
Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City


Abstract
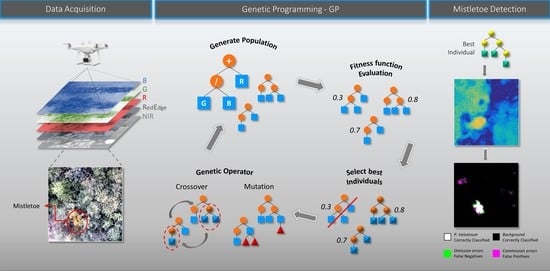
:
1. Introduction
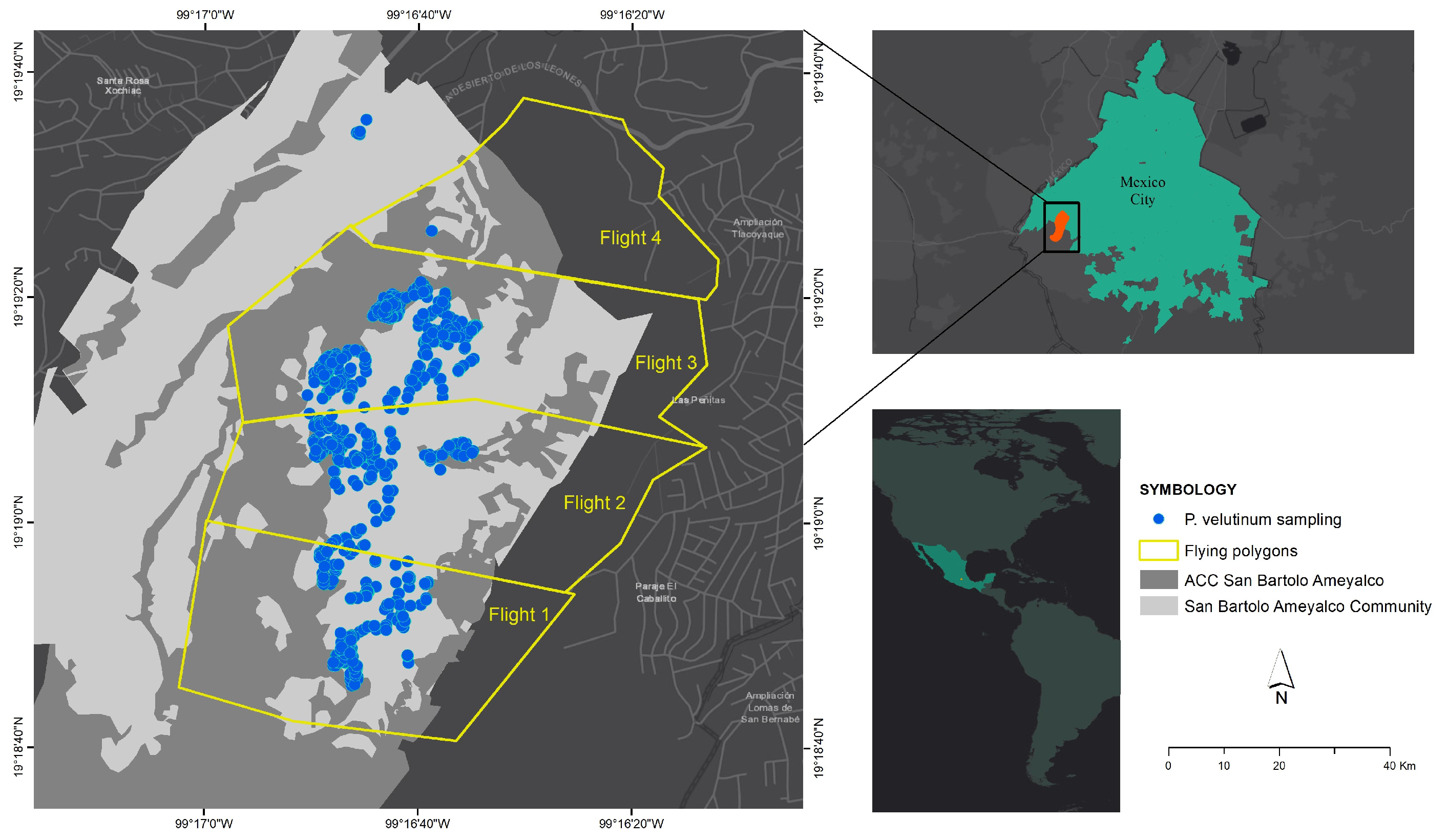
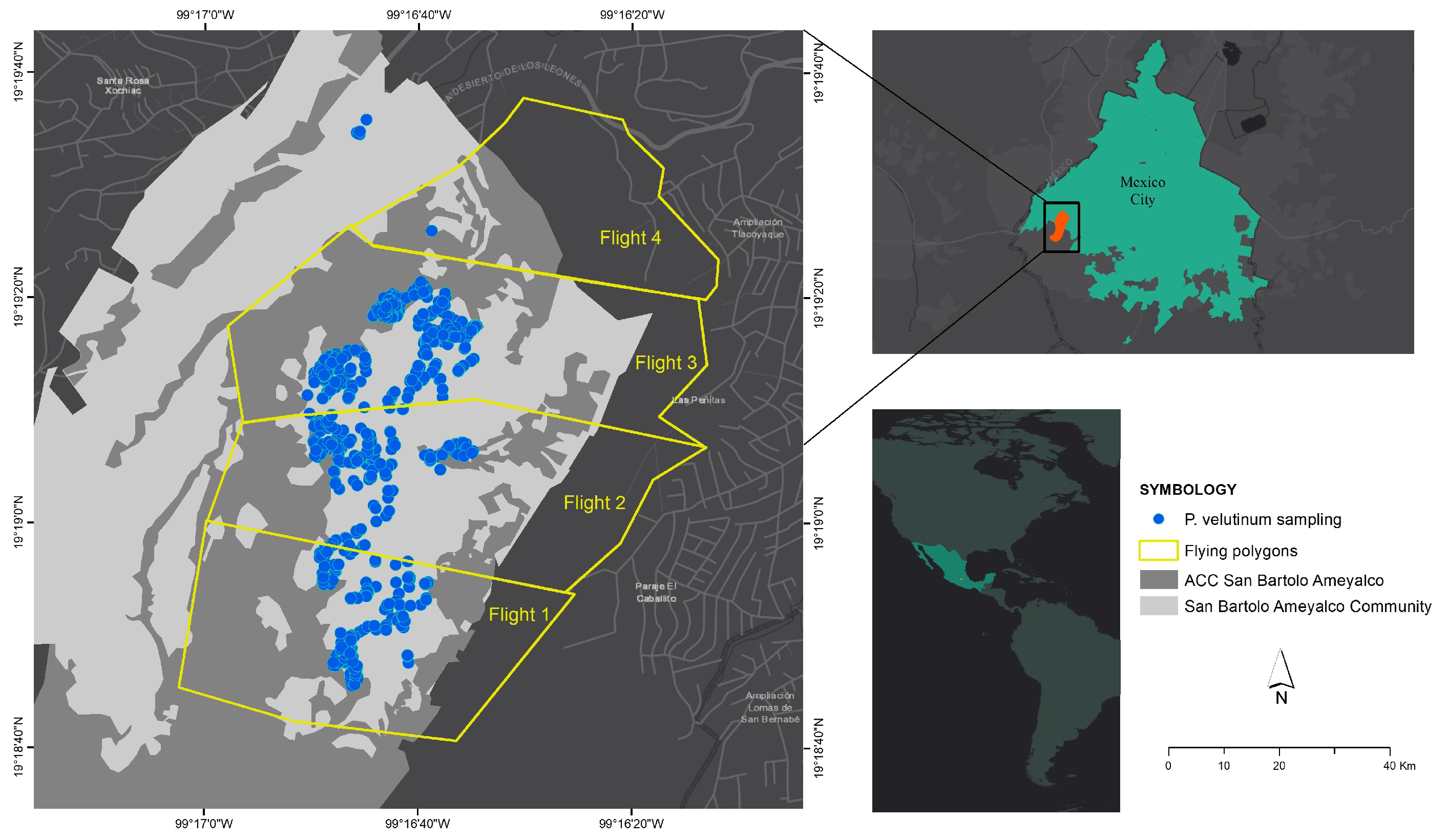
2. Study Area
3. Materials and Methods
3.1. Multispectral Image Collection
