
C3 Vegetation Mapping and CO2 Fertilization Effect in the Arid Lower Heihe River Basin, Northwestern China



Abstract
:
1. Introduction
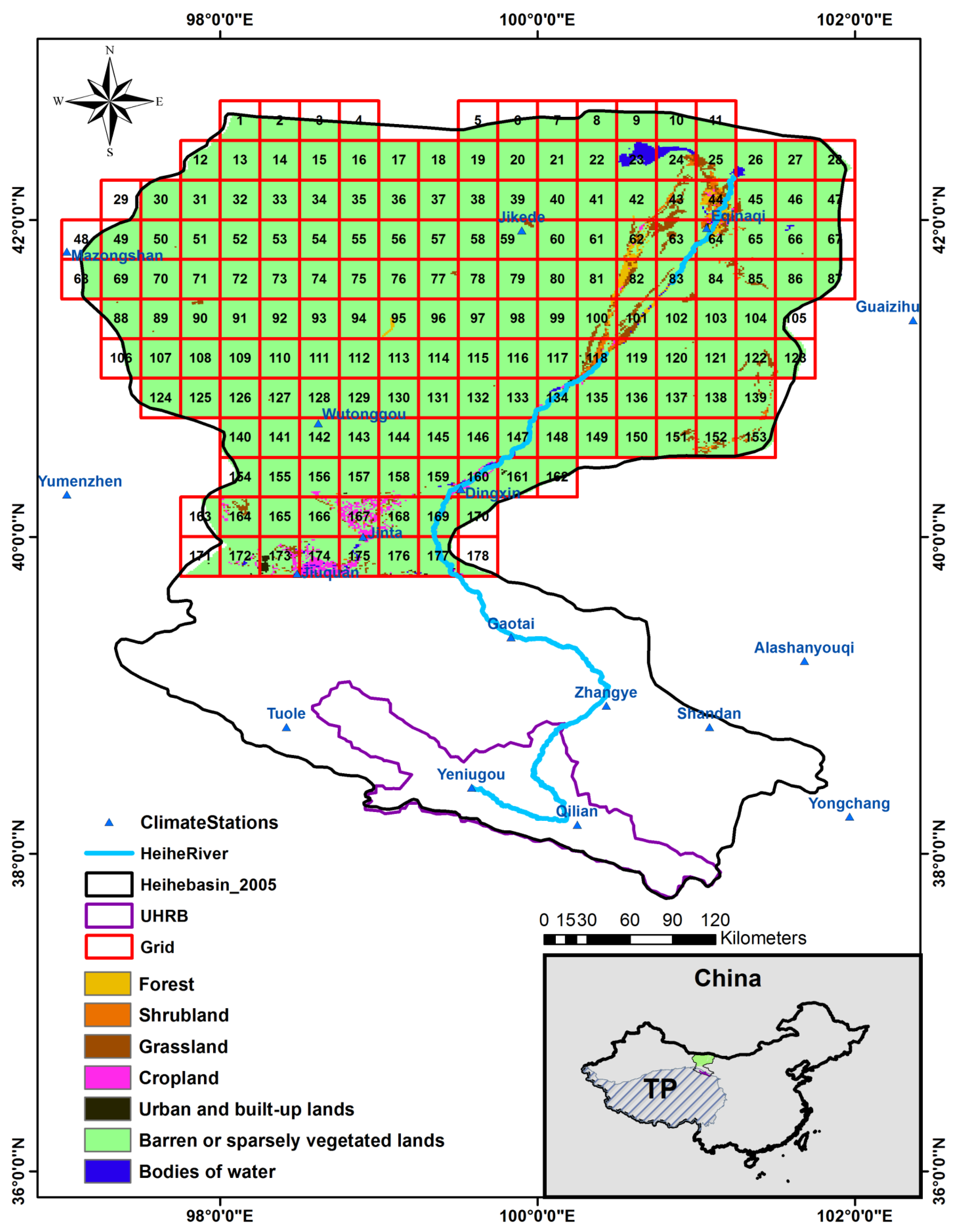
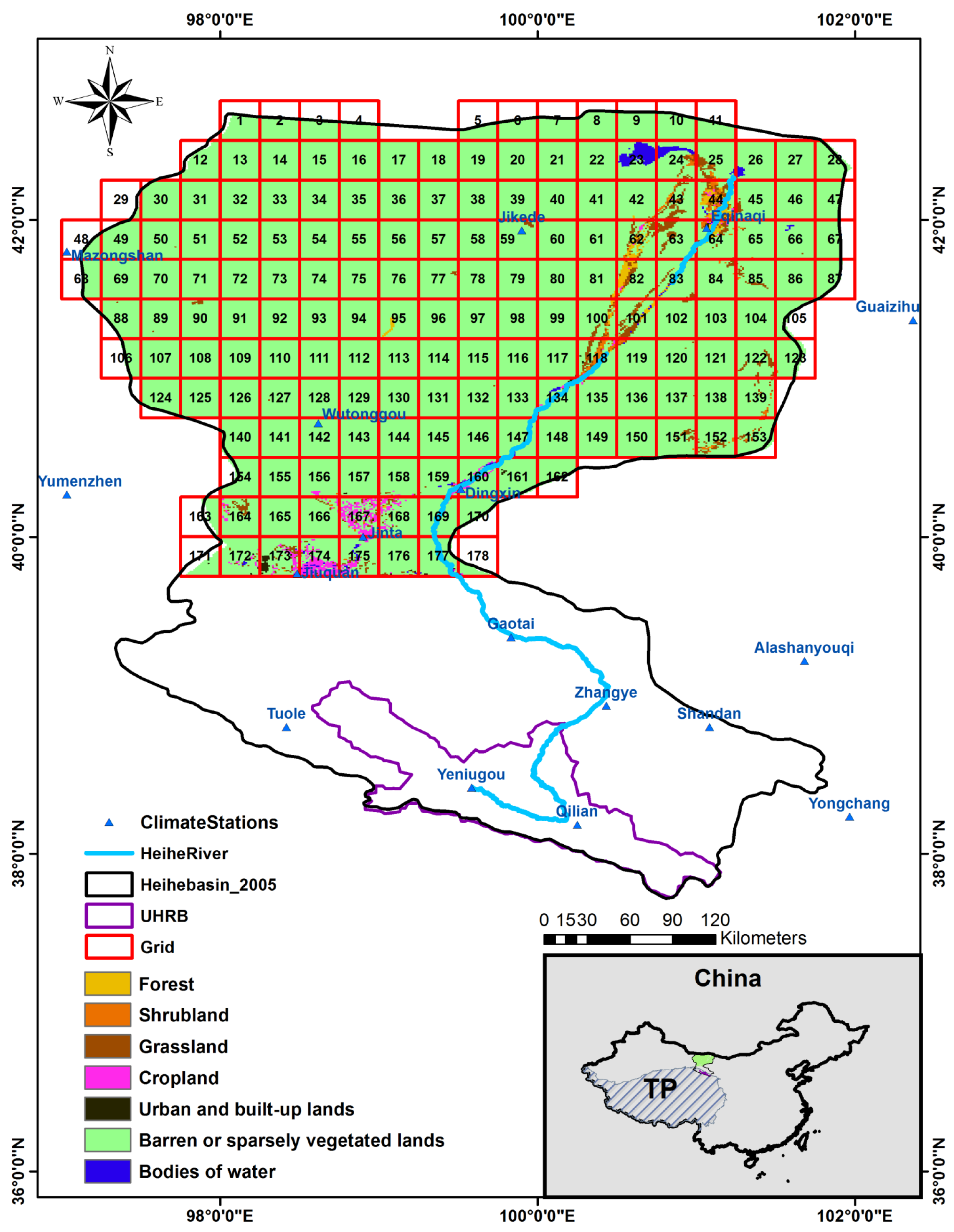
2. Study Area
3. Data and Methodology
3.1. Precipitation Data
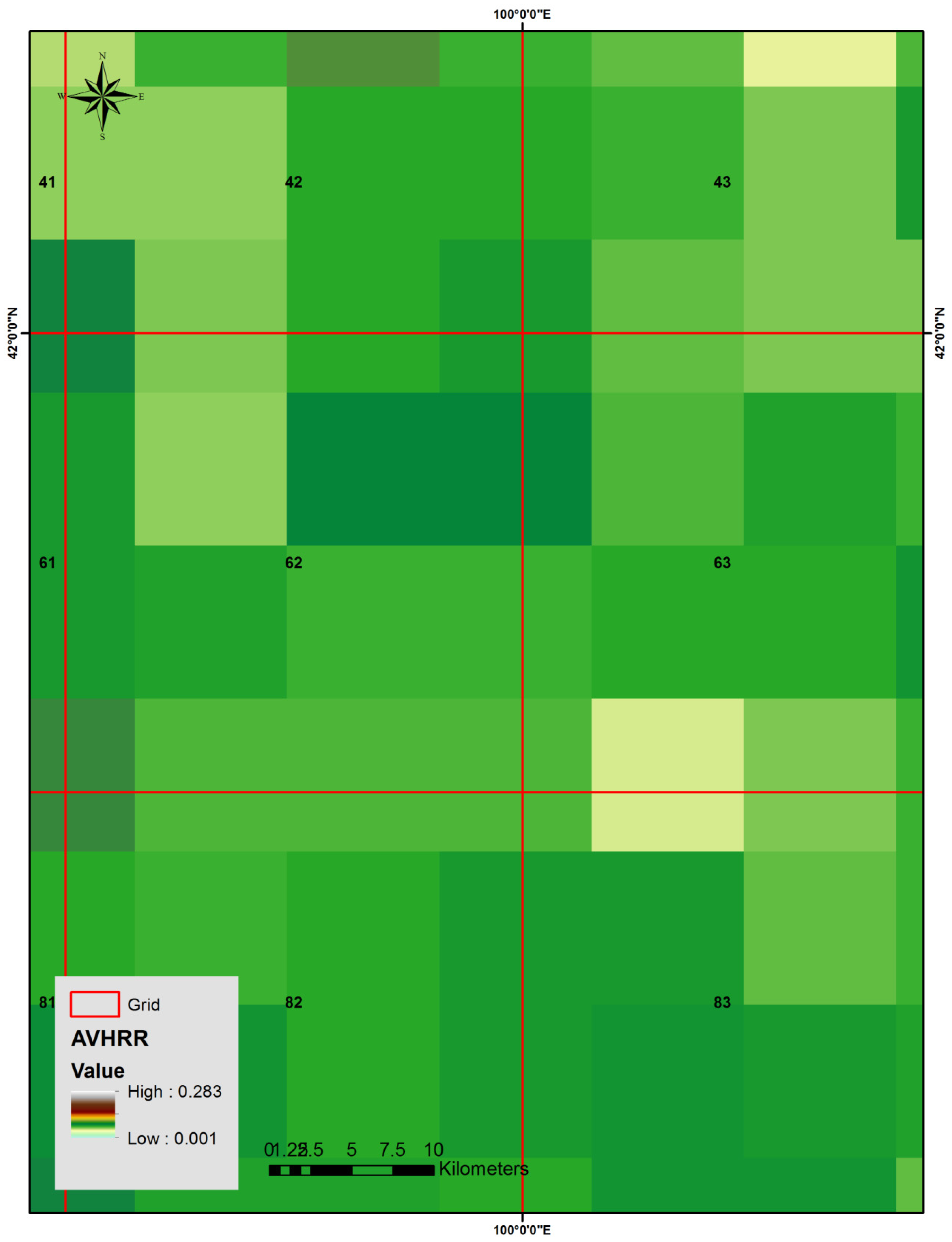
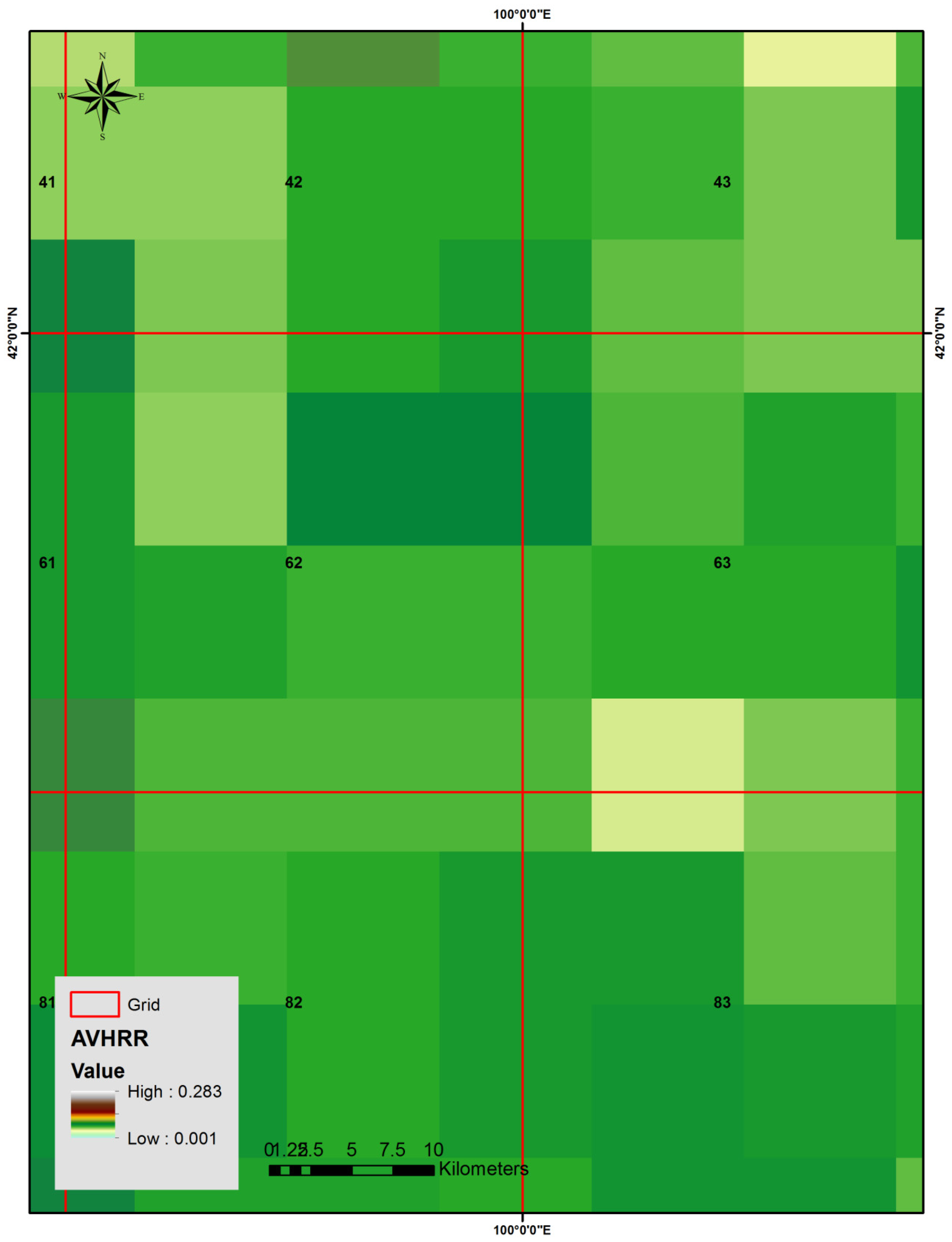
3.2. NOAA/AVHRR-NDVI Data
3.3. Air Temperature Data
3.4. Correlation between Climate Factors and NDVI
3.5. Discrimination of C3 and C4 Vegetation based on Different Methods
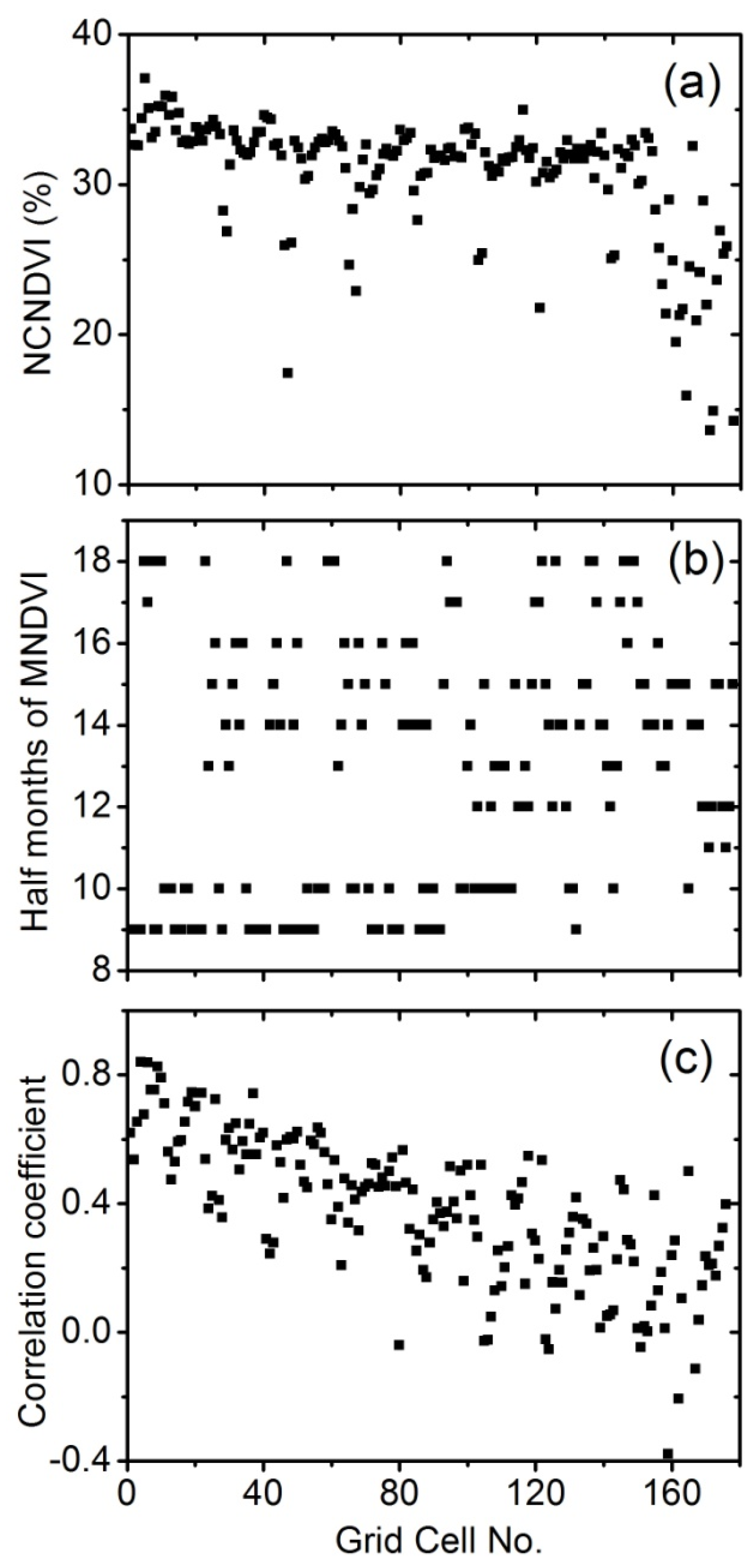
3.5.1. Method 1-Based on Normalized Cumulative NDVI
3.5.2. Method 2-Based on Date of Maximum NDVI

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Half Month No. | Half Month No. | ||
|---|---|---|---|
| 1st half of January | 1 | 1st half of July | 13 |
| 2nd half of January | 2 | 2nd half of July | 14 |
| 1st half of February | 3 | 1st half of August | 15 |
| 2nd half of February | 4 | 2nd half of August | 16 |
| 1st half of March | 5 | 1st half of September | 17 |
| 2nd half of March | 6 | 2nd half of September | 18 |
| 1st half of April | 7 | 1st half of October | 19 |
| 2nd half of April | 8 | 2nd half of October | 20 |
| 1st half of May | 9 | 1st half of November | 21 |
| 2nd half of May | 10 | 2nd half of November | 22 |
| 1st half of June | 11 | 1st half of December | 23 |
| 2nd half of June | 12 | 2nd half of December | 24 |
3.5.3. Method 3-Based on Correlation Between MNDVI and Precipitation
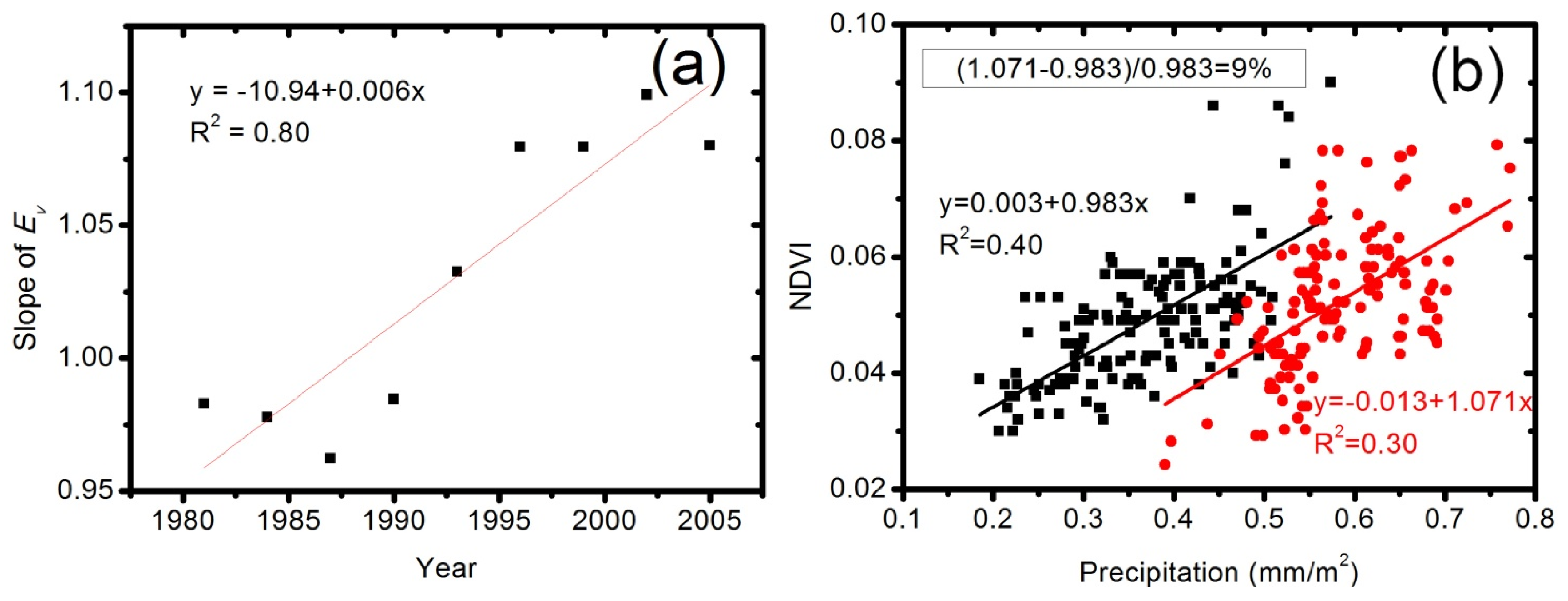
3.6. Quantification of the CO2 Fertilization Effect
4. Results
4.1. Characteristics of Temporal Variations of NDVI
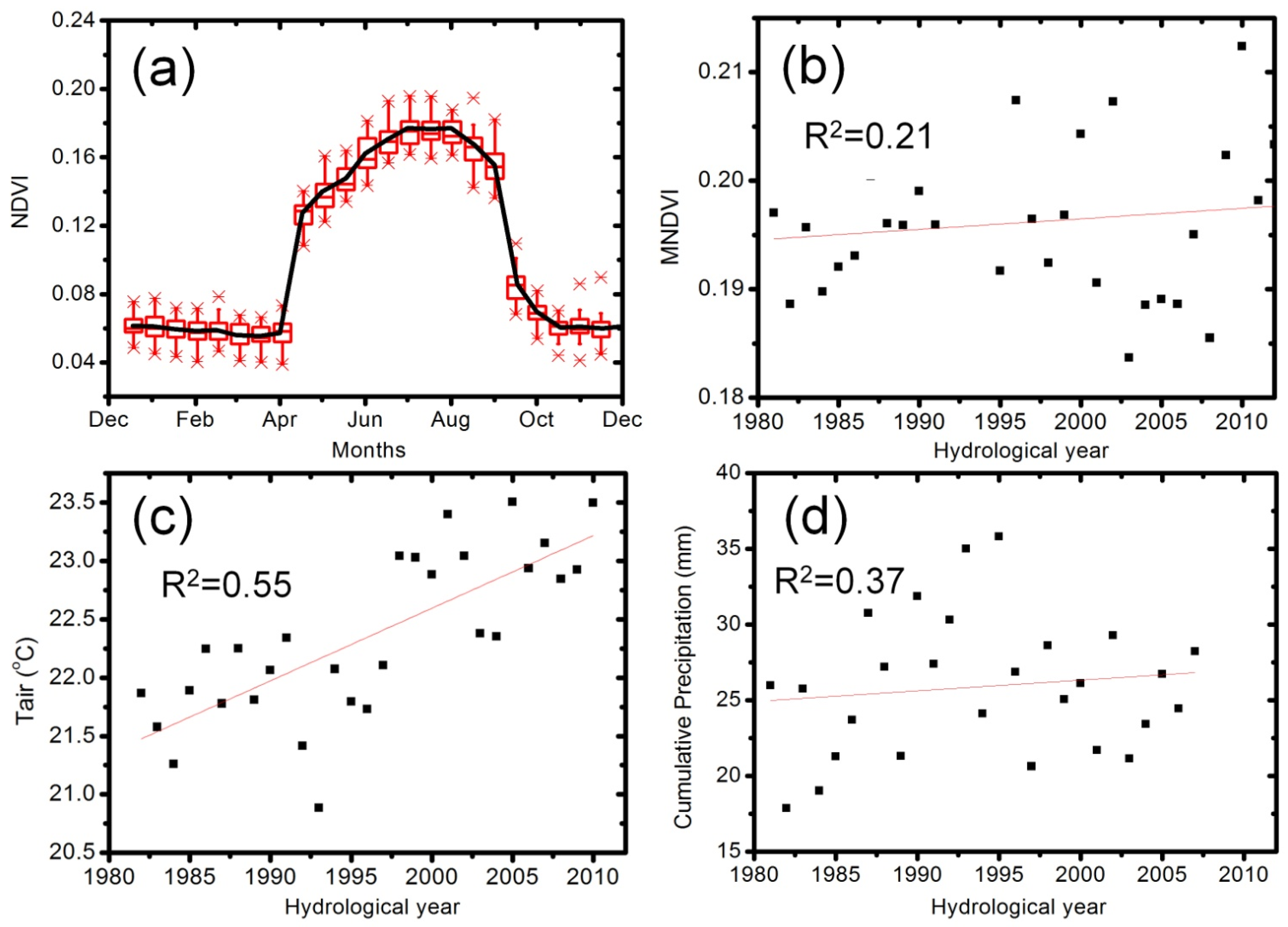
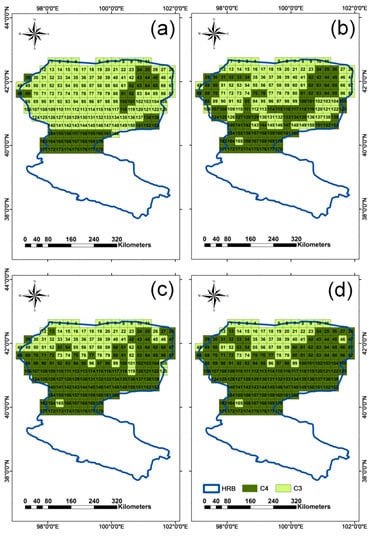
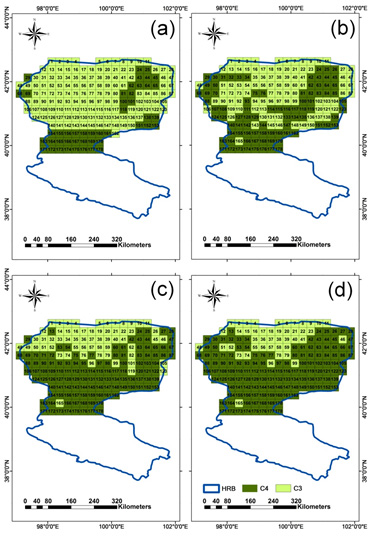
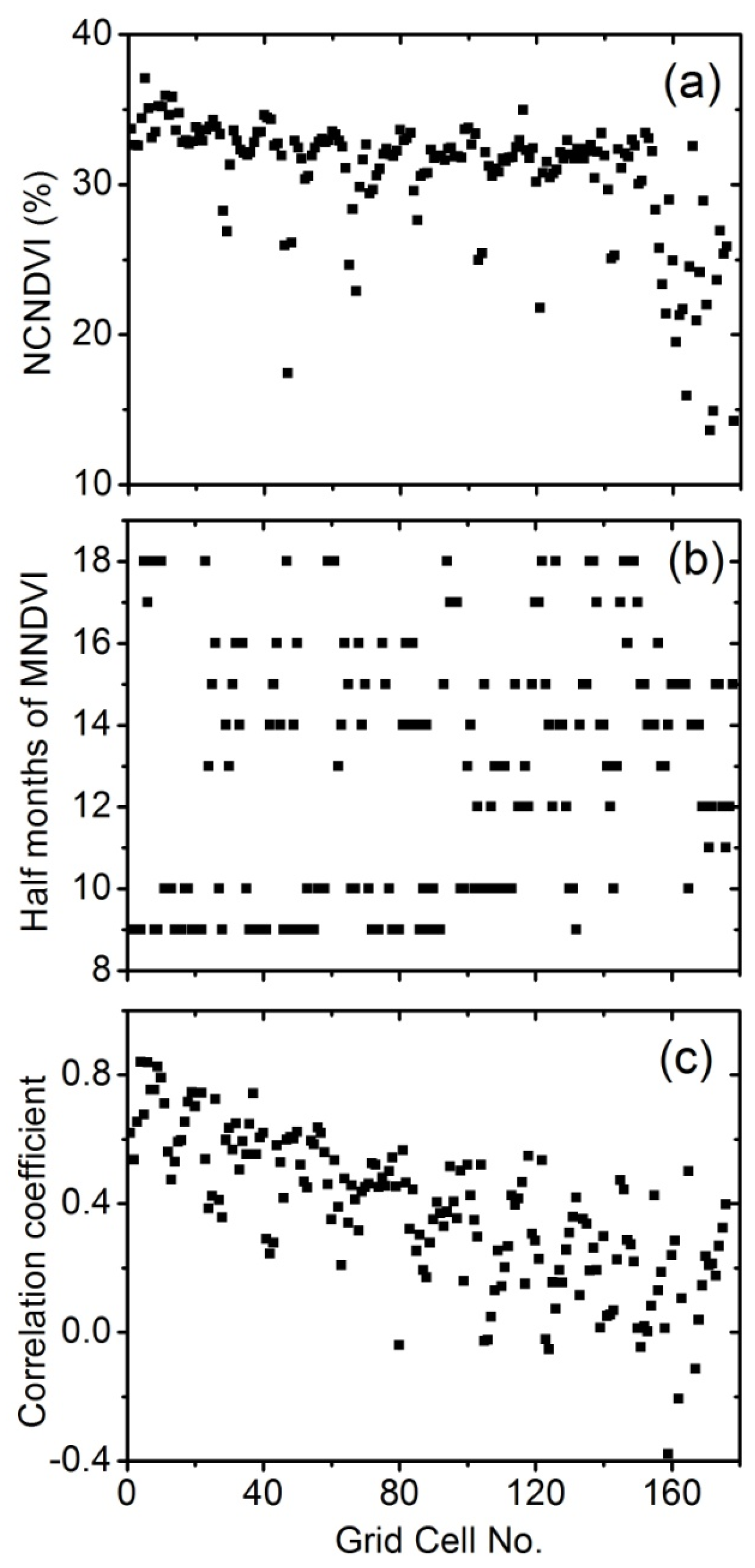
4.2. Discrimination of C3 and C4 Vegetation Based on Three Methods
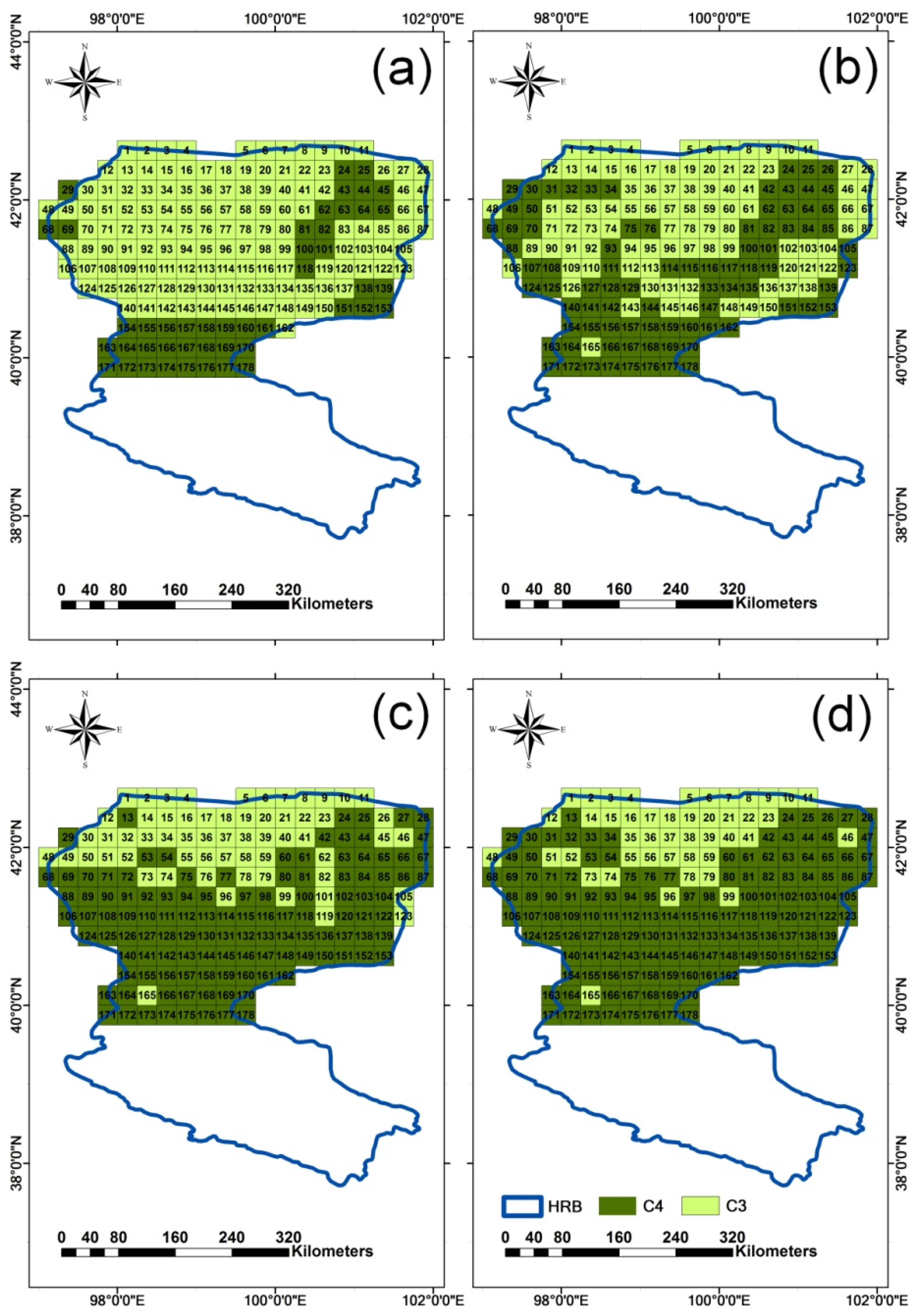
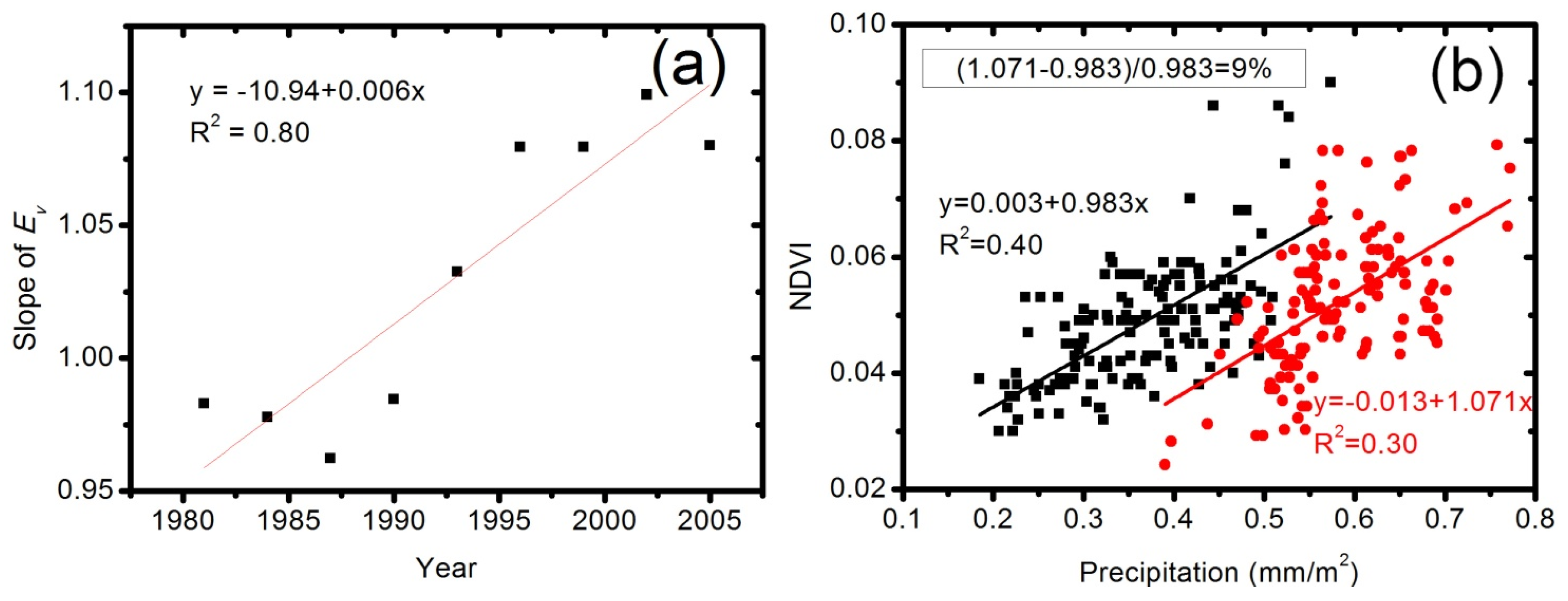
