
Elevation Regimes Modulated the Responses of Canopy Structure of Coastal Mangrove Forests to Hurricane Damage

Abstract
:1. Introduction
2. Materials and Methods
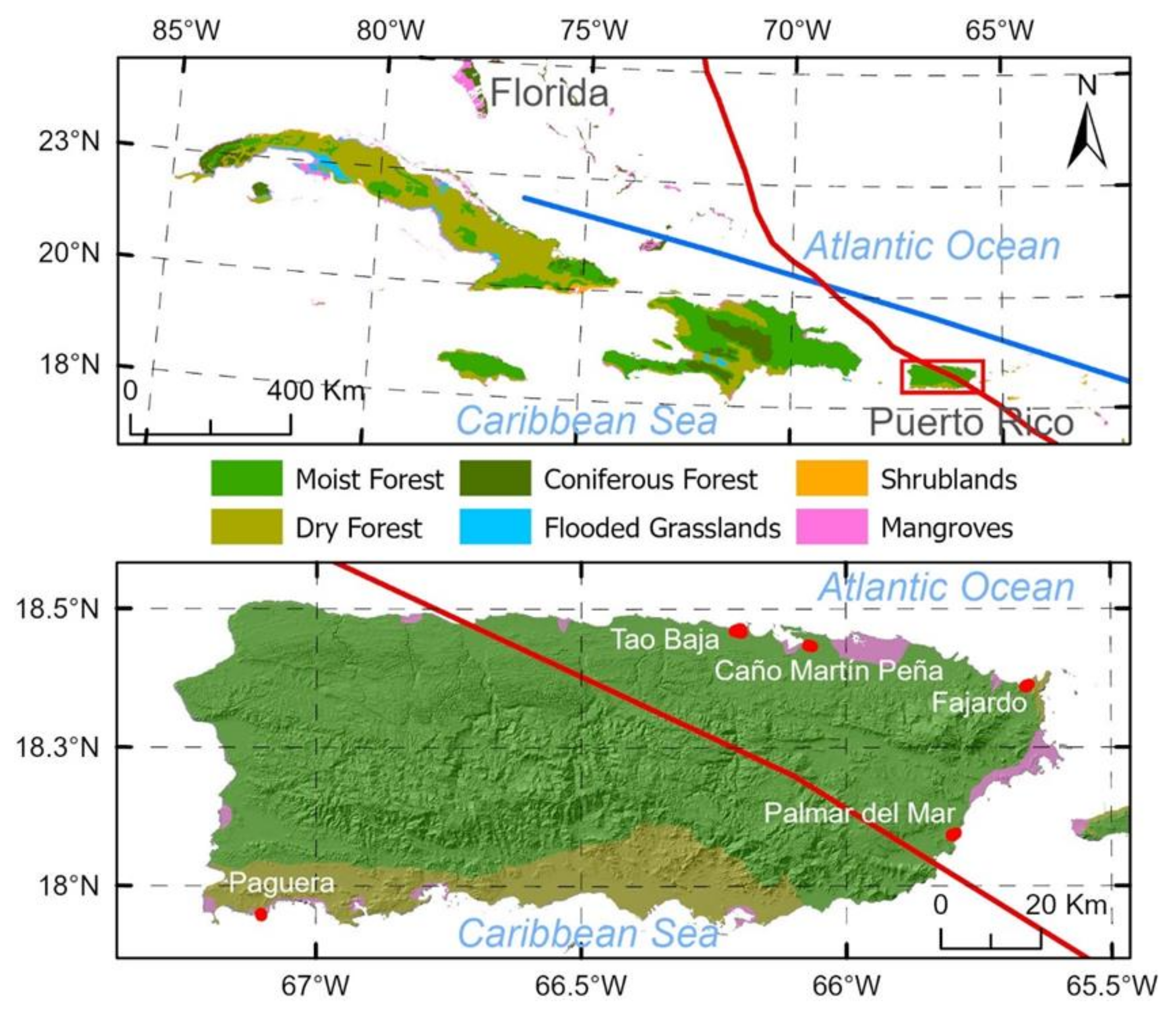
2.1. Site Description
2.2. Data Source
2.3. Analysis of Pre-Hurricane Canopy Height and Canopy Height Reduction by the Hurricanes
3. Results
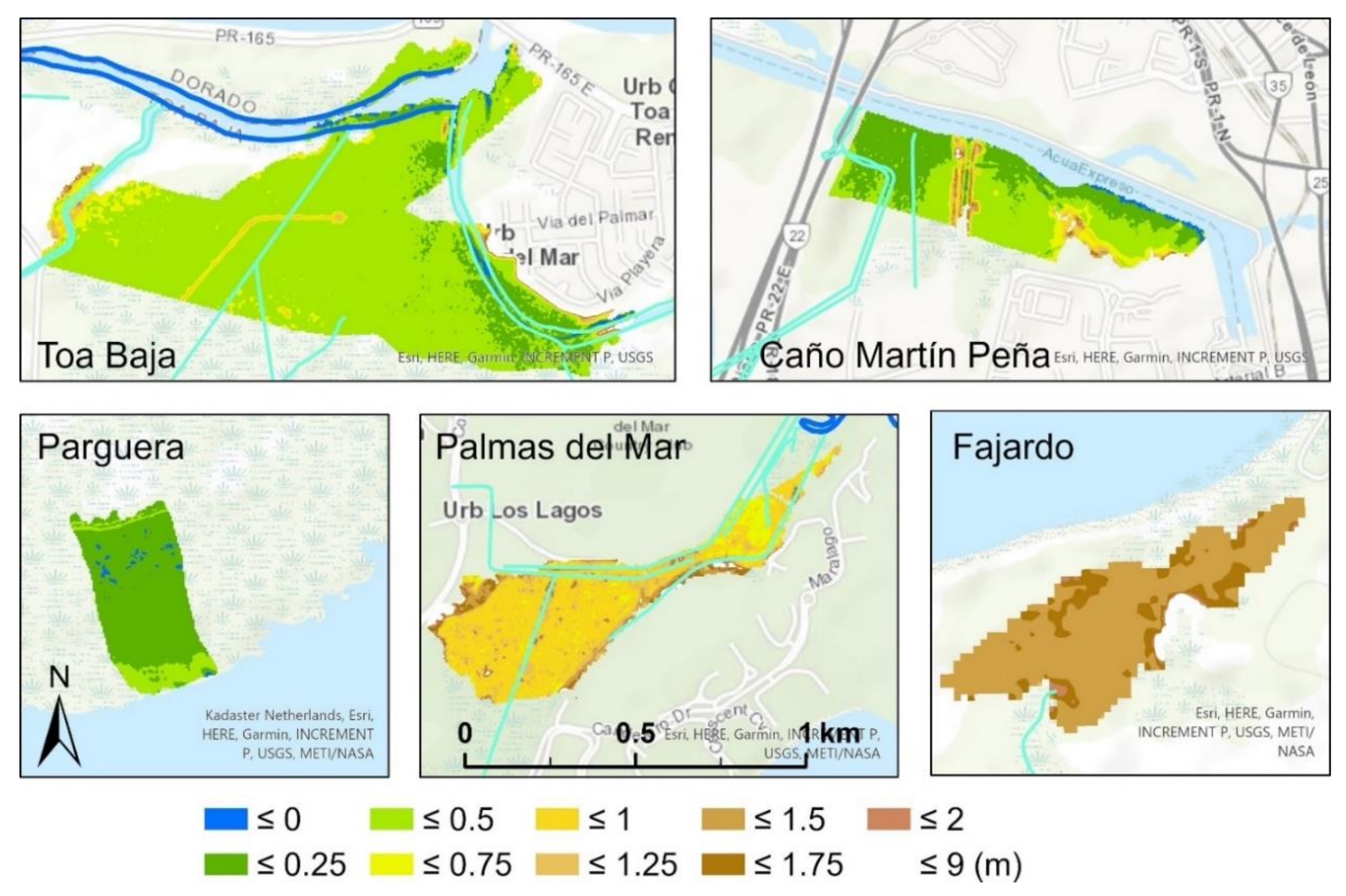
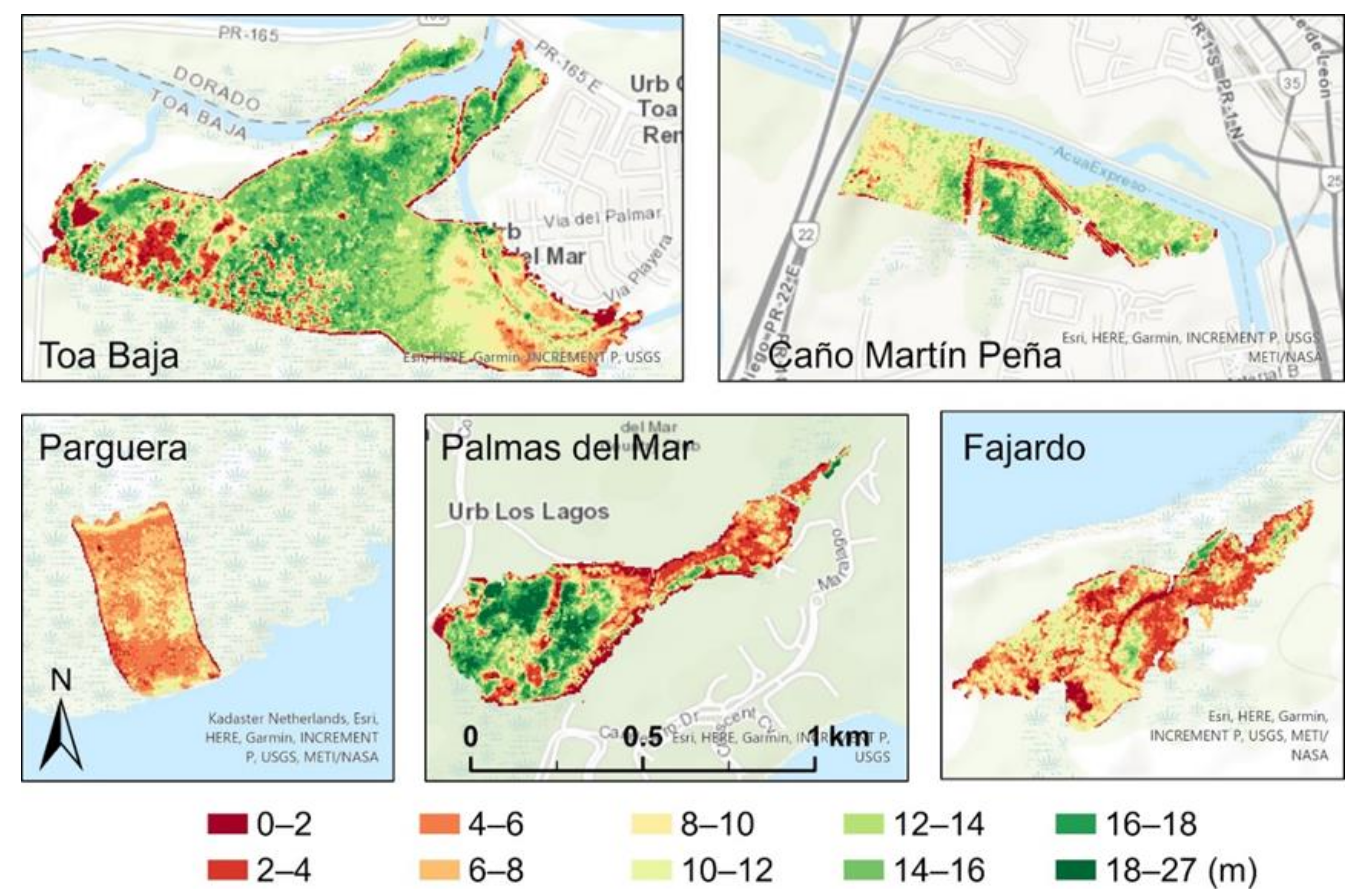
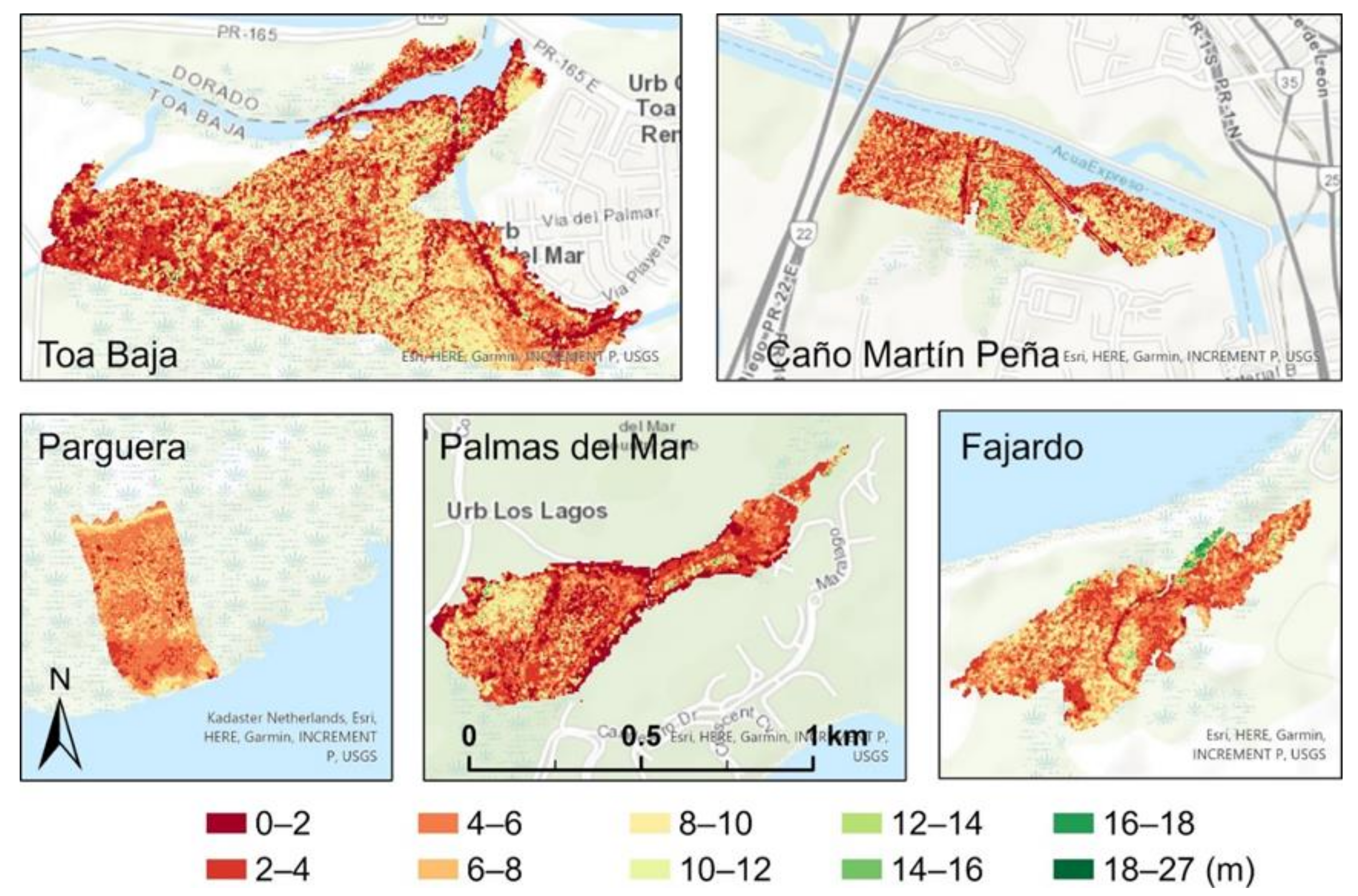
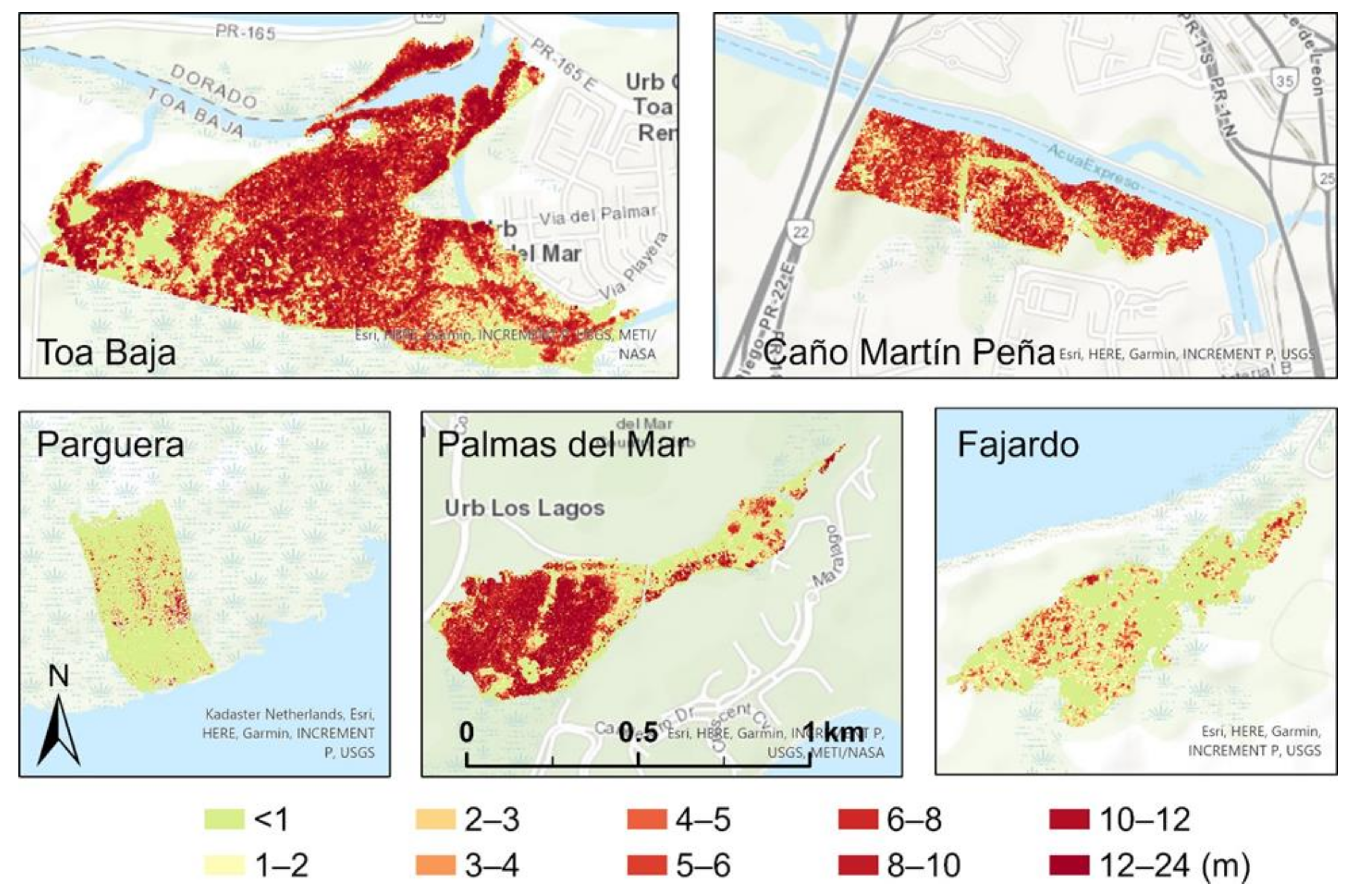
3.1. Spatial Variation and Summary Statistics of the Canopy Heights before and after the Hurricanes
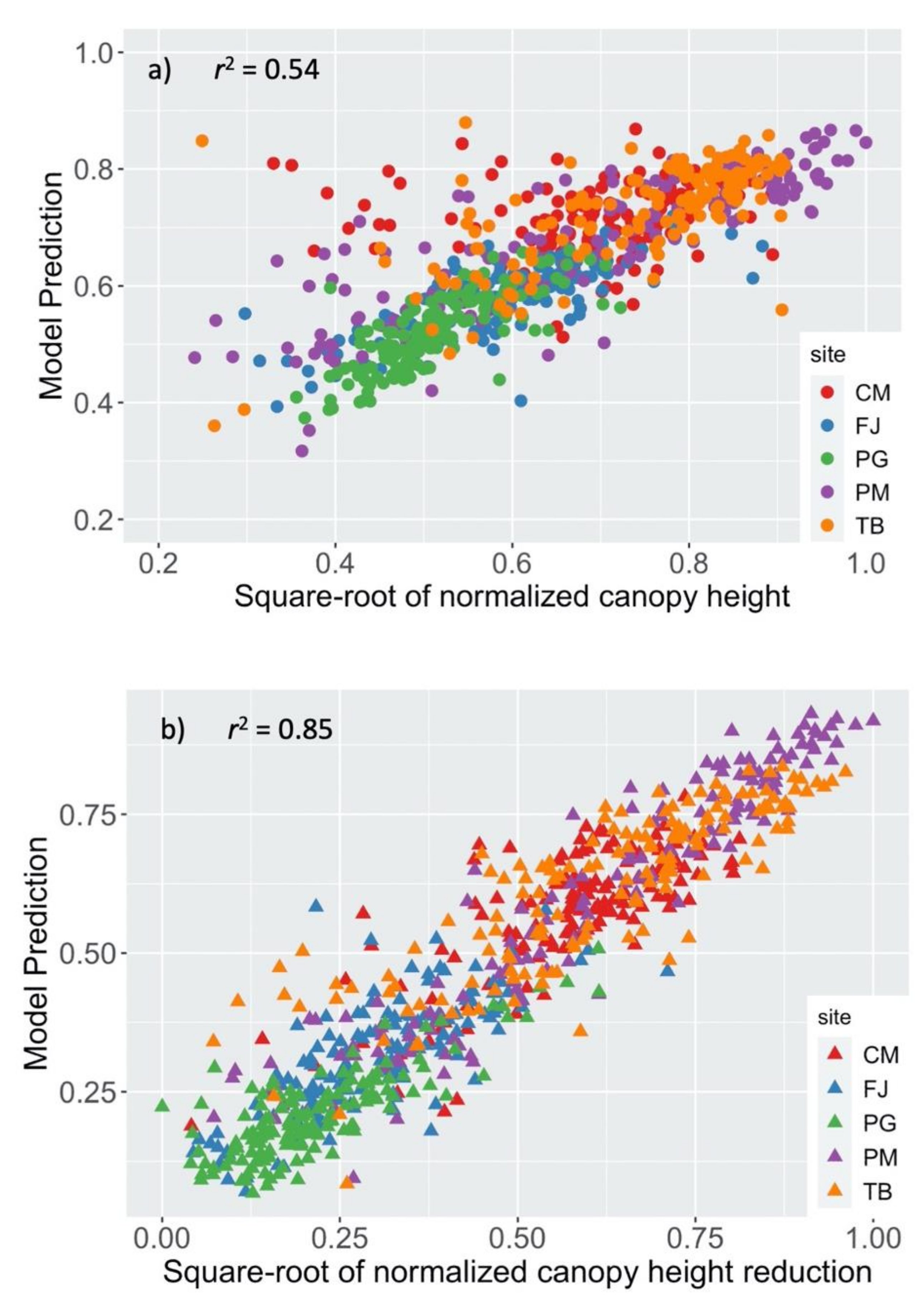
3.2. Spatial Regression Models of the Pre-Hurricane Canopy Height and the Canopy Height Reduction by the Hurricanes
4. Discussion
4.1. Canopy Height before Hurricanes
4.2. Elevation
4.3. Distance to Rivers/Canals and to Coast
4.4. Tree Density and Unevenness of Canopy
4.5. Hurricane Winds and Rainfall
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Alongi, D.M. Carbon Cycling and Storage in Mangrove Forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Atwood, T.B.; Connolly, R.M.; Almahasheer, H.; Carnell, P.E.; Duarte, C.M.; Ewers Lewis, C.J.; Irigoien, X.; Kelleway, J.J.; Lavery, P.S.; Macreadie, P.I.; et al. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Chang. 2017, 7, 523. [Google Scholar] [CrossRef]