3.2. Hardware and Software
3.3. Image Preprocessing
3.3.1. Image Registration
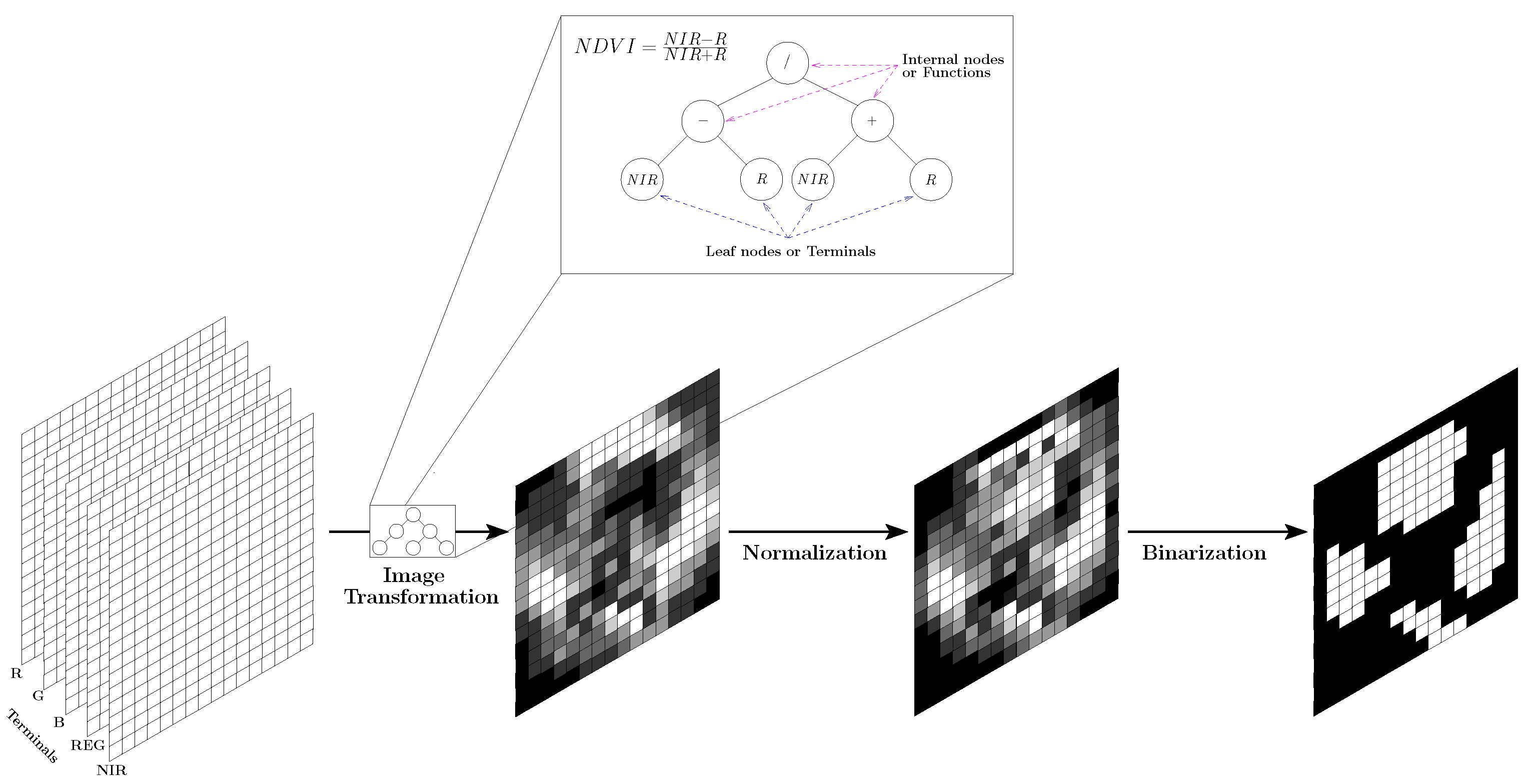
3.3.2. Mask and Data Set Creation
3.4. Algorithm to Detect Mistletoe
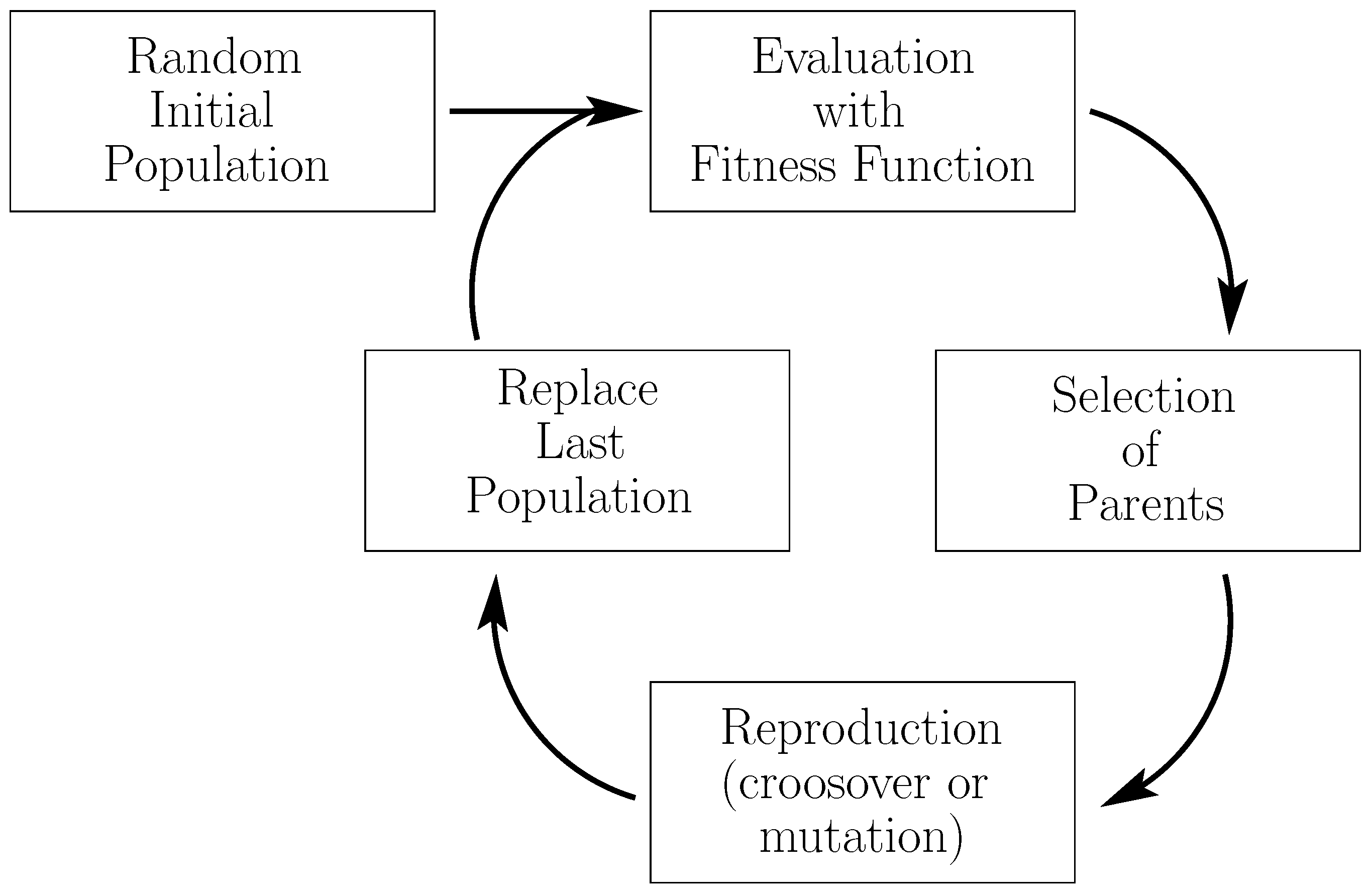
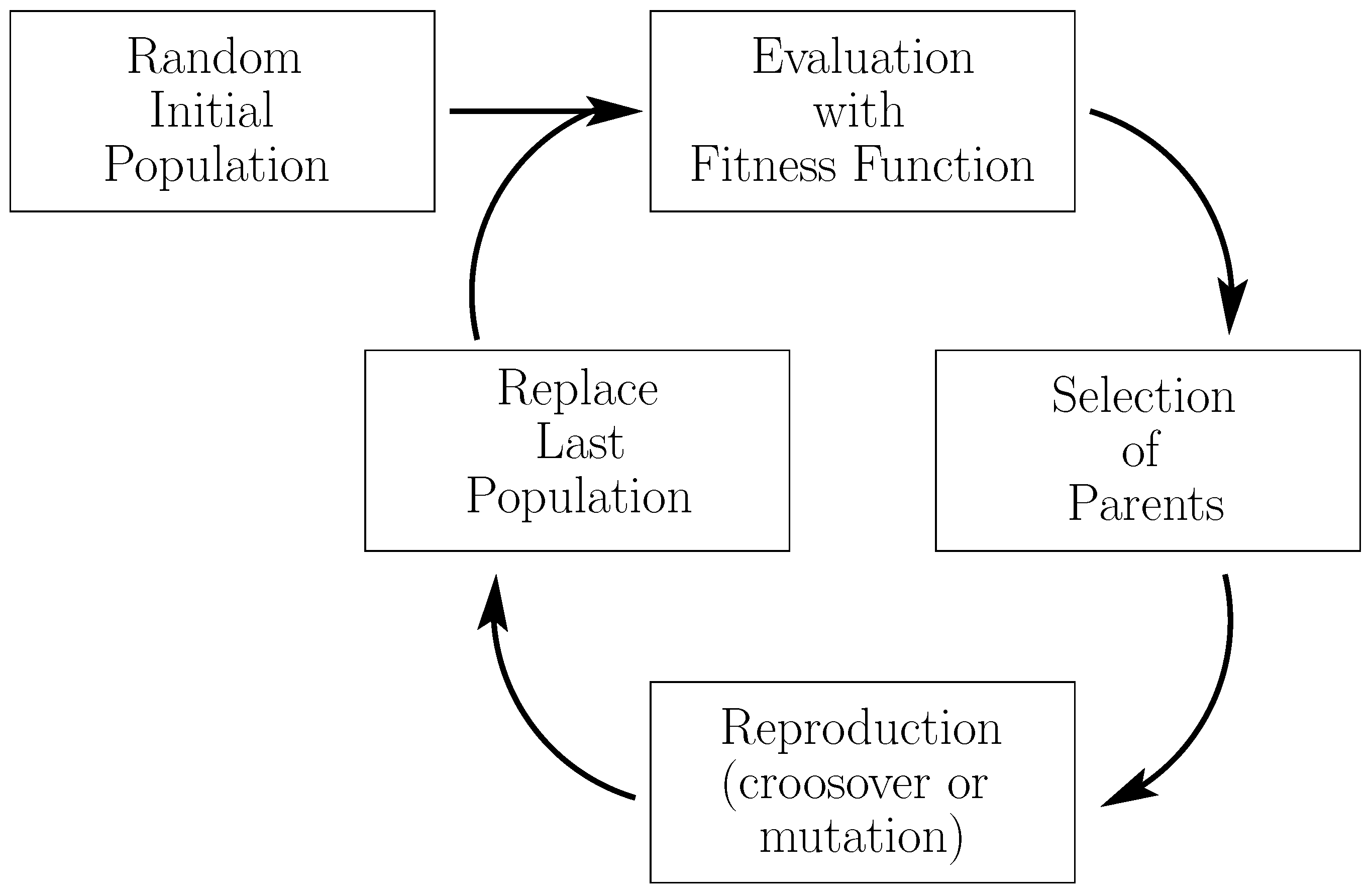
