
Monitoring Tamarix Changes Using WorldView-2 Satellite Imagery in Grand Canyon National Park, Arizona



Abstract
1. Introduction
2. Materials and Methods
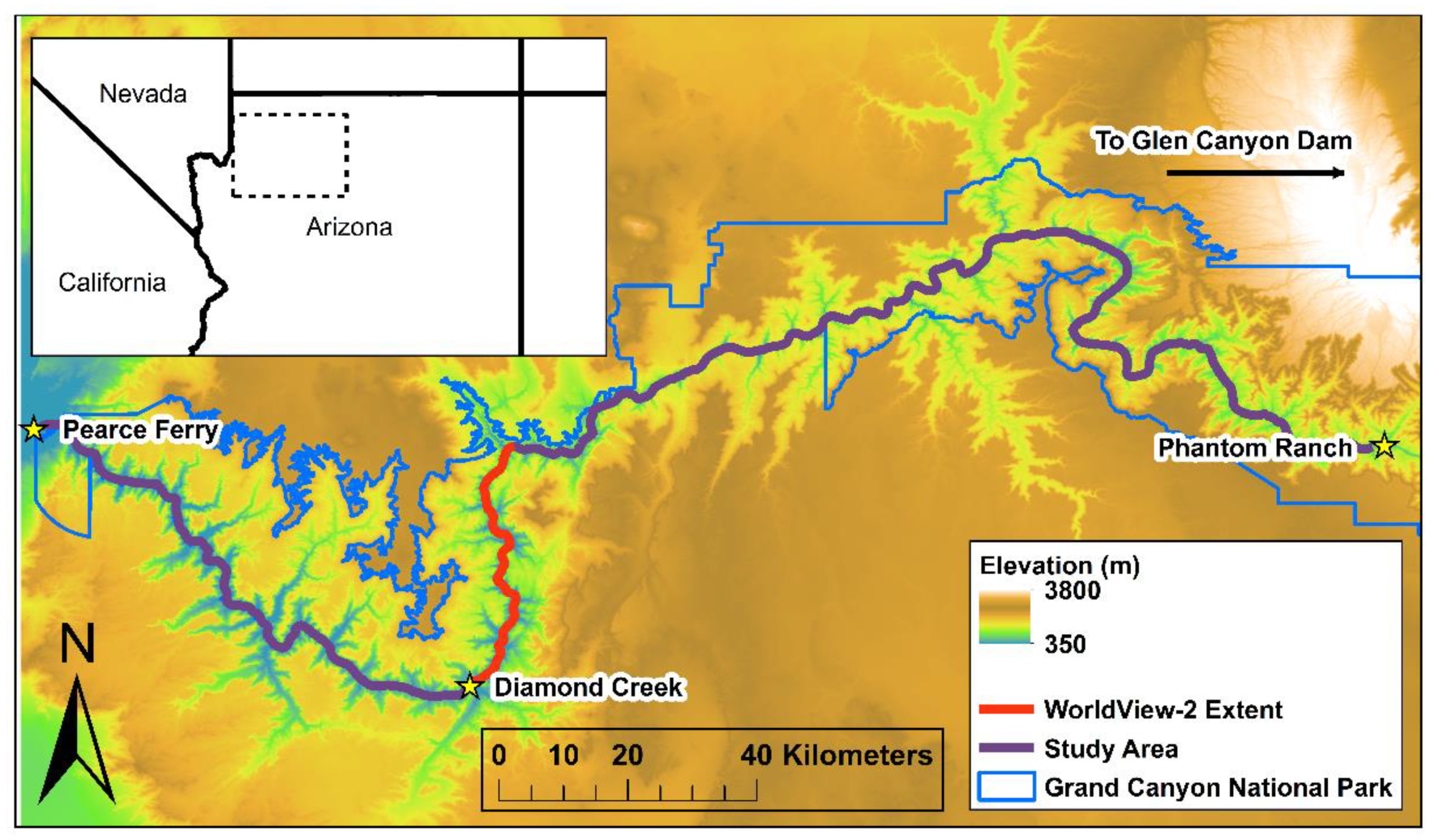