- Cannicci, S.; Burrows, D.; Fratini, S.; Smith, T.J.; Offenberg, J.; Dahdouh-Guebas, F. Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat. Bot. 2008, 89, 186–200. [Google Scholar] [CrossRef]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How mangrove forests adjust to rising sea level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Rivera-Ocasio, E.; Heartsill-Scalley, T.; Davila-Casanova, D.; Rios-López, N.; Gao, Q. Landscape-Level Consequences of Rising Sea-Level on Coastal Wetlands: Saltwater Intrusion Drives Displacement and Mortality in the Twenty-First Century. Wetlands 2019, 39, 1343–1355. [Google Scholar] [CrossRef]
- Badola, R.; Hussain, S.A. Valuing ecosystem functions: An empirical study on the storm protection function of Bhitarkanika mangrove ecosystem, India. Environ. Conserv. 2005, 32, 85–92. [Google Scholar] [CrossRef]
- Cartier, K. Hurricanes Hit Puerto Rico’s Mangroves Harder Than Florida’s. EOS 2019, 100. [Google Scholar] [CrossRef]
- Duryea, M.L.; Kamp, E. Wind and Trees: Lessons Learned from Hurricanes; School of Forest Resources and Conservation, University of Florida FAS Extension: Gainesville, FL, USA, 2017. [Google Scholar]
- Smith, T.J.; Anderson, G.H.; Balentine, K.; Tiling, G.; Ward, G.A.; Whelan, K.R.T. Cumulative impacts of hurricanes on Florida mangrove ecosystems: Sediment deposition, storm surges and vegetation. Wetlands 2009, 29, 24. [Google Scholar] [CrossRef]