3.4.1. Genetic Programming for Feature Extraction
3.5. Comparison Methods
4. Experiments and Results
4.1. Experimental Setup
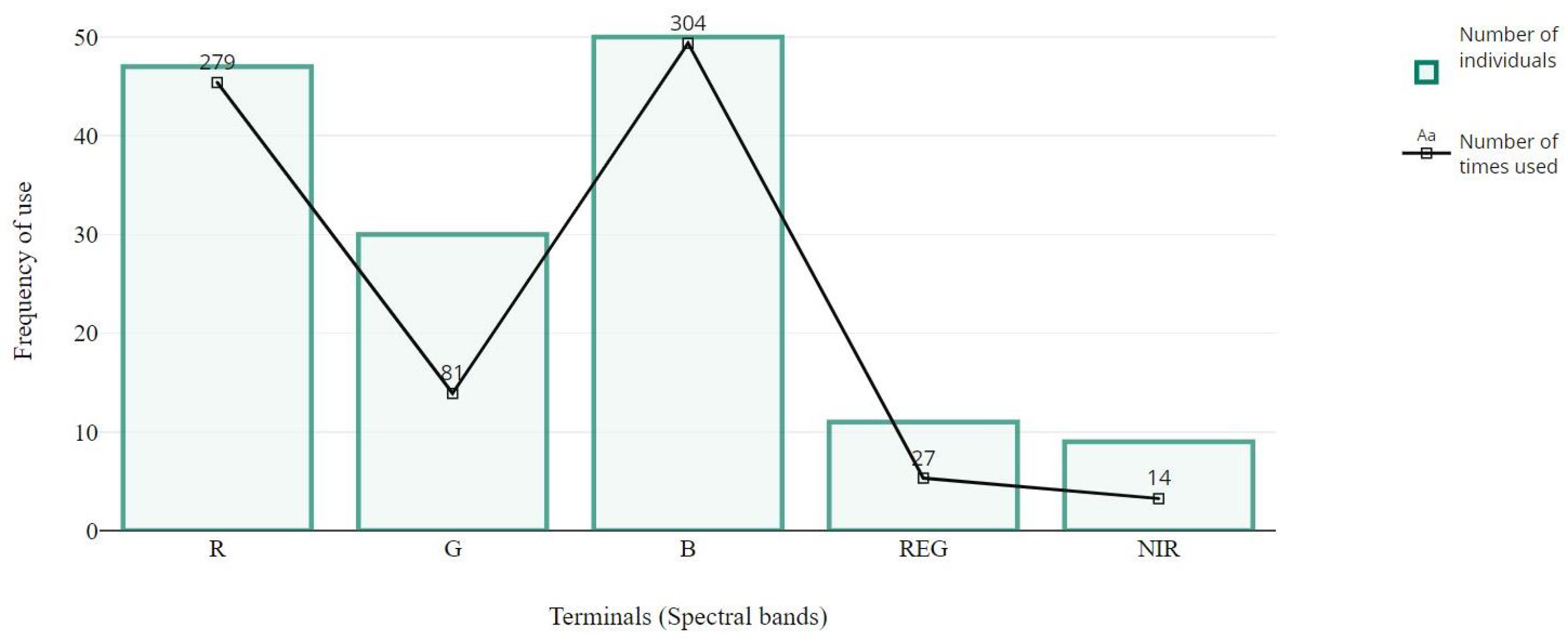
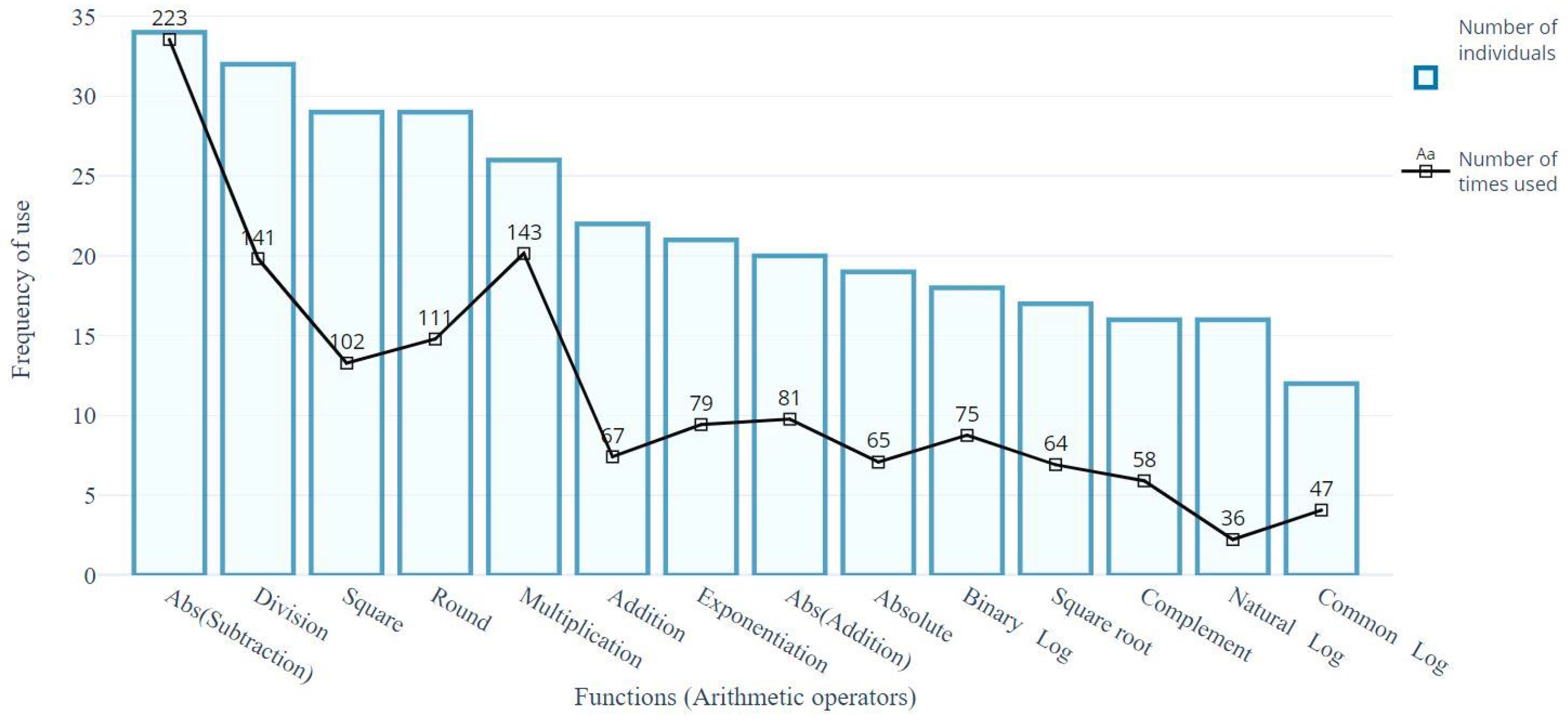
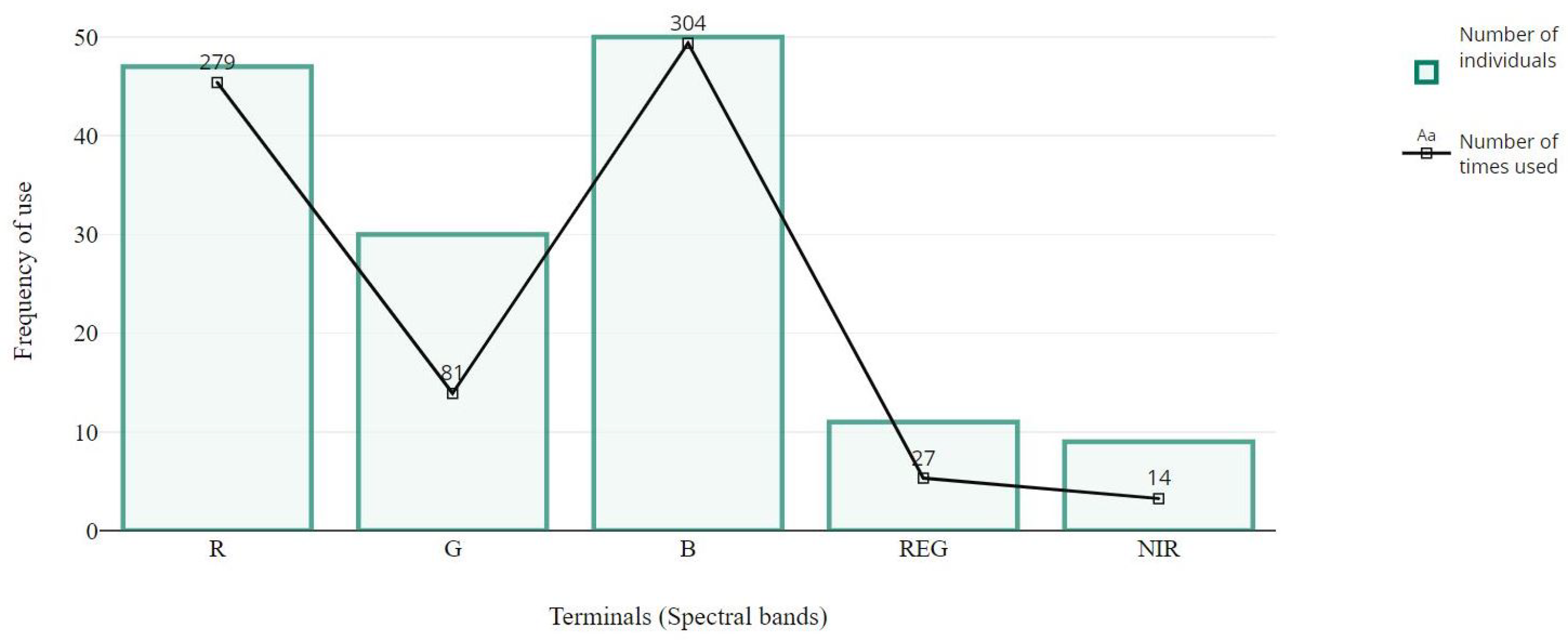
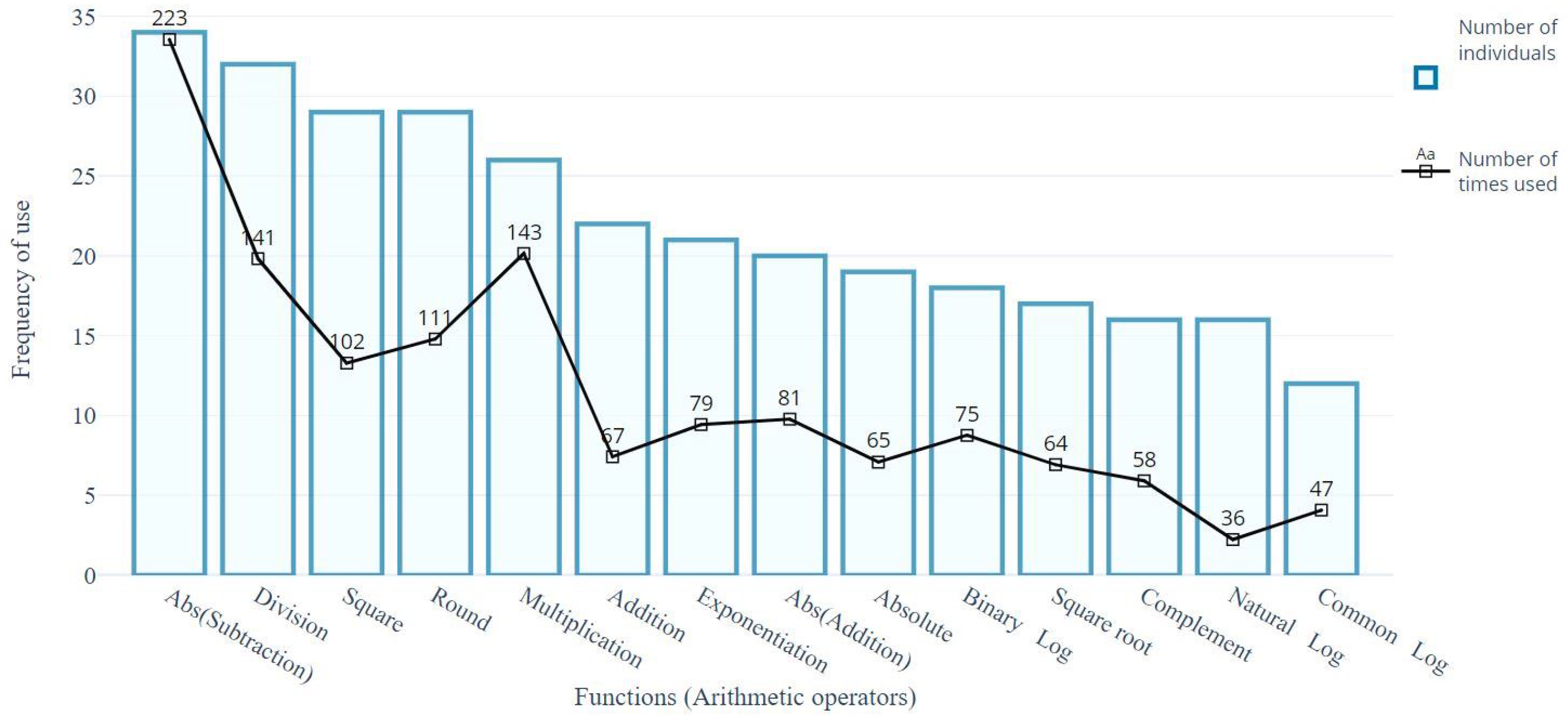