4.3. Quantification of CO2 Fertilization Effect
5. Discussion
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Han, H.; Ma, M.; Yan, P.; Song, Y. Periodicity analysis of NDVI time series and its relationship with climatic factors in the Heihe River Basin in China. Proc. SPIE 2011. [Google Scholar] [CrossRef]
- Deil, U. A review on habitats, plant traits and vegetation of ephemeral wetlands—A global perspective. Phytocoenologia 2005, 35, 533–705. [Google Scholar] [CrossRef]
- Bennett, K.D.; Willis, K.J. Effect of global atmospheric carbon dioxide on glacial-interglacial vegetation change. Glob. Ecol. Biogeogr. 2000, 9, 355–361. [Google Scholar] [CrossRef]
- Kidane, Y.; Stahlmann, R.; Beierkuhnlein, C. Vegetation dynamics, and land use and land cover change in the Bale Mountains, Ethiopia. Environ. Monit. Assess. 2012, 184, 7473–7489. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Piao, S.L.; He, J.S.; Ma, W.H. Increasing terrestrial vegetation activity in China, 1982–1999. Sci. China Ser. C Life Sci. 2004, 47, 229–240. [Google Scholar]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Porporato, A.; Daly, E.; Rodriguez-Iturbe, I. Soil water balance and ecosystem response to climate change. Am. Nat. 2004, 164, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Canon, J.; Dominguez, F.; Valdes, J.B. Vegetation responses to precipitation and temperature: A spatiotemporal analysis of ecoregions in the Colorado River Basin. Int. J. Remote Sens. 2011, 32, 5665–5687. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Port, U.; Brovkin, V.; Claussen, M. The influence of vegetation dynamics on anthropogenic climate change. Earth Syst. Dyn. 2012, 3, 233–243. [Google Scholar] [CrossRef]
- Qiu, L.; Liu, X. Sensitivity analysis of modelled responses of vegetation dynamics on the Tibetan Plateau to doubled CO2 and associated climate change. Theor. Appl. Climatol. 2015, 1–11. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; de Colstoun, E.B.; McMurtrey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Song, X.Y.; Wang, J.H.; Yang, G.J.; Feng, H.K. Winter wheat cropland grain protein content evaluation through remote sensing. Intell. Autom. Soft Comput. 2014, 20, 599–609. [Google Scholar] [CrossRef]