- Gardiner, B.; Byrne, K.; Hale, S.; Kamimura, K.; Mitchell, S.J.; Peltola, H.; Ruel, J.-C. A review of mechanistic modelling of wind damage risk to forests. For. Int. J. For. Res. 2008, 81, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. For. Int. J. For. Res. 2012, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Ye, F.; Huang, W.; Zhang, Y.J.; Moghimi, S.; Myers, E.; Pe'eri, S.; Yu, H.C. A cross-scale study for compound flooding processes during Hurricane Florence. Nat. Hazards Earth Syst. Sci. 2021, 21, 1703–1719. [Google Scholar] [CrossRef]
- Choy, S.C.; Booth, W.E. Prolonged inundation and ecological changes in an Avicennia mangrove: Implications for conservation and management. Hydrobiologia 1994, 285, 237–247. [Google Scholar] [CrossRef]
- Krauss, K.W.; Osland, M.J. Tropical cyclones and the organization of mangrove forests: A review. Ann. Bot. 2019, 125, 213–234. [Google Scholar] [CrossRef] [PubMed]
- Lovelock, C.E.; Krauss, K.W.; Osland, M.J.; Reef, R.; Ball, M.C. The Physiology of Mangrove Trees with Changing Climate. In Tropical Tree Physiology: Adaptations and Responses in a Changing Environment; Goldstein, G., Santiago, L.S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 149–179. [Google Scholar] [CrossRef]
- Peel, J.R.; Sanchez, M.C.M.; Lopez-Portillo, J.; Golubov, J. Stomatal density, leaf area and plant size variation of Rhizophora mangle (Malpighiales: Rhizophoraceae) along a salinity gradient in the Mexican Caribbean. Rev. Biol. Trop. 2017, 65, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Kodikara, K.A.S.; Jayatissa, L.P.; Huxham, M.; Dahdouh-Guebas, F.; Koedam, N. The