
Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data




Abstract
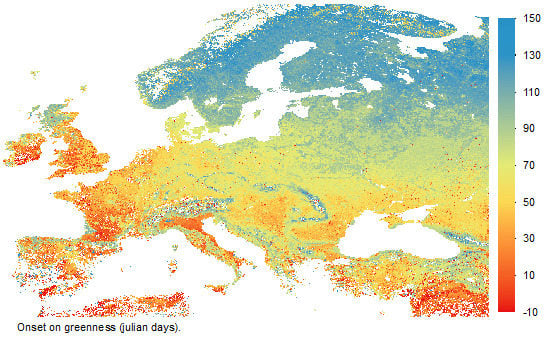
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Year of Publication | Study Area | Type | Resolution | Period | Composite Period | Vegetation Index |
|---|---|---|---|---|---|---|---|
| Myneni, et al. [29] | 1997 | Global | GIMMS AVHRR | 8 km | 1981–1991 | 10 days | NDVI |
| Tucker, et al. [30] | 2001 | Northern Hemisphere | AVHRR | 8 km | 1982–1999 | 7 days | NDVI |
| Zhou, et al. [31] | 2001 | Eurasia | GIMMS AVHRR | 9 km | 1981–1999 | 15 days | NDVI |
| Stockli and Vidale [32] | 2004 | Europe | PAL AVHRR | 8 Km | 1982–2001 | 10 days | NDVI |
| Tateishi and Ebata [33] | 2004 | Global | PAL AVHRR | 8 Km | 1982–2000 | 10 days | NDVI |
| Beck, Atzberger, Høgda, Johansen and Skidmore [22] | 2006 | Finland Norway and Sweden | MODIS | 250 m | 2000–2004 | 16 days | NDVI |
| Karlsen, et al. [34] | 2007 | Fennoscandia | GIMMS AVHRR | 8 Km | 1982–2002 | 15 days | NDVI |
| Delbart, Picard, Le Toans, Kergoat, Quegan, Woodward, Dye and Fedotova [24] | 2008 | Boreal Eurasia | SPOT VGT | 1 Km | 1998–2005 | 10 days | NDVI |
| Maignan, et al. [35] | 2008 | Europe | PAL AVHRR | 8 Km | 1982–1999 | 1 day | DVI |
| Julien and Sobrino [36] | 2009 | Global | GIMMS AVHRR | 8 Km | 1981–2003 | 10 days | NDVI |
| De Beurs and Henebry [37] | 2010 | Eurasia | PAL AVHRR | 8 km | 1981–1999 | 10 days | NDVI |
| Hogda, et al. [38] | 2011 | Fennoscandia | GIMMS AVHRR | 8 Km | 1982–2011 | 15 days | NDVI |
| Jeong, et al. [39] | 2011 | Northern Hemisphere | GIMMS AVHRR | 8 km | 1982–2008 | 15 days | NDVI |
| Ivits, et al. [40] | 2012 | Europe | GIMMS AVHRR | 8 Km | 1982–2006 | 15 days | NDVI |
| O’Connor, Dwyer, Cawkwell and Eklundh [23] | 2012 | Ireland | MERIS | 1.2 Km | 2003–2009 | 10 days | MGVI |
