
Climatic Controls on Spring Onset of the Tibetan Plateau Grasslands from 1982 to 2008


Abstract
:
1. Introduction
2. Data and Methods
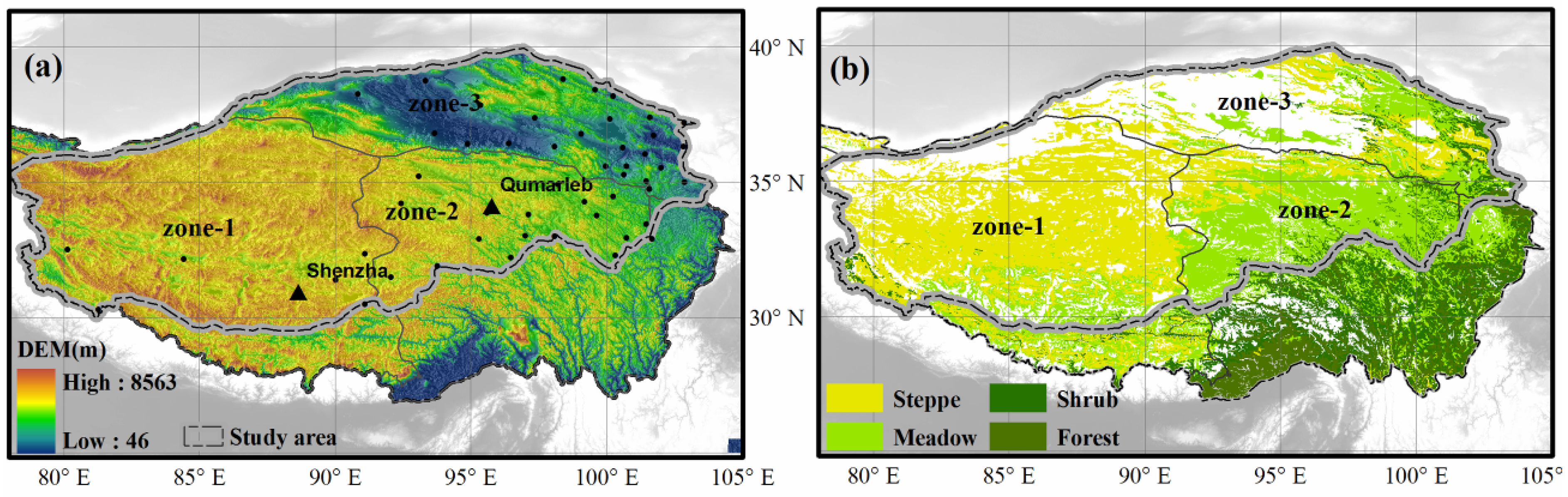
2.1. Study Area

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
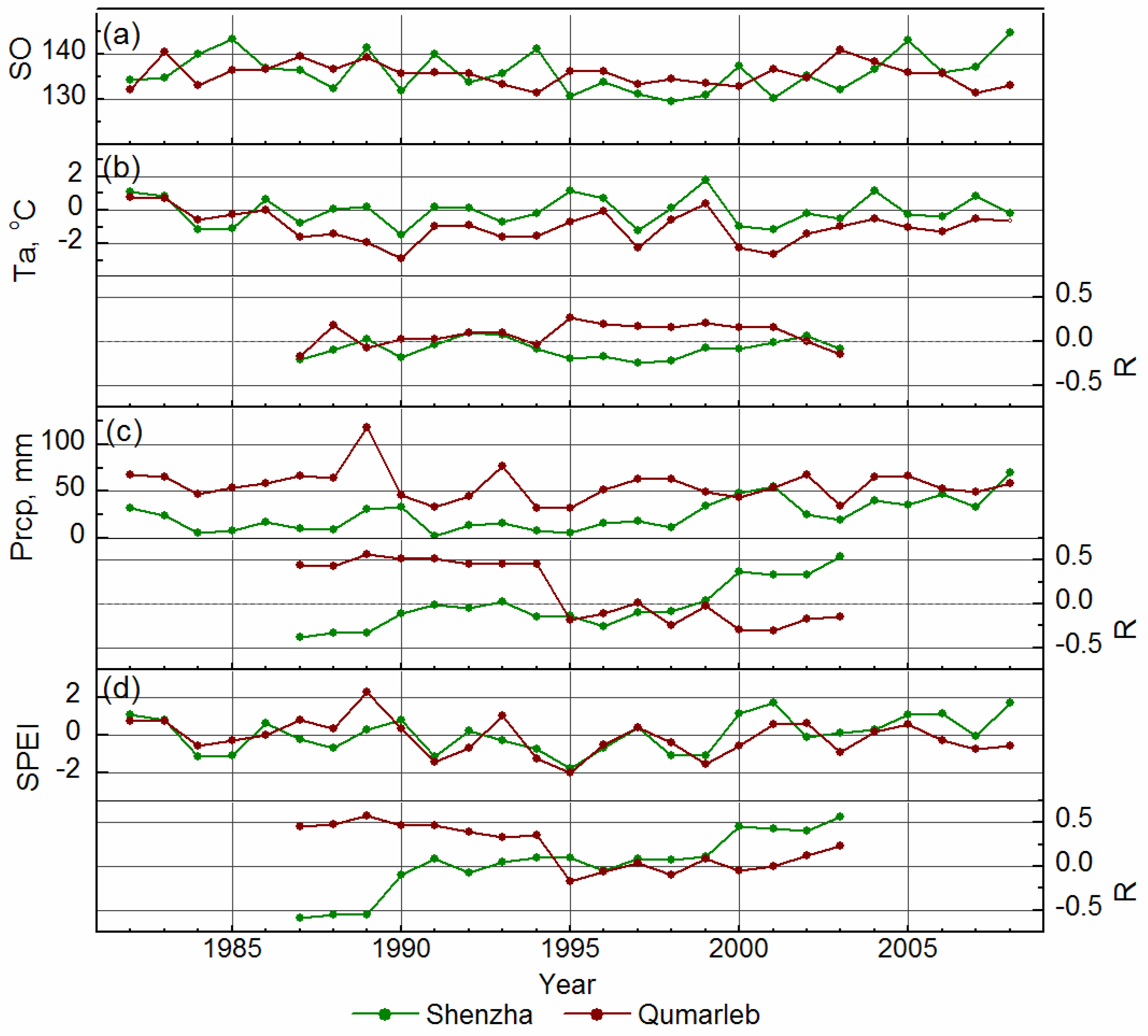{kind=link}
| Region | Period | Ta | Prcp | SPEI | |||
|---|---|---|---|---|---|---|---|
| Mean ± SD (°C) | Trend (°C·year−1) | Mean ± SD (mm) | Trend (mm·year−1) | Mean ± SD | Trend (year−1) | ||
| Zone-1 | 1982–2008 | −3.27 ± 0.87 | 0.047 ** | 22.8 ± 11.1 | 0.50 * | 0.035 ± 0.486 | −0.003 * |
| 1982–1995 | −3.49 ± 0.82 | 0.066 ** | 18.4 ± 8.9 | −0.68 * | −0.004 ± 0.476 | −0.057 ** | |
| 1996–2008 | −2.96 ± 0.81 | 0.038 * | 27.4 ± 11.3 | 1.01 ** | 0.077 ± 0.494 | 0.001 ** | |
| Zone-2 | 1982–2008 | −3.51 ± 0.77 | 0.045 ** | 73.0 ± 11.5 | −0.06 | 0.081 ± 0.426 | −0.016 * |
| 1982–1995 | −3.80 ± 0.64 | 0.054 * | 71.9 ± 2.6 | −0.79 * | 0.128 ± 0.405 | −0.037 ** | |
| 1996–2008 | −3.16 ± 0.77 | 0.039 | 73.8 ± 10.1 | −0.67 | 0.022 ± 0.441 | −0.049 * | |
| Zone-3 | 1982–2008 | −2.18 ± 0.80 | 0.073 ** | 69.6 ± 16.7 | −0.92 ** | 0.065 ± 0.475 | −0.039 ** |
| 1982–1995 | −2.61 ± 0.63 | 0.062 ** | 74.6 ± 17.2 | −2.08 ** | 0.280 ± 0.412 | −0.058 ** | |
| 1996–2008 | −1.61 ± 0.58 | 0.035 | 65.1 ± 14.9 | −1.23 | −0.167 ± 0.431 | −0.056 * | |
2.2. Datasets
2.3. Methods
2.3.1. Satellite Data Analysis
2.3.2. Statistical Analysis
3. Results
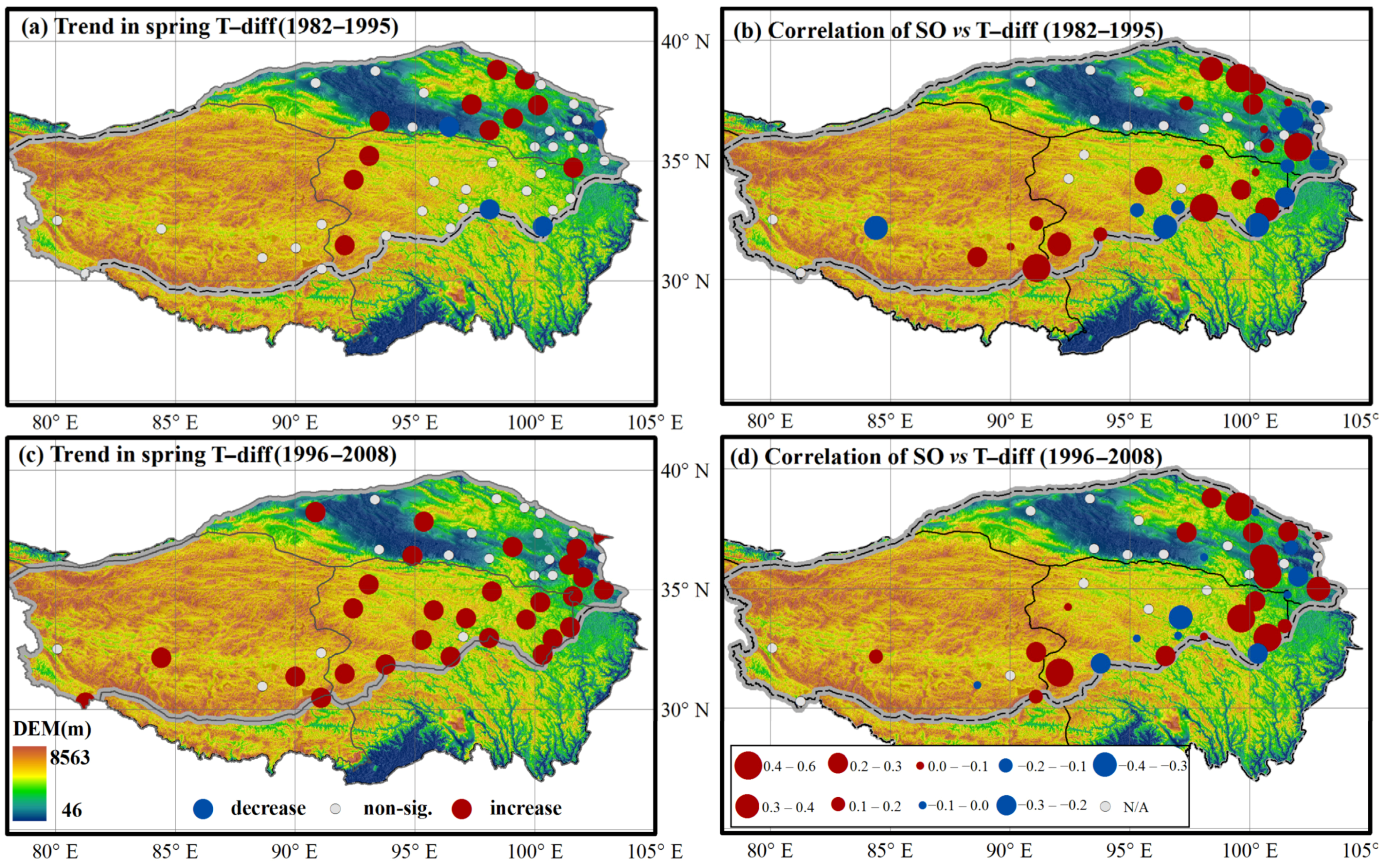
3.1. Variations in Environmental Indicators
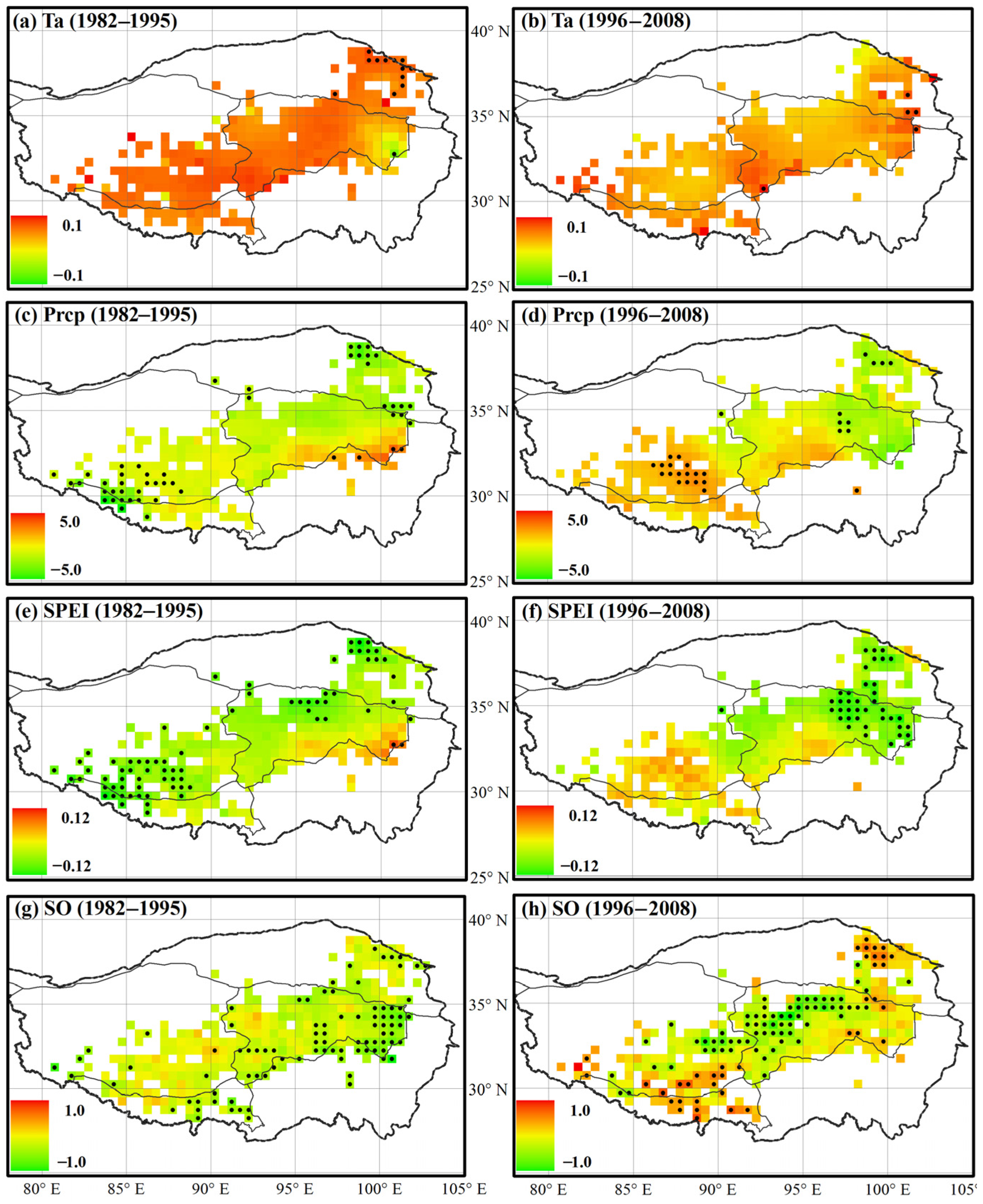
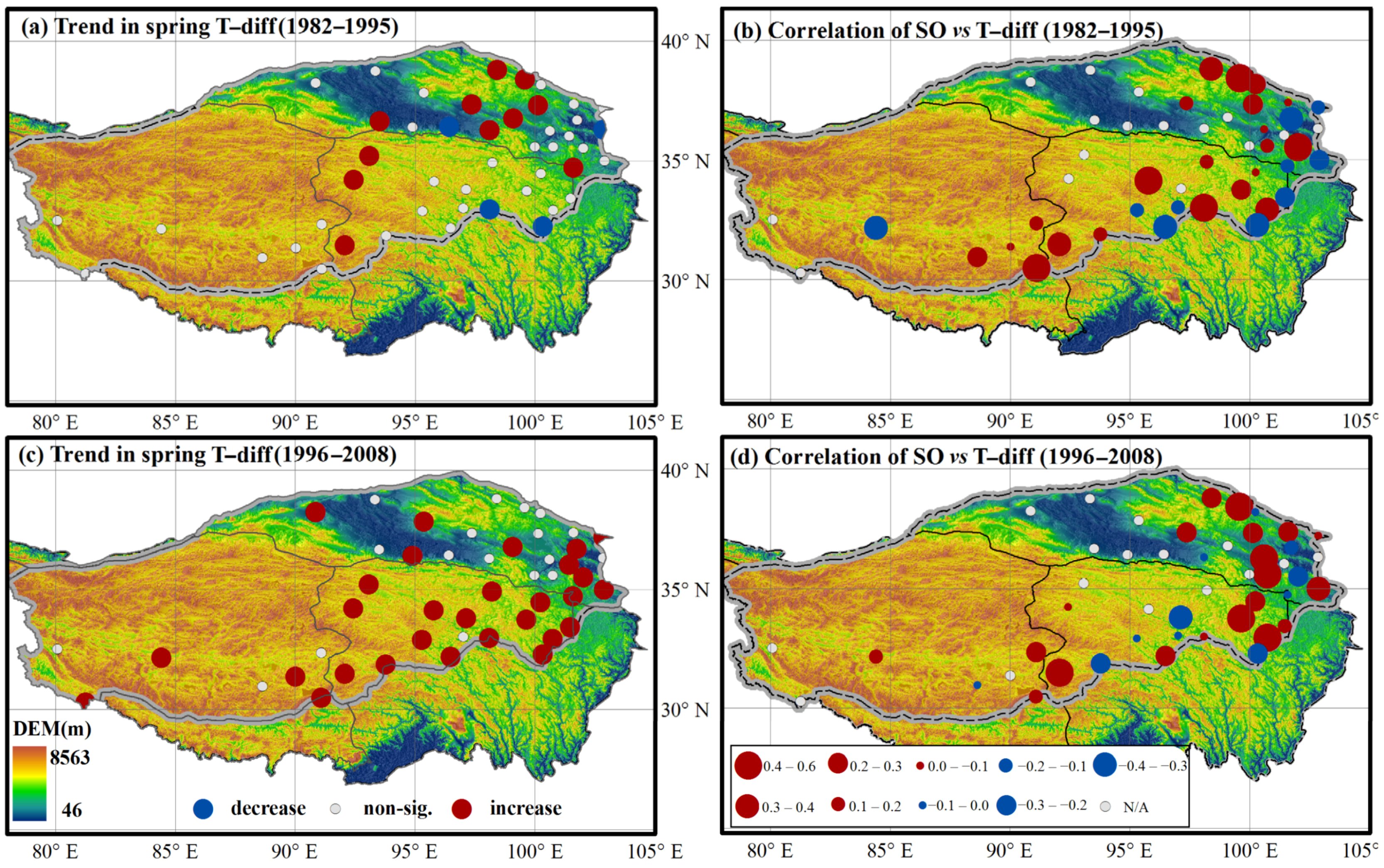
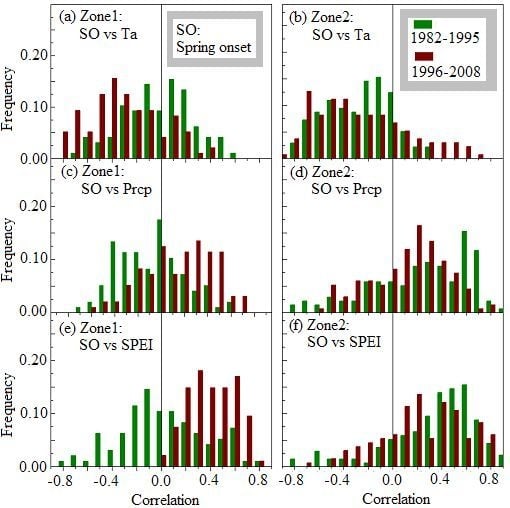
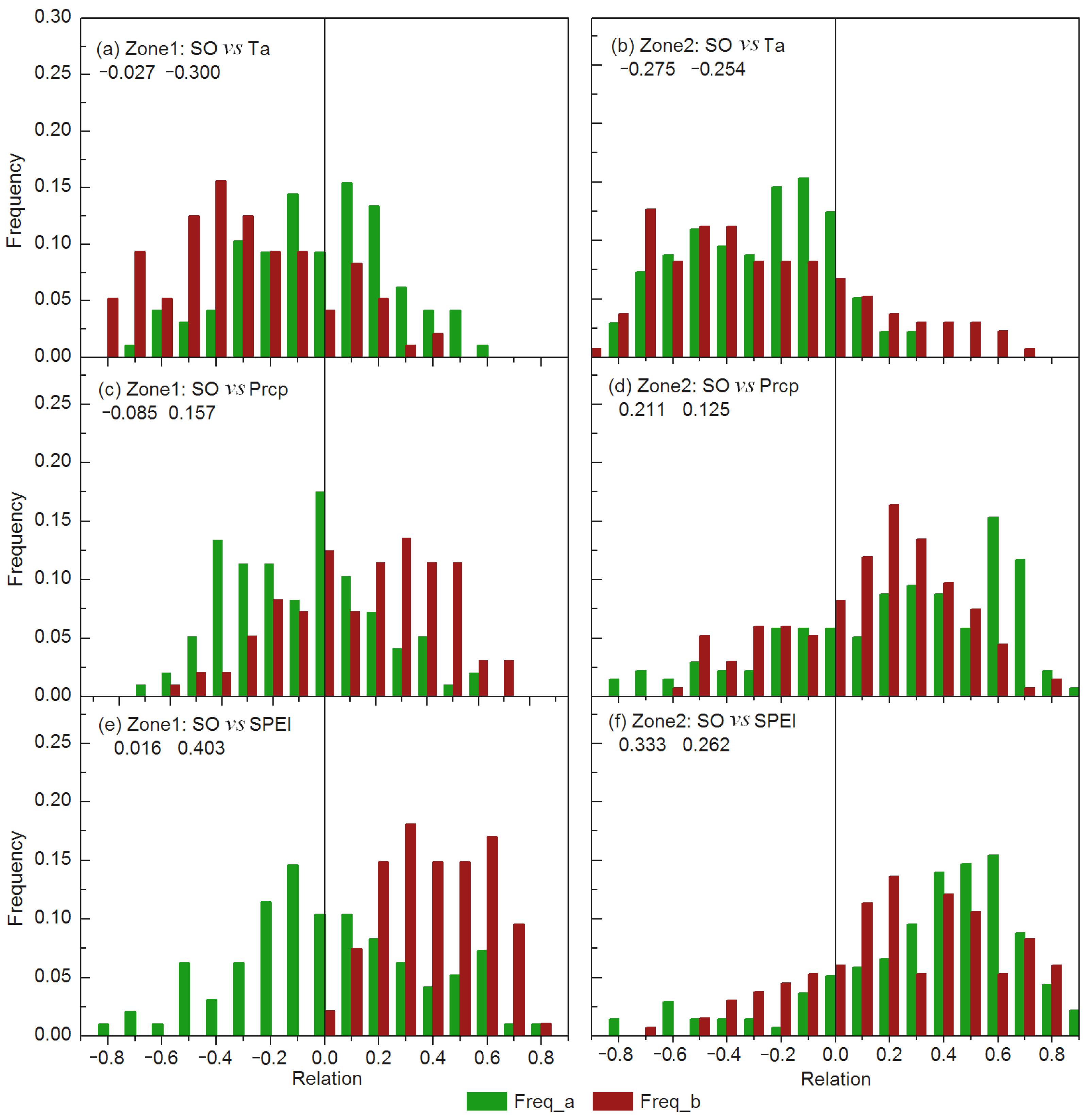
3.2. Variations in Spring Onset and Associated Climate Controls