effects of salinity on growth and survival of mangrove seedlings changes with age. Acta Bot. Bras. 2018, 32, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Waisel, Y.; Eshel, A.; Agami, M. Salt balance of leaves of the mangrove Avicennia marina. Physiol. Plant. 2006, 67, 67–72. [Google Scholar] [CrossRef]
- Reef, R.; Lovelock, C.E. Regulation of water balance in mangroves. Ann.Bot. 2015, 115, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Lugo, A.E.; Medina, E. Mangrove Forests. In Encyclopedia of Natural Resources–Land; Wang, Y., Ed.; Taylor & Francis Group: New York, NY, USA, 2014; Volume 1. [Google Scholar]
- Branoff, B. Urban Mangrove Biology and Ecology: Emergent Patterns and Management Implications; University of Puerto Rico: San Juan, PR, USA, 2018. [Google Scholar]
- Martinuzzi, S.; Gould, W.A.; Lugo, A.E.; Medina, E. Conversion and recovery of Puerto Rican mangroves: 200 years of change. For. Ecol. Manag. 2009, 257, 75–84. [Google Scholar] [CrossRef]
- Lugo, A.E.; Medina, E.; Cuevas, E.; Cintrón, G.; Nieves, E.N.L.; Novelli, Y.S. Ecophysiology of a Mangrove Forest in Jobos Bay, Puerto Rico. Caribb. J. Sci. 2007, 43, 200–219. [Google Scholar] [CrossRef] [Green Version]
- Quadros, A.F.; Zimmer, M. Dataset of “true mangroves” plant species traits. Biodivers. Data J. 2017, 5, e22089. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L.; Lugo, A.E. Guide to the Ecological Systems of Puerto Rico; U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry: San Juan, PR, USA, 2009; p. 437. [Google Scholar]