| Atzberger, et al. [41] | 2013 | Europe | GIMMS AVHRR MODIS | 8 Km | 2003–2011 | 15 days | NDVI |
| Hamunyela, Verbesselt, Roerink and Herold [25] | 2013 | Western Europe | MODIS | 250 m | 2001–2011 | 16 days | NDVI |
| Han, Luo and Li [26] | 2013 | Europe | SPOT VGT | 1 Km | 1999–2005 | 10 days | NDVI |
| Klisch, Atzberger and Luminari [27] | 2014 | Europe | MODIS | 250 m | 2003–2011 | 16 days | NDVI |
| Zhang, et al. [42] | 2014 | Global | AVHRR and MODIS | 5 Km | 1982–2010 | 3 days | EVI |
2. Data and Methods
2.1. Dataset
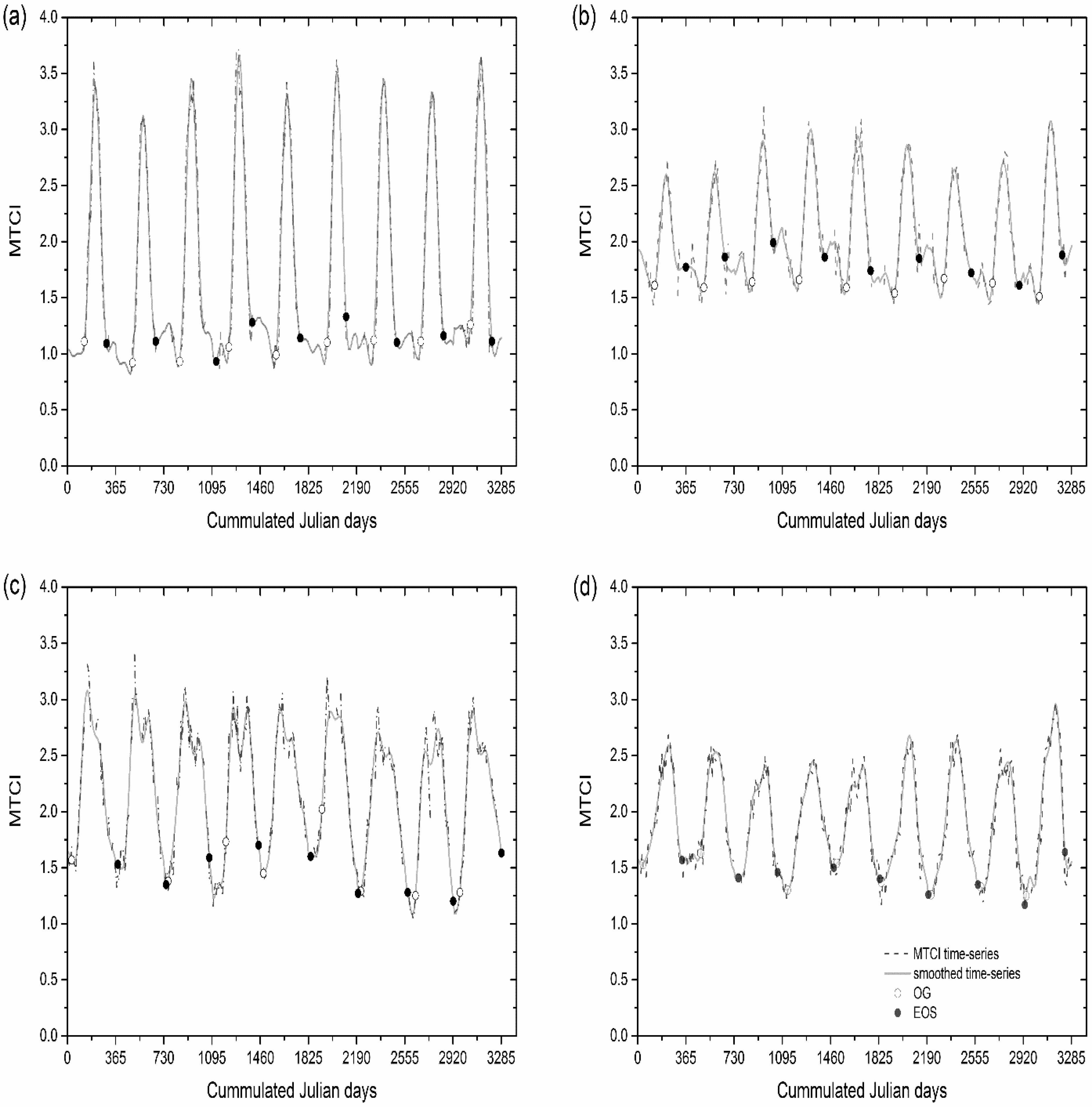
2.2. Phenology Estimates
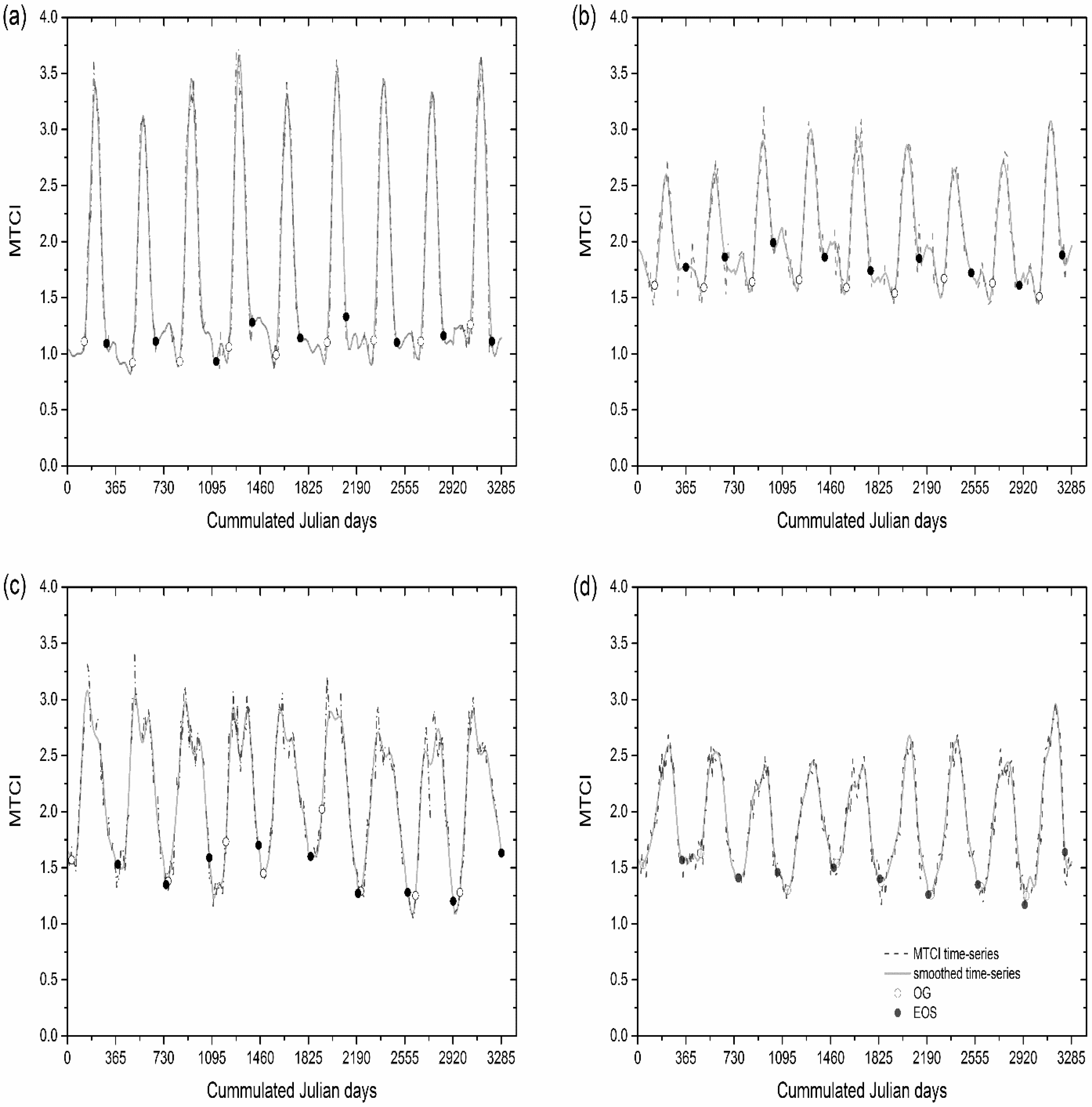
3. Results
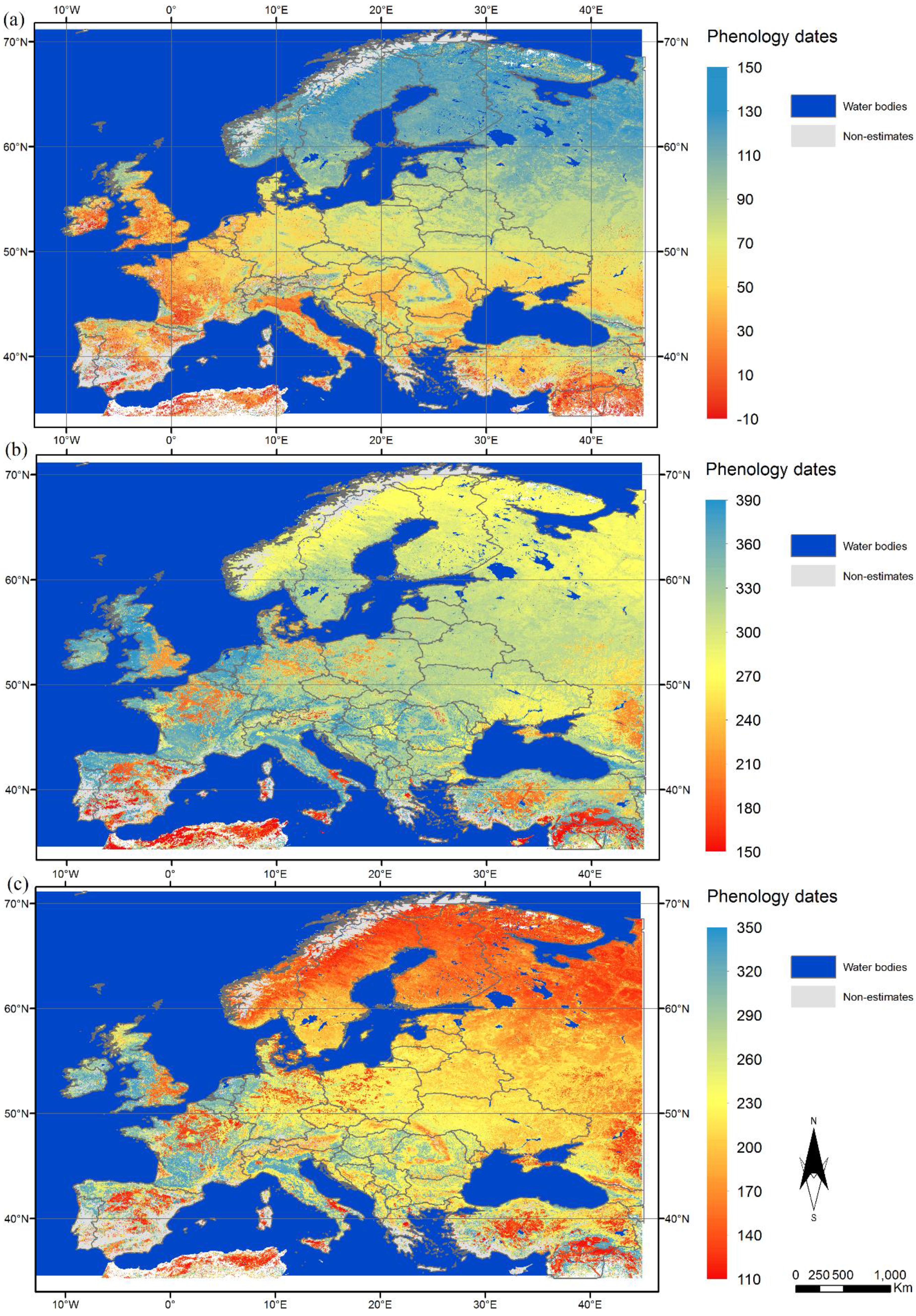
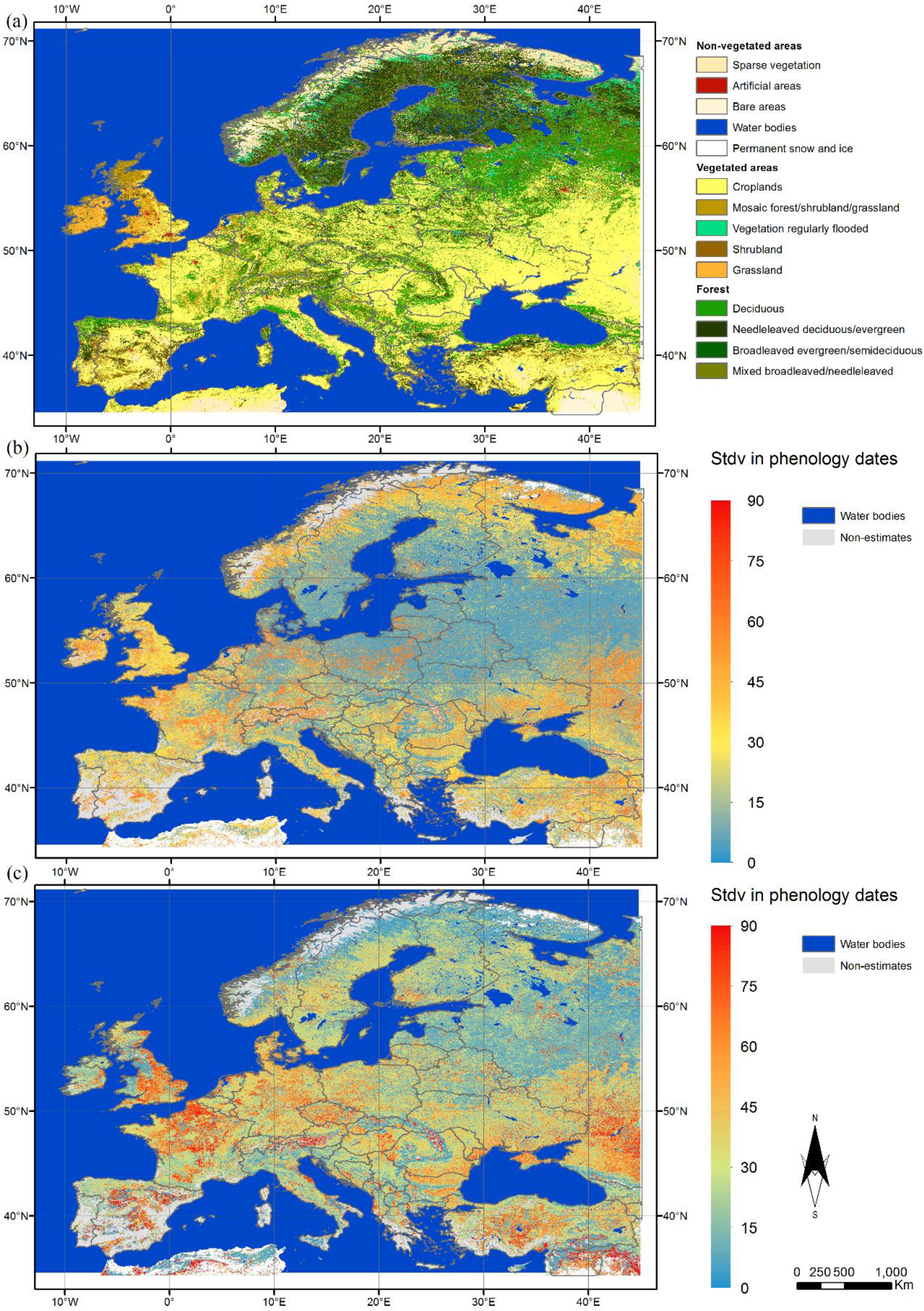
3.1. Spatial Variation in Phenological Metrics
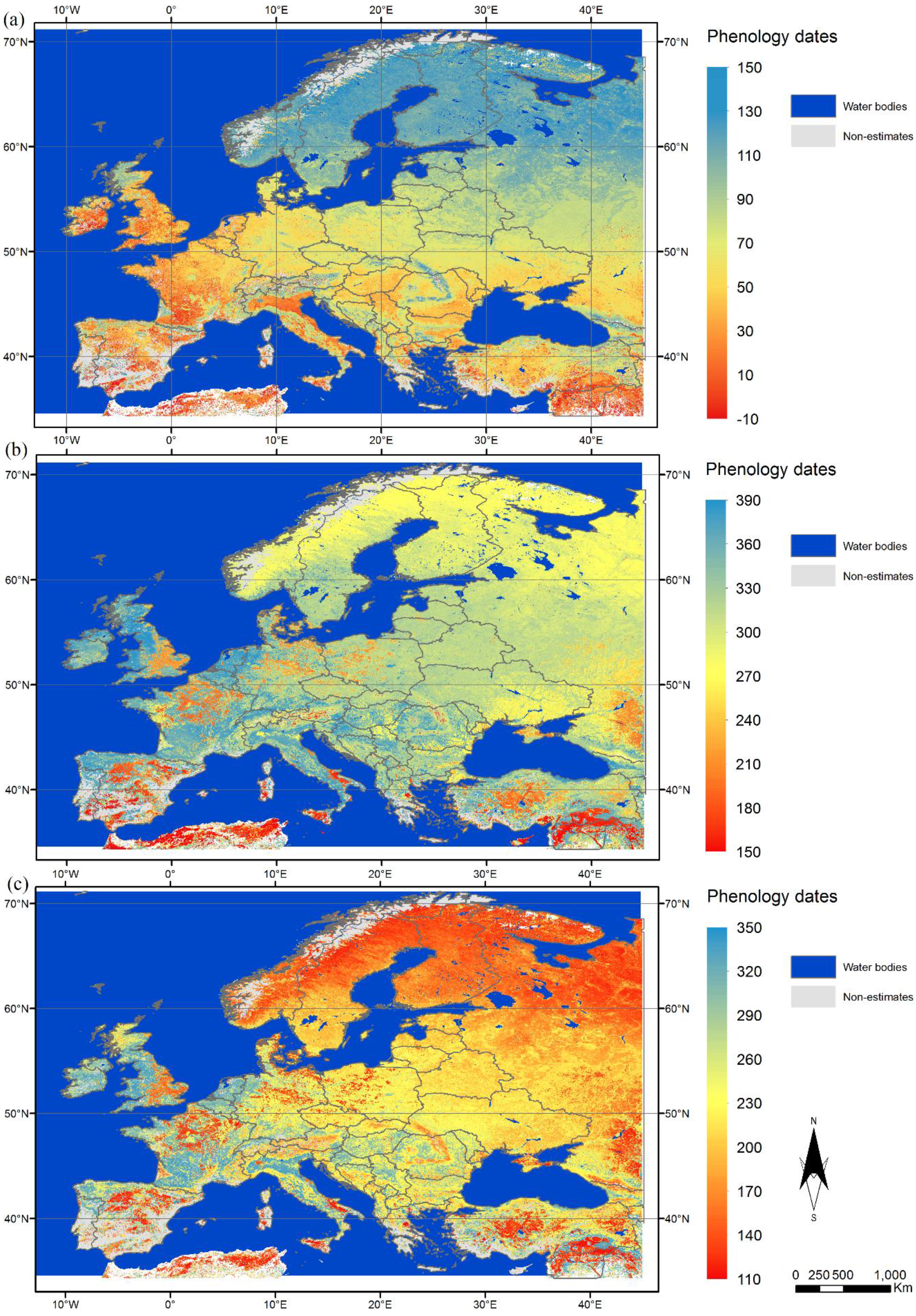
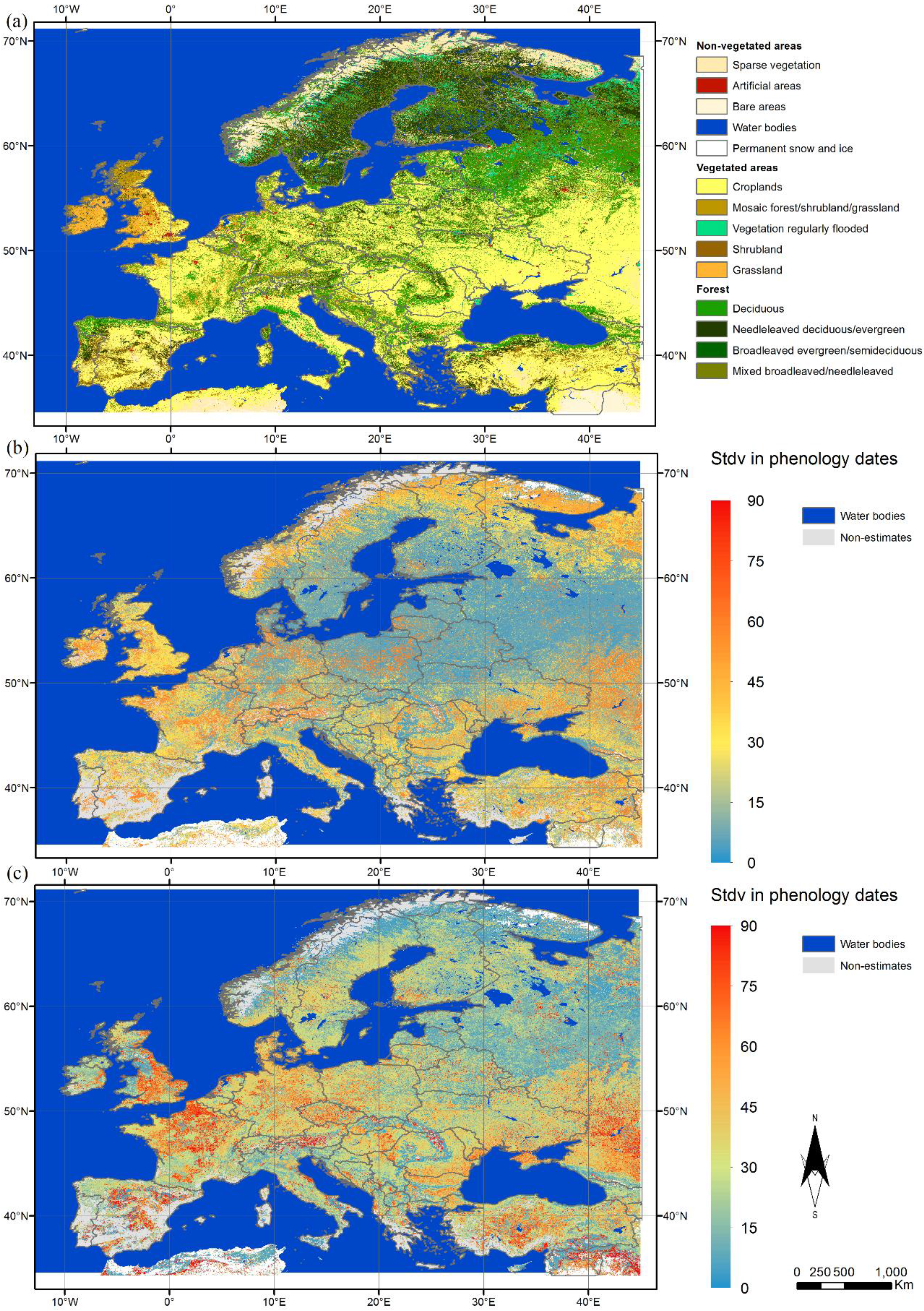
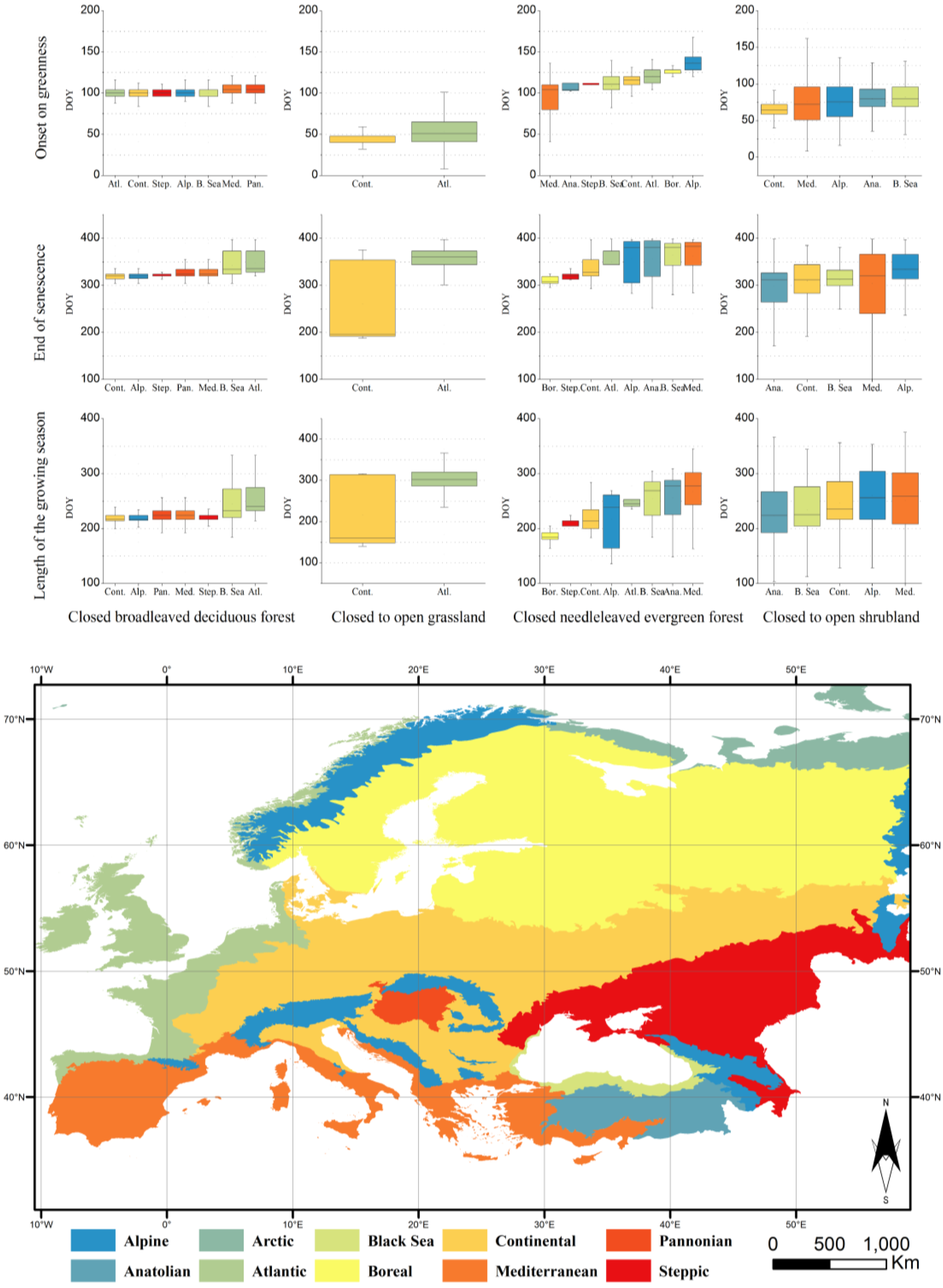
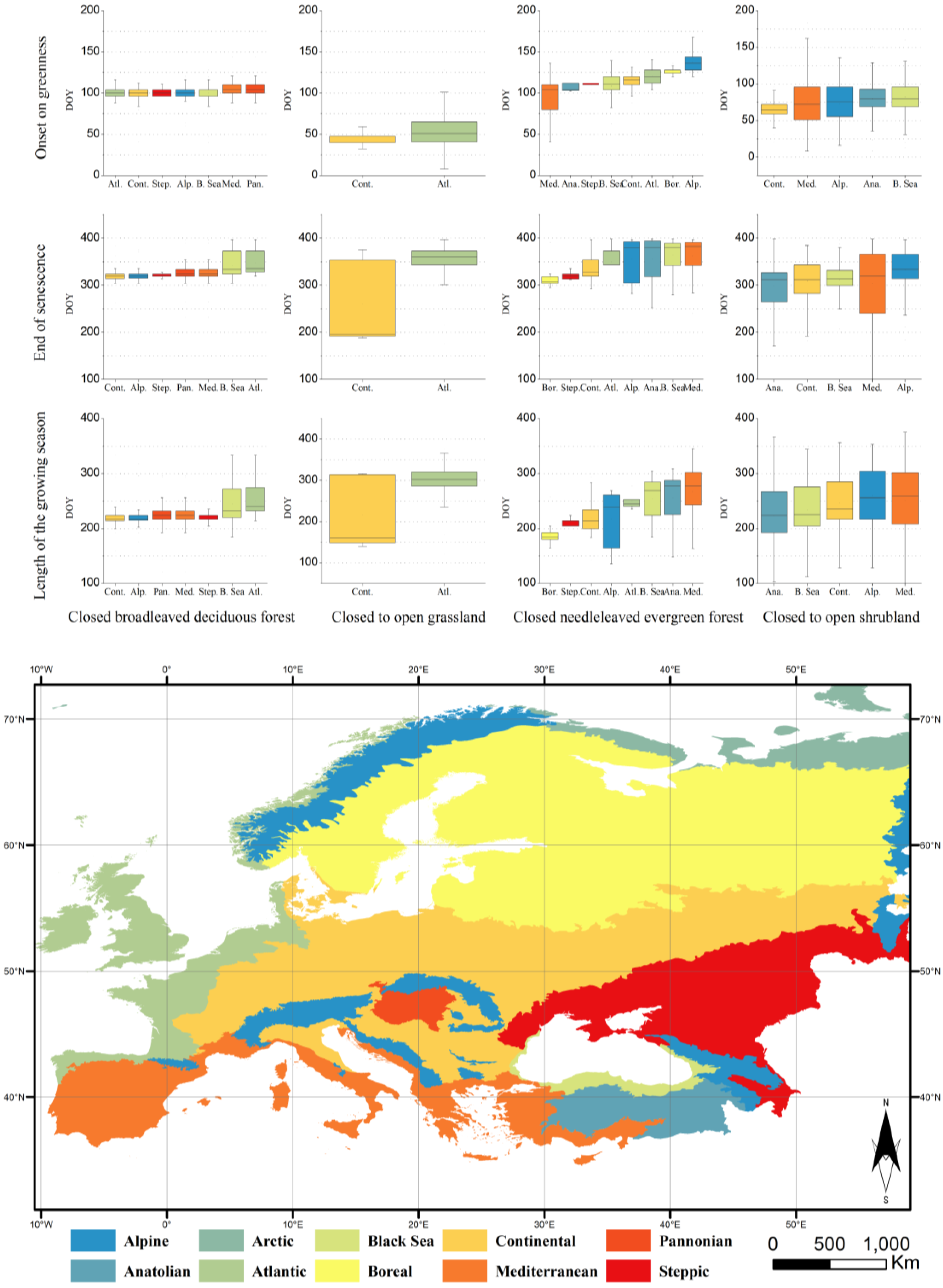
3.2. Characterisation of the Phenology of the Main Biogeographical Regions in Europe
4. Discussion
| Broadleaved Deciduous Forest | ||||||
| Alpine | Anatolian | Black Sea | Continental | Mediterranean | Panonian | |
| Anatolian | 0.067 | -- | ||||
| Black Sea | 0 * | 0.1 | -- | |||
| Continental | 0.808 | 0.066 | 0 * | -- | ||
| Mediterranean | 0 * | 0.027 * | 0 * | 0 * | -- | |
| Panonian | 0 * | 0.105 | 0.699 | 0 * | 0 * | -- |
| Steppic | 0.467 | 0.062 | 0 * | 0.507 | 0 * | 0 * |
| Needleleaved Evergreen Forest | ||||||
| Alpine | Anatolian | Black Sea | Continental | Mediterranean | ||
| Anatolian | 0 * | -- | ||||
| Black Sea | 0 * | 0.03 * | -- | |||
| Continental | 0 * | 0 * | 0* | -- | ||
| Mediterranean | 0 * | 0.691 | 0.001 * | 0 * | -- | |
| Steppic | 0 * | 0.03 * | 0.816 | 0.021 * | 0.002 * | |
| Shrublands | ||||||
| Alpine | Anatolian | Black Sea | Continental | |||