| Region | Period | Spring Onset | Mean Correlation with Spring Onset | |||
|---|---|---|---|---|---|---|
| Mean ± SD (day of year) | Trend (day·year−1) | Ta Mean ± SD | Prcp Mean ± SD | SPEI Mean ± SD | ||
| Zone-1 | 1982–2008 | 141.6 ± 2.1 | −0.187 ** | −0.121 ± 0.206 | 0.044 ± 0.203 | 0.185 ± 0.207 |
| 1982–1995 | 143.0 ± 1.0 | −0.121 ** | −0.027 ± 0.287 | −0.085 ± 0.283 | 0.016 ± 0.345 | |
| 1996–2008 | 140.1 ± 1.7 | −0.015 * | −0.300 ± 0.294 | 0.157 ± 0.299 | 0.403 ± 0.190 | |
| Zone-2 | 1982–2008 | 139.6 ± 2.5 | −0.257 ** | −0.268 ± 0.218 | 0.149 ± 0.259 | 0.266 ± 0.234 |
| 1982–1995 | 141.3 ± 2.0 | −0.274 ** | −0.275 ± 0.263 | 0.211 ± 0.432 | 0.333 ± 0.362 | |
| 1996–2008 | 137.8 ± 1.5 | −0.207 * | −0.254 ± 0.377 | 0.125 ± 0.312 | 0.262 ± 0.348 | |
| Zone-3 | 1982–2008 | 140.4 ± 1.9 | −0.123 ** | −0.166 ± 0.163 | −0.099 ± 0.261 | −0.109 ± 0.248 |
| 1982–1995 | 141.2 ± 1.8 | −0.211 ** | −0.054 ± 0.264 | −0.181 ± 0.317 | −0.076 ± 0.298 | |
| 1996–2008 | 139.4 ± 1.2 | 0.119 ** | −0.190 ± 0.319 | −0.050 ± 0.350 | −0.101 ± 0.329 | |
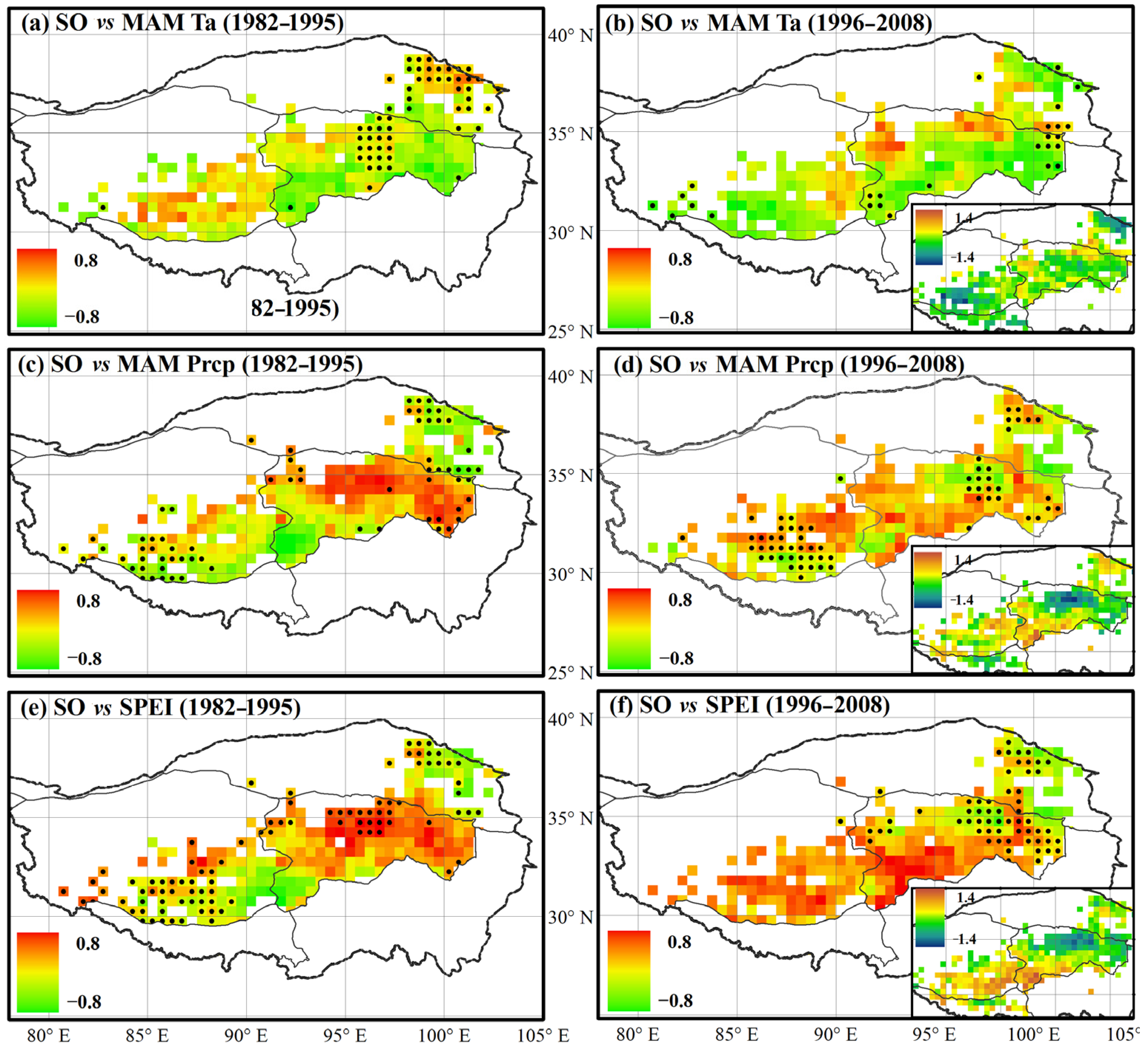
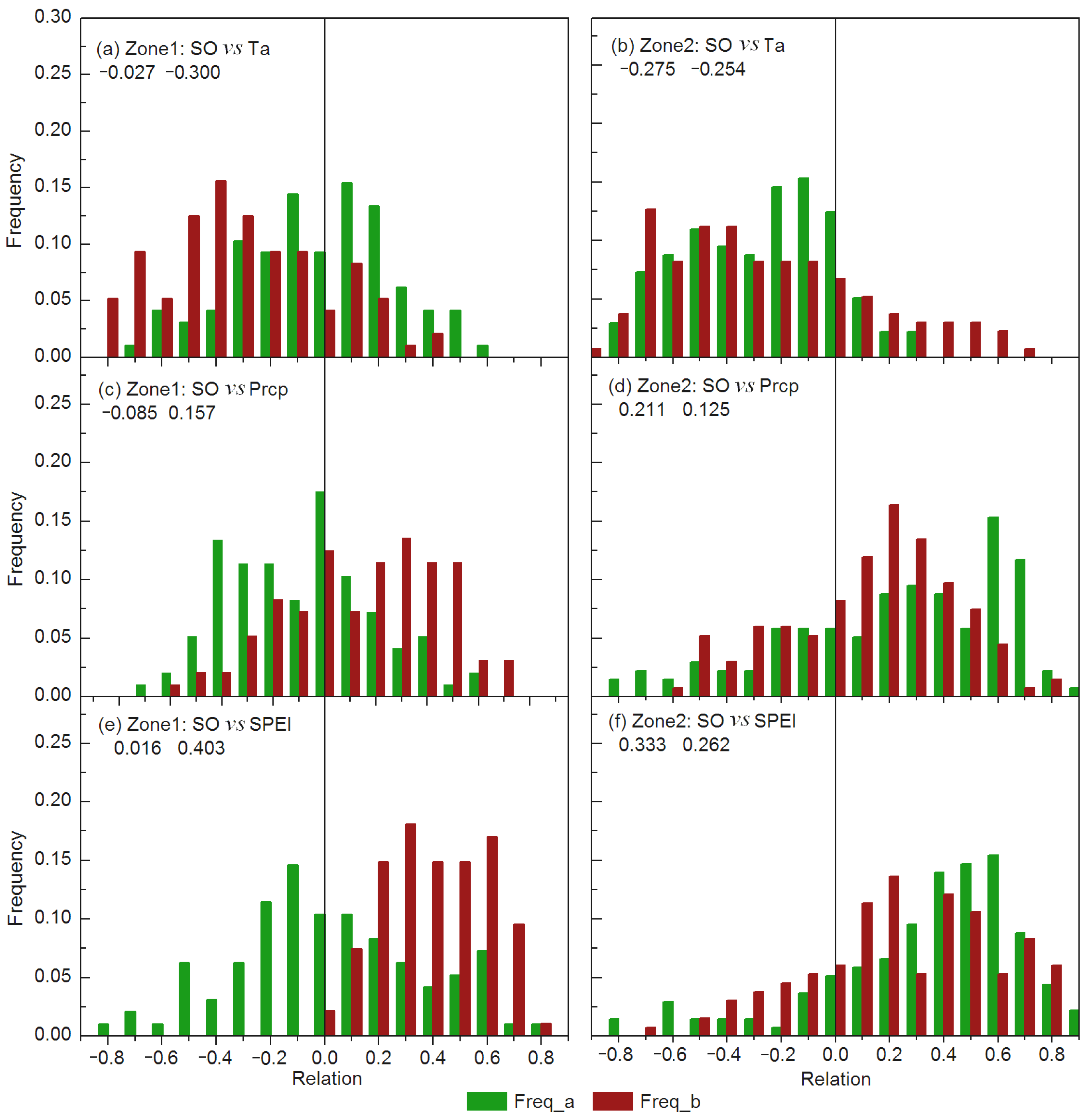
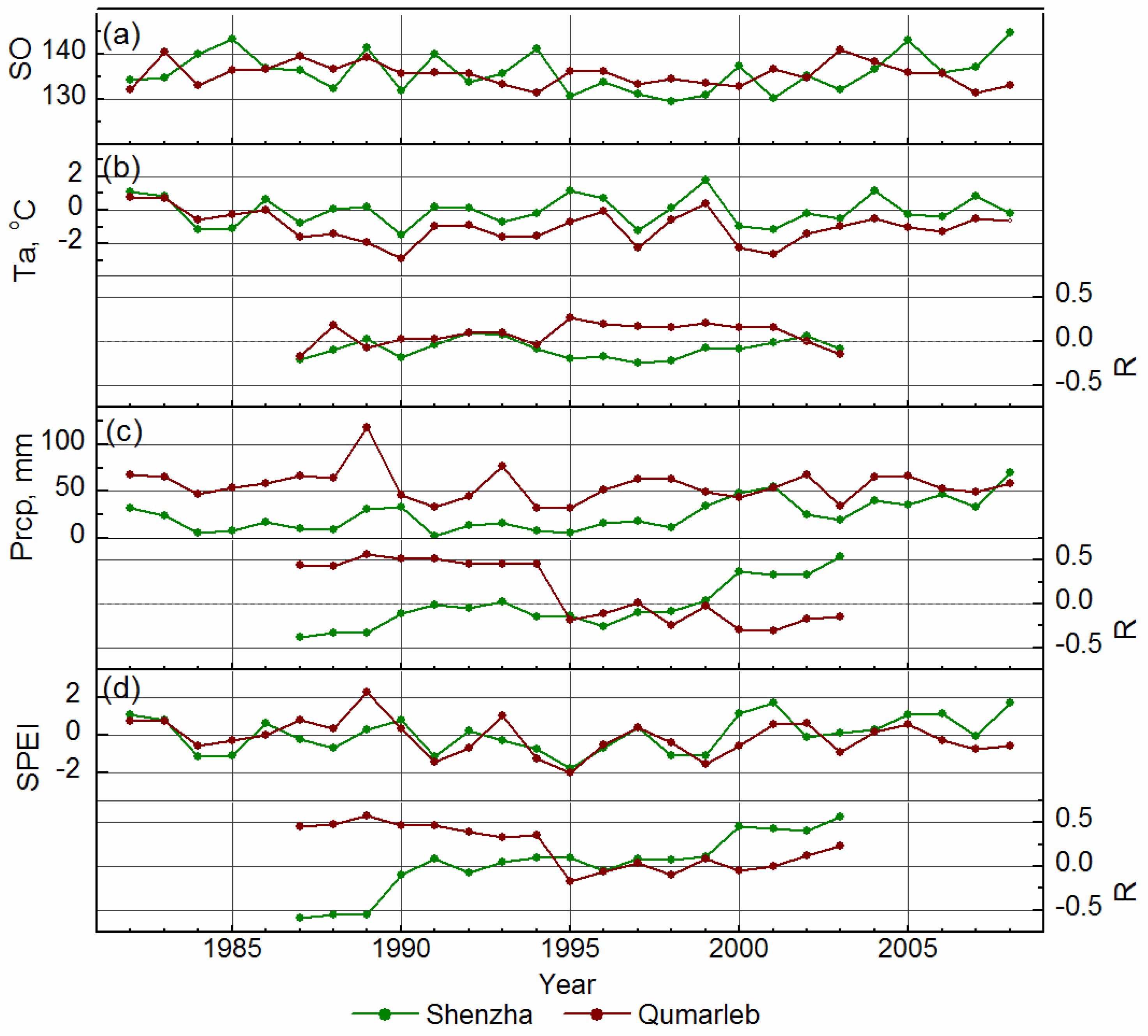
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, K.; Wu, H.; Qin, J.; Lin, C.; Tang, W.; Chen, Y. Recent climate changes over the Tibetan Plateau and their impacts on energy and water cycle: A review. Glob. Planet. Chang. 2014, 112, 79–91. [Google Scholar] [CrossRef]
- Yao, T.; Masson-Delmotte, V.; Gao, J.; Yu, W.; Yang, X.; Risi, C.; Sturm, C.; Werner, M.; Zhao, H.; He, Y.; et al. A review of climatic controls on δ18o in precipitation over the Tibetan Plateau: Observations and simulations. Rev. Geophys. 2013, 51, 525–548. [Google Scholar] [CrossRef]
