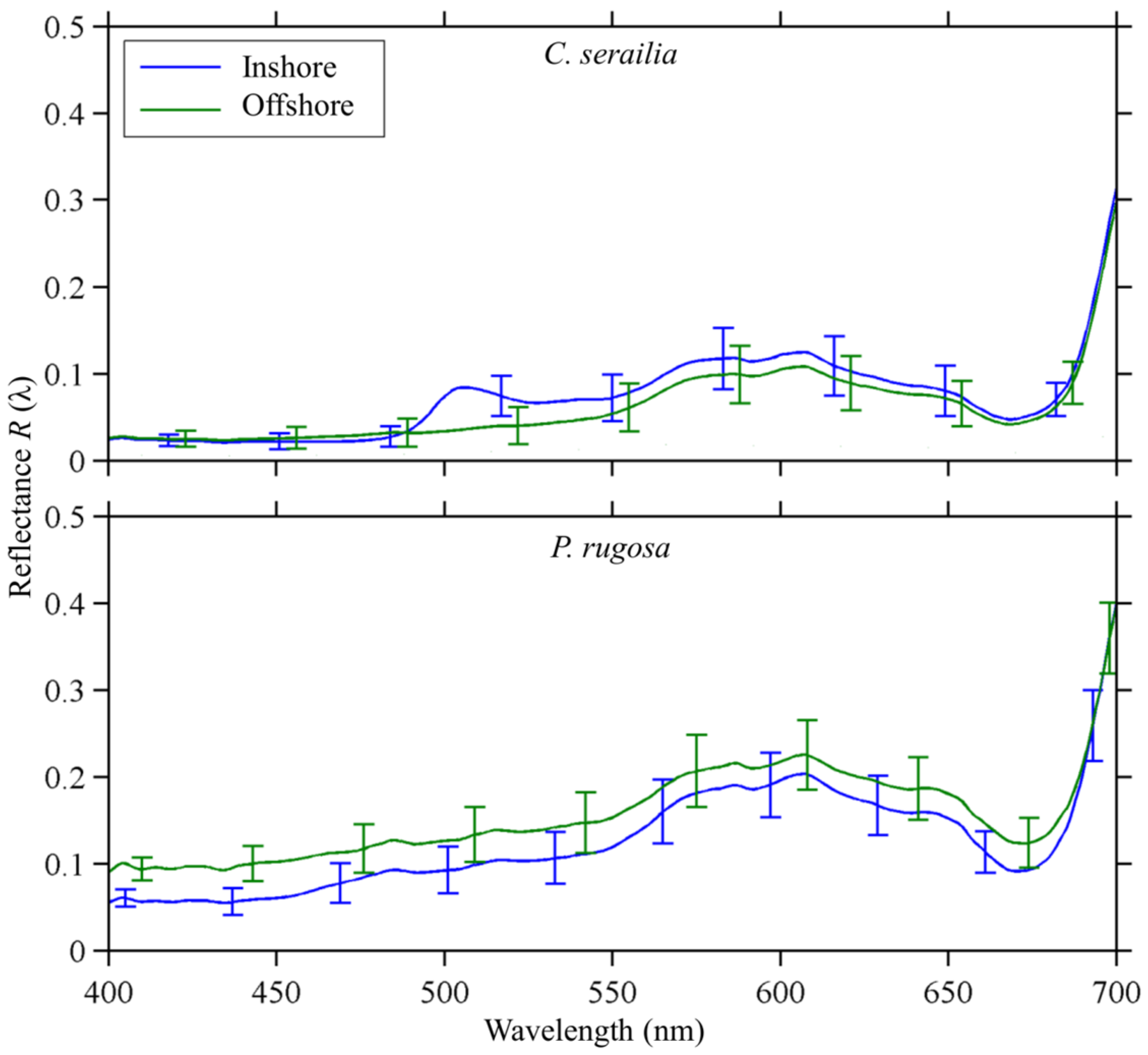
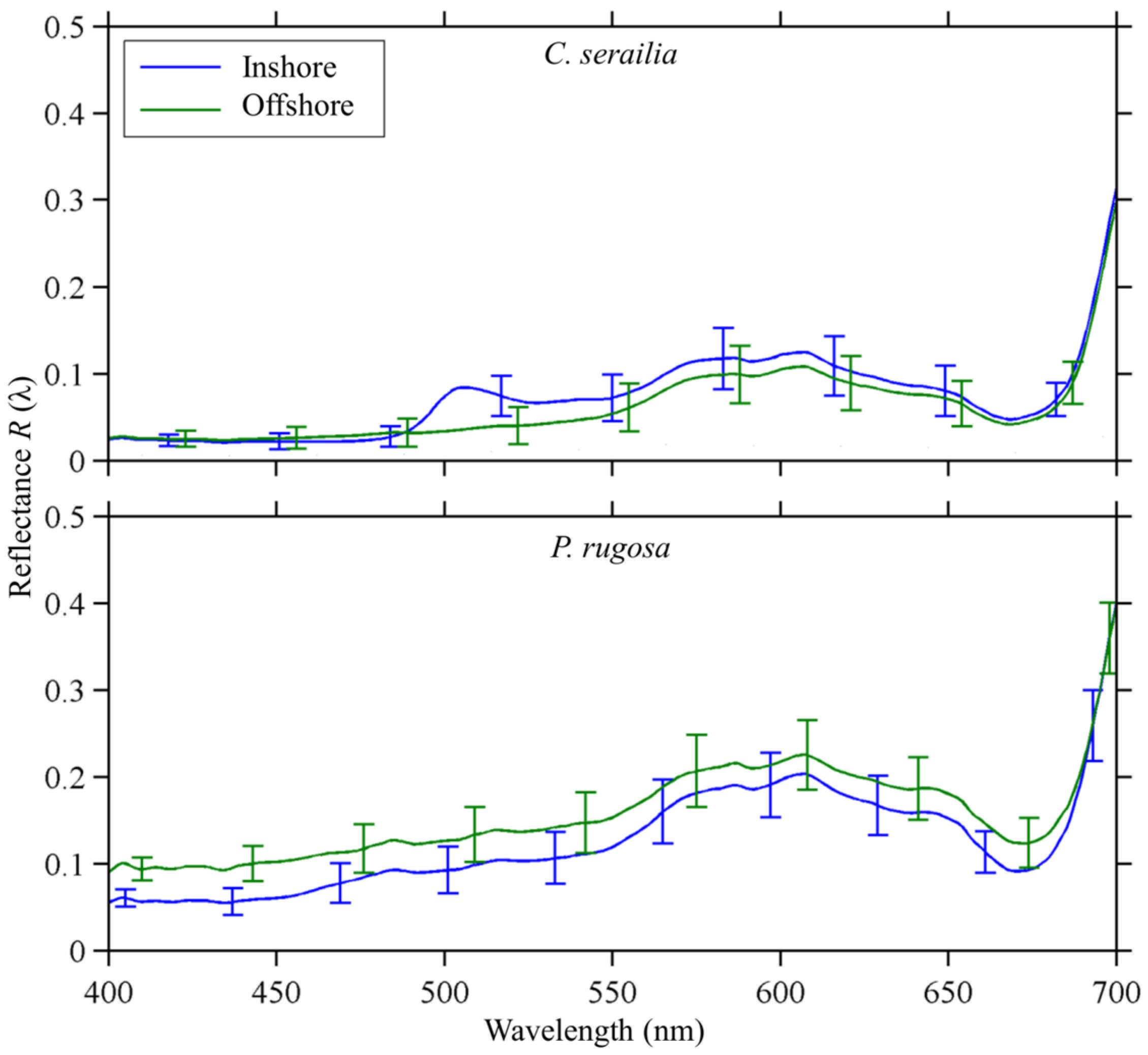
3.2. Spectral Reflectance
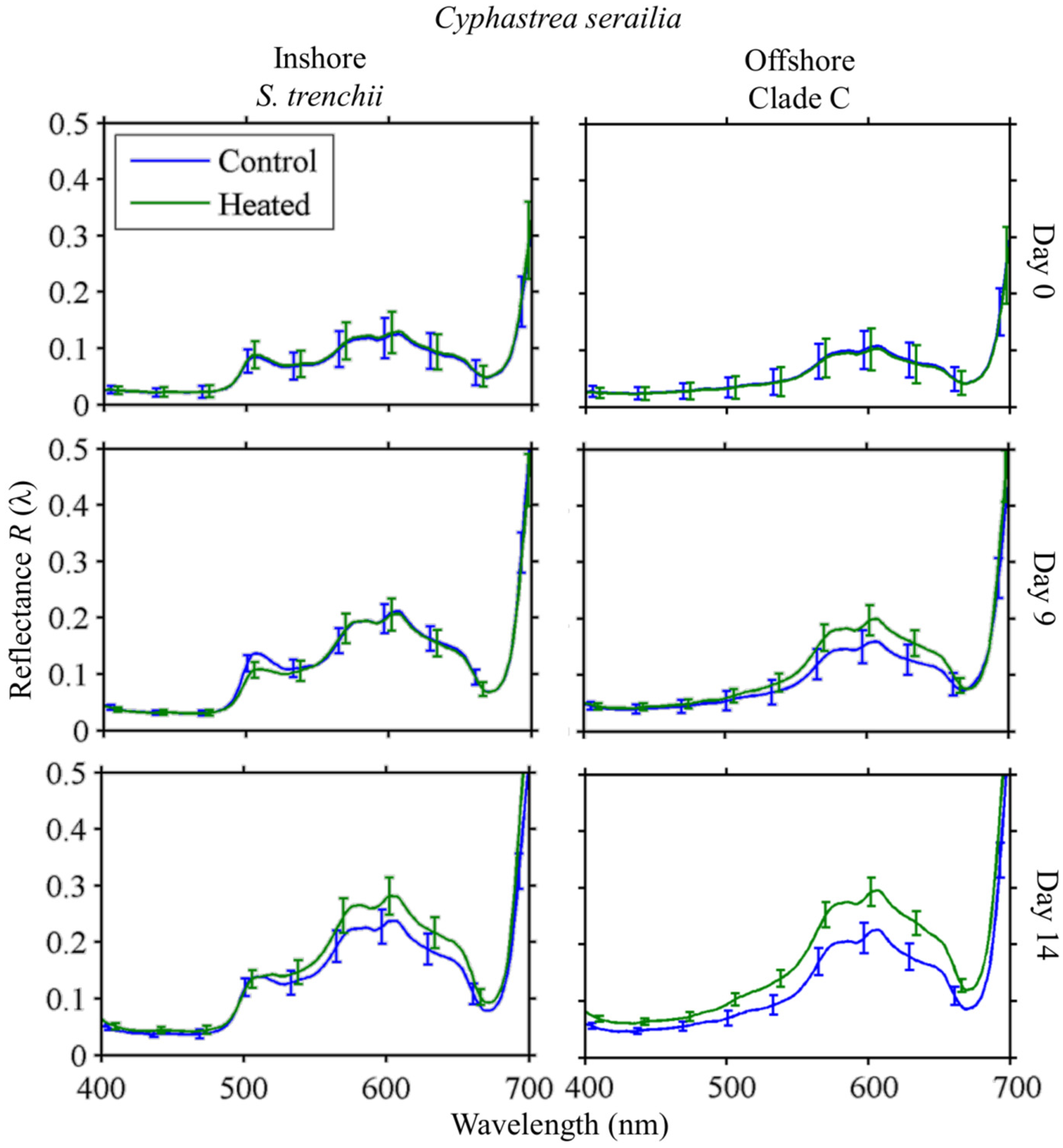
3.2.1. Cyphastrea serailia
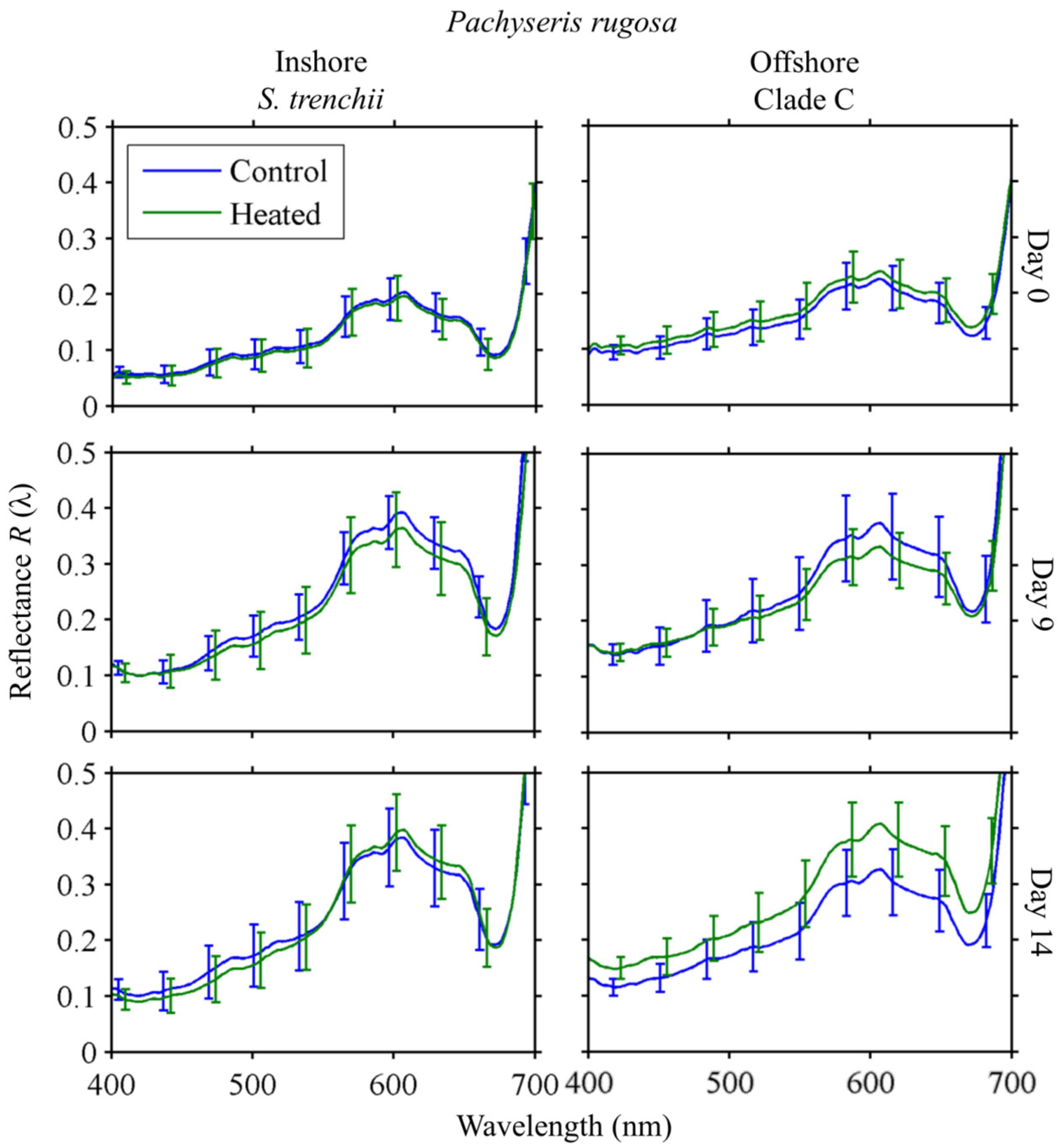
3.2.2. Pachyseris rugosa
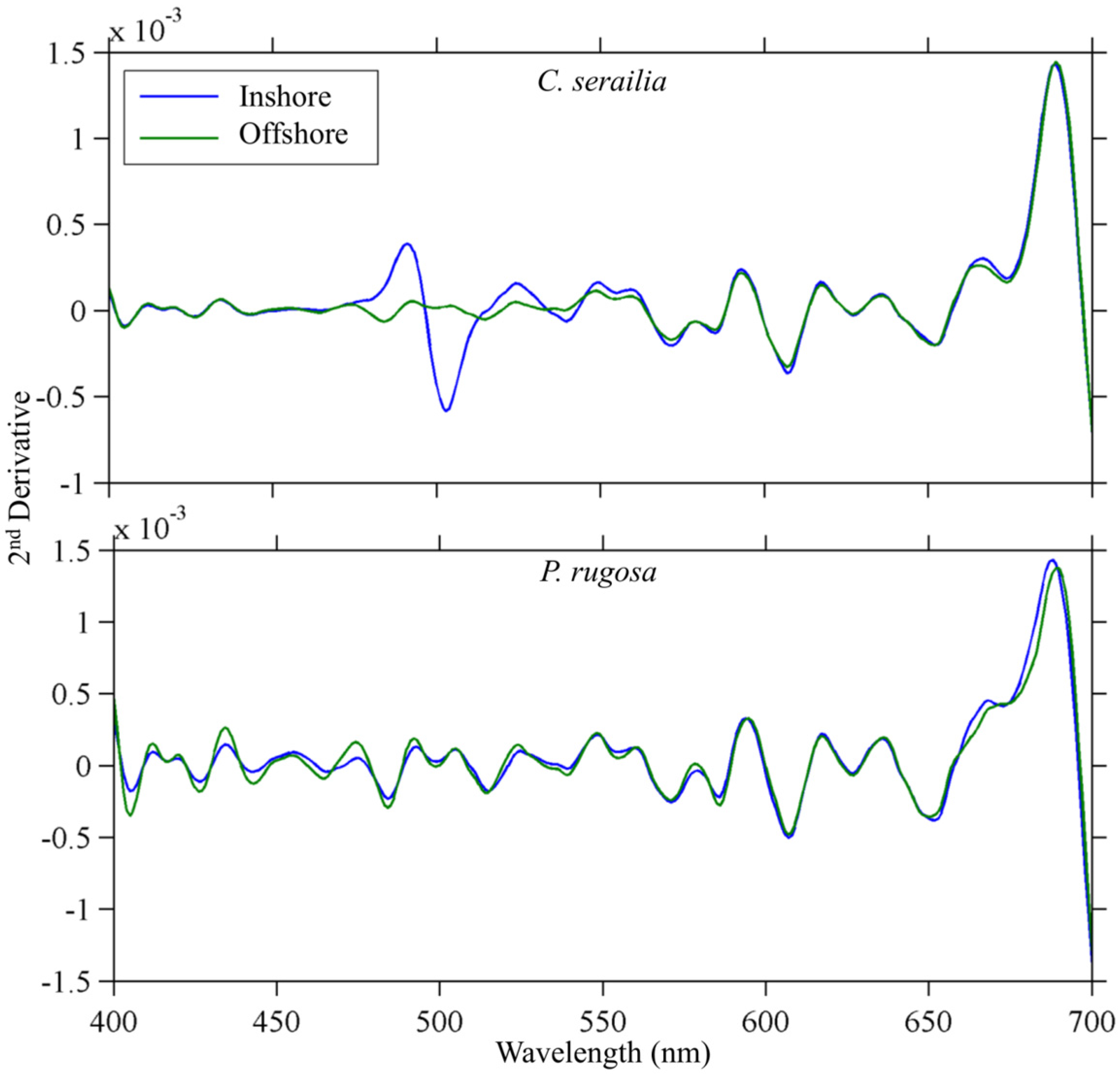
3.2.3. Derivative Analysis
3.2.4. Inshore vs. Offshore
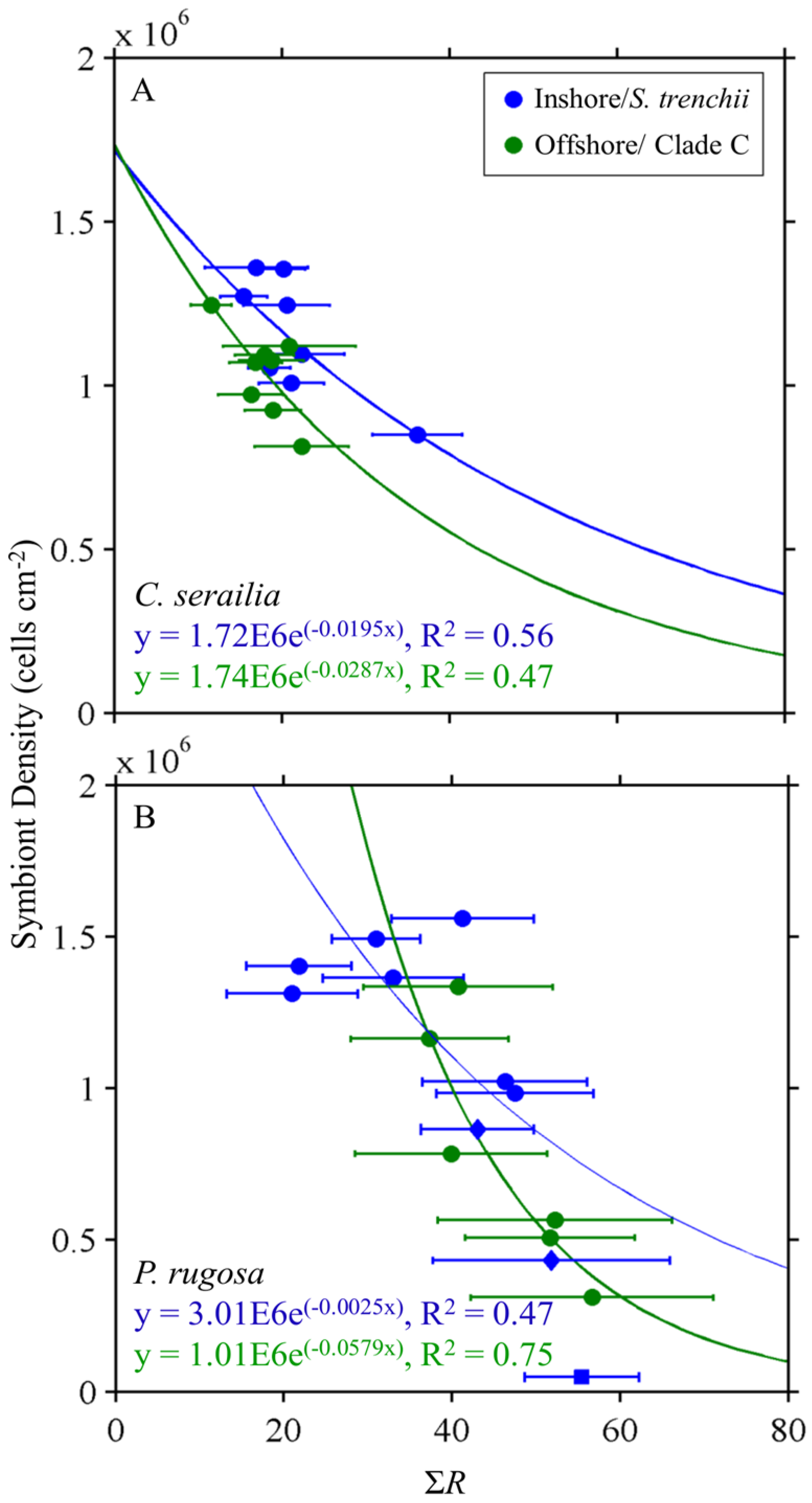
3.3. Reflectance and Symbiont Density Comparison
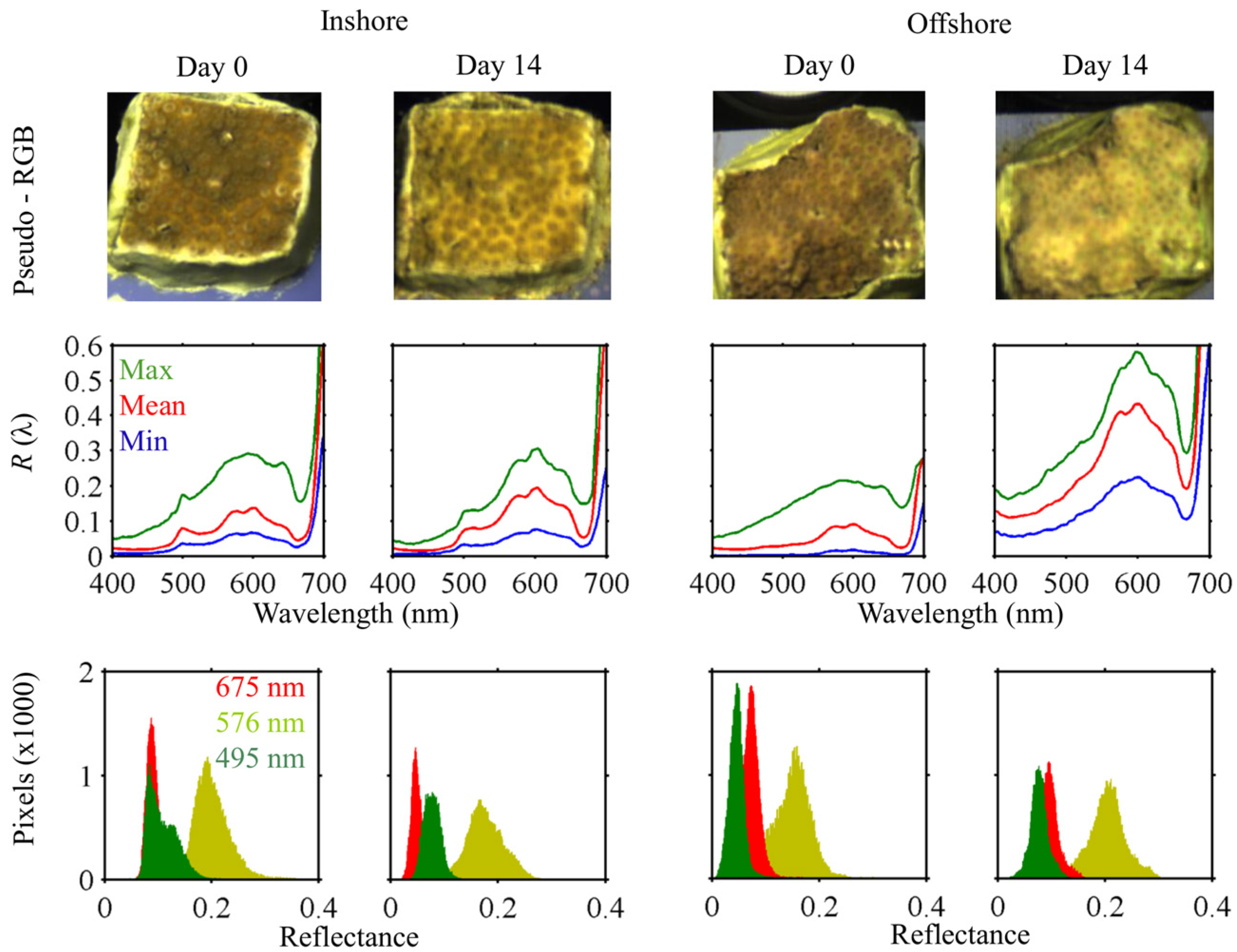
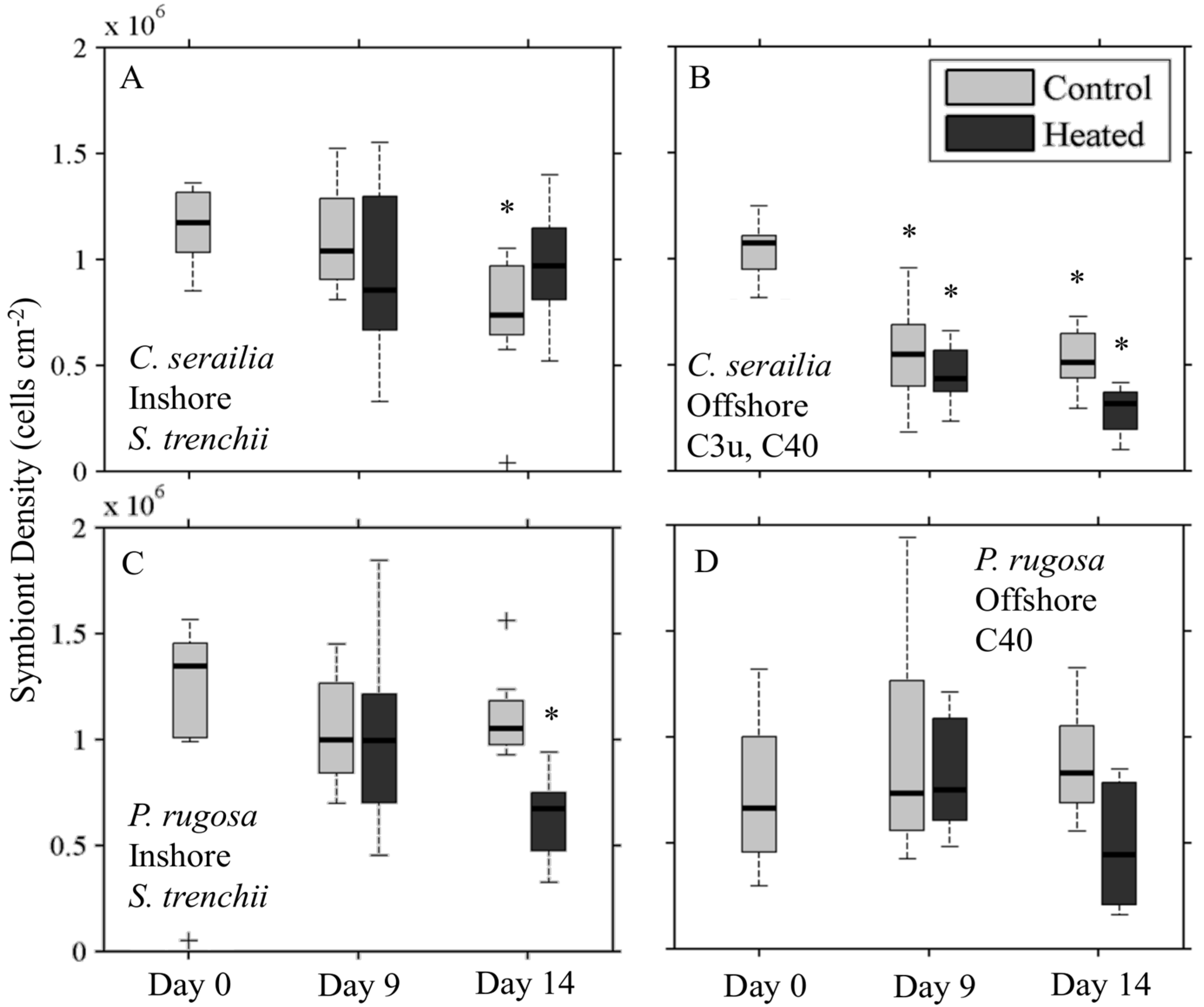
3.4. Response to Heating
3.4.1. Symbiont Density
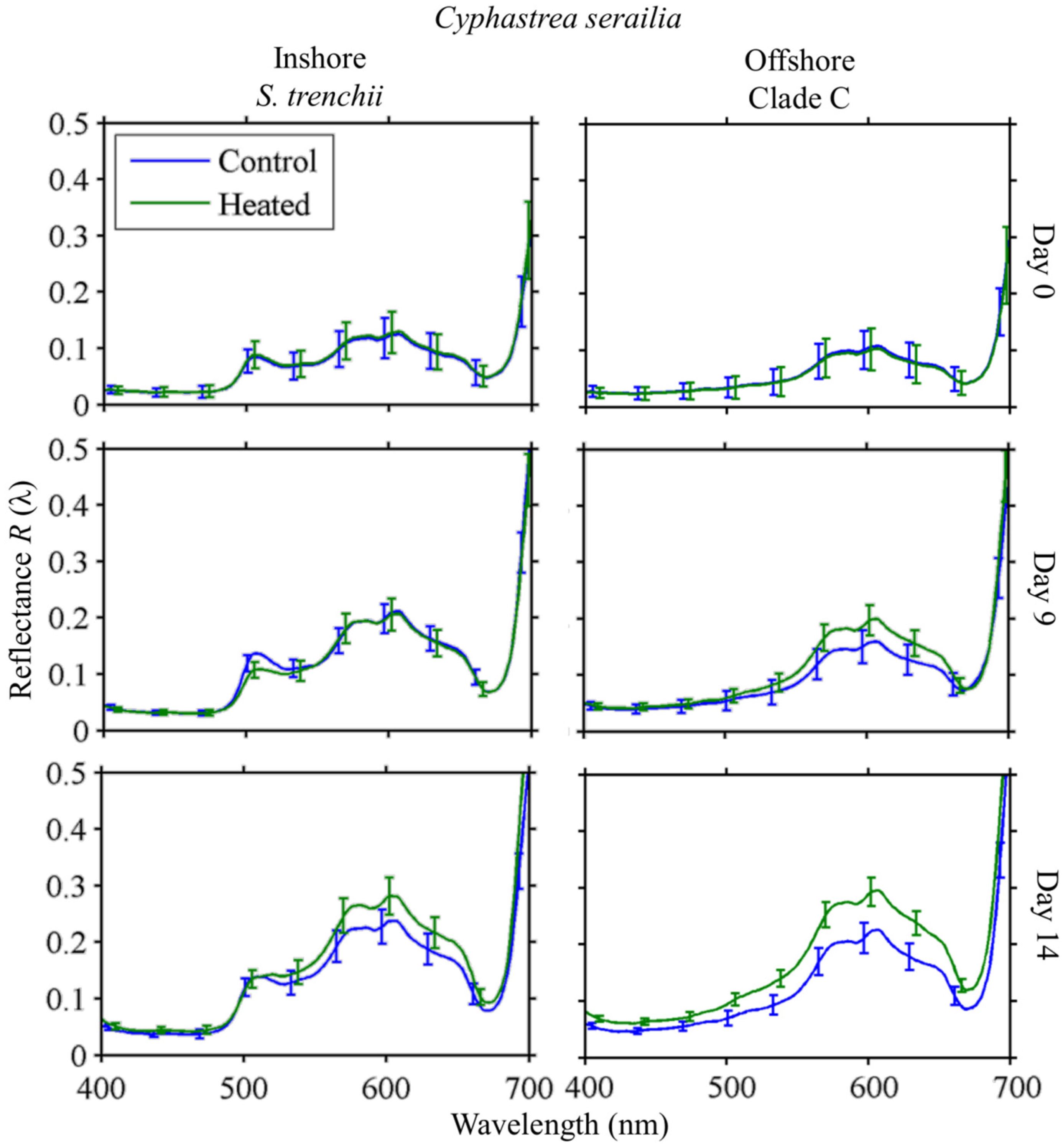
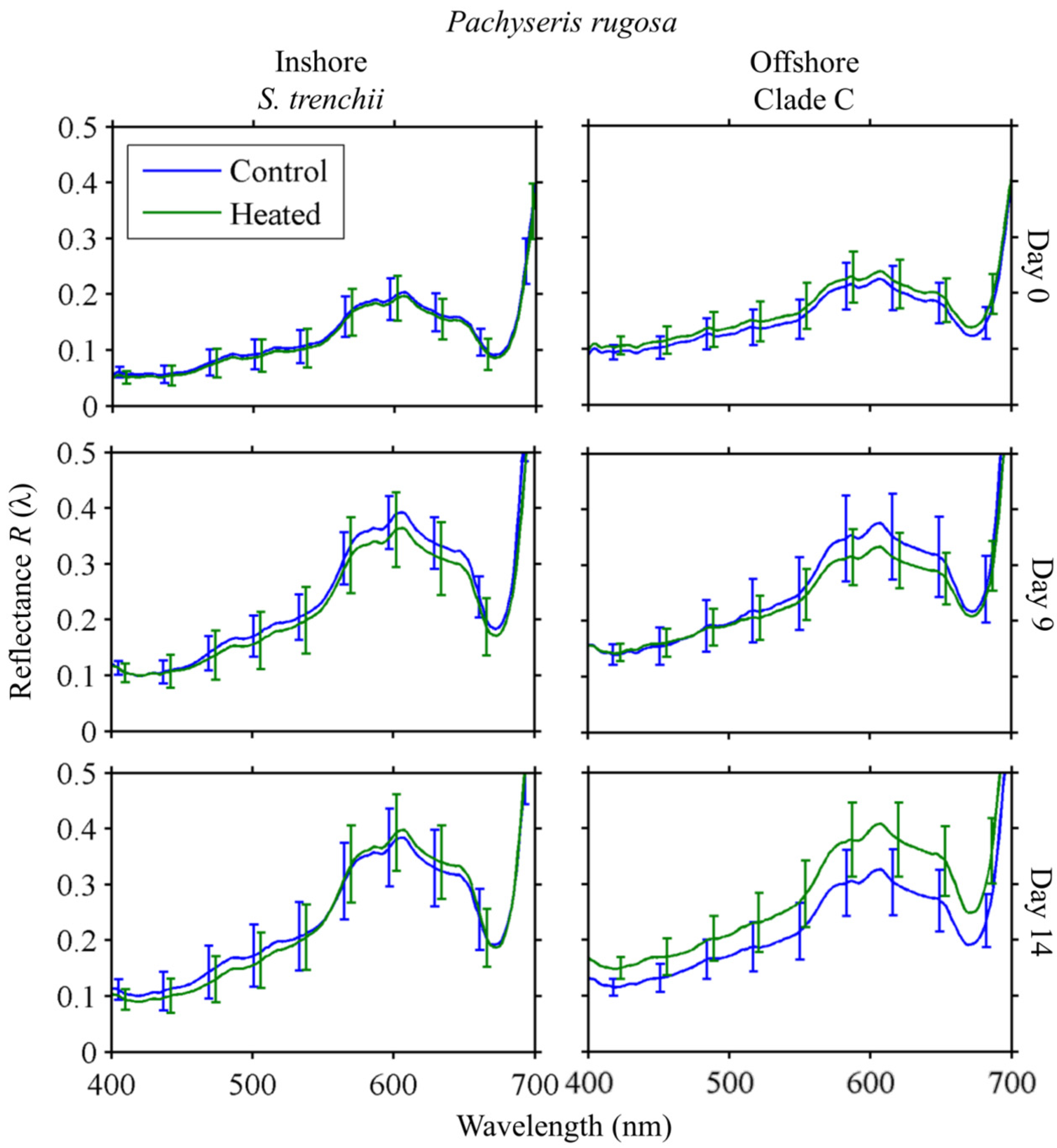
3.4.2. Reflectance
Cyphastrea serailia
Pachyseris rugosa
4. Discussion
4.1. Reflectance of Different Coral/Symbiont Systems
4.2. Spectral Variability with Symbiont Concentration
4.3. Spectral Response to Heating
5. Conclusions and Outlook for Remote Sensing
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DGGE | Denaturing gradient gel electrophoresis |
| GFP | Green fluorescent protein |
| HSI | Hyperspectral imaging |
| PCHIP | Piecewise cubic hermite interpolating polynomial |
| PICRC | Palau International Coral Reef Center |
| ROI | Region of interest |
References
- Andréfouét, S.; Payri, C.; Hochberg, E.J.; Hu, C.; Atkinson, M.J.; Muller-Karger, F.E. Use of in situ and airborne reflectance for scaling-up spectral discrimination of coral reef macroalgae from species to communities. Mar. Ecol. Prog. Ser. 2004, 283, 161–177. [Google Scholar] [CrossRef]
- Mumby, P.J.; Skirving, W.; Strong, A.E.; Hardy, J.T.; LeDrew, E.F.; Hochberg, E.J.; Stumpf, R.P.; David, L.T. Remote sensing of coral reefs and their physical environment. Mar. Pollut. Bull. 2004, 48, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J.; Ustin, S.L. Classification of benthic composition in a coral reef environment using spectral unmixing. J. Appl. Remote Sens. 2007, 1. [Google Scholar] [CrossRef]
- Scopélitis, J.; Andréfouët, S.; Phinn, S.; Chabanet, P.; Naim, O.; Tourrand, C.; Done, T. Changes of coral communities over 35 years: Integrating in situ and remote-sensing data on Saint-Leu Reef (la Réunion, Indian Ocean). Estuar. Coast. Shelf Sci. 2009, 84, 342–352. [Google Scholar] [CrossRef]
- Hochberg, E. Spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. Remote Sens. Environ. 2003, 85, 159–173. [Google Scholar] [CrossRef]
- Freudenthal, H.D. Symbiodinium gen. nov. and Symbiodinium microadriaticum sp. nov., a Zooxanthella: Taxonomy, Life Cycle, and Morphology. J. Protozool. 1962, 9, 45–52. [Google Scholar] [CrossRef]