| Anatolian | 0 * | -- | ||||
| Black Sea | 0 * | 0 * | -- | |||
| Continental | 0 * | 0 * | 0 * | -- | ||
| Mediterranean | 0 * | 0 * | 0 * | 0 * | ||
| Grasslands | ||||||
| Continental | ||||||
| Atlantic | 0.826 * | |||||
| Broadleaved Deciduous Forest | ||||||
| Alpine | Anatolian | Black Sea | Continental | Mediterranean | Panonian | |
| Anatolian | 0.271 | -- | ||||
| Black Sea | 0 * | 0.137 | -- | |||
| Continental | 0.324 | 0.25 | 0 * | -- | ||
| Mediterranean | 0 * | 0.999 | 0 * | 0 * | -- | |
| Panonian | 0.168 | 0.364 | 0 * | 0.086 | 0 * | -- |
| Steppic | 0.021 * | 0.366 | 0 * | 0.001 * | 0 * | 0.949 |
| Needleleaved Evergreen Forest | ||||||
| Alpine | Anatolian | Black Sea | Continental | Mediterranean | ||
| Anatolian | 0.836 | -- | ||||
| Black Sea | 0.154 | 0.202 | -- | |||
| Continental | 0.048 * | 0.328 | 0 * | -- | ||
| Mediterranean | 0 * | 0 * | 0 * | 0 * | -- | |
| Steppic | 0 * | 0.018 * | 0 * | 0.006 * | 0 * | |
| Shrublands | ||||||
| Alpine | Anatolian | Black Sea | Continental | |||
| Alpine | -- | |||||
| Anatolian | 0 * | -- | ||||
| Black Sea | 0 * | 0 * | -- | |||
| Continental | 0 * | 0 * | 0.001 * | -- | ||
| Mediterranean | 0 * | 0 * | 0.38 | 0 * | ||
| Grasslands | ||||||
| Continental | ||||||
| Atlantic | 0 * | |||||
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aaasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Change Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.B.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, J.A.; Julien, Y.; Morales, L. Changes in vegetation spring dates in the second half of the twentieth century. Int. J. Remote Sens. 2011, 32, 5247–5265. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Menzel, A. Phenology: Its importance to the global change community. Clim. Change 2002, 54, 379–385. [Google Scholar] [CrossRef]