- Klein, J.A.; Hopping, K.A.; Yeh, E.T.; Nyima, Y.; Boone, R.B.; Galvin, K.A. Unexpected climate impacts on the Tibetan Plateau: Local and scientific knowledge in findings of delayed summer. Glob. Environ. Chang. 2014, 28, 141–152. [Google Scholar] [CrossRef]
- Piao, S.; Ciais, P.; Huang, Y.; Shen, Z.; Peng, S.; Li, J.; Zhou, L.; Liu, H.; Ma, Y.; Ding, Y.; et al. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, Y.; Dong, J.; Xiao, X. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Natl. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Nelson, F.E.; Shiklomanov, N.I.; Guo, D.; Wan, G. Permafrost degradation and its environmental effects on the Tibetan Plateau: A review of recent research. Earth Sci. Rev. 2010, 103, 31–44. [Google Scholar] [CrossRef]
- Henebry, G.; de Beurs, K. Remote sensing of land surface phenology: A prospectus. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 385–411. [Google Scholar]
- Inouye, D.; Wielgolaski, F. Phenology at high altitudes. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 249–272. [Google Scholar]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-KÜBler, K.; Bissolli, P.; BraslavskÁ, O.G.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.F.; Dash, J.; Atkinson, P.M. Intercomparison of satellite sensor land surface phenology and ground phenology in Europe. Geophys. Res. Lett. 2015, 42, 2253–2260. [Google Scholar] [CrossRef]
- Donnelly, A.; Jones, M.; Sweeney, J. A review of indicators of climate change for use in Ireland. Int. J. Biometeorol. 2004, 49, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Chang. Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- De Beurs, K.; Henebry, G. Vegetation phenology in global change studies. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 483–502. [Google Scholar]
