
Assessing Spatial Variation in Algal Productivity in a Tropical River Floodplain Using Satellite Remote Sensing



Abstract

1. Introduction
2. Materials and Methods
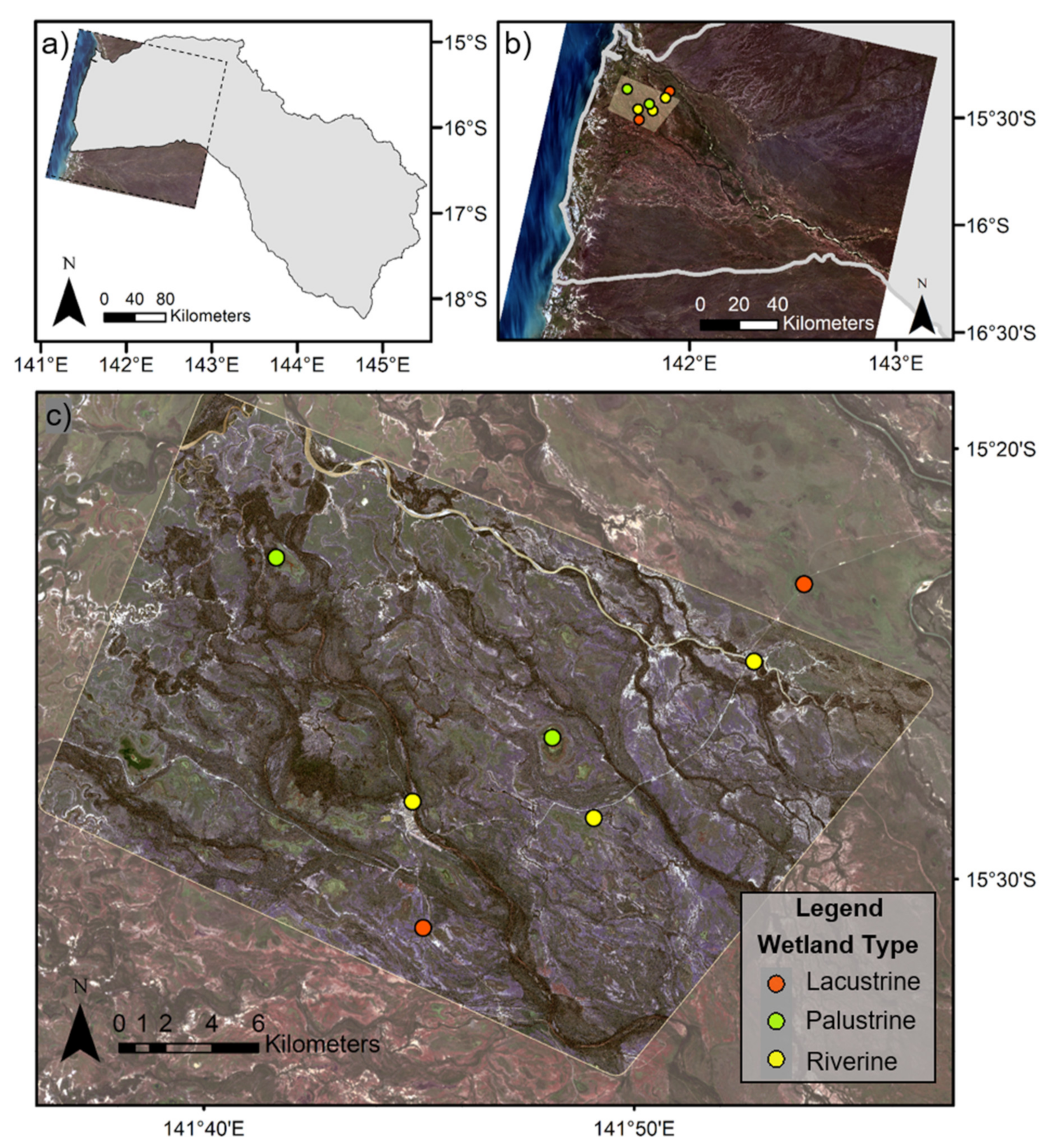
2.1. Study Area
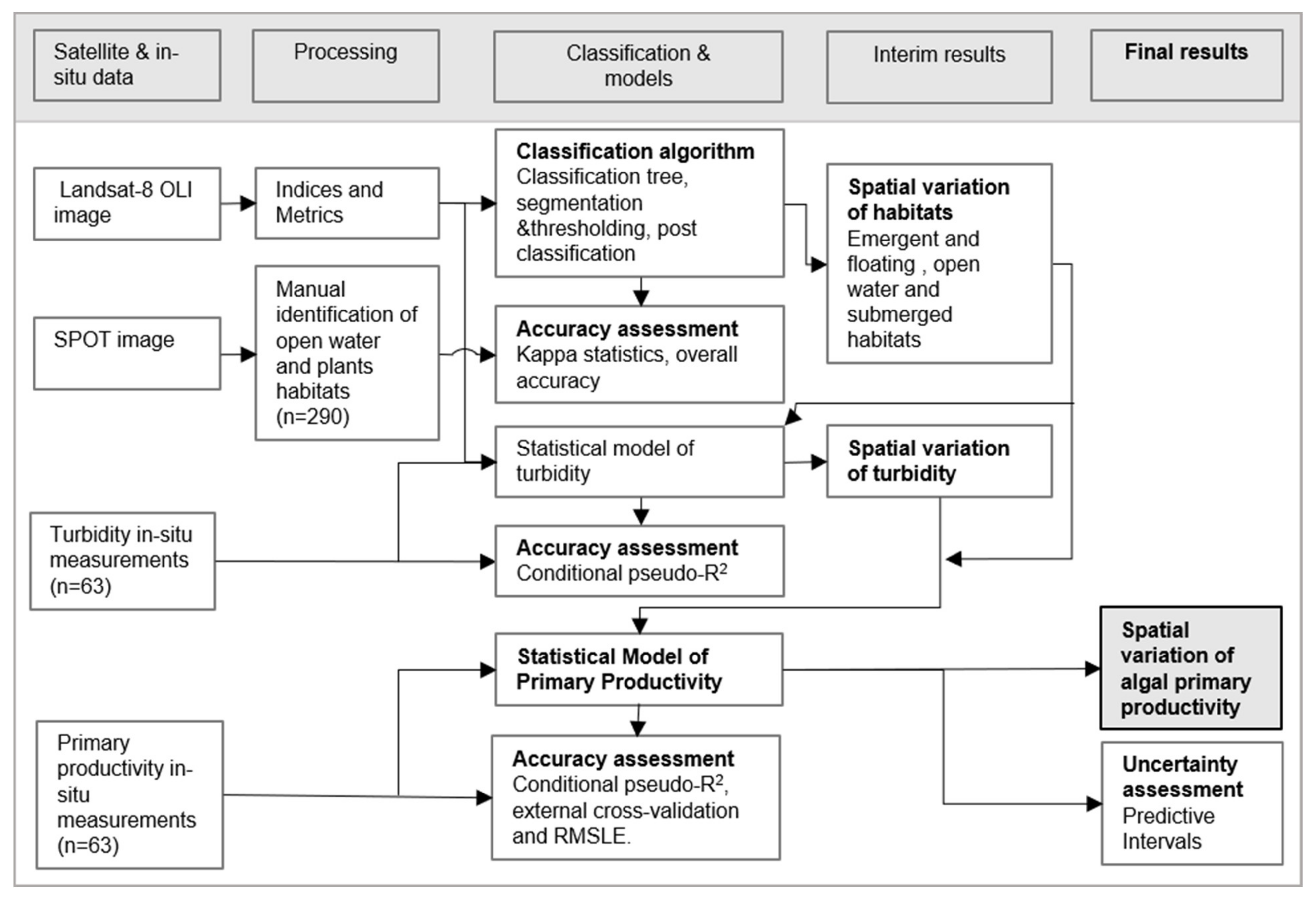
2.2. Methodological Framework
2.3. Data Acquisition
2.3.1. Field Campaign
2.3.2. Optical Imagery
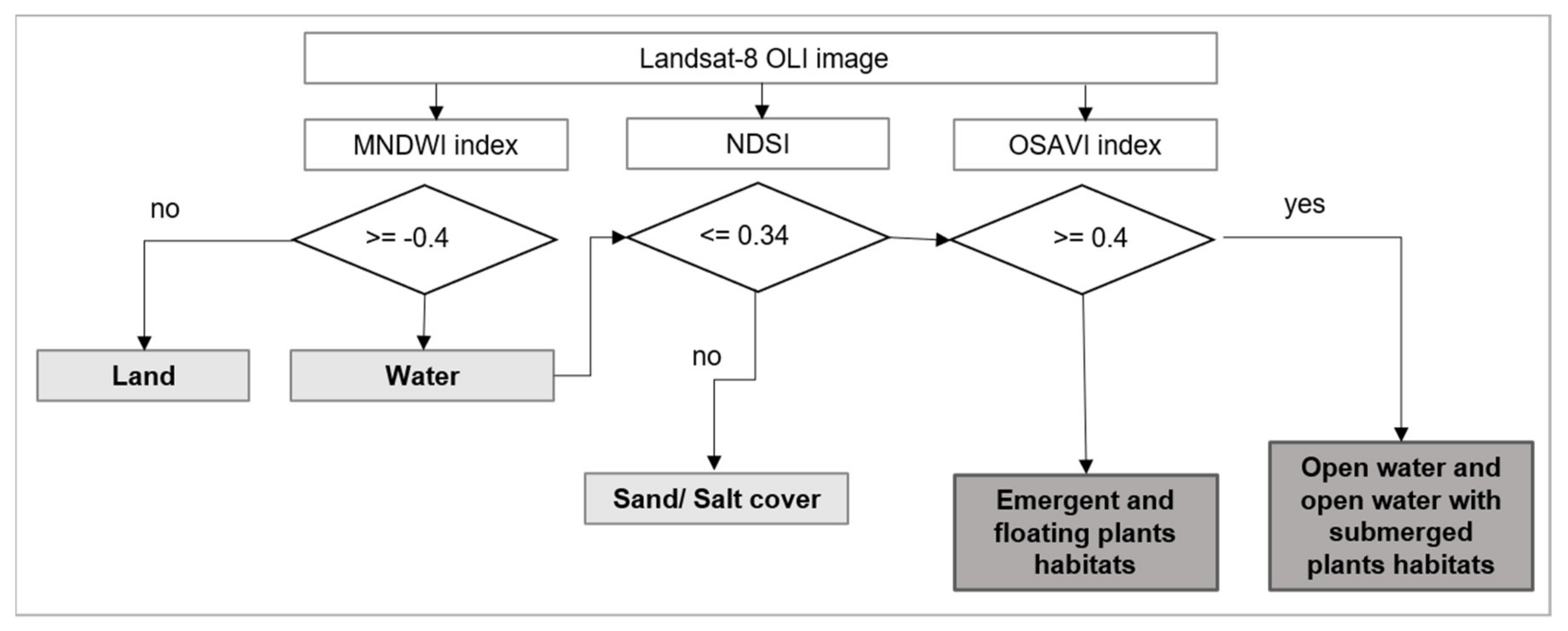
2.4. Statistical Modelling of Habitats
2.5. Statistical Modelling of Turbidity
2.6. Statistical Modelling of Algal Primary Productivity
3. Results
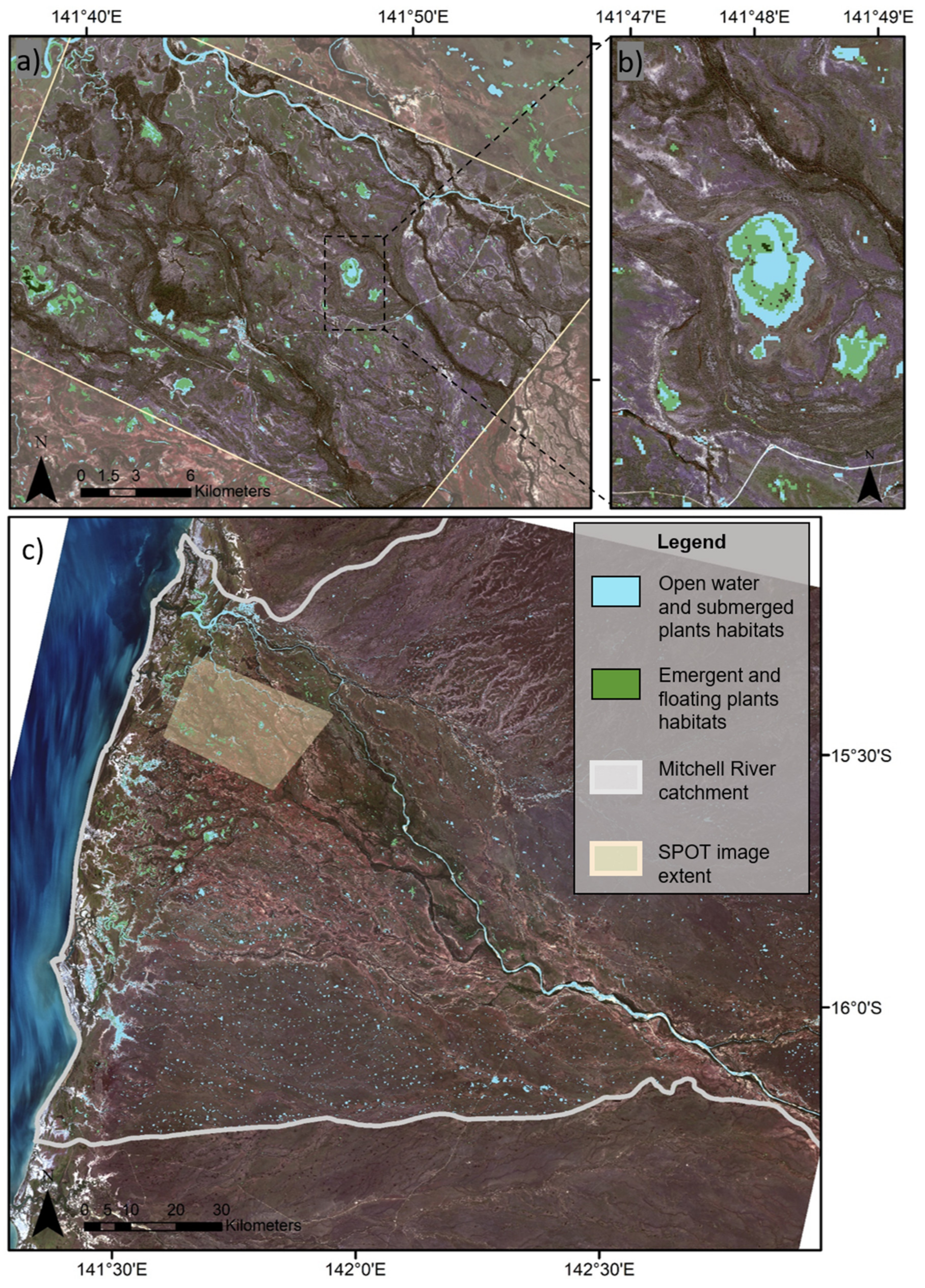
3.1. Statistical Modelling of Habitat Type
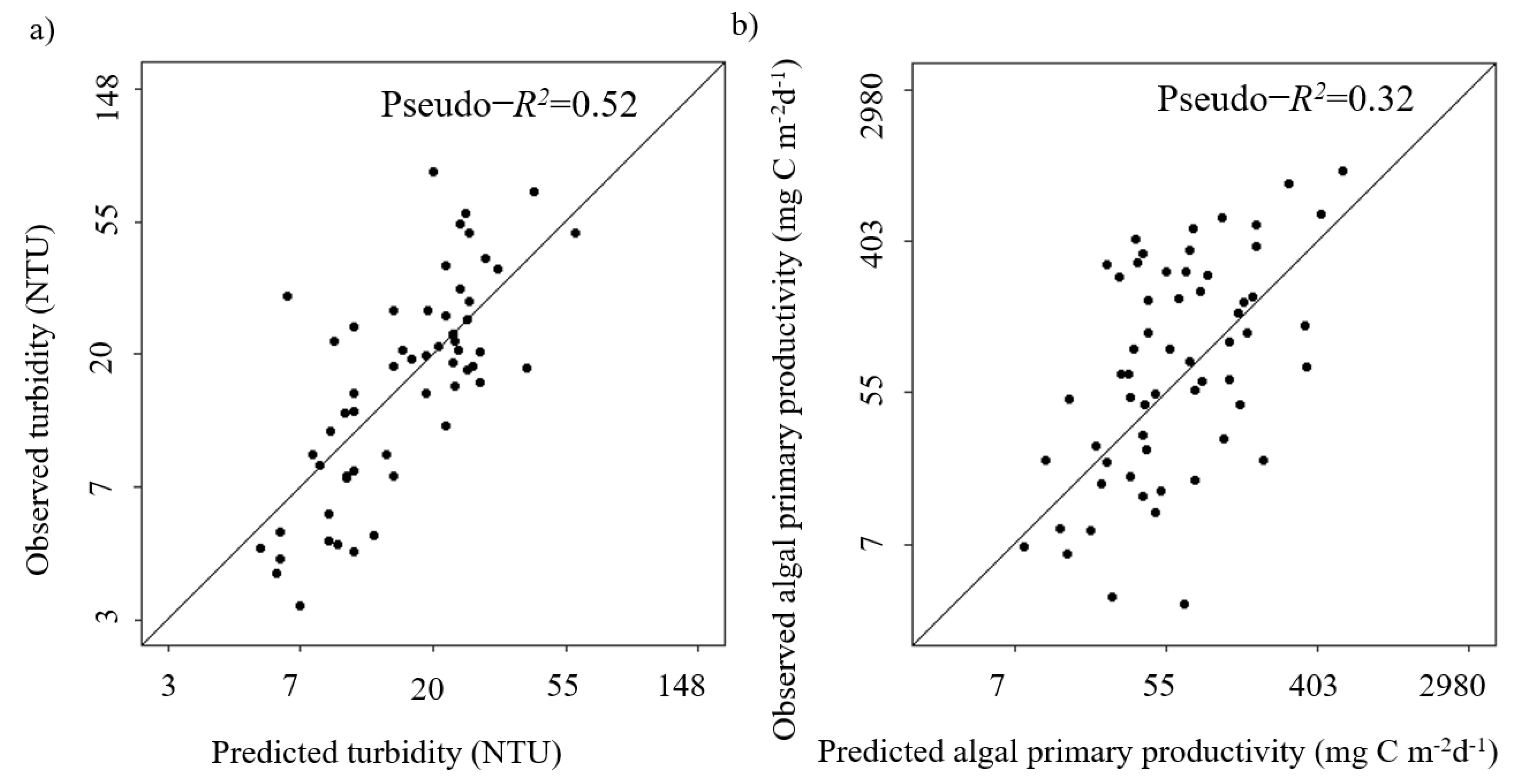
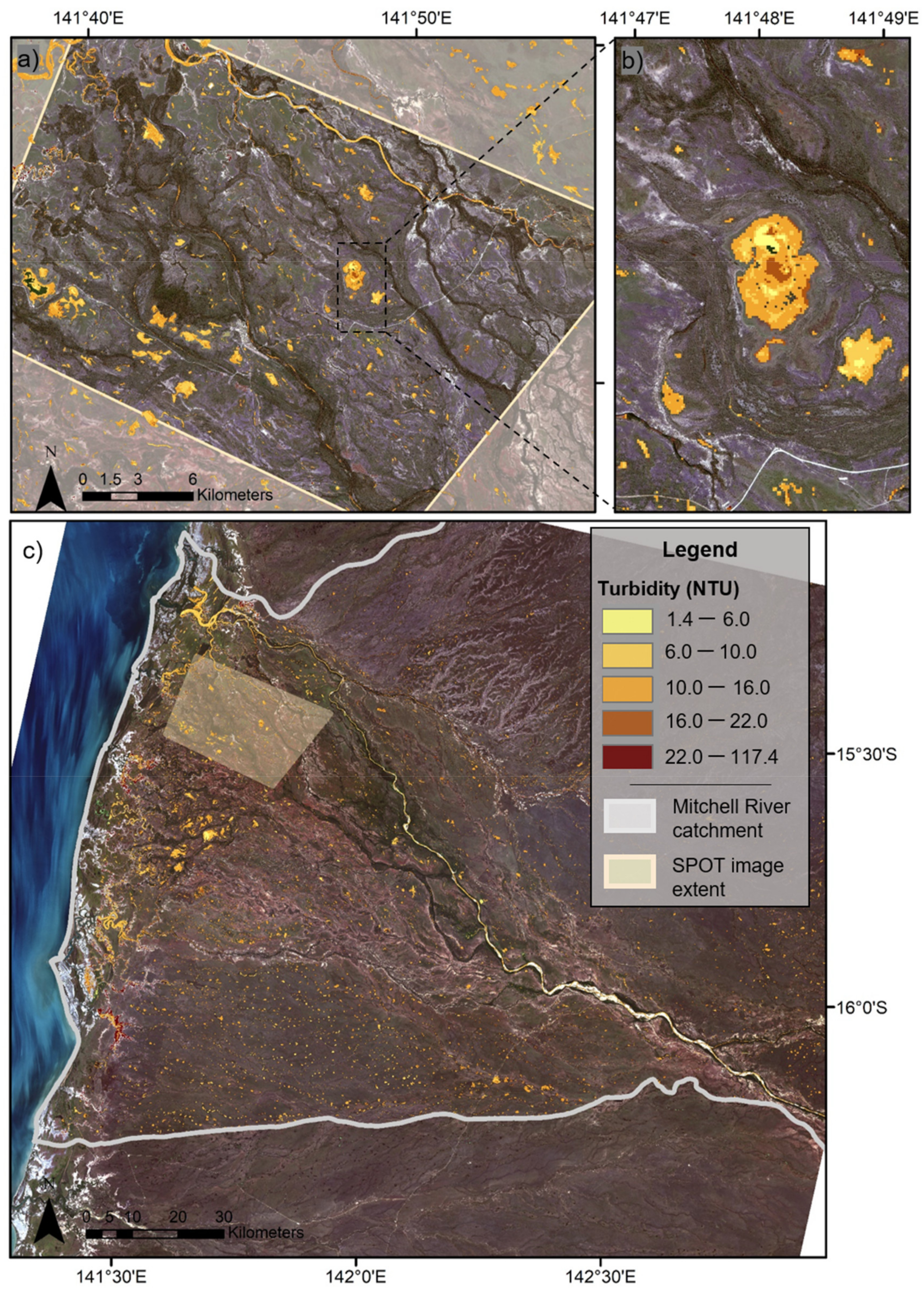
3.2. Statistical Modelling of Turbidity
3.3. Statistical Modelling of Algal Primary Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Bunn, S.; Ward, D.; Crook, D.A.; Jardine, T.; Adame, F.; Pettit, N.; Douglas, M.M.; Valdez, D.; Kyne, P.M. Tropical Floodplain Food Webs—Connectivity and Hotspots—Final Report; Charles Darwin University: Darwin, Australia, 2015. [Google Scholar]
- Jardine, T.D.; Pusey, B.J.; Hamilton, S.K.; Pettit, N.E.; Davies, P.M.; Douglas, M.M.; Sinnamon, V.; Halliday, I.A.; Bunn, S.E. Fish mediate high food web connectivity in the lower reaches of a tropical floodplain river. Oecologia 2012, 168, 829–838. [Google Scholar] [CrossRef]
- Jardine, T.D.; Hunt, R.; Faggotter, S.; Valdez, D.; Burford, M.; Bunn, S. Carbon from periphyton supports fish biomass in waterholes of a wet-dry tropical river. River Res. Appl. 2013, 29, 560–573. [Google Scholar] [CrossRef]