- Castañeda-Moya, E.; Rivera-Monroy, V.H.; Chambers, R.M.; Zhao, X.; Lamb-Wotton, L.; Gorsky, A.; Gaiser, E.E.; Troxler, T.G.; Kominoski, J.S.; Hiatt, M. Hurricanes fertilize mangrove forests in the Gulf of Mexico (Florida Everglades, USA). Proc. Natl. Acad. Sci. USA 2020, 117, 4831–4841. [Google Scholar] [CrossRef]
- Taillie, P.J.; Roman-Cuesta, R.; Lagomasino, D.; Cifuentes-Jara, M.; Fatoyinbo, T.; Ott, L.E.; Poulter, B. Widespread mangrove damage resulting from the 2017 Atlantic mega hurricane season. Environ. Res. Lett. 2020, 15, 064010. [Google Scholar] [CrossRef]
- Branoff, B.; Martinuzzi, S. Mangrove forest structure and composition along urban gradients in Puerto Rico. bioRxiv 2018, 504928. [Google Scholar] [CrossRef] [Green Version]
- Field, C.; Osborn, J.; Hoffman, L.; Polsenberg, J.; Ackerly, D.; Berry, J.; BjÖRkman, O.; Held, A.; Matson, P.; Mooney, H. Mangrove biodiversity and ecosystem function. Glob. Ecol. Biogeogr. Lett. 1998, 7, 3–14. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Yu, M.; Gao, Q. Topography, drainage capability, and legacy of drought differentiate tropical ecosystem response to and recovery from major hurricanes. Environ. Res. Lett. 2020, 15, 104046. [Google Scholar] [CrossRef]