- Wang, X.W.; Xie, H.J.; Guan, H.D.; Zhou, X.B. Different responses of MODIS-derived NDVI to root-zone soil moisture in semi-arid and humid regions. J. Hydrol. 2007, 340, 12–24. [Google Scholar] [CrossRef]
- Schnur, M.T.; Xie, H.J.; Wang, X.W. Estimating root zone soil moisture at distant sites using MODIS NDVI and EVI in a semi-arid region of southwestern USA. Ecol. Inform. 2010, 5, 400–409. [Google Scholar] [CrossRef]
- Murray, S.J.; Foster, P.N.; Prentice, I.C. Future global water resources with respect to climate change and water withdrawals as estimated by a dynamic global vegetation model. J. Hydrol. 2012, 448, 14–29. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef]
- Gessner, U.; Naeimi, V.; Klein, I.; Kuenzer, C.; Klein, D.; Dech, S. The relationship between precipitation anomalies and satellite-derived vegetation activity in Central Asia. Glob. Planet. Chang. 2013, 110, 74–87. [Google Scholar] [CrossRef]
- Barichivich, J.; Briffa, K.R.; Myneni, R.B.; Osborn, T.J.; Melvin, T.M.; Ciais, P.; Piao, S.L.; Tucker, C. Large-scale variations in the vegetation growing season and annual cycle of atmospheric CO2 at high northern latitudes from 1950 to 2011. Glob. Chang. Biol. 2013, 19, 3167–3183. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.Q.; Mao, F.; Sun, H.; Hou, Y.Y. Study of normalized difference vegetation index variation and its correlation with climate factors in the three-river-source region. Int. J. Appl. Earth Observ. Geoinf. 2011, 13, 24–33. [Google Scholar] [CrossRef]
- Meier, G.A.; Brown, J.F. Remote sensing of land surface phenology. US Geol. Surv. 2014. [Google Scholar] [CrossRef]