- Saigo, M.; Zilli, F.L.; Marchese, M.R.; Demonte, D. Trophic level, food chain length and omnivory in the Paraná River: A food web model approach in a floodplain river system. Ecol. Res. 2015, 30, 843–852. [Google Scholar] [CrossRef]
- Brett, M.T.; Bunn, S.E.; Chandra, S.; Galloway, A.W.E.; Guo, F.; Kainz, M.J.; Kankaala, P.; Lau, D.C.P.; Moulton, T.P.; Power, M.E.; et al. How important are terrestrial organic carbon inputs for secondary production in freshwater ecosystems? Freshw. Biol. 2017, 62, 833–853. [Google Scholar] [CrossRef]
- Kuhn, C.D.; Bogard, M.; Johnston, S.E.; John, A.; Vermote, E.F.; Spencer, R.; Dornblaser, M.; Wickland, K.P.; Striegl, R.G.; Butman, D. Satellite and airborne remote sensing of gross primary productivity in boreal Alaskan lakes. Environ. Res. Lett. 2020, 15, 105001. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Devlin, S.; McIntyre, P.; Vander Zanden, J. Is there light after depth? Distribution of periphyton chlorophyll and productivity in lake littoral zones. Freshw. Sci. 2014, 33, 524–536. [Google Scholar] [CrossRef]
- Adame, M.F.; Pettit, N.E.; Valdez, D.; Ward, D.; Burford, M.A.; Bunn, S.E. The contribution of epiphyton to the primary production of tropical floodplain wetlands. Biotropica 2017, 49, 461–471. [Google Scholar] [CrossRef]
- Molinari, B.; Stewart-Koster, B.; Adame, M.F.; Campbell, M.D.; McGregor, G.; Schulz, C.; Malthus, T.J.; Bunn, S. Relationships between algal primary productivity and environmental variables in tropical floodplain wetlands. Inland Waters 2021, 1–11. [Google Scholar] [CrossRef]