- Betts, R.A. Offset of the potential carbon sink from boreal forestation by decreases in surface albedo. Nature 2000, 408, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using Phenocam imagery. Biogeosciences Discuss. 2014, 11, 2305–2342. [Google Scholar] [CrossRef]
- Kirbyshire, A.L.; Bigg, G.R. Is the onset of the English summer advancing? Clim. Change 2010, 100, 419–431. [Google Scholar] [CrossRef]
- Fitter, A.H.; Fitter, R.S.R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, T.; Wittenzeller, M.; Haeckel, H.; Nekovar, J. Phenology in Central Europe differences and trends of spring phenophases in urban and rural areas. Int. J. Biometeorol. 2000, 44, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Defila, C.; Clot, B. Phytophenological trends in Switzerland. Int. J. Biometeorol. 2001, 45, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Ahas, R.; Aasa, R.; Menzel, A.; Fedotova, V.G.; Scheifinger, H. Changes in European spring phenology. Int. J. Climatol. 2002, 22, 1727–1738. [Google Scholar] [CrossRef]
- Studer, S.; Stöckli, R.; Appenzeller, C.; Vidale, P.L. A comparative study of satellite and ground-based phenology. Int. J. Biometeorol. 2007, 51, 405–414. [Google Scholar] [CrossRef] [PubMed]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Change Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Jeganathan, C.; Dash, J.; Atkinson, P. Characterising the spatial pattern of phenology for the tropical vegetation of India using multi-temporal MERIS chlorophyll data. Landsc. Ecol. 2010, 25, 1125–1141. [Google Scholar] [CrossRef]
- Dash, J.; Jeganathan, C.; Atkinson, P.M. The use of MERIS terrestrial chlorophyll index to study spatio-temporal variation in vegetation phenology over India. Remote Sens. Environ. 2010, 114, 1388–1402. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Maignan, F.; Bréon, F.M.; Vermote, E.; Ciais, P.; Viovy, N. Mild winter and spring 2007 over western Europe led to a widespread early vegetation onset. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef]