- Rigge, M.; Smart, A.; Wylie, B.; Gilmanov, T.; Johnson, P. Linking phenology and biomass productivity in South Dakota mixed-grass prairie. Rangel. Ecol. Manag. 2013, 66, 579–587. [Google Scholar] [CrossRef]
- Ichii, K.; Kawabata, A.; Yamaguchi, Y. Global correlation analysis for NDVI and climatic variables and NDVI trends: 1982–1990. Int. J. Remote Sens. 2002, 23, 3873–3878. [Google Scholar] [CrossRef]
- Maselli, F.; di Gregorio, A.; Capecchi, V.; Breda, F. Enrichment of land-cover polygons with eco-climatic information derived from MODIS NDVI imagery. J. Biogeogr. 2009, 36, 639–650. [Google Scholar] [CrossRef]
- Song, Y.; Ma, M.G. A statistical analysis of the relationship between climatic factors and the Normalized Difference Vegetation Index in China. Int. J. Remote Sens. 2011, 32, 3947–3965. [Google Scholar] [CrossRef]
- Beck, H.E.; McVicar, T.R.; van Dijk, A.I.J.M.; Schellekens, J.; de Jeu, R.A.M.; Bruijnzeel, L.A. Global evaluation of four AVHRR-NDVI data sets: Intercomparison and assessment against Landsat imagery. Remote Sens. Environ. 2011, 115, 2547–2563. [Google Scholar] [CrossRef]
- Fensholt, R.; Langanke, T.; Rasmussen, K.; Reenberg, A.; Prince, S.D.; Tucker, C.; Scholes, R.J.; Le, Q.B.; Bondeau, A.; Eastman, R.; et al. Greenness in semi-arid areas across the globe 1981–2007—An Earth Observing Satellite based analysis of trends and drivers. Remote Sens. Environ. 2012, 121, 144–158. [Google Scholar] [CrossRef]
- Houghton, R.A. Revised estimates of the annual net flux of carbon to the atmosphere from changes in land use and land management 1850–2000. Tellus Ser. B Chem. Phys. Meteorol. 2003, 55, 378–390. [Google Scholar] [CrossRef]
- Ghannoum, O. C-4 photosynthesis and water stress. Ann. Bot. 2009, 103, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.E.P.; Moran, M.S.; Huete, A.; Zhang, Y.; Bresloff, C.; Huxman, T.E.; Eamus, D.; Bosch, D.D.; Buda, A.R.; Gunter, S.A.; et al. Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature 2013, 494, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Hogan, C.M. Respiration. In Encyclopedia of Earth; McGinley, M., Cleveland, C.J., Eds.; National Council for Science and the Environment: Washington, DC, USA, 2011. [Google Scholar]