- Jia, J.; Gao, Y.; Zhou, F.; Shi, K.; Johnes, P.J.; Dungait, J.A.J.; Ma, M.; Lu, Y. Identifying the main drivers of change of phytoplankton community structure and gross primary productivity in a river-lake system. J. Hydrol. 2020, 583, 124633. [Google Scholar] [CrossRef]
- Vis, C.; Hudon, C.; Carignan, R.; Gagnon, P. Spatial Analysis of Production by Macrophytes, Phytoplankton and Epiphyton in a Large River System under Different Water-Level Conditions. Ecosystems 2007, 10, 293–310. [Google Scholar] [CrossRef]
- Garcia, E.A.; Townsend, S.A.; Douglas, M.M. Context dependency of top-down and bottom-up effects in a Northern Australian tropical river. Freshw. Sci. 2015, 34, 679–690. [Google Scholar] [CrossRef]
- Polis, G.A. Why Are Parts of the World Green? Multiple Factors Control Productivity and the Distribution of Biomass. Oikos 1999, 86, 3–15. [Google Scholar] [CrossRef]
- Thompson, R.M.; Townsend, C.R. Energy Availability, Spatial Heterogeneity and Ecosystem Size Predict Food-Web Structure in Streams. Oikos 2005, 108, 137–148. [Google Scholar] [CrossRef]
- Palmer, S.C.J.; Kutser, T.; Hunter, P.D. Remote sensing of inland waters: Challenges, progress and future directions. Remote Sens. Environ. 2015, 157, 1–8. [Google Scholar] [CrossRef]
- Zolfaghari, K.; Duguay, C.R. Estimation of water quality parameters in Lake Erie from MERIS using Linear mixed effect models. Remote Sens. 2016, 8, 473. [Google Scholar] [CrossRef]