- Shen, M.; Zhang, G.; Cong, N.; Wang, S.; Kong, W.; Piao, S. Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai–Tibetan Plateau. Agric. For. Meteorol. 2014, 189, 71–80. [Google Scholar] [CrossRef]
- Liu, L.; Liu, L.; Liang, L.; Donnelly, A.; Park, I.; Schwartz, M. Effects of elevation on spring phenological sensitivity to temperature in Tibetan Plateau grasslands. Chin. Sci. Bull. 2014, 59, 4856–4863. [Google Scholar] [CrossRef]
- Yao, T.; Thompson, L.; Yang, W.; Yu, W.; Gao, Y.; Guo, X.; Yang, X.; Duan, K.; Zhao, H.; Xu, B.; et al. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nat. Clim. Chang. 2012, 2, 663–667. [Google Scholar] [CrossRef]
- Wu, X.; Liu, H. Consistent shifts in spring vegetation green-up date across temperate biomes in China, 1982–2006. Glob. Chang. Biol. 2013, 19, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Wu, T. Responses of permafrost to climate change and their environmental significance, Qinghai-Tibet Plateau. J. Geophys. Res. Earth Surf. 2007. [Google Scholar] [CrossRef]
- Li, X.; Jin, R.; Pan, X.; Zhang, T.; Guo, J. Changes in the near-surface soil freeze-thaw cycle on the Qinghai-Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2012, 17, 33–42. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, T.; Liu, Y. Permafrost temperatures and thickness on the Qinghai-Tibet Plateau. Glob. Planet. Chang. 2010, 72, 32–38. [Google Scholar] [CrossRef]
- Wang, G.; Guangsheng, L.; Chunjie, L.; Yan, Y. The variability of soil thermal and hydrological dynamics with vegetation cover in a permafrost region. Agric. For. Meteorol. 2012, 162, 44–57. [Google Scholar]