- LaJeunesse, T. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs. Mar. Biol. 2002, 141, 387–400. [Google Scholar]
- LaJeunesse, T.C.; Pettay, D.T.; Sampayo, E.M.; Phongsuwan, N.; Brown, B.; Obura, D.O.; Hoegh-Guldberg, O.; Fitt, W.K. Long-standing environmental conditions, geographic isolation and host-symbiont specificity influence the relative ecological dominance and genetic diversification of coral endosymbionts in the genus Symbiodinium. J. Biogeogr. 2010, 37, 785–800. [Google Scholar] [CrossRef]
- Thornhill, D.J.; Lewis, A.M.; Wham, D.C.; LaJeunesse, T.C. Host-specialist lineages dominate the adaptive radidation of reef coral endosymbionts. Evolution 2014, 68, 352–367. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Loh, W.K.; Van Woesik, R.; Hoegh-Guldberg, O.; Schmidt, G.W.; Fitt, W.K. Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Limnol. Oceanogr. 2003, 48, 2046–2054. [Google Scholar] [CrossRef]
- Tonk, L.; Sampayo, E.M.; LaJeunesse, T.C.; Schrameyer, V.; Hoegh-Guldberg, O. Symbiodinium (Dinophyceae) diversity in reef-invertebrates along an offshore to inshore reef gradient near Lizard Island, Great Barrier Reef. J. Phycol. 2014, 50, 552–563. [Google Scholar] [CrossRef]
- Torres-Pérez, J.L.; Guild, L.S.; Armstrong, R.A. Hyperspectral distinction of two Caribbean shallow-water corals based on their pigments and corresponding reflectance. Remote Sens. 2012, 4, 3813–3832. [Google Scholar] [CrossRef]
- Kemp, D.W.; Hernandez-Pech, X.; Iglesias-Prieto, R.; Fitt, W.K.; Schmidt, G.W. Community dynamics and physiology of Symbiodinium spp. before, during, and after a coral bleaching event. Limnol. Oceanogr. 2014, 59, 788–797. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Smith, R.T.; Finney, J.; Oxenford, H. Outbreak and persistence of opportunistic symbiotic dinoflagellates during the 2005 Caribbean mass coral “bleaching”event. Proc. R. Soc. B Biol. Sci. 2009, 276, 4139–4148. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Wham, D.C.; Pettay, D.T.; Parkinson, J.E.; Keshavmurthy, S.; Chen, C.A. Ecologically differentiated stress-tolerant endosymbionts in the dinoflagellate genus Symbiodinium (Dinophyceae) Clade D are different species. Phycologia 2014, 53, 305–319. [Google Scholar] [CrossRef]
- Pettay, D.T.; Wham, D.C.; Smith, R.T.; Iglesias-Prieto, R.; LaJeunesse, T.C. Microbial invasion of the Caribbean by an Indo-Pacific coral zooxanthella. Proc. Natl. Acad. Sci. USA 2015, 112, 7513–7518. [Google Scholar] [CrossRef] [PubMed]