- Tyler, A.N.; Svab, E.; Preston, T.; Présing, M.; Kovács, W.A. Remote sensing of the water quality of shallow lakes: A mixture modelling approach to quantifying phytoplankton in water characterized by high-suspended sediment. Int. J. Remote Sens. 2006, 27, 1521–1537. [Google Scholar] [CrossRef]
- Watanabe, F.S.Y.; Alcantara, E.; Rodrigues, T.W.P.; Imai, N.N.; Barbosa, C.C.F.; Rotta, L.H.D. Estimation of Chlorophyll-a Concentration and the Trophic State of the Barra Bonita Hydroelectric Reservoir Using OLI/Landsat-8 Images. Int. J. Environ. Res. Public Health 2015, 12, 10391–10417. [Google Scholar] [CrossRef]
- Rotta, L.; Alcântara, E.; Park, E.; Bernardo, N.; Watanabe, F. A single semi-analytical algorithm to retrieve chlorophyll-a concentration in oligo-to-hypereutrophic waters of a tropical reservoir cascade. Ecol. Indic. 2021, 120, 106913. [Google Scholar] [CrossRef]
- Ward, D.P.; Pettit, N.E.; Adame, M.; Douglas, M.M.; Setterfield, S.A.; Bunn, S.E. Seasonal spatial dynamics of floodplain macrophyte and periphyton abundance in the Alligator Rivers region (Kakadu) of northern Australia: Kakadu Seasonal Dynamics of Macrophytes and Epiphytic Algae. Ecohydrology 2016, 9, 1675–1686. [Google Scholar] [CrossRef]
- Pettit, N.E.; Bayliss, P.; Davies, P.M.; Hamilton, S.K.; Warfe, D.M.; Bunn, S.E.; Douglas, M.M. Seasonal contrasts in carbon resources and ecological processes on a tropical floodplain. Freshw. Biol. 2011, 56, 1047–1064. [Google Scholar] [CrossRef]
- Pettit, N.E.; Ward, D.P.; Adame, M.F.; Valdez, D.; Bunn, S.E. Influence of aquatic plant architecture on epiphyte biomass on a tropical river floodplain. Aquat. Bot. 2016, 129, 35–43. [Google Scholar] [CrossRef]