- Dash, J.; Curran, P.J. Evaluation of the MERIS terrestrial chlorophyll index (MTCI). Adv. Space Res. 2007, 39, 100–104. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- O’Connor, B.; Dwyer, E.; Cawkwell, F.; Eklundh, L. Spatio-temporal patterns in vegetation start of season across the island of Ireland using the MERIS global vegetation index. ISPRS J. Photogramm. Remote Sens. 2012, 68, 79–94. [Google Scholar] [CrossRef]
- Delbart, N.; Picard, G.; Le Toans, T.; Kergoat, L.; Quegan, S.; Woodward, I.; Dye, D.; Fedotova, V. Spring phenology in Boreal Eurasia over a nearly century time scale. Glob. Change Biol. 2008, 14, 603–614. [Google Scholar] [CrossRef]
- Hamunyela, E.; Verbesselt, J.; Roerink, G.; Herold, M. Trends in spring phenology of western European deciduous forests. Remote Sens. 2013, 5, 6159–6179. [Google Scholar] [CrossRef]
- Han, Q.; Luo, G.; Li, C. Remote sensing-based quantification of spatial variation in canopy phenology of four dominant tree species in Europe. J. Appl. Remote Sens. 2013, 7. [Google Scholar] [CrossRef]
- Klisch, A.; Atzberger, C.; Luminari, L. Satellite-based drought monitoring in Kenya in an operational setting. ISPRS Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, 2015, 433–439. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.; Dash, J.; Atkinson, P.M. Inter-comparison of satellite sensor land surface phenology and ground phenology in Europe. Geophys. Res. Lett. 2015, 42, 2253–2260. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Tucker, C.J.; Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Myneni, R.B.; Taylor, M.G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int. J. Biometeorol. 