- Foody, G.M.; Dash, J. Discriminating and mapping the C3 and C4 composition of grasslands in the northern Great Plains, USA. Ecol. Inform. 2007, 2, 89–93. [Google Scholar] [CrossRef]
- Ehleringer, J. The influence of atmospheric CO2, temperature, and water on the abundance of C3/C4 taxa. In A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems; Baldwin, I.T., Caldwell, M.M., Heldmaier, G., Jackson, R., Lange, O.L., Mooney, H.A., Schulze, E.D., Sommer, U., Ehleringer, J., Dearing, M., et al., Eds.; Springer: New York, NY, USA, 2005; pp. 214–231. [Google Scholar]
- Kakani, V.G.; Reddy, K.R. Temperature response of C4 species big bluestem (Andropogon gerardii) is modified by growing carbon dioxide concentration. Environ. Exp. Bot. 2007, 61, 281–290. [Google Scholar] [CrossRef]
- Mishra, R.S.; Abdin, M.Z.; Uprety, D.C. Interactive effects of elevated CO2 and moisture stress on the photosynthesis, water relation and growth of Brassica species. J. Agron. Crop Sci. 1999, 182, 223–229. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Yu, D.; Zhang, N.; Lin, J.; Zhang, J.; Tang, J.; Wang, J.; Mu, C. The influence of precipitation regimes and elevated CO2 on photosynthesis and biomass accumulation and partitioning in seedlings of the rhizomatous perennial grass Leymus chinensis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Adjorlolo, C.; Mutanga, O.; Cho, M.A.; Ismail, R. Challenges and opportunities in the use of remote sensing for C-3 and C-4 grass species discrimination and mapping. Afr. J. Range Forage Sci. 2012, 29, 47–61. [Google Scholar] [CrossRef]
- Ricotta, C.; Reed, B.C.; Tieszen, L.T. The role of C-3 and C-4 grasses to interannual variability in remotely sensed ecosystem performance over the US Great Plains. Int. J. Remote Sens. 2003, 24, 4421–4431. [Google Scholar] [CrossRef]