4.2. Structural Analysis of Gp Solutions
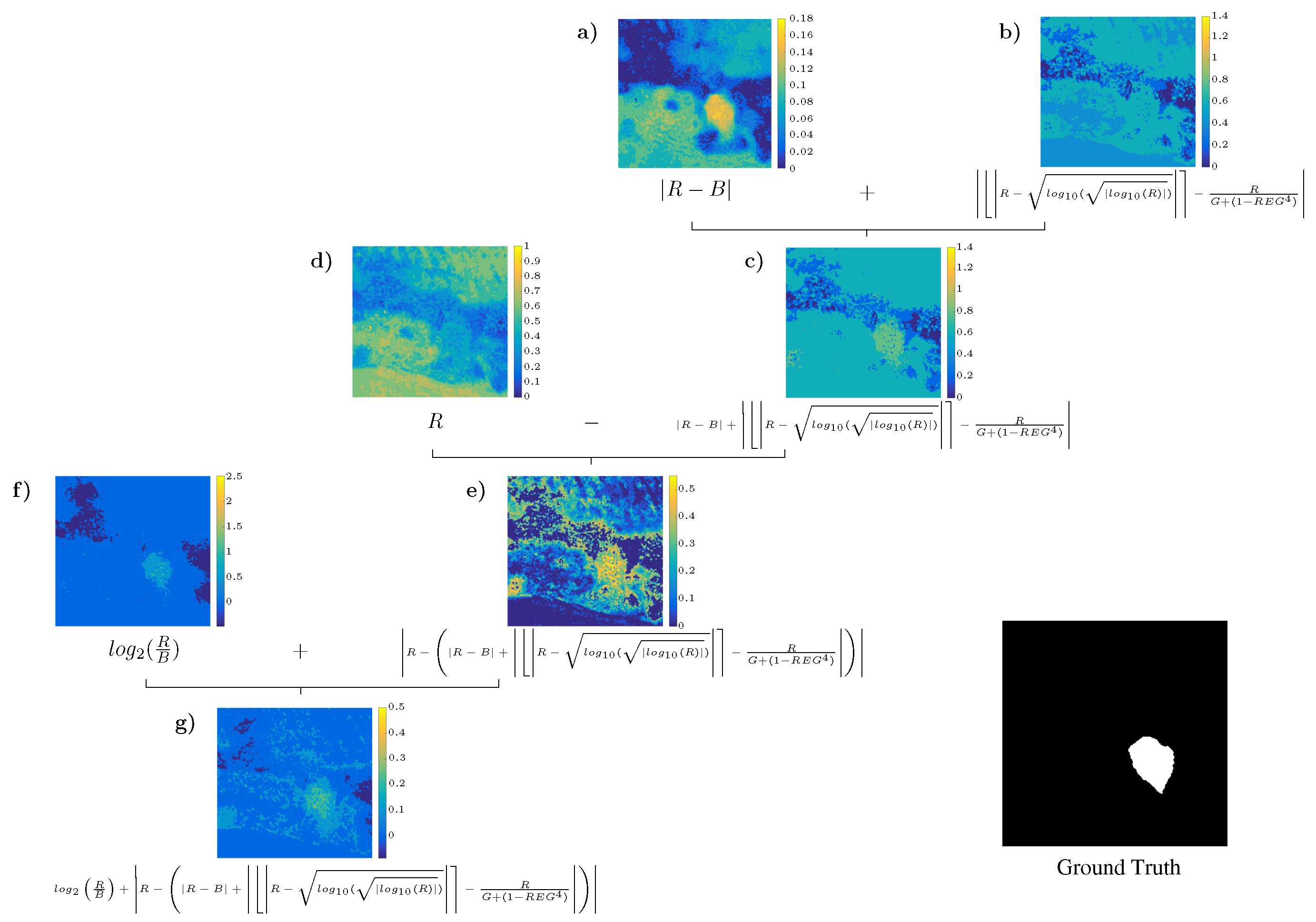
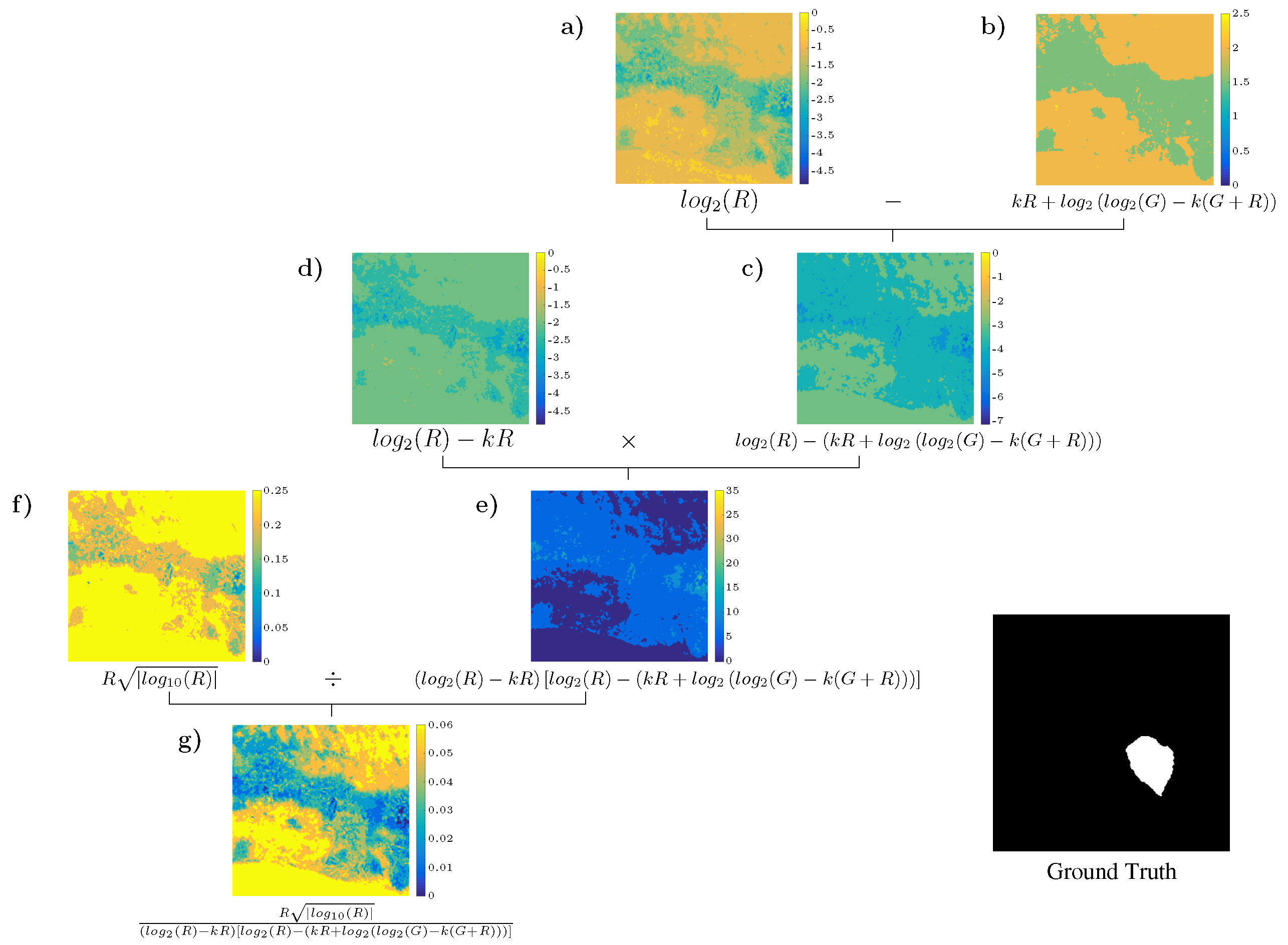
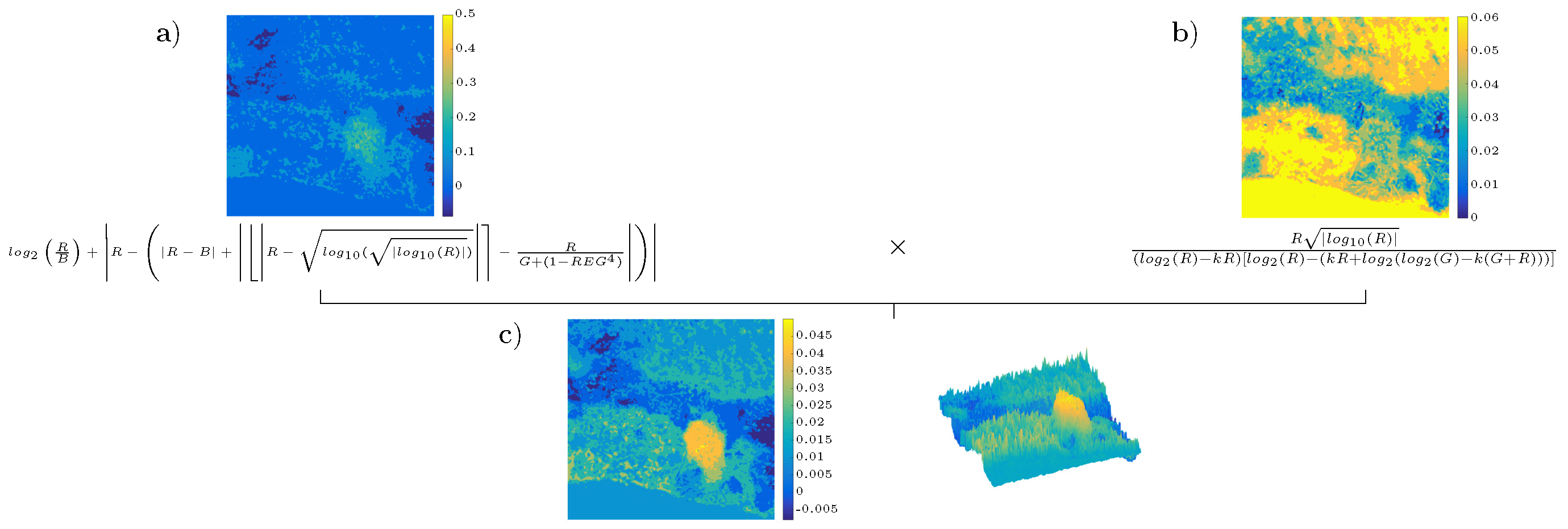
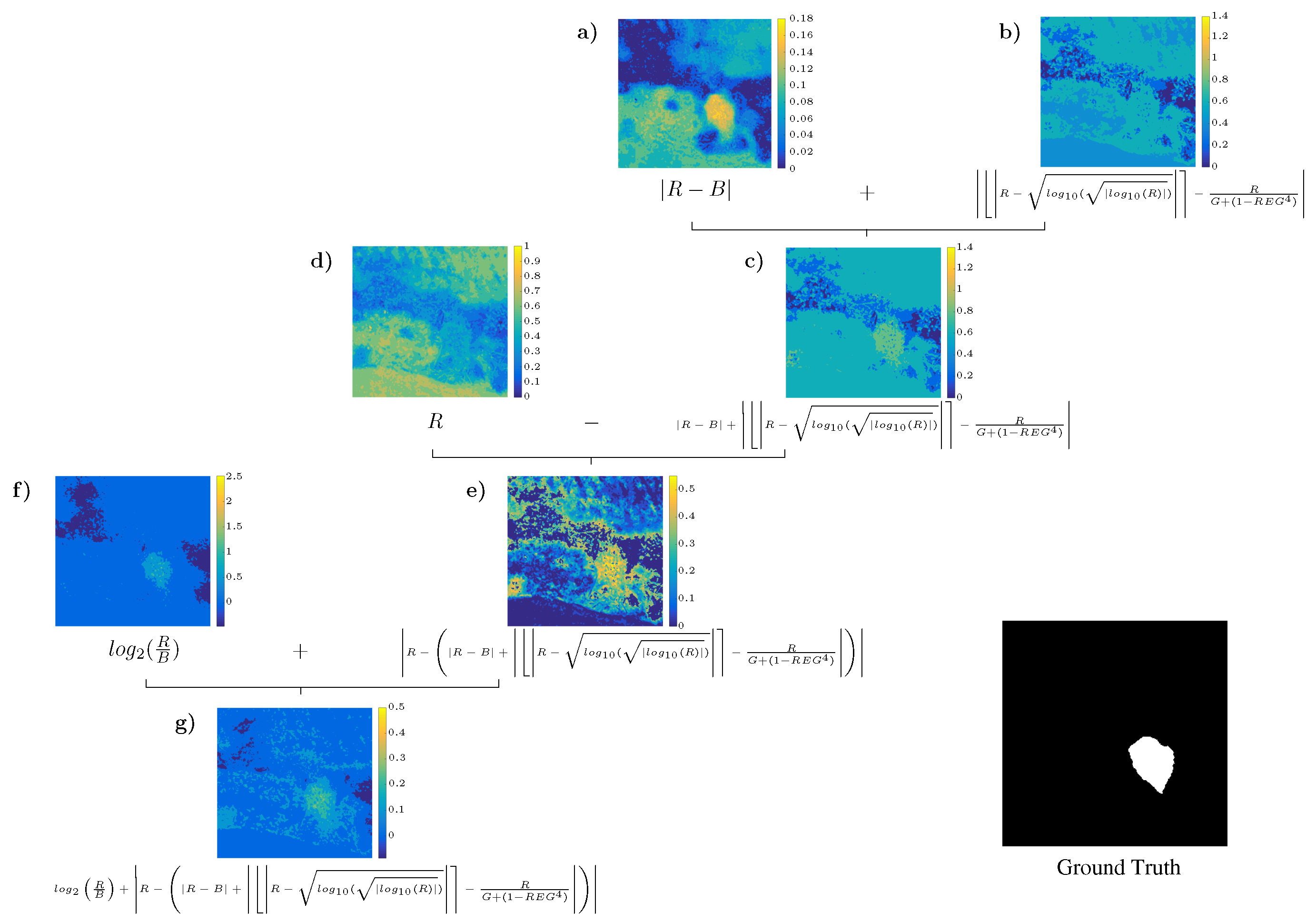
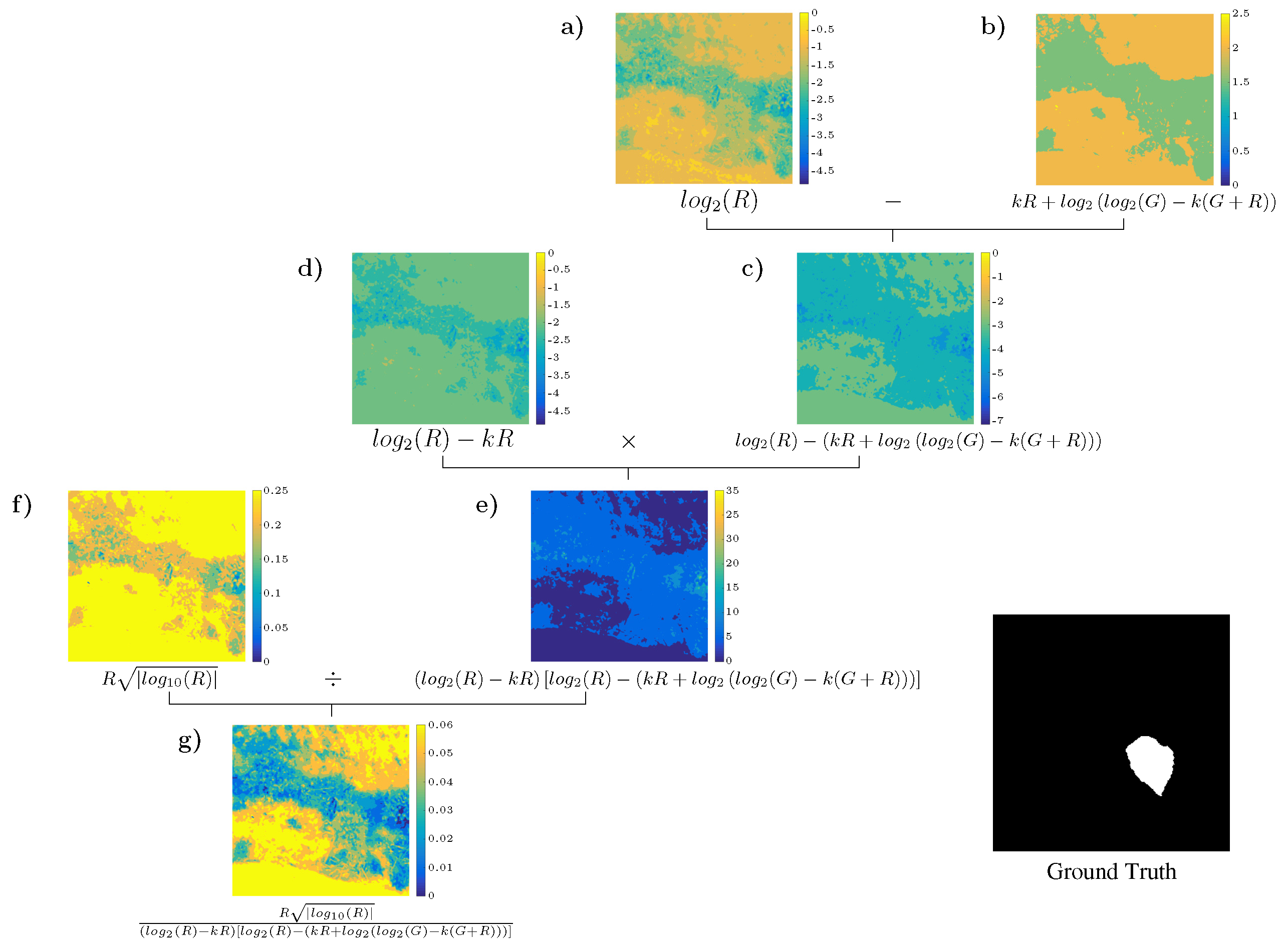