2001, 45, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.M.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Stockli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-year AVHRR land-surface parameter dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- Tateishi, R.; Ebata, M. Analysis of phenological change patterns using 1982–2000 advanced very high resolution radiometer (AVHRR) data. Int. J. Remote Sens. 2004, 25, 2287–2300. [Google Scholar] [CrossRef]
- Karlsen, S.R.; Solheim, I.; Beck, P.S.A.; Hogda, K.A.; Wielgolaski, F.E.; Tommervik, H. Variability of the start of the growing season in Fennoscandia, 1982–2002. Int. J. Biometeorol. 2007, 51, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Maignan, F.; Bréon, F.M.; Bacour, C.; Demarty, J.; Poirson, A. Interannual vegetation phenology estimates from global AVHRR measurements. Comparison with in situ data and applications. Remote Sens. Environ. 2008, 112, 496–505. [Google Scholar] [CrossRef]
- Julien, Y.; Sobrino, J.A. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. A land surface phenology assessment of the Northern Polar regions using MODIS reflectance time series. Can. J. Remote Sens. 2010, 36, S87–S110. [Google Scholar] [CrossRef]
- Hogda, K.A.; Tommervik, H.; Karlsen, S.R. Trends in the start of the growing season in Fennoscandia 1982–2011. Remote Sens. 2013, 5, 4304–4318. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Ho, C.-H.; Gim, H.-J.; Brown, M.E. Phenology shifts at start vs. End of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Ivits, E.; Cherlet, M.; Tóth, G.; Sommer, S.; Mehl, W.; Vogt, J.; Micale, F. Combining satellite derived phenology with climate data for climate change impact assessment. Glob. Planet. Change 2012, 88–89, 85–97. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the European continent from NDVI3G data and MODIS time series. Remote Sens. 2013, 6, 257–284. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, B.; Yu, Y. Interannual variations and trends in global land surface phenology derived from enhanced vegetation index during 1982–2010. Int. J. Biometeorol. 2014, 58, 1–18. [Google Scholar] [CrossRef]
- Defourny, P.; Vancutsem, C.; Bicheron, P.; Brockmann, C.; Nino, F.; Schouten, L.; Leroy, M. Globcover: A 300 m Global Land Cover Product for 2005 Using Envisat MERIS Time Series. Available online: http://dup.esrin.esa.int/files/131-176-131-25_2007510152728.pdf (accessed on 21 July 2015).
- Bicheron, P.; Amberg, V.; Bourg, L.; Petit, D.; Huc, M.; Miras, B.; Brockmann, C.; Hagolle, O.; Delwart, S.; Ranera, F.; et al. Geolocation assessment of MERIS globcover orthorectified products. IEEE Trans. Geosci. Remote Sens. 2011, 49, 2972–2982. [Google Scholar] [CrossRef] [Green Version]
- Verhoef, W.; Menenti, M.; Azzali, S. Cover a colour composite of NOAA-AVHRR-NDVI based on time series analysis (1981–1992). Int. J. Remote Sens. 1996, 17, 231–235. [Google Scholar] [CrossRef]
- Roerink, G.J.; Menenti, M.; Verhoef, W. Reconstructing cloudfree NDVI composites using fourier analysis of time series. Int. J. Remote Sens. 2000, 21, 1911–1917. [Google Scholar] [CrossRef]
- Atkinson, P.M.; Jeganathan, C.; Dash, J.; Atzberger, C. Inter-comparison of four models for smoothing satellite sensor time-series data to estimate vegetation phenology. Remote Sens. Environ. 2012, 123, 400–417. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Change Biol. 2006, 12, 343–351. [Google Scholar] [CrossRef]
- Delbart, N.; Picard, G.; Le Toan, T.; Kergoat, L.; Quegan, S.; Woodward, I.; Dye, D.; Fedotova, V. Spring phenology in Boreal Eurasia over a nearly century time scale. Glob. Change Biol. 2008, 14, 603–614. [Google Scholar] [CrossRef]