- Goodin, D.G.; Henebry, G.M. A technique for monitoring ecological disturbance in tallgrass prairie using seasonal NDVI trajectories and a discriminant function mixture model. Remote Sens. Environ. 1997, 61, 270–278. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The temperature response of C-3 and C-4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.H.; Li, X.; Lu, L.; Li, Z.Y. Large-scale land cover mapping with the integration of multi-source information based on the Dempster-Shafer theory. Int. J. Geogr. Inf. Sci. 2012, 26, 169–191. [Google Scholar] [CrossRef]
- Zhou, B.; Zheng, H.; Shen, C.; Wei, G.; Li, L. Effect of temperature and precipitation on C3/C4 plants evolutions since late Quaternary in the East Asian Monsoon Area: Evidence from carbon isotopes of alkanes and elemental carbon. AGU Fall Meet. Abstr. 2009, 1, 1394. [Google Scholar]
- Guo, Q.; Feng, Q.; Li, J. Environmental changes after ecological water conveyance in the lower reaches of Heihe River, northwest China. Environ. Geol. 2009, 58, 1387–1396. [Google Scholar] [CrossRef]
- Qin, D.; Zhao, Z.; Han, L.; Qian, Y.; Ou, L.; Wu, Z.; Wang, M. Determination of groundwater recharge regime and flowpath in the Lower Heihe River basin in an arid area of Northwest China by using environmental tracers: Implications for vegetation degradation in the Ejina Oasis. Appl. Geochem. 2012, 27, 1133–1145. [Google Scholar] [CrossRef]
- Yan, H.; Zhan, J.; Liu, B.; Yuan, Y. Model estimation of water use efficiency for soil conservation in the lower Heihe River Basin, Northwest China during 2000–2008. Sustainability 2014, 6, 6250–6266. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Y.; Yu, J.; Fu, G.; Ao, F. Vegetation dynamics induced by groundwater fluctuations in the lower Heihe River Basin, northwestern China. J. Plant Ecol. 2011, 4, 77–90. [Google Scholar] [CrossRef]