- Piao, S.; Cui, M.; Chen, A.; Wang, X.; Ciais, P.; Liu, J.; Tang, Y. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Dorji, T.; Totland, Ø.; Moe, S.R.; Hopping, K.A.; Pan, J.; Klein, J.A. Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Glob. Chang. Biol. 2013, 19, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Luedeling, E.; Xu, J. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, C.; Duan, J.; Zhu, X.; Xu, G.; Luo, C.; Zhang, Z.; Meng, F.; Li, Y.; Du, M. Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau. Agric. For. Meteorol. 2014, 189, 220–228. [Google Scholar] [CrossRef]
- Che, M.; Chen, B.; Innes, J.L.; Wang, G.; Dou, X.; Zhou, T.; Zhang, H.; Yan, J.; Xu, G.; Zhao, H. Spatial and temporal variations in the end date of the vegetation growing season throughout the Qinghai-Tibetan Plateau from 1982 to 2011. Agric. For. Meteorol. 2014, 189, 81–90. [Google Scholar] [CrossRef]
- Shen, M. Spring phenology was not consistently related to winter warming on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2011, 108, E91–E92. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D. The Zoning of Ecographical Systems in China; The Commercial Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Chen, X.; An, S.; Inouye, D.; Schwartz, M. Temperature and snowfall trigger alpine vegetation green-up on the world’s roof. Glob. Chang. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Xiong, D.; Su, Z.; Li, J.; Yang, D.; Shi, L.; Liu, G. The distribution of and factors influencing the vegetation in a gully in the dry-hot valley of Southwest China. CATENA 2014, 116, 60–67. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Wang, C.; Ji, L.; Li, J.; Wang, K.; Dai, L. The long-term trends (1982–2006) in vegetation greenness of the alpine ecosystem in the Qinghai-Tibetan Plateau. Environ. Earth Sci. 2014, 72, 1–15. [Google Scholar] [CrossRef]