- Ragni, M.; Ruth, A.L.; Hennige, S.J.; Suggett, D.J.; Warner, M.E.; Geider, R.J. PSII photoinhibition and photorepair in Symbiodinium (Pyrrhophyta) differs between thermally tolerant and sensitive phylotypes. Mar. Ecol. Prog. Ser. 2010, 406, 57–70. [Google Scholar] [CrossRef]
- Robison, J.D.; Warner, M.E. Differential impacts of photoacclimation and thermal stress on the photobiology of four different phylotypes of Symbiodinium (Pyrrhophyta). J. Phycol. 2006, 42, 568–579. [Google Scholar] [CrossRef]
- Steinke, M.; Brading, P.; Kerrison, P.; Warner, M.E.; Suggett, D.J. Concentrations of dimethylsulfoniopropionate and dimethyl sulfide are strain-specific in symbiotic dinoflagellates (Symbiodinium sp., Dinophyceae). J. Phycol. 2011, 47, 775–783. [Google Scholar] [CrossRef]
- Berkelmans, R.; van Oppen, M.J. The role of zooxanthellae in the thermal tolerance of corals: A “nugget of hope” for coral reefs in an era of climate change. Proc. R. Soc. B Biol. Sci. 2006, 273, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Grottoli, A.G.; Warner, M.E.; Levas, S.J.; Aschaffenburg, M.D.; Schoepf, V.; McGinley, M.; Baumann, J.; Matsui, Y. The cumulative impact of annual coral bleaching can turn some coral species winners into losers. Glob. Chang. Biol. 2014, 20, 3823–3833. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, R.N.; Cunning, R.; Baker, A.C. Change in algal symbiont communities after bleaching, not prior heat exposure, increases heat tolerance of reef corals. Glob. Change Biol. 2015, 21, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Enríquez, S.; Méndez, E.R.; Prieto, R.I. Multiple scattering on coral skeletons enhances light absorption by symbiotic algae. Limnol. Oceanogr. 2005, 50, 1025–1032. [Google Scholar] [CrossRef]
- Hedley, J.D.; Mumby, P.J. Biological and remote sensing perspectives of pigmentation in coral reef organisms. Adv. Mar. Biol. 2002, 43, 277–317. [Google Scholar] [PubMed]
- Holden, H.; LeDrew, E. Spectral discrimination of healthy and non-healthy corals based on cluster analysis, principal components analysis, and derivative spectroscopy. Remote Sens. Environ. 1998, 65, 217–224. [Google Scholar] [CrossRef]
- Joyce, K.E.; Phinn, S.R. Hyperspectral analysis of chlorophyll content and photosynthetic capacity of coral reef substrates. Limnol. Oceanogr. 2003, 48, 489–496. [Google Scholar] [CrossRef]