- Xi, H.; Feng, Q.; Si, J.; Chang, Z.; Cao, S. Impacts of river recharge on groundwater level and hydrochemistry in the lower reaches of Heihe River Watershed, northwestern China. Hydrogeol. J. 2010, 18, 791–801. [Google Scholar] [CrossRef]
- Su, P.; Yan, Q.; Xie, T.; Zhou, Z.; Gao, S. Associated growth of C3 and C4 desert plants helps the C3 species at the cost of the C4 species. Acta Physiol. Plant. 2012, 34, 2057–2068. [Google Scholar] [CrossRef]
- Su, P.; Yan, Q. Stable carbon isotope variation in plants and their indicating significances along the inland Heihe River basin of northwestern China. Acta Ecol. Sin. 2008, 28, 1616–1624. [Google Scholar]
- Yin, D.; Li, X.; Huang, Y.; Si, Y.; Bai, R. Identifying vegetation dynamics and sensitivities in response to water resources management in the Heihe River Basin in China. Adv. Meteorol. 2015, 2015, 12. [Google Scholar] [CrossRef]
- Wen, X.; Wu, Y.; Su, J.; Zhang, Y.; Liu, F. Hydrochemical characteristics and salinity of groundwater in the Ejina Basin, Northwestern China. Environ. Geol. 2005, 48, 665–675. [Google Scholar] [CrossRef]
- Wang, J.H.; Gai, C.M.; Zhao, J.; Hu, X.L. Landuse/Landcover Data of the Heihe River Basin in 2011; Heihe Plan Science Data Center: Lanzhou, China, 2014. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; el Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Liu, G.; Liu, H.; Yin, Y. Global patterns of NDVI-indicated vegetation extremes and their sensitivity to climate extremes. Environ. Res. Lett. 2013, 8. [Google Scholar] [CrossRef]
- Pinzon, J.E.; Tucker, C.J. A Non-Stationary 1981–2012 AVHRR NDVI3g Time Series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Wong, S.; Cowan, I.; Farquhar, G. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282. [Google Scholar] [CrossRef]
- Tans, P.; Keeling, R. Trends in Atmospheric Carbon Dioxide; National Oceanic & Atmospheric Administration, Earth system Research Laboratory of Global Monitoring Division: Boulder, CO, USA, 2011. [Google Scholar]