- Costa, M. Estimate of net primary productivity of aquatic vegetation of the Amazon floodplain using Radarsat and JERS-1. Int. J. Remote Sens. 2005, 26, 4527–4536. [Google Scholar] [CrossRef]
- Güttler, F.N.; Niculescu, S.; Gohin, F. Turbidity retrieval and monitoring of Danube Delta waters using multi-sensor optical remote sensing data: An integrated view from the delta plain lakes to the western–northwestern Black Sea coastal zone. Remote Sens. Environ. 2013, 132, 86–101. [Google Scholar] [CrossRef]
- Ward, D.; Hamilton, S.; Jardine, T.; Pettit, N.; Tews, K.; Olley, J.; Bunn, S. Assessing the seasonal dynamics of inundation, turbidity, and aquatic vegetation in the Australian wet–dry tropics using optical remote sensing. Ecohydrology 2013, 6, 312–323. [Google Scholar] [CrossRef]
- Malthus, T.J. Chapter 9—Bio-optical Modeling and Remote Sensing of Aquatic Macrophytes. In Bio-Optical Modeling and Remote Sensing of Inland Waters; Mishra, D.R., Ogashawara, I., Gitelson, A.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 263–308. [Google Scholar]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Karim, F.; Peña-Arancibia, J.; Ticehurst, C.; Marvanek, S.; Gallant, J.; Hughes, J.; Dutta, D.; Vaze, J.; Petheram, C.; Seo, L.; et al. Floodplain Inundation Mapping and Modelling for the Fitzroy, Darwin and Mitchell Catchments; A Technical Report to the Australian Government from the CSIRO Northern Australia Water Resource Assessment, Part of the National Water Infrastructure Development Fund: Water Resource Assessments; CSIRO: Canberra, Australia, 2018.