- Zhang, X. Vegetation Map of the People's Republic of China (1:1,000,000); China Geological Publishing House: Beijing, China, 2007. (in Chinese) [Google Scholar]
- Zhou, H.K.; Yao, B.Q.; Xu, W.X.; Ye, X.; Fu, J.J.; Jin, Y.X.; Zhao, X.Q. Field evidence for earlier leaf-out dates in alpine grassland on the eastern Tibetan Plateau from 1990 to 2006. Biol. Lett. 2014. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Buermann, W.; Parida, B.; Jung, M.; MacDonald, G.M.; Tucker, C.J.; Reichstein, M. Recent shift in Eurasian boreal forest greening response may be associated with warmer and drier summers. Geophys. Res. Lett. 2014, 41, 1995–2002. [Google Scholar] [CrossRef]
- Hart, R.; Salick, J.; Ranjitkar, S.; Xu, J. Herbarium specimens show contrasting phenological responses to Himalayan climate. Proc. Natl. Acad. Sci. USA 2014, 111, 10615–10619. [Google Scholar] [CrossRef] [PubMed]
- Julien, Y.; Sobrino, J.A. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- Pinzon, J.; Tucker, C. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of earth observation based global long term vegetation trends—Comparing GIMMS and MODIS global NDVI time series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Eastman, J.; Sangermano, F.; Machado, E.; Rogan, J.; Anyamba, A. Global trends in seasonality of normalized difference vegetation index (NDVI), 1982–2011. Remote Sens. 2013, 5, 4799. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the european continent from NDVI 3g data and MODIS time series. Remote Sens. 2013, 6, 257. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, S.; Wang, J.; Sun, P.; Liu, W.; Hartley, D.S. Effects of seasonal snow on the growing season of temperate vegetation in China. Glob. Chang. Biol. 2013, 19, 2182–2195. [Google Scholar] [CrossRef] [PubMed]