- Huang, W.; Sun, G.; Dubayah, R.; Cook, B.; Montesano, P.; Ni, W.; Zhang, Z. Mapping biomass change after forest disturbance: Applying LiDAR footprint-derived models at key map scales. Remote Sens. Environ. 2013, 134, 319–332. [Google Scholar] [CrossRef]
- Eisemann, E.; Dunkin, L.; Hartman, M.; Wozencraft, J. JALBTCX/NCMP emergency-response airborne Lidar coastal mapping & quick response data products for 2016/2017/2018 hurricane impact assessments. Shore Beach 2019, 87, 31–40. [Google Scholar] [CrossRef]
- Gao, Q.; Yu, M. Elevation and Distribution of Freshwater and Sewage Canals Regulate Canopy Structure and Differentiate Hurricane Damages to a Basin Mangrove Forest. Remote Sens. 2021, 13, 3387. [Google Scholar] [CrossRef]
- Kennaway, T.; Helmer, E.H. The forest types and ages cleared for land development in Puerto Rico. Gisci. Remote Sens. 2007, 44, 356–382. [Google Scholar] [CrossRef]
- Gao, Q.; Yu, M. Discerning Fragmentation Dynamics of Tropical Forest and Wetland during Reforestation, Urban Sprawl, and Policy Shifts. Plos ONE 2014, 9, e113140. [Google Scholar] [CrossRef]
- Cook, B.; Corp, L.; Nelson, R.; Middleton, E.; Morton, D.; McCorkel, J.; Masek, J.; Ranson, K.; Ly, V.; Montesano, P. NASA Goddard’s LiDAR, Hyperspectral and Thermal (G-LiHT) Airborne Imager. Remote Sens. 2013, 5, 4045–4066. [Google Scholar] [CrossRef] [Green Version]
- Office for Coastal Management. C-CAP Land Cover, Puerto Rico. 2010. Available online: https://www.fisheries.noaa.gov/inport/item/48301 (accessed on 15 March 2022).