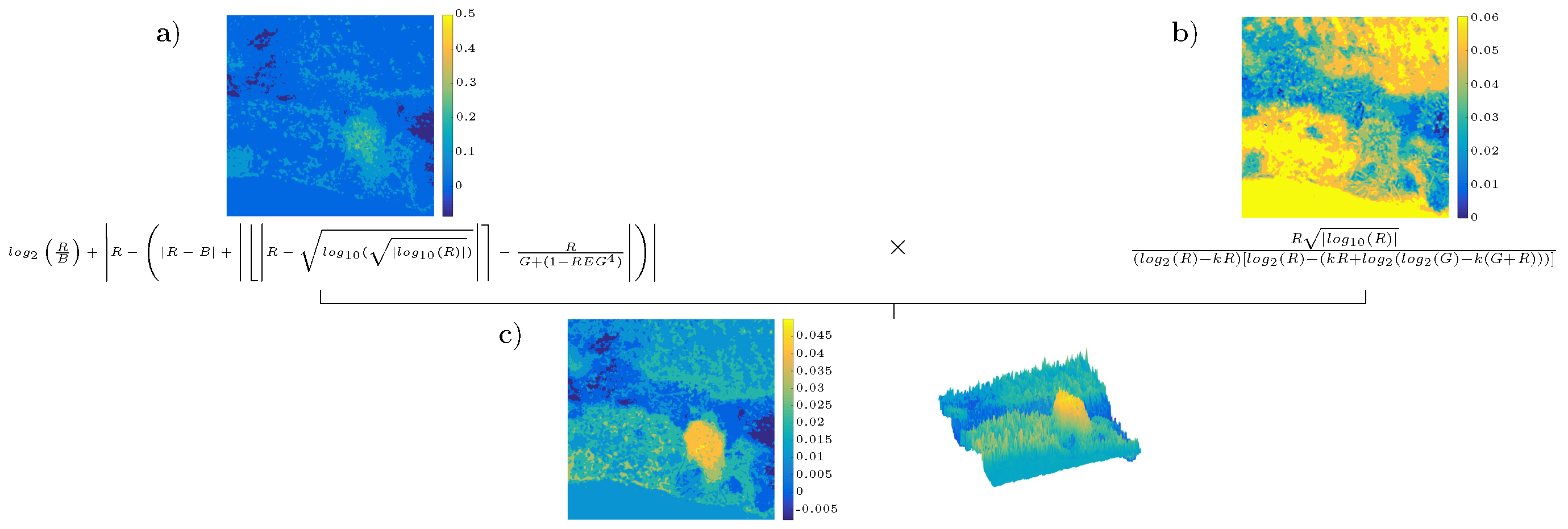
4.3. Analysis of the Best Solution
4.4. Comparison with Other Methods
4.5. Mistletoe Map Detection
5. Discussion
6. Conclusions and Future Work
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Griebel, A.; Watson, D.; Pendall, E. Mistletoe, friend and foe: Synthesizing ecosystem implications of Mistletoe infection. Environ. Res. Lett. 2017, 12, 115012. [Google Scholar] [CrossRef]
- Secretaría del Medio Ambiente de la Ciudad de México—SEDEMA. Suelo de Conservación, 1st ed.; SEDEMA: Mexico City, Mexico, 2016; pp. 89–95.