2.1. Field Observations
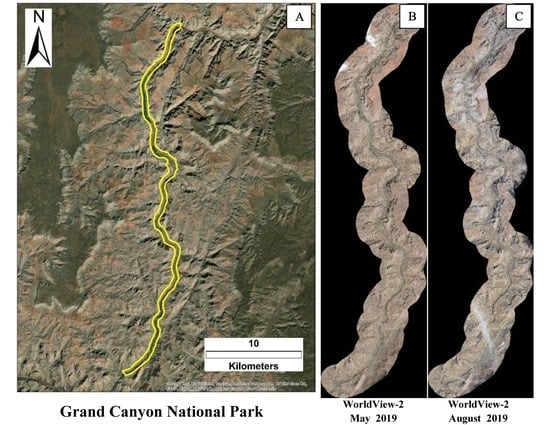
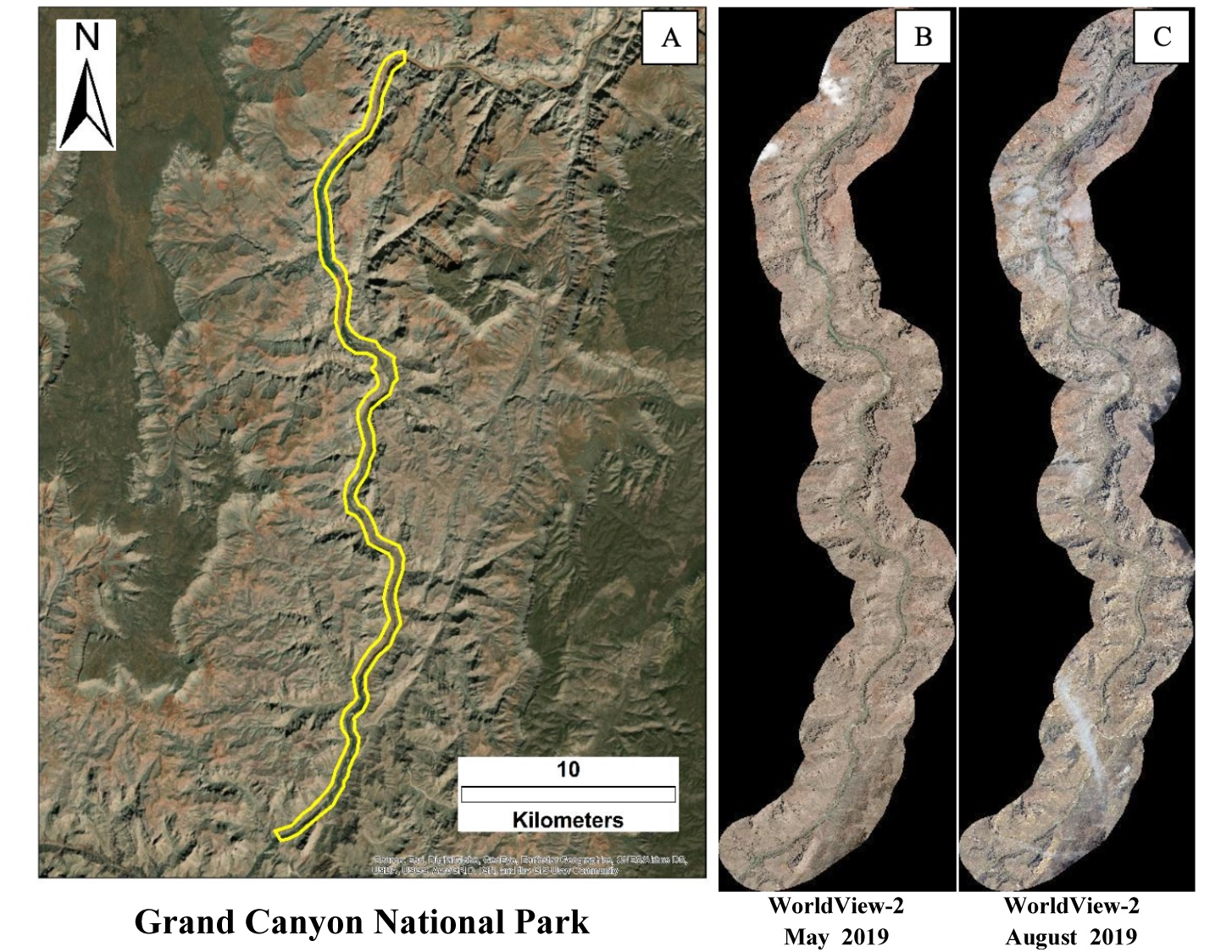