- Roussel, J.-R.; Auty, D.; Boissieu, F.D.; Meador, A.S. lidR: Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. 2019. Available online: https://cran.r-project.org/package=lidR (accessed on 24 August 2021).
- Pasch, R.J.; Penny, A.B.; Berg, R. National Hurricane Center Tropical Cyclone Report–Hurricane Maria (AL152017) September 16–30, 2017; National Hurricane Center: Miami, FL, USA, 2019; pp. 1–48. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 March 2022).
- Angelou, N.; Dellwik, E.; Mann, J. Wind load estimation on an open-grown European oak tree. For. Int. J. For. Res. 2019, 92, 381–392. [Google Scholar] [CrossRef]
- Peterson, C.J.; Ribeiro, G.H.P.d.M.; Negrón-Juárez, R.; Marra, D.M.; Chambers, J.Q.; Higuchi, N.; Lima, A.; Cannon, J.B. Critical wind speeds suggest wind could be an important disturbance agent in Amazonian forests. For. Int. J. For. Res. 2019, 92, 444–459. [Google Scholar] [CrossRef]
- Wang, H.; Hsieh, Y.P.; Harwell, M.A.; Huang, W. Modeling soil salinity distribution along topographic gradients in tidal salt marshes in Atlantic and Gulf coastal regions. Ecol. Model. 2007, 201, 429–439. [Google Scholar] [CrossRef]
- Jiang, J.; Gao, D.; DeAngelis, D.L. Towards a theory of ecotone resilience: Coastal vegetation on a salinity gradient. Theor. Popul. Biol. 2012, 82, 29–37. [Google Scholar] [CrossRef]
- Chen, Y.; Ye, Y. Effects of Salinity and Nutrient Addition on Mangrove Excoecaria agallocha. PLoS ONE 2014, 9, e93337. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Stanton, D.E.; Schmitz, N.; Farquhar, G.D.; Ball, M.C. Growth responses of the mangrove Avicennia marina to salinity: Development and function of shoot hydraulic systems require saline conditions. Ann. Bot. 2015, 115, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.; Vovides, A.G.; Luna, S.; Gruters, U.; Berger, U. Changes in allometric relations of mangrove trees due to resource availability—A new mechanistic modelling approach. Ecol. Model. 2014, 283, 53–61. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. Leaf Life-Span in Relation to Leaf, Plant, and Stand Characteristics among Diverse Ecosystems. Ecol. Monogr. 1992, 62, 365–392. [Google Scholar] [CrossRef]
- Yang, S.-C.; Shih, S.-S.; Hwang, G.-W.; Adams, J.B.; Lee, H.-Y.; Chen, C.-P. The salinity gradient influences on the inundation tolerance thresholds of mangrove forests. Ecol. Eng. 2013, 51, 59–65. [Google Scholar] [CrossRef]
- Talkkari, A.; Peltola, H.; Kellomäki, S.; Strandman, H. Integration of component models from the tree, stand and regional levels to assess the risk of wind damage at forest margins. For. Ecol. Manag. 2000, 135, 303–313. [Google Scholar] [CrossRef]
- Jimenez-Rodríguez, D.L.; Alvarez-Añorve, M.Y.; Pineda-Cortes, M.; Flores-Puerto, J.I.; Benítez-Malvido, J.; Oyama, K.; Avila-Cabadilla, L.D. Structural and functional traits predict short term response of tropical dry forests to a high intensity hurricane. For. Ecol. Manag. 2018, 426, 101–114. [Google Scholar] [CrossRef]
- Hale, S.E.; Gardiner, B.; Peace, A.; Nicoll, B.; Taylor, P.; Pizzirani, S. Comparison and validation of three versions of a forest wind risk model. Environ. Model. Softw. 2015, 68, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Krogstadt, P.Å.; Antonia, R.A. Surface roughness effects in turbulent boundary layers. Exp. Fluids 1999, 27, 450–460. [Google Scholar] [CrossRef]
- Patterson, P.L.; Healey, S.P.; Ståhl, G.; Saarela, S.; Holm, S.; Andersen, H.-E.; Dubayah, R.O.; Duncanson, L.; Hancock, S.; Armston, J.; et al. Statistical properties of hybrid estimators proposed for GEDI—NASA’s global ecosystem dynamics investigation. Environ. Res. Lett. 2019, 14, 065007. [Google Scholar] [CrossRef]