- Lee, H.S.; Seo, W.W.; Lee, K.S. Detection of Oak Wilt Disease Using Convolutional Neural Network From Uav Natural Color Imagery. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Yokohama, Japan, 28 July–2 August 2019; pp. 6622–6624. [Google Scholar]
- Dwivedi, R.; Dey, S.; Chakraborty, C.; Tiwari, S. Grape Disease Detection Network based on Multi-task Learning and Attention Features. IEEE Sens. J. 2021, 21, 17573–17580. [Google Scholar] [CrossRef]
- Hari-Shankar, R.L.; Veeraraghavan, A.K.; Uvais; Sivaraman, K.; Shreyas, S. Application of UAV for pest, weeds and disease detection using open computer vision. In Proceedings of the International Conference on Smart Systems and Inventive Technology, Tirunelveli, India, 13–14 December 2018; pp. 287–292. [Google Scholar]
- Akila, M.; Deepan, P. Detection and classification of plant leaf diseases by using Deep Learning algorithm. Int. J. Eng. Res. Technol. 2018, 6. Available online: https://www.ijert.org/detection-and-classification-of-plant-leaf-diseases-by-using-deep-learning-algorithm (accessed on 30 November 2021).
- Minařík, R.; Langhammer, J.; Lendzioch, T. Automatic tree crown extraction from uas multispectral imagery for the detection of bark beetle disturbance in mixed forests. Remote Sens. 2020, 12, 4081. [Google Scholar] [CrossRef]
- Nasi, R.; Honkavaara, E.; Blomqvist, M.; Lyytikäinen-Saarenmaa, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Holopainen, M. Remote sensing of bark beetle damage in urban forests at individual tree level using a novel hyperspectral camera from UAV and aircraf. Urban For. Urban Green. 2018, 30, 72–83. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, Y.; Zhang, X. Extraction of tree crowns damaged by Dendrolimus tabulaeformis Tsai et Liu via spectral-spatial classification using UAV-based hyperspectral images. Plant Methods 2020, 16, 135. [Google Scholar] [CrossRef] [PubMed]
- Pernar, R.; Bajic, M.; Ancic, M.; Seletkovic, A.; Idzojtic, M. Detection of mistletoe in digital colour infrared images of infested fir trees. Period. Biol. 2007, 109, 67–75. [Google Scholar]
- Ančić, M.; Pernar, R.; Bajić, M.; Seletković, A.; Kolić, J. Detecting mistletoe infestation on Silver fir using hyperspectral images. iForest-Biogeosciences For. 2013, 7, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Maes, W.H.; Huete, A.R.; Avino, M.; Boer, M.M.; Dehaan, R.; Pendall, E.; Griebel, A.; Steppe, K. Can UAV-based infrared thermography be used to study plant-parasite interactions between mistletoe and eucalypt trees? Remote Sens. 2018, 10, 2062. [Google Scholar] [CrossRef] [Green Version]
- Sabrina, F.; Sohail, S.; Thakur, S.; Azad, S.; Wasimi, S. Use of Deep Learning Approach on UAV imagery to Detect Mistletoe Infestation. In Proceedings of the IEEE Region 10 Symposium, TENSYMP, Dhaka, Bangladesh, 5–7 June 2020; pp. 556–559. [Google Scholar]
- León-Bañuelos, L.A.; Endara-Agramont, A.R.; Gómez-Demetrio, W.; Martínez-García, C.G.; Nava-Bernal, E.G. Identification of Arceuthobium globosum using unmanned aerial vehicle images in a high mountain forest of central Mexico. J. For. Res. 2020, 31, 1759–1771. [Google Scholar] [CrossRef]
- Miraki, M.; Sohrabi, H.; Fatehi, P.; Kneubuehler, M. Detection of mistletoe infected trees using UAV high spatial resolution images. J. Plant Dis. Prot. 2021, 128, 1679–1689. [Google Scholar] [CrossRef]
- Mejia-Zuluaga, P.; Dosal, L.; Valdiviezo-N, J.C. Detection of Phoradendron velutinum. In Proceedings of the iGISc2021: International Conference on Geospatial Information Sciences, Mexico City, Mexico, 3–5 November 2021. [Google Scholar]
- Chen, J.; Li, Z.; Liao, Z.; Wang, Y. Distributed service performance management based on linear regression and genetic programming. In Proceedings of the 2005 International Conference on Machine Learning and Cybernetics, Guangzhou, China, 18–21 August 2005; pp. 560–563. [Google Scholar]
- Zhu, H.-R.; Li, Y.M.; Meng, L.-M. Based on meteorological factors and short-term load forecasting genetic programming. In Proceedings of the 2010 International Conference on Computer Design and Applications, Qinhuangdao, China, 25–27 June 2010; pp. 465–467. [Google Scholar]
- Huo, L.; Fan, X.; Xie, Y.; Yin, J. Short-term load forecasting based on the method of genetic programming. In Proceedings of the 2007 IEEE International Conference on Mechatronics and Automation, Harbin, China, 5–8 August 2007; pp. 839–843. [Google Scholar]
- Brameier, M.; Banzhaf, W. A comparison of linear genetic programming and neural networks in medical data mining. IEEE Trans. Evol. Comput. 2001, 5, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Icke, I.; Bongard, J.C. Improving genetic programming based symbolic regression using deterministic machine learning. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 1763–1770. [Google Scholar]
- Yuan, Y.; Fan, W.; Wang, W.; Liu, H. Robust collaborative optimization of a multi-finger micro-accelerometer based on genetic algorithm. In Proceedings of the 2nd International Conference on Genetic and Evolutionary Computing, Jinzhou, China, 3 October 2008; pp. 105–108. [Google Scholar]
- Phoradendron velutinum (DC.) Oliv. 2014. Available online: https://repositorio.unam.mx/contenidos/34phoradendron-velutinum34-dc-oliv-2590151?c=4EQE34&d=false&q=*:*&i=1&v=1&t=search_0&as=0 (accessed on 30 November 2021).
- González Gaona, E.; Serrano Gómez, C.; De Lira Ramos, K.V.; Quiñonez Barraza, S.; Sánchez Martínez, G.; López Pérez, I.; Sánchez Lucio, R. Identificacion, Distribucion y Control de Muerdago Enano (Arceuthobium spp.) en Bosques de Coniferas; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias: Aguascalientes, Mexico, 2017.