2.2. WorldView-2 Satellite Images and Analysis
2.3. Accuracy Assessment
2.4. Change Detection
3. Results
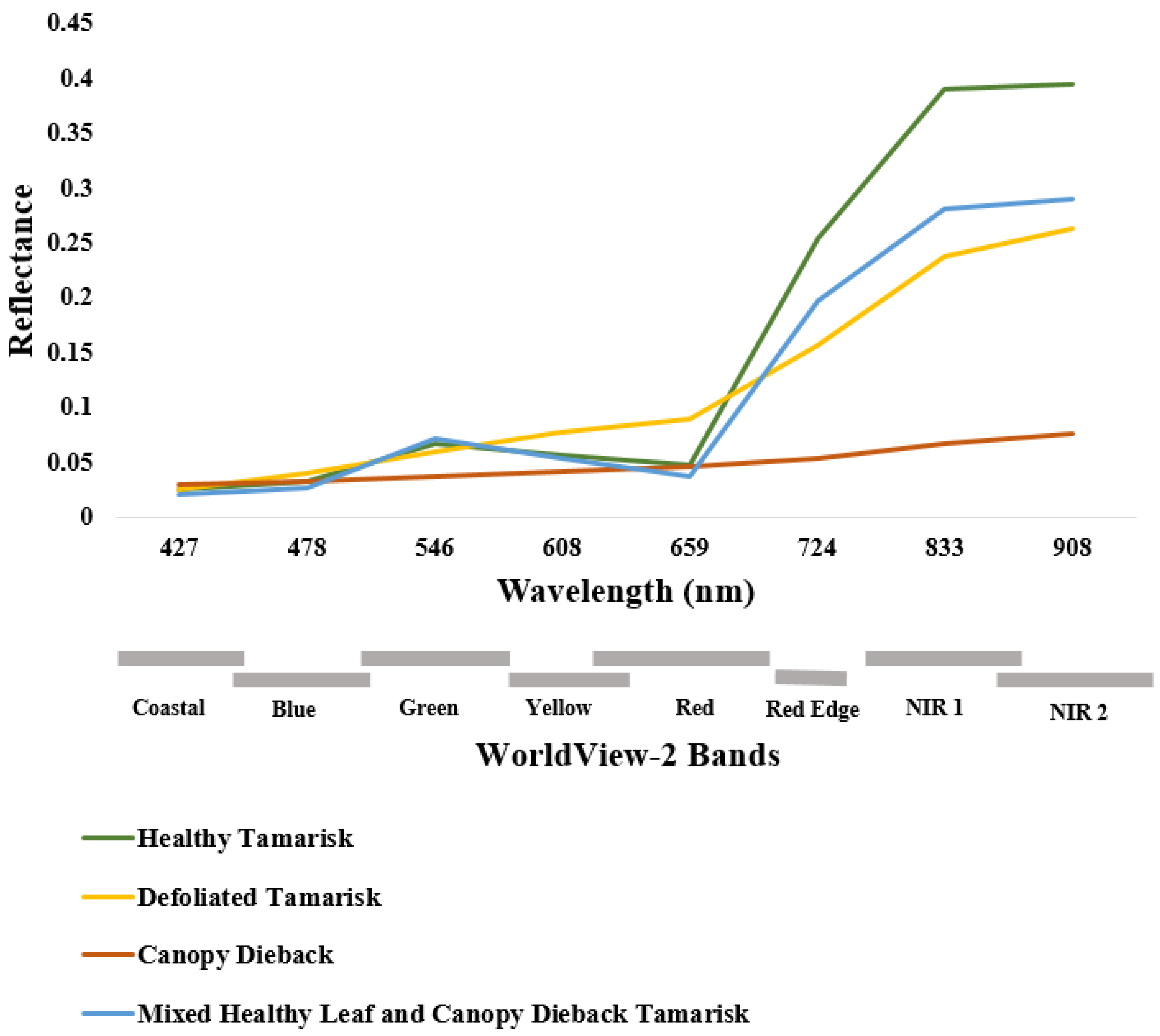
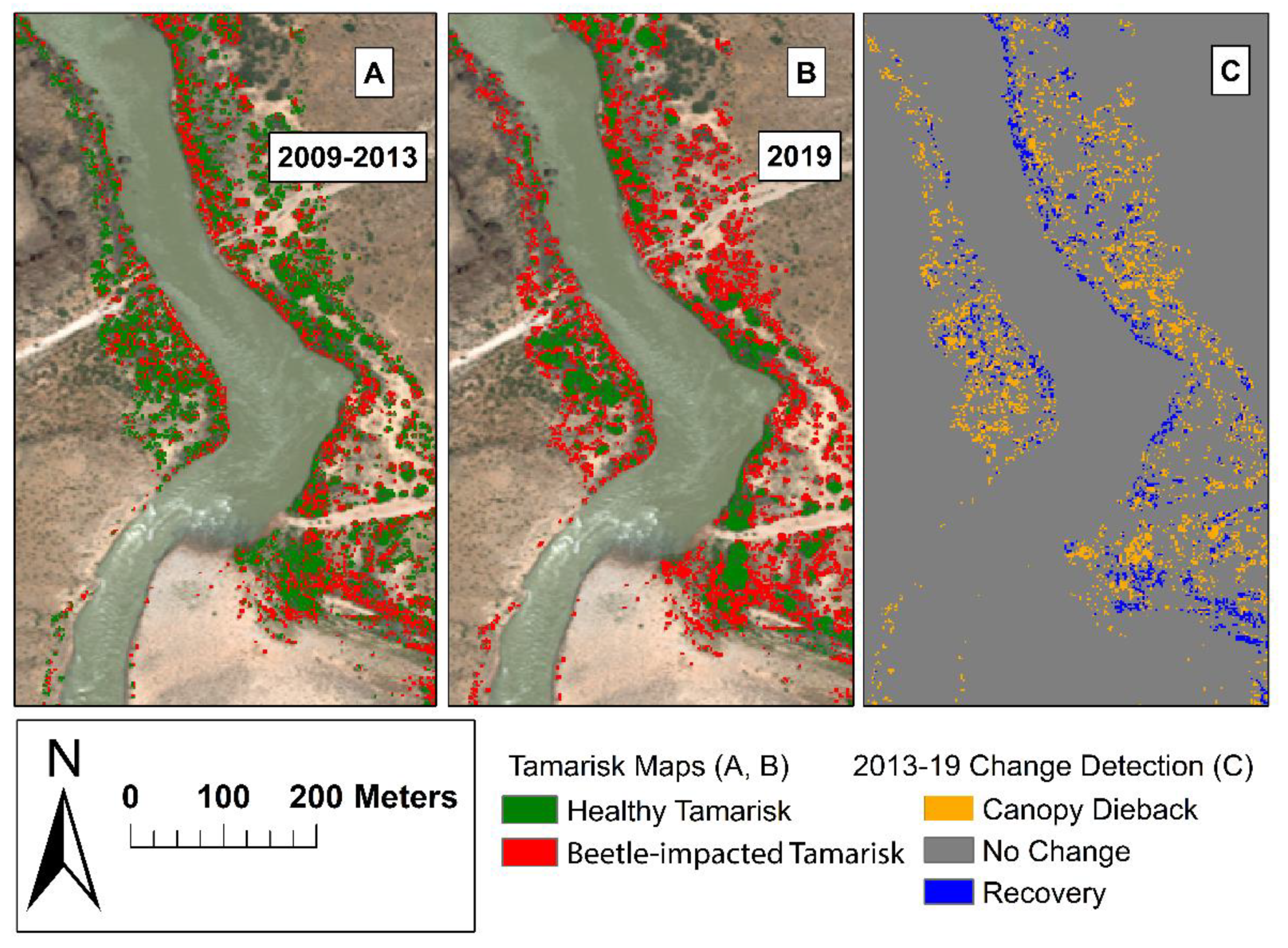