- Stoeckl, N.; Stanley, O. Key Industries in Australia’s Tropical Savanna. Australas. J. Reg. Stud. 2007, 13, 255–286. [Google Scholar]
- Australian Government. Our North, Our Future: White Paper on Developing Northern Australia; Department of the Prime Minister and Cabinet: Canberra, Australia, 2015.
- Ndehedehe, C.E.; Onojeghuo, A.O.; Stewart-Koster, B.; Bunn, S.E.; Ferreira, V.G. Upstream flows drive the productivity of floodplain ecosystems in tropical Queensland. Ecol. Indic. 2021, 125, 107546. [Google Scholar] [CrossRef]
- Burford, M.A.; Webster, I.T.; Revill, A.T.; Kenyon, R.A.; Whittle, M.; Curwen, G. Controls on phytoplankton productivity in a wet–dry tropical estuary. Estuar. Coast. Shelf Sci. 2012, 113, 141–151. [Google Scholar] [CrossRef]
- Flood, N.; Danaher, T.; Gill, T.; Gillingham, S. An operational scheme for deriving standardised surface reflectance from landsat TM/ETM+ and SPOT HRG imagery for eastern Australia. Remote Sens. 2013, 5, 83–109. [Google Scholar] [CrossRef]
- Pohl, C.; Van Genderen, J.L. Review article Multisensor image fusion in remote sensing: Concepts, methods and applications. Int. J. Remote Sens. 2010, 19, 823–854. [Google Scholar] [CrossRef]
- Breiman, L. Classification and Regression Trees; Wadsworth International Group: Belmont, CA, USA, 1984. [Google Scholar]
- Therneau, T.; Atkinson, B. Package ‘rpart’: Recursive Partitioning and Regression Trees. R Package Version 4.1-15. 2019. Available online: https://CRAN.R-project.org/package=rpart (accessed on 6 June 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Davranche, A.; Lefebvre, G.; Poulin, B. Wetland monitoring using classification trees and SPOT-5 seasonal time series. Remote Sens. Environ. 2010, 114, 552–562. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, H.; Yang, T.; Cai, Y.; Xu, D.; An, S. Remote sensing of aquatic vegetation distribution in Taihu Lake using an improved classification tree with modified thresholds. J. Environ. Manag. 2012, 95, 98–107. [Google Scholar] [CrossRef]
- Ndehedehe, C.E.; Burford, M.A.; Stewart-Koster, B.; Bunn, S.E. Satellite-derived changes in floodplain productivity and freshwater habitats in northern Australia (1991–2019). Ecol. Indic. 2020, 114, 106320. [Google Scholar] [CrossRef]
- Feyisa, G.L.; Meilby, H.; Fensholt, R.; Proud, S.R. Automated Water Extraction Index: A new technique for surface water mapping using Landsat imagery. Remote Sens. Environ. 2014, 140, 23–35. [Google Scholar] [CrossRef]
- Gao, B.-C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- McFeeters, S.K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Sims, N.; Anstee, J.; Barron, O.; Botha, E.; Lehmann, E.; Li, L.; McVicar, T.; Paget, M.; Ticehurst, C.; Van Niel, T.; et al. Earth Observation Remote Sensing; A Technical Report to the Australian Government from the CSIRO Northern Australia Water Resource Assessment, Part of the National Water Infrastructure Development Fund: Water Resource Assessments; CSIRO: Canberra, Australia, 2016.
- Steven, M.D. The Sensitivity of the OSAVI Vegetation Index to Observational Parameters. Remote Sens. Environ. 1998, 63, 49–60. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indices: A Review of Developments and Applications. J. Sens. 2017, 2017, 1–17. [Google Scholar] [CrossRef]
- Fern, R.R.; Foxley, E.A.; Bruno, A.; Morrison, M.L. Suitability of NDVI and OSAVI as estimators of green biomass and coverage in a semi-arid rangeland. Ecol. Indic. 2018, 94, 16–21. [Google Scholar] [CrossRef]
- Pan, X.; Zhu, X.; Yang, Y.; Cao, C.; Zhang, X.; Shan, L. Applicability of Downscaling Land Surface Temperature by Using Normalized Difference Sand Index. Sci. Rep. 2018, 8, 9530. [Google Scholar] [CrossRef] [PubMed]
- Lymburner, L.; Botha, E.; Hestir, E.; Anstee, J.; Sagar, S.; Dekker, A.; Malthus, T. Landsat 8: Providing continuity and increased precision for measuring multi-decadal time series of total suspended matter. Remote Sens. Environ. 2016, 185, 108–118. [Google Scholar] [CrossRef]
- Lillesand, T.M.; Kiefer, R.W.; Chipman, J.W. Remote Sensing and Image Interpretation, 5th ed.; John Wiley: New York, NY, USA, 2004. [Google Scholar]
- Burman, P. A Comparative Study of Ordinary Cross-Validation, v-Fold Cross-Validation and the Repeated Learning-Testing Methods. Biometrika 1989, 76, 503–514. [Google Scholar] [CrossRef]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning: With Applications in R; Springer: New York, NY, USA, 2013. [Google Scholar]
- Petheram, C.; Watson, I.; Bruce, C.; Chilcott, C. Water Resource Assessment for the Mitchell Catchment; A Report to the Australian Government from the CSIRO Northern Australia Water Resource Assessment, Part of the National Water Infrastructure Development Fund: Water Resource Assessments; CSIRO: Canberra, Australia, 2018.