- Ralph, P.J.; Larkum, A.W.D.; Kühl, M. Photobiology of endolithic microorganisms in living coral skeletons: 1. Pigmentation, spectral reflectance and variable chlorophyll fluorescence analysis of endoliths in the massive corals Cyphastrea serailia, Porites lutea and Goniastrea australensis. Mar. Biol. 2007, 152, 395–404. [Google Scholar] [CrossRef]
- Rodríguez-Román, A.; Hernández-Pech, X.; E Thome, P.; Enríquez, S.; Iglesias-Prieto, R. Photosynthesis and light utilization in the Caribbean coral Montastraea faveolata recovering from a bleaching event. Limnol. Oceanogr. 2006, 51, 2702–2710. [Google Scholar] [CrossRef]
- Clark, C.D.; Mumby, P.J.; Chisholm, J.R.M.; Jaubert, J.; Andréfouét, S. Spectral discrimination of coral mortality states following a severe bleaching event. Int. J. Remote Sens. 2000, 21, 2321–2327. [Google Scholar] [CrossRef]
- Hochberg, E.J.; Atkinson, M.J. Spectral discrimination of coral reef benthic communities. Coral Reefs 2000, 19, 164–171. [Google Scholar] [CrossRef]
- Myers, M.R.; Hardy, J.T.; Mazel, C.H.; Dustan, P. Optical spectra and pigmentation of Caribbean reef corals and macroalgae. Coral Reefs 1999, 18, 179–186. [Google Scholar] [CrossRef]
- Caras, T.; Karnieli, A. Ground-level spectroscopy analyses and classification of coral reefs using a hyperspectral camera. Coral Reefs 2013, 32, 825–834. [Google Scholar] [CrossRef]
- Caras, T.; Karnieli, A. Ground-level classification of a coral reef using a hyperspectral camera. Remote Sens. 2015, 7, 7521–7544. [Google Scholar] [CrossRef]
- Leiper, I.; Phinn, S.; Dekker, A.G. Spectral reflectance of coral reef benthos and substrate assemblages on Heron Reef, Australia. Int. J. Remote Sens. 2012, 33, 3946–3965. [Google Scholar] [CrossRef]
- Lucas, M.; Goodman, J. Linking coral reef remote sensing and field ecology: It’s a matter of scale. J. Mar. Sci. Eng. 2014, 3, 1–20. [Google Scholar] [CrossRef]
- Anderson, D.A.; Armstrong, R.A.; Weil, E. Hyperspectral sensing of disease stress in the Caribbean reef-building coral, Orbicella faveolata—Perspectives for the field of coral disease monitoring. PLoS ONE 2013, 8, e81478. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, E.J.; Apprill, A.M.; Atkinson, M.J.; Bidigare, R.R. Bio-optical modeling of photosynthetic pigments in corals. Coral Reefs 2006, 25, 99–109. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Chlus, A.; Russell, B. Hyperspectral discrimination of floating mats of seagrass wrack and the macroalgae sargassum in coastal waters of Greater Florida Bay using airborne remote sensing. Remote Sens. Environ. 2015, 167, 247–258. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Steiner, U.; Hillnhütter, C.; Dehne, H.W.; Oerke, E.C. Hyperspectral imaging for small-scale analysis of symptoms caused by different sugar beet diseases. Plant Methods 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Nansen, C.; Ribeiro, L.P.; Dadour, I.; Roberts, J.D. Detection of temporal changes in insect body reflectance in response to killing agents. PLoS ONE 2015, 10, e0124866. [Google Scholar] [CrossRef] [PubMed]
- Rascher, U.; Nichol, C.J.; Small, C.; Hendricks, L. Monitoring spatio-temporal dynamics of photosynthesis with a portable hyperspectral imaging system. Photogramm. Eng. Remote Sens. 2007, 73, 45–56. [Google Scholar] [CrossRef]
- Römer, C.; Wahabzada, M.; Ballvora, A.; Pinto, F.; Rossini, M.; Panigada, C.; Behmann, J.; Léon, J.; Thurau, C.; Bauckhage, C.; et al. Early drought stress detection in cereals: Simplex volume maximisation for hyperspectral image analysis. Funct. Plant Biol. 2012, 39, 878–890. [Google Scholar] [CrossRef]
- Russell, B.J.; Dierssen, H.M. Use of hyperspectral imagery to assess cryptic color matching in sargassum associated crabs. PLoS ONE 2015, 10, e0136260. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, L.; Larkum, A.W.; Norman, A.; Qvortrup, K.; Chen, M.; Ralph, P.; Sørensen, S.J.; Trampe, E.; Kühl, M. Endolithic chlorophyll d-containing phototrophs. ISME J. 2011, 5, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Kühl, M.; Polerecky, L. Functional and structural imaging of phototrophic microbial communities and symbioses. Aquat. Microb. Ecol. 2008, 53, 99–118. [Google Scholar] [CrossRef]
- Mehrubeoglu, M.; Smith, D.K.; Smith, S.W.; Strychar, K.B.; McLauchlan, L. Investigating coral hyperspectral properties across coral species and coral state using hyperspectral imaging. Proc. SPIE 2013. [Google Scholar] [CrossRef]
- Johnsen, G.; Volent, Z.; Dierssen, H.; Pettersen, R.; Ardelan, M.V.; Søreide, F.; Fearns, P.; Ludvigsen, M.; Moline, M. Underwater hyperspectral imagery to create biogeochemical maps of seafloor properties. In Subsea Optics and Imaging; Elsevier: Amsterdam, The Netherlands, 2013; pp. 508–540. [Google Scholar]
- Dierssen, H.M. Overview of hyperspectral remote sensing for mapping marine benthic habitats from airborne and underwater sensors. Proc. SPIE 2013. [Google Scholar] [CrossRef]
- Chennu, A.; Färber, P.; Volkenborn, N.; Al-Najjar, M.A.A.; Janssen, F.; de Beer, D.; Polerecky, L. Hyperspectral imaging of the microscale distribution and dynamics of microphytobenthos in intertidal sediments: Hyperspectral imaging of MPB biofilms. Limnol. Oceanogr. Methods 2013, 11, 511–528. [Google Scholar] [CrossRef]