- Monmany Garzia, A.C.; Yu, M.; Zimmerman, J.K. Effects of vegetation structure and landscape complexity on insect parasitism across an agricultural frontier in Argentina. Basic Appl. Ecol. 2018, 29, 69–78. [Google Scholar] [CrossRef]
- Mccormack, J.J.; Cotoras, D.D. Beetle Diversity Across Micro-habitats on Lizard Island Group (Great Barrier Reef, Australia). Zool. Stud. 2021, 60, 12. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Pre-Hurricane GLiHT Transects | Post-Hurricane USGS Tiles |
|---|---|---|
| Caño Martín Peña | PR_10March2017_118 | E_2018_19QHA37506600, E_2018_19QHA39006600 |
| Fajardo | PR_15March2017_75 | H_2018_20QKF81005700, H_2018_20QKF81005850 |
| Palma del Mar | PR_12March2017_58a, PR_12March2017_58b | H_2018_20QKF66002850, H_2018_20QKF67502700, H_2018_20QKF67502850 |
| La Parguera | PR_8March2017_114 | A_2018_19QFV28001200 |
| Toa Baja | PR_10March2017_117, PR_17March2017_199 | G_2018_19QGA22506750, G_2018_19QGA22506900, G_2018_19QGA24006900, G_2018_19QGA25506900 |
| Site | Toa Baja | Caño Martín Peña | La Parguera | Palma del Mar | Fajardo |
|---|---|---|---|---|---|
| Area (ha) | 71.1 | 22.8 | 13.9 | 24.0 | 22.0 |
| CHM_2017 (m) | 11.7 ± 4.3 | 10.8 ± 3.7 | 6.2 ± 1.8 | 9.3 ± 5.6 | 6.7 ± 2.8 |
| CHM_2018 (m) | 5.5 ± 3.1 | 6.1 ± 3.3 | 5.7 ± 1.1 | 4.2 ± 2.2 | 6.0 ± 2.2 |
| Reduction (m) | 6.1 ± 4.4 (52%) | 4.8 ± 3.4 (44%) | 0.5 ± 1.4 (8%) | 5.1 ± 4.2 (55%) | 0.7 ± 1.8 (10%) |
| Tree density (ha−1) | 2132 ± 938 | 2135 ± 863 | 1674 ± 1410 | 1682 ± 718 | 1988 ± 585 |
| DEM (m) | 0.39 ± 0.31 | 0.36 ± 0.32 | 0.16 ± 0.07 | 1.03 ± 0.38 | 1.61 ± 1.07 |
| To river/canal (m) | 341.5 ± 173.6 | 294.9 ± 201.0 | 948.1 ± 157.9 | 57.8 ± 55.3 | 83.8 ± 60.8 |
| To coast(m) | 516.9 ± 209.2 | 101.2 ± 67.7 | 247.9 ± 151.4 | 710.3 ± 188.0 | 291.1 ± 98.5 |
| Gust speed (m·s−1) | 44.7 | 45.0 | 34.1 | 49.5 | 44.3 |
| Rainfall (mm) | 497.2 | 427.8 | 91.1 | 286.5 | 148.2 |
| Site | Intercept | z | z2 | d | sc | λ | r2 |
|---|---|---|---|---|---|---|---|
| Toa Baja | 0.294 | 0.377 | −0.490 | 0.700 | −0.080 | 0.79 | 0.80 |
| Caño Martín Peña | 0.585 | 0.539 | −1.191 | −0.212 | 0.87 | 0.43 | |
| La Parguera | 0.183 | 0.762 | −0.623 | −0.295 | 0.91 | 0.62 | |
| Palma del Mar | 0.803 | −0.785 | 0.467 | −0.646 | 0.90 | 0.68 | |
| Fajardo | 0.684 | −0.684 | 0.64 | 0.58 |
| Site | Intercept | h | h2 | z | z2 | d | sh | s | sc | λ | r2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Toa Baja | 0.097 | 0.331 | 0.536 | −0.426 | 0.360 | 0.112 | −0.126 | 0.75 | 0.78 | ||
| Caño Martín Peña | 0.182 | 1.101 | −0.328 | −0.768 | 0.947 | −0.088 | −0.167 | 0.69 | 0.60 | ||
| La Parguera | 0.267 | −0.451 | 1.076 | −0.316 | 0.75 | 0.69 | |||||
| Palma del Mar | 0.022 | 0.884 | 0.161 | −0.151 | 0.84 | 0.91 | |||||
| Fajardo | 0.166 | 0.440 | −0.238 | −0.255 | 0.54 | 0.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Q.; Yu, M. Elevation Regimes Modulated the Responses of Canopy Structure of Coastal Mangrove Forests to Hurricane Damage. Remote Sens. 2022, 14, 1497. https://doi.org/10.3390/rs14061497
Gao Q, Yu M. Elevation Regimes Modulated the Responses of Canopy Structure of Coastal Mangrove Forests to Hurricane Damage. Remote Sensing. 2022; 14(6):1497. https://doi.org/10.3390/rs14061497
Chicago/Turabian StyleGao, Qiong, and Mei Yu. 2022. "Elevation Regimes Modulated the Responses of Canopy Structure of Coastal Mangrove Forests to Hurricane Damage" Remote Sensing 14, no. 6: 1497. https://doi.org/10.3390/rs14061497
APA StyleGao, Q., & Yu, M. (2022). Elevation Regimes Modulated the Responses of Canopy Structure of Coastal Mangrove Forests to Hurricane Damage. Remote Sensing, 14(6), 1497. https://doi.org/10.3390/rs14061497