- Bustamante, J.; Pacios, F.; Díaz-Delgado, R.; Aragonés, D. Predictive models of turbidity and water depth in the Doñana marshes using Landsat TM and ETM+ images. J. Environ. Manag. 2009, 90, 2219–2225. [Google Scholar] [CrossRef]
- Chen, Z.; Hu, C.; Muller-Karger, F. Monitoring turbidity in Tampa Bay using MODIS/Aqua 250-m imagery. Remote Sens. Environ. 2007, 109, 207–220. [Google Scholar] [CrossRef]
- Xing, L.W.; Niu, Z.G. Mapping and analyzing China’s wetlands using MODIS time series data. Wetl. Ecol. Manag. 2019, 27, 693–710. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C.-L. Regression and Time Series Model Selection in Small Samples. Biometrika 1989, 76, 297. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H.; O’Hara, R.B. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Halekoh, U.; Højsgaard, S. A Kenward-Roger Approximation and Parametric Bootstrap Methods for Tests in Linear Mixed Models—The R Package pbkrtest. J. Stat. Softw. 2014, 59, 32. [Google Scholar] [CrossRef]
- Jachner, S.; Boogaart, G.; Petzoldt, T. Statistical Methods for the Qualitative Assessment of Dynamic Models with Time Delay (R Package qual V). J. Stat. Softw. 2007, 22, 1–30. [Google Scholar] [CrossRef]
- Knowles, J.E.; Frederick, C. merTools: Tools for Analyzing Mixed Effect Regression Models. 2020. Available online: https://CRAN.R-project.org/package=merTools (accessed on 25 September 2020).
- Dei Rossi, J.A. Prediction Intervals for Summed Totals; Rand Corporation: Santa Monica, CA, USA, 1968. [Google Scholar]
- Lane, D.M.; David, S.; Mikki, H.; Rudy, G.; Dan, O.; Zimmer, H. Introduction to Statistics; Rice University: Houston, TX, USA; University of Houston Clear Lake: Houston, TX, USA; Tufts University: Medford, MA, USA, 2013. [Google Scholar]
- Reis, V.; Hermoso, V.; Hamilton, S.K.; Bunn, S.E.; Linke, S. Conservation planning for river-wetland mosaics: A flexible spatial approach to integrate floodplain and upstream catchment connectivity. Biol. Conserv. 2019, 236, 356–365. [Google Scholar] [CrossRef]
- Malthus, T.J.; Hestir, E.L.; Dekker, A.G.; Brando, V.E. The Case for a Global Inland Water Quality Product. In Proceedings of the 2012 IEEE International Geoscience and remote sensing Symposium, Munich, Germany, 22–27 July 2012; pp. 5234–5237. [Google Scholar]
- Huang, C.; Wu, J.; Chen, Y.; Yu, J. Detecting Floodplain Inundation Frequency Using MODIS Time-Series Imagery. In Proceedings of the 2012 First International Conference on Agro- Geoinformatics (Agro-Geoinformatics), Shanghai, China, 2–4 August 2012; pp. 1–6. [Google Scholar]
- Ndehedehe, C.E.; Stewart-Koster, B.; Burford, M.A.; Bunn, S.E. Predicting hot spots of aquatic plant biomass in a large floodplain river catchment in the Australian wet-dry tropics. Ecol. Indic. 2020, 117, 106616. [Google Scholar] [CrossRef]
- Quang, N.H.; Sasaki, J.; Higa, H.; Huan, N.H. Spatiotemporal variation of turbidity based on landsat 8 OLI in Cam Ranh Bay and Thuy Trieu Lagoon, Vietnam. Water 2017, 9, 570. [Google Scholar] [CrossRef]
- Bid, S.; Siddique, G. Identification of seasonal variation of water turbidity using NDTI method in Panchet Hill Dam, India. Modeling Earth Syst. Environ. 2019, 5, 1179–1200. [Google Scholar] [CrossRef]
- Pham, Q.; Ha, N.; Pahlevan, N.; Oanh, L.; Nguyen, T.; Nguyen, N. Using Landsat-8 Images for Quantifying Suspended Sediment Concentration in Red River (Northern Vietnam). Remote Sens. 2018, 10, 1841. [Google Scholar] [CrossRef]
- McCarthy, M.A.; Masters, P. Profiting from prior information in Bayesian analyses of ecological data. J. Appl. Ecol. 2005, 42, 1012–1019. [Google Scholar] [CrossRef]
- Hinojosa-Garro, D.; Mason, C.F.; Underwood, G.J.C. Influence of macrophyte spatial architecture on periphyton and macroinvertebrate community structure in shallow water bodies under contrasting land management. Fundam. Appl. Limnol. 2010, 177, 19–37. [Google Scholar] [CrossRef]
- Leibowitz, S.G. Isolated wetlands and their functions: An ecological perspective. Wetlands 2003, 23, 517–531. [Google Scholar] [CrossRef]
- Sparks, R.E. Need for ecosystem management of large rivers and their floodplains—These phenomenally productive ecosystems produce fish and wildlife and preserve species. Bioscience 1995, 45, 168–182. [Google Scholar] [CrossRef]
- Pettit, N.E.; Naiman, R.J.; Warfe, D.M.; Jardine, T.D.; Douglas, M.M.; Bunn, S.E.; Davies, P.M. Productivity and Connectivity in Tropical Riverscapes of Northern Australia: Ecological Insights for Management. Ecosystems 2017, 20, 492–514. [Google Scholar] [CrossRef]