- Golbuu, Y.; Victor, S.; Penland, L.; Idip, D.; Emaurois, C.; Okaji, K.; Yukihira, H.; Iwase, A.; van Woesik, R. Palau’s coral reefs show differential habitat recovery following the 1998-bleaching event. Coral Reefs 2007, 26, 319–332. [Google Scholar] [CrossRef]
- Van Woesik, R.; Houk, P.; Isechal, A.L.; Idechong, J.W.; Victor, S.; Golbuu, Y. Climate-change refugia in the sheltered bays of Palau: Analogs of future reefs. Ecol. Evol. 2012, 2, 2474–2484. [Google Scholar] [CrossRef] [PubMed]
- Golbuu, Y.; Bauman, A.; Kuartei, J.; Victor, S. The state of coral reef ecosystems of Palau. In The State of Coral Reef Ecosystems of the United States and Pacific Freely Associated States; National Oceanic and Atmospheric Administration, National Ocean Service, National Centers for Coastal Ocean Science: Silver Spring, MD, USA, 2005; pp. 488–507. [Google Scholar]
- Barkley, H.C.; Cohen, A.L.; Golbuu, Y.; Starczak, V.R.; DeCarlo, T.M.; Shamberger, K.E.F. Changes in coral reef communities across a natural gradient in seawater pH. Sci. Adv. 2015. [Google Scholar] [CrossRef] [PubMed]
- Szmant, A.M.; Gassman, N.J. The effects of prolonged “bleaching” on the tissue biomass and reproduction of the reef coral Montastrea annularis. Coral Reefs 1990, 8, 217–224. [Google Scholar] [CrossRef]
- Fitt, W.K.; McFarland, F.K.; Warner, M.E.; Chilcoat, G.C. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnol. Oceanogr. 2000, 45, 677–685. [Google Scholar] [CrossRef]
- Suggett, D.J.; Prášil, O.; Borowitzka, M.A. Chlorophyll a Fluorescence in Aquatic Sciences: Methods and Applications; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Marsh, J.A. Primary productivity of reef-building calcareous red algae. Ecology 1970, 51, 255–263. [Google Scholar] [CrossRef]
- Zhang, H.; Voss, K.J. Bi-directional reflectance of dry and submerged Labsphere Spectralon plaque. Appl. Opt. 2006, 45, 7924–7927. [Google Scholar]
- Hochberg, E.J.; Atkinson, M.J.; Apprill, A.; Andréfouét, S. Spectral reflectance of coral. Coral Reefs 2004, 23, 84–95. [Google Scholar] [CrossRef]
- Torres-Pérez, J.L.; Guild, L.S.; Armstrong, R.A.; Corredor, J.; Zuluaga-Montero, A.; Polanco, R. Relative pigment composition and remote sensing reflectance of Caribbean shallow-water corals. PLoS ONE 2015, 10, e0143709. [Google Scholar] [CrossRef] [PubMed]
- Mazel, C.H. Coral fluorescence characteristics: Excitation/emmission spectra, fluorescence efficiences, and contribution to apparent reflectance. Proc. SPIE 1997. [Google Scholar] [CrossRef]