- Ryu, Y.; Lee, G.; Jeon, S.; Song, Y.; Kimm, H. Monitoring multi-layer canopy spring phenology of temperate deciduous and evergreen forests using low-cost spectral sensors. Remote Sens. Environ. 2014, 149, 227–238. [Google Scholar] [CrossRef]
- Karnieli, A. Natural vegetation phenology assessment by ground spectral measurements in two semi-arid environments. Int. J. Biometeorol. 2003, 47, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Atzberger, C.; Eilers, P.H.C. Evaluating the effectiveness of smoothing algorithms in the absence of ground reference measurements. Int. J. Remote Sens. 2011, 32, 3689–3709. [Google Scholar] [CrossRef]
- Seixas, J.; Carvalhais, N.; Nunes, C.; Benali, A. Comparative analysis of MODIS-FAPAR and MERIS–MGVI datasets: Potential impacts on ecosystem modeling. Remote Sens. Environ. 2009, 113, 2547–2559. [Google Scholar] [CrossRef]
- Chmielewski, F.-M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Galiano, V.F.; Dash, J.; Atkinson, P.M. Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data. Remote Sens. 2015, 7, 9390-9409. https://doi.org/10.3390/rs70709390
Rodriguez-Galiano VF, Dash J, Atkinson PM. Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data. Remote Sensing. 2015; 7(7):9390-9409. https://doi.org/10.3390/rs70709390
Chicago/Turabian StyleRodriguez-Galiano, Victor F., Jadunandan Dash, and Peter M. Atkinson. 2015. "Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data" Remote Sensing 7, no. 7: 9390-9409. https://doi.org/10.3390/rs70709390
APA StyleRodriguez-Galiano, V. F., Dash, J., & Atkinson, P. M. (2015). Characterising the Land Surface Phenology of Europe Using Decadal MERIS Data. Remote Sensing, 7(7), 9390-9409. https://doi.org/10.3390/rs70709390