- Furst, D.J.; Aldridge, K.T.; Shiel, R.J.; Ganf, G.G.; Mills, S.; Brookes, J.D. Floodplain connectivity facilitates significant export of zooplankton to the main River Murray channel during a flood event. Inland Waters 2014, 4, 413–424. [Google Scholar] [CrossRef]
- Duan, H.; Ma, R.; Xu, J.; Zhang, Y.; Zhang, B. Comparison of different semi-empirical algorithms to estimate chlorophyll-a concentration in inland lake water. Environ. Monit. Assess. 2010, 170, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Gosselain, V.R.; Hudon, C.; Cattaneo, A.; Gagnon, P.; Planas, D.; Rochefort, D. Physical variables driving epiphytic algal biomass in a dense macrophyte bed of the St. Lawrence River (Quebec, Canada). Hydrobiologia 2005, 534, 11–22. [Google Scholar] [CrossRef]
- Burford, M.A.; Valdez, D.; Curwen, G.; Faggotter, S.J.; Ward, D.P.; O’Brien, K.R. Inundation of saline supratidal mudflats provides an important source of carbon and nutrients in an aquatic system. Mar. Ecol. Prog. Ser. 2016, 545, 21–33. [Google Scholar] [CrossRef]
- Saravia, L.A.; Momo, F.; Boffi Lissin, L.D. Modelling periphyton dynamics in running water. Ecol. Model. 1998, 114, 35–47. [Google Scholar] [CrossRef]
- Rowan, G.S.L.; Kalacska, M. A Review of Remote Sensing of Submerged Aquatic Vegetation for Non-Specialists. Remote Sens. 2021, 13, 623. [Google Scholar] [CrossRef]
- Tockner, K.; Pusch, M.; Borchardt, D.; Lorang, M.S. Multiple stressors in coupled river-floodplain ecosystems. Freshw. Biol. 2010, 55, 135–151. [Google Scholar] [CrossRef]
- Mukherjee, K.; Pal, S. Hydrological and landscape dynamics of floodplain wetlands of the Diara region, Eastern India. Ecol. Indic. 2021, 121, 106961. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landsat 8 OLI | SPOT 7 | |
|---|---|---|
| Date of Acquisition | 25 April 2018 | 25 April 2018 |
| Spatial resolution | 30 m | 6 m |
| Spectral bands: | Wavelengths (nm) | |
| Coastal Aerosol (CA) | 435–451 | NA |
| Blue | 452–512 | 450–520 |
| Green | 533–590 | 530–590 |
| Red | 636–673 | 625–695 |
| Near Infrared (NIR) | 851–879 | 760–890 |
| Shortwave NIR 1 (SWIR 1) | 1566–1651 | NA |
| Shortwave NIR 2 (SWIR 2) | 2107–2294 | NA |
| Spectral Indices | Formula | Reference |
|---|---|---|
| Automated water extraction index (AWEI) | 4(Green − SWIR) − 0.25(0.25 × NIR + 2.75 × SWIR) | [41] |
| Normalized difference water index (NDWI) | (Green − NIR)/(Green + NIR) | [43] |
| Modified normalized difference water index (MNDWI) | (Green − SWIR)/(Green + SWIR) | [44] |
| Optimized soil adjusted vegetation index (OSAVI) | (NIR − Red)/(NIR + Red + 0.16) | [46] |
| Normalized difference vegetation index (NDVI) | (NIR − Red)/(NIR + Red) | [42] |
| Normalized difference sand index (NDSI) | (Red − Coastal aerosol)/(Red − Coastal aerosol) | [49] |
| Total suspended matter (TSM) | 3957 × (TSM_index) ^ 1.6436; TSM_index = (green_band/10,000 + red_band/10,000)/2 | [50] |
| Turbidity Model | PP Model | ||||||
|---|---|---|---|---|---|---|---|
| Parameter Estimate | Std. Error | p Value | Parameter Estimate | Std. Error | p Value | ||
| Intercept | 2.898486 | 0.413495 | <0.001 | 3.1256 | 0.7505 | <0.001 | |
| Habitat Type | Floating + Emergent | 0.434920 | 0.176097 | 0.02 | −1.7609 | 0.4182 | <0.001 |
| Green Band | −0.004759 | 0.002077 | 0.04 | - | - | ||
| Red Band | 0.004176 | 0.001745 | 0.04 | - | - | ||
| Log (turbidity-NTU) | - | - | 0.7435 | 0.2605 | 0.02 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinari, B.; Stewart-Koster, B.; Malthus, T.J.; Bunn, S.E. Assessing Spatial Variation in Algal Productivity in a Tropical River Floodplain Using Satellite Remote Sensing. Remote Sens. 2021, 13, 1710. https://doi.org/10.3390/rs13091710
Molinari B, Stewart-Koster B, Malthus TJ, Bunn SE. Assessing Spatial Variation in Algal Productivity in a Tropical River Floodplain Using Satellite Remote Sensing. Remote Sensing. 2021; 13(9):1710. https://doi.org/10.3390/rs13091710
Chicago/Turabian StyleMolinari, Bianca, Ben Stewart-Koster, Tim J. Malthus, and Stuart E. Bunn. 2021. "Assessing Spatial Variation in Algal Productivity in a Tropical River Floodplain Using Satellite Remote Sensing" Remote Sensing 13, no. 9: 1710. https://doi.org/10.3390/rs13091710
APA StyleMolinari, B., Stewart-Koster, B., Malthus, T. J., & Bunn, S. E. (2021). Assessing Spatial Variation in Algal Productivity in a Tropical River Floodplain Using Satellite Remote Sensing. Remote Sensing, 13(9), 1710. https://doi.org/10.3390/rs13091710