- Gutierrez-Vilchis, L.H.; Resendiz-Martinez, J.F. Fenologia del muerdago enano en el desierto de los leones, D.F. Rev. Cienc. For. Mex. 1994, 19, 41–62. [Google Scholar]
- Rzedowski, J.; Calderon-de-Rzedowski, G. Flora del Bajio y de Regiones Adyacentes; Insituto de Ecologia a.c.: Michoacan, Mexico, 2011. [Google Scholar]
- Mathiasen, R.L.; Nickrent, D.L.; Shaw, D.C.; Watson, D.M. Mistletoes: Pathology, systematics, ecology, and management. Plant Dis. 2008, 92, 988–1006. [Google Scholar] [CrossRef] [Green Version]
- Alvarado-Rosales, D.; Equihua-Martínez, A.; Tagle, E.L.-G.; Rodríguez-Ortega, A. Situacion actual de la declinacion del oyamel en el parque Desierto de los Leones, D.F. In Proceedings of the XIV Simposio Nacional de Parasitologia Forestal, Aguascalientes, Mexico, 7–9 November 2007; pp. 24–28. [Google Scholar]
- Silva, S.; Almeida, J. GPLAB A Genetic Programming Toolbox for MATLAB. In Proceedings of the Nordic MATLAB Conference, Stockholm, Sweden, April 2007. [Google Scholar]
- Bay, H.; Tuytelaars, T.; Van Gool, L. SURF: Speeded Up Robust Features. In Proceedings of the Computer Vision—ECCV, Graz, Austria, 7–13 May 2006; pp. 404–417. [Google Scholar] [CrossRef]
- Wang, Z.; Bovik, A.C.; Sheikh, H.R.; Simoncelli, E.P. Image quality assessment: From error visibility to structural similarity. IEEE Trans. Image Process. 2004, 13, 600–612. [Google Scholar] [CrossRef] [Green Version]
- Itti, L.; Koch, C. A saliency-based search mechanism for overt and covert shifts of visual attention. Vis. Res. 2000, 40, 1489–1506. [Google Scholar] [CrossRef] [Green Version]
- Koza, J.R. Genetic Programming: On the Programming of Computers by Means of Natural Selection, 1st ed.; A Bradford Book: Cambridge, MA, USA, 1992; p. 840. [Google Scholar]
- Cohen, J. Weighted kappa: Nominal scale agreement with provision for scaled disagreement or partial credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Luke, S.; Panait, L. Lexicographic parsimony pressure. In Proceedings of the 4th Annual Conference on Genetic and Evolutionary Computation, New York, NY, USA, 9–13 July 2002; pp. 829–836. [Google Scholar]
- Blackburn, G.A. Spectral indices for estimating photosynthetic pigment concentrations: A test using senescent tree leaves. Int. J. Remote Sens. 1998, 19, 657–675. [Google Scholar] [CrossRef]
- Pu, R.; Gong, P.; Yu, Q. Comparative Analysis of EO-1 ALI and Hyperion, and Landsat ETM+ Data for Mapping Forest Crown Closure and Leaf Area Index. Sensors 2008, 8, 3744–3766. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Wu, Z.; Zhang, L.; Li, J.; Zhou, J.; Jun, Z.; Zhu, B. Monitoring of peanut leaves chlorophyll content based on drone-based multispectral image feature extraction. Comput. Electron. Agric. 2021, 187, 106292. [Google Scholar] [CrossRef]
- Mountrakis, G.; Im, J.; Ogole, C. Support vector machines in remote sensing: A review. ISPRS J. Photogramm. Remote Sens. 2011, 66, 247–259. [Google Scholar] [CrossRef]
- Gamon, J.; Surfus, J. Assessing Leaf Pigment Content and Activity With a Reflectometer. New Phytol. 1999, 143, 105–117. [Google Scholar] [CrossRef]
- Delegido, J.; Verrelst, J.; Meza, C.M.; Rivera, J.P.; Alonso, L.; Moreno, J. A red-edge spectral index for remote sensing estimation of green LAI over agroecosystems. Eur. J. Agron. 2013, 46, 42–52. [Google Scholar] [CrossRef]
- Kawashima, S.; Nakatani, M. An algorithm for estimating chlorophyll content in leaves using a video camera. Ann. Bot. 1998, 81, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Alasta, A.F. Using Remote Sensing data to identify iron deposits in central western Libya. In Proceedings of the International Conference on Emerging Trends in Computer and Image Processing (ICETCIP’2011); pp. 56–61. Available online: http://psrcentre.org/images/extraimages/122.%201211924.pdf (accessed on 30 November 2021).
- Deng, G.; Cahill, L.W.; Tobin, G.R. The study of logarithmic image processing model and its application to image enhancement. IEEE Trans. Image Process. 1995, 4, 506–512. [Google Scholar] [CrossRef]
- Negru, M.; Nedevschi, S.; Peter, R.I. Exponential image enhancement in daytime fog conditions. In Proceedings of the 17th International IEEE Conference on Intelligent Transportation Systems (ITSC), Qingdao, China, 8–11 October 2014; pp. 1675–1681. [Google Scholar]
- Varma, M.K.S.; Rao, N.K.K.; Raju, K.K.; Varma, G.P.S. Pixel-Based Classification Using Support Vector Machine Classifier. In Proceedings of the 2016 IEEE 6th International Conference on Advanced Computing (IACC), Bhimavaram, India, 27–28 February 2016; pp. 51–55. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polygon | Area (ha) | Flying Height (m) | GSD (cm/px) | Number of Images |
|---|---|---|---|---|
| 1 | 31.40 | 110 | 5.8 | 474 |
| 2 | 46.34 | 80 | 4.2 | 1244 |
| 3 | 49.14 | 90 | 4.8 | 522 |
| 4 | 20.37 | 110 | 5.8 | 325 |
| Protected Function | Input Arguments | Output Argument |
|---|---|---|
| Division | a, b | a (if b ) |
| a/b (otherwise) | ||
| Square root | a | 0 (if a ≤ 0 ) |
| sqrt (a) (otherwise) | ||
| Base 2 | a | 0 (if a ) |
| Logarithm | a (otherwise) | |
| Base 10 | a | 0 (if a ) |
| Logarithm | a (otherwise) |
| Feature | Feature | ||
|---|---|---|---|
| Generations | 50 | Selection | Lexictour |
| Population size | 100 | Elitism criteria | Keepbest |
| Initialization | Rampedinit | Functions set | +, , , ÷, ×, , , , , , , ¬, , |
| Crossover | Probability 0.7 | Terminals set | |
| Mutation | Probability 0.3 | Fitness function | Weighted Cohen’s Kappa |
| Predicted | |||
|---|---|---|---|
| Infested | Other | ||
| Real | Infested | 0 | 1 |
| Other | 2 | 0 | |
| Method | Training | Test | Diff. | ||||
|---|---|---|---|---|---|---|---|
| k | OA | R | P | ||||
| 0.465 | 0.448 | 0.442 | 0.966 | 0.482 | 0.498 | −0.017 | |
| 0.436 | 0.416 | 0.395 | 0.964 | 0.389 | 0.524 | −0.021 | |
| 0.298 | 0.294 | 0.275 | 0.963 | 0.269 | 0.412 | −0.004 | |
| 0.296 | 0.282 | 0.284 | 0.952 | 0.357 | 0.319 | −0.014 | |
| SIPI2 | 0.0025 | −0.005 | -0.006 | 0.543 | 0.426 | 0.028 | −0.007 |
| 0.267 | 0.276 | 0.261 | 0.913 | 0.326 | 0.373 | 0.009 | |
| 0.236 | 0.197 | 0.203 | 0.732 | 0.649 | 0.247 | −0.039 | |
| SVM | Validation | ||||||
| 0.198 | 0.168 | 0.207 | 0.881 | 0.146 | 0.704 | −0.030 |
| Original Image | Testing Results | ||
|---|---|---|---|
| GP | SIPI2 | SVM | |
 | 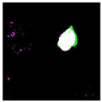 OA P R |  OA P R |  OA P R |
 |  OA P R |  OA P R |  OA P R |
 |  OA P R |  OA P R |  OA P R |
 | 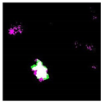 OA P R |  OA P R |  OA P R |
 | |||
| Original Image | Testing Results | |
|---|---|---|
 |  |  |
| OA P R | OA P R | |
 |  |  |
| OA P R | OA P R | |
 |  |  |
| OA P R | OA P R | |
 |  |  |
| OA P R | OA P R | |
 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejia-Zuluaga, P.A.; Dozal, L.; Valdiviezo-N., J.C. Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City. Remote Sens. 2022, 14, 801. https://doi.org/10.3390/rs14030801
Mejia-Zuluaga PA, Dozal L, Valdiviezo-N. JC. Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City. Remote Sensing. 2022; 14(3):801. https://doi.org/10.3390/rs14030801
Chicago/Turabian StyleMejia-Zuluaga, Paola Andrea, León Dozal, and Juan C. Valdiviezo-N. 2022. "Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City" Remote Sensing 14, no. 3: 801. https://doi.org/10.3390/rs14030801
APA StyleMejia-Zuluaga, P. A., Dozal, L., & Valdiviezo-N., J. C. (2022). Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City. Remote Sensing, 14(3), 801. https://doi.org/10.3390/rs14030801