- Delbart, N.; Kergoat, L.; Le Toan, T.; Lhermitte, J.; Picard, G. Determination of phenological dates in boreal regions using normalized difference water index. Remote Sens. Environ. 2005, 97, 26–38. [Google Scholar] [CrossRef]
- China Meteorological Administration. China Meteorological Data Sharing Service System. Available online: http://cdc.nmic.cn/home.do (accessed on 15 March 2014). (In Chinese)
- Majorowicz, J.A.; Skinner, W.R. Potential causes of differences between ground and surface air temperature warming across different ecozones in Alberta, Canada. Glob. Planet. Chang. 1997, 15, 79–91. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Eklundh, L.; Hellström, M.; Bärring, L.; Jönsson, P. Annual changes in MODIS vegetation indices of swedish coniferous forests in relation to snow dynamics and tree phenology. Remote Sens. Environ. 2010, 114, 2719–2730. [Google Scholar] [CrossRef]
- Yi, Y.; Kimball, J.S.; Reichle, R.H. Spring hydrology determines summer net carbon uptake in northern ecosystems. Environ. Res. Lett. 2014, 9, 064003. [Google Scholar] [CrossRef]
- Lioubimtseva, E.; Milanova, E.V.; Tcherkashin, P.; Solntsev, V.N. VEGETATION/SPOT4 applications for macro-regional landscape mapping. In Proceedings of VEGETATION 2000 Conference, Belgirate, Italy, 3–6 April 2000.
- Huete, A.; Didan, K.; van Leeuwen, W.; Miura, T.; Glenn, E. MODIS vegetation indices. In Land Remote Sensing and Global Environmental Change; Ramachandran, B., Justice, C.O., Abrams, M.J., Eds.; Springer: New York, NY, USA, 2011; Volume 11, pp. 579–602. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2009, 23, 1696–1718. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar]