- Mazel, C.H.; Fuchs, E. Contribution of fluorescence to the spectral signature and perceived color of corals. Limnol. Oceanogr. 2003, 48, 390–401. [Google Scholar] [CrossRef]
- Salih, A.; Larkum, A.W.; Cox, G.; Kühl, M.; Hoegh-Guldberg, O. Fluorescent pigments in corals are photoprotective. Nature 2000, 408, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Bou-Abdallah, F.; Chasteen, N.D.; Lesser, M.P. Quenching of superoxide radicals by green fluorescent protein. Biochim. Biophys. Acta 2006, 1760, 1690–1695. [Google Scholar] [CrossRef] [PubMed]
- Mazel, C.H.; Strand, M.P.; Lesser, M.P.; Crosby, M.P.; Coles, B.; Nevis, A.J. High-resolution determination of coral reef bottom cover from multispectral fluorescence laser line scan imagery. Limnol. Oceanogr. 2003, 48, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Iglesias-Prieto, R.; Trench, R.K. Acclimation and adaptation to irradiance in symbiotic dinoflagellates. I. Responses of the photosynthetic unit to changes in photon flux density. Mar. Ecol. Prog. Ser. 1994, 113, 163–175. [Google Scholar] [CrossRef]
- Iglesias-Prieto, R.; Beltran, V.H.; LaJeunesse, T.C.; Reyes-Bonilla, H.; Thome, P.E. Different algal symbionts explain the vertical distribution of dominant reef corals in the eastern Pacific. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; van Woesik, R.; Yamasaki, H. Photoinhibition of photosynthesis is reduced by water flow in the reef-building coral Acropora digitifera. Mar. Ecol. Prog. Ser. 2005, 301, 109–118. [Google Scholar] [CrossRef]
- Takahashi, S.; Nakamura, T.; Sakamizu, M.; van Woesik, R.; Yamasaki, H. Repair machinery of symbiotic photosynthesis as the primary target of heat stress for reef-building corals. Plant Cell Physiol. 2004, 45, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Warner, M.E.; Fitt, W.K.; Schmidt, G.W. The effects of elevated temperature on the photosynthetic efficiency of zooxanthellae in hospite from four different species of reef coral: A novel approach. Plant Cell Environ. 1996, 19, 291–299. [Google Scholar] [CrossRef]
- Reichstetter, M.; Fearns, P.; Weeks, S.; McKinna, L.; Roelfsema, C.; Furnas, M. Bottom reflectance in ocean color satellite remote sensing for coral reef environments. Remote Sens. 2015, 7, 16756–16777. [Google Scholar] [CrossRef]
- Garcia, R.; Hedley, J.; Tin, H.; Fearns, P. A method to analyze the potential of optical remote sensing for benthic habitat mapping. Remote Sens. 2015, 7, 13157–13189. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | ||||