3.1. Tamarisk Detection
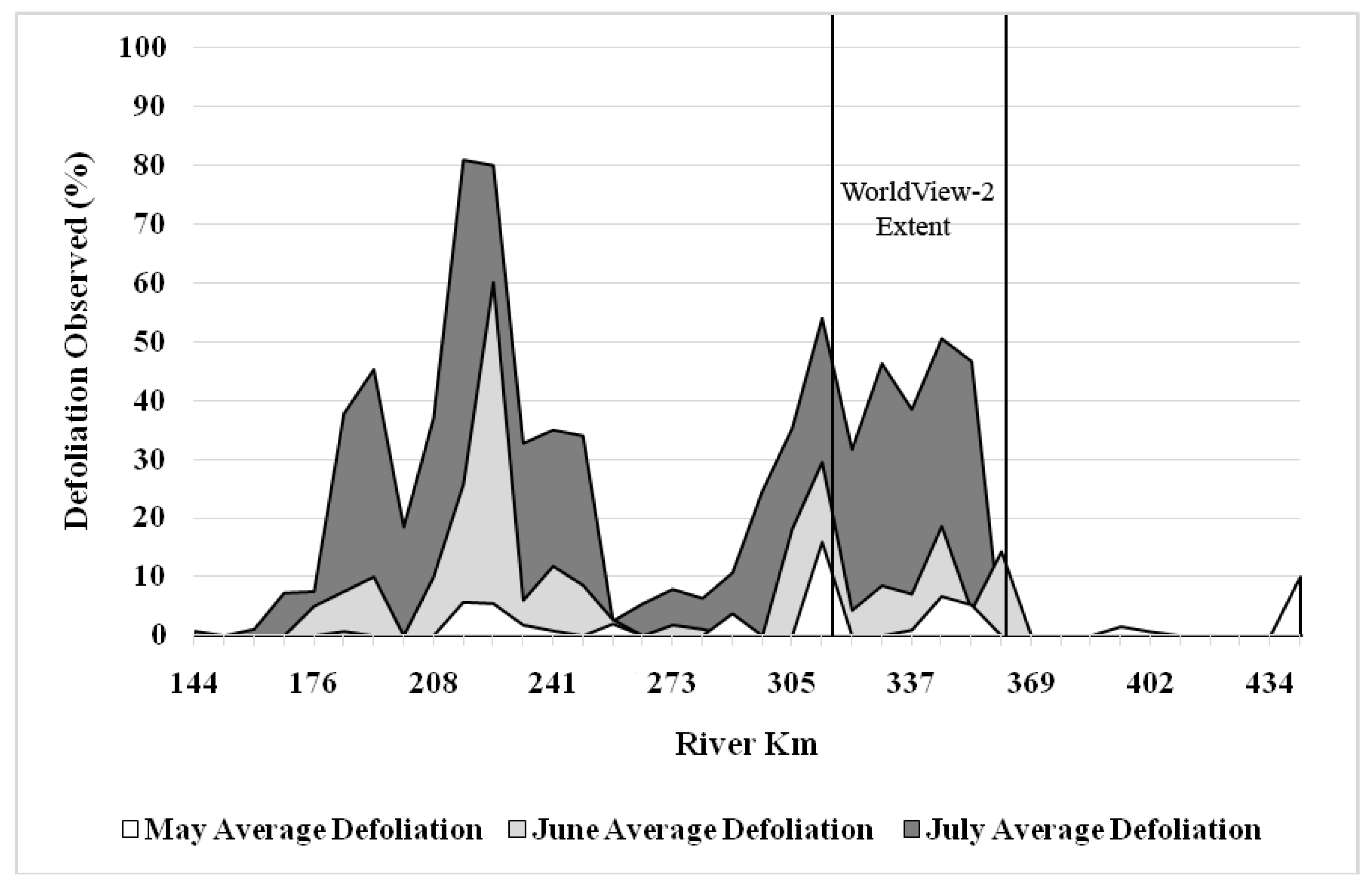
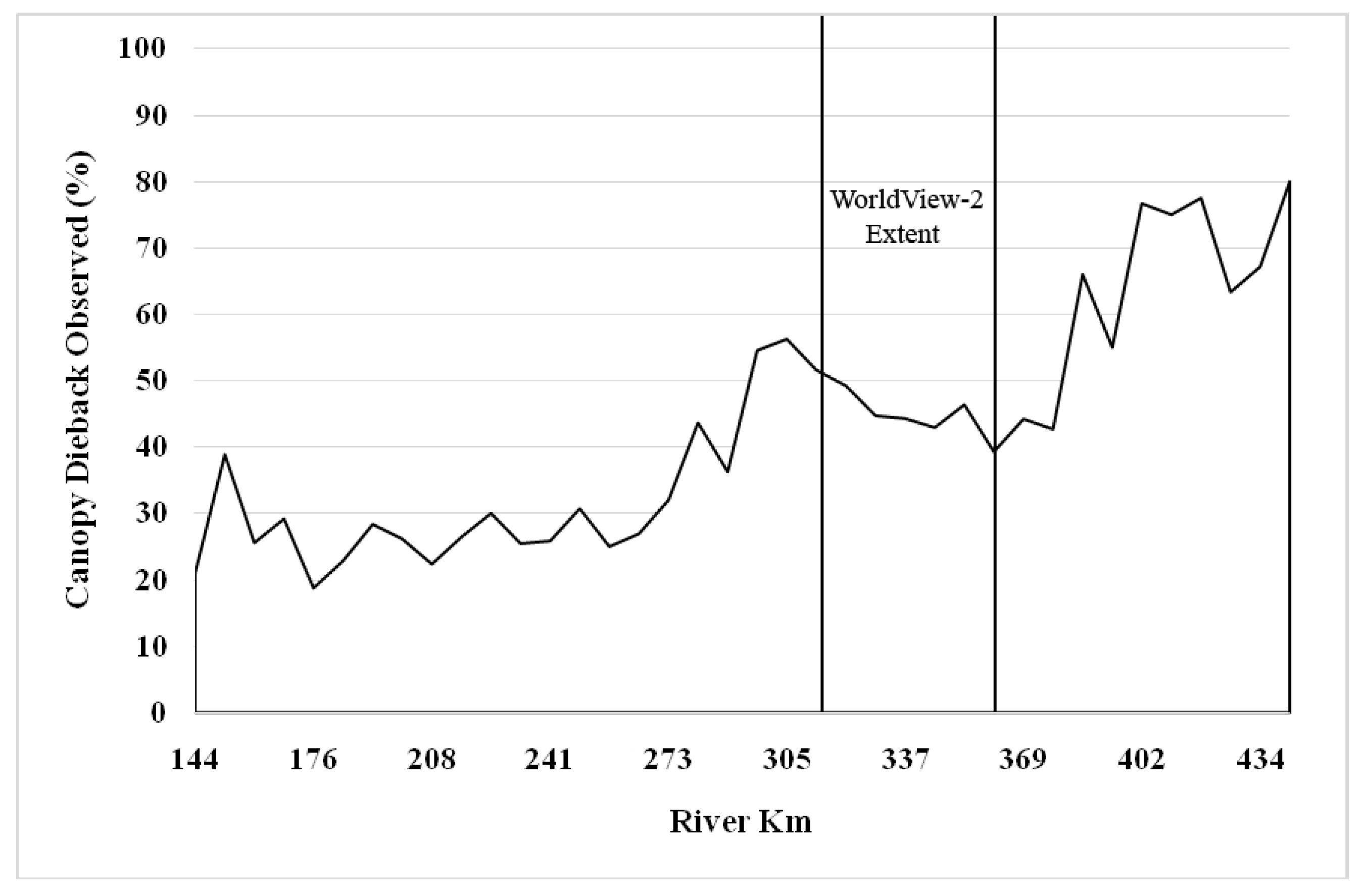