- Chen, X. East Asia. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 9–22. [Google Scholar]
- Peng, S.; Piao, S.; Ciais, P.; Fang, J.; Wang, X. Change in winter snow depth and its impacts on vegetation in China. Glob. Chang. Biol. 2010, 16, 3004–3013. [Google Scholar] [CrossRef]
- Wang, C.; Guo, H.; Zhang, L.; Liu, S.; Qiu, Y.; Sun, Z. Assessing phenological change and climatic control of alpine grasslands in the Tibetan Plateau with MODIS time series. Int. J. Biometeorol. 2015, 59, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Entekhabi, D.; Njoku, E.G.; O’Neill, P.E.; Kellogg, K.H.; Crow, W.T.; Edelstein, W.N.; Entin, J.K.; Goodman, S.D.; Jackson, T.J.; Johnson, J.; et al. The soil moisture active and passive (SMAP) mission. Proc. IEEE 2010, 98, 704–716. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Yi, Y.; Kimball, J.S.; Kim, Y.; Song, K. Climatic Controls on Spring Onset of the Tibetan Plateau Grasslands from 1982 to 2008. Remote Sens. 2015, 7, 16607-16622. https://doi.org/10.3390/rs71215847
Zhang W, Yi Y, Kimball JS, Kim Y, Song K. Climatic Controls on Spring Onset of the Tibetan Plateau Grasslands from 1982 to 2008. Remote Sensing. 2015; 7(12):16607-16622. https://doi.org/10.3390/rs71215847
Chicago/Turabian StyleZhang, Wenjiang, Yonghong Yi, John S. Kimball, Youngwook Kim, and Kechao Song. 2015. "Climatic Controls on Spring Onset of the Tibetan Plateau Grasslands from 1982 to 2008" Remote Sensing 7, no. 12: 16607-16622. https://doi.org/10.3390/rs71215847
APA StyleZhang, W., Yi, Y., Kimball, J. S., Kim, Y., & Song, K. (2015). Climatic Controls on Spring Onset of the Tibetan Plateau Grasslands from 1982 to 2008. Remote Sensing, 7(12), 16607-16622. https://doi.org/10.3390/rs71215847