- Guan, L.; Liu, L.; Peng, D.; Hu, Y.; Jiao, Q.; Liu, L. Monitoring the distribution of C3 and C4 grasses in a temperate grassland in northern China using moderate resolution imaging spectroradiometer normalized difference vegetation index trajectories. J. Appl. Remote Sens. 2012, 6. [Google Scholar] [CrossRef]
- National Research Council (US); Committee on Climate Uncertainty, and Water Resources Management. Managing Water Resources in the West Under Conditions of Climate Uncertainty: Proceedings of a Colloquium, November 14–16, 1990, Scottsdale, Arizona; National Academies Press: Washington, DC, USA, 1991. [Google Scholar]
- Boelman, N.T.; Stieglitz, M.; Griffin, K.L.; Shaver, G.R. Inter-annual variability of NDVI in response to long-term warming and fertilization in wet sedge and tussock tundra. Oecologia 2005, 143, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Boelman, N.T.; Stieglitz, M.; Rueth, H.M.; Sommerkorn, M.; Griffin, K.L.; Shaver, G.R.; Gamon, J.A. Response of NDVI, biomass, and ecosystem gas exchange to long-term warming and fertilization in wet sedge tundra. Oecologia 2003, 135, 414–421. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, Y.; Xie, H. C3 Vegetation Mapping and CO2 Fertilization Effect in the Arid Lower Heihe River Basin, Northwestern China. Remote Sens. 2015, 7, 16384-16397. https://doi.org/10.3390/rs71215836
Bi Y, Xie H. C3 Vegetation Mapping and CO2 Fertilization Effect in the Arid Lower Heihe River Basin, Northwestern China. Remote Sensing. 2015; 7(12):16384-16397. https://doi.org/10.3390/rs71215836
Chicago/Turabian StyleBi, Yunbo, and Hongjie Xie. 2015. "C3 Vegetation Mapping and CO2 Fertilization Effect in the Arid Lower Heihe River Basin, Northwestern China" Remote Sensing 7, no. 12: 16384-16397. https://doi.org/10.3390/rs71215836
APA StyleBi, Y., & Xie, H. (2015). C3 Vegetation Mapping and CO2 Fertilization Effect in the Arid Lower Heihe River Basin, Northwestern China. Remote Sensing, 7(12), 16384-16397. https://doi.org/10.3390/rs71215836