|---|---|---|---|---|
| Site | Temp (°C) | pH (Total) [53] | DIC (μmol·kg−1) [53] | KPAR (m−1) [50] |
| Inshore—Nikko Bay | 30.3 ± 0.8 | 7.84 ± 0.00 | 1782.7 ± 4.1 | 0.129 |
| Offshore—Barrier Reefs | 29.0 ± 0.7 | 8.05 ± 0.01 | 1835.3 ± 15.6 | 0.085 |
| Symbol | Definition | Unit |
|---|---|---|
| Lcrl | Radiance of coral pixels | W·m−1·sr−1 |
| Lglr | Radiance from surface reflectance | W·m−1·sr−1 |
| Lstd | Radiance of Spectralon pixels | W·m−1·sr−1 |
| Rstd | Reflectance of Spectralon | Dimensionless number |
| Rblk | Reflectance of black background | Dimensionless number |
| Rcrl | Reflectance of coral | Dimensionless number |
| C. serailia | P. rugosa | |||
|---|---|---|---|---|
| Wavelength (nm) | Inshore | Offshore | Inshore | Offshore |
| S. trenchii | C Type | S. trenchii | C Type | |
| 412 | + | + | + | + |
| 420 | + | + | + | + |
| 434 | + | + | + | + |
| 455 | + | + | + | + |
| 474 | - | + | + | + |
| 492 | + | + | + | + |
| 504 | - | + | + | + |
| 524 | + | + | + | + |
| 536 | - | + | + | - |
| 549 | + | + | + | + |
| 559 | + | + | + | + |
| 579 | + | + | + | + |
| 593 | + | + | + | + |
| 617 | + | + | + | + |
| 636 | + | + | + | + |
| 666 | + | + | + | - |
| 689 | + | + | - | + |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, B.J.; Dierssen, H.M.; LaJeunesse, T.C.; Hoadley, K.D.; Warner, M.E.; Kemp, D.W.; Bateman, T.G. Spectral Reflectance of Palauan Reef-Building Coral with Different Symbionts in Response to Elevated Temperature. Remote Sens. 2016, 8, 164. https://doi.org/10.3390/rs8030164
Russell BJ, Dierssen HM, LaJeunesse TC, Hoadley KD, Warner ME, Kemp DW, Bateman TG. Spectral Reflectance of Palauan Reef-Building Coral with Different Symbionts in Response to Elevated Temperature. Remote Sensing. 2016; 8(3):164. https://doi.org/10.3390/rs8030164
Chicago/Turabian StyleRussell, Brandon J., Heidi M. Dierssen, Todd C. LaJeunesse, Kenneth D. Hoadley, Mark E. Warner, Dustin W. Kemp, and Timothy G. Bateman. 2016. "Spectral Reflectance of Palauan Reef-Building Coral with Different Symbionts in Response to Elevated Temperature" Remote Sensing 8, no. 3: 164. https://doi.org/10.3390/rs8030164
APA StyleRussell, B. J., Dierssen, H. M., LaJeunesse, T. C., Hoadley, K. D., Warner, M. E., Kemp, D. W., & Bateman, T. G. (2016). Spectral Reflectance of Palauan Reef-Building Coral with Different Symbionts in Response to Elevated Temperature. Remote Sensing, 8(3), 164. https://doi.org/10.3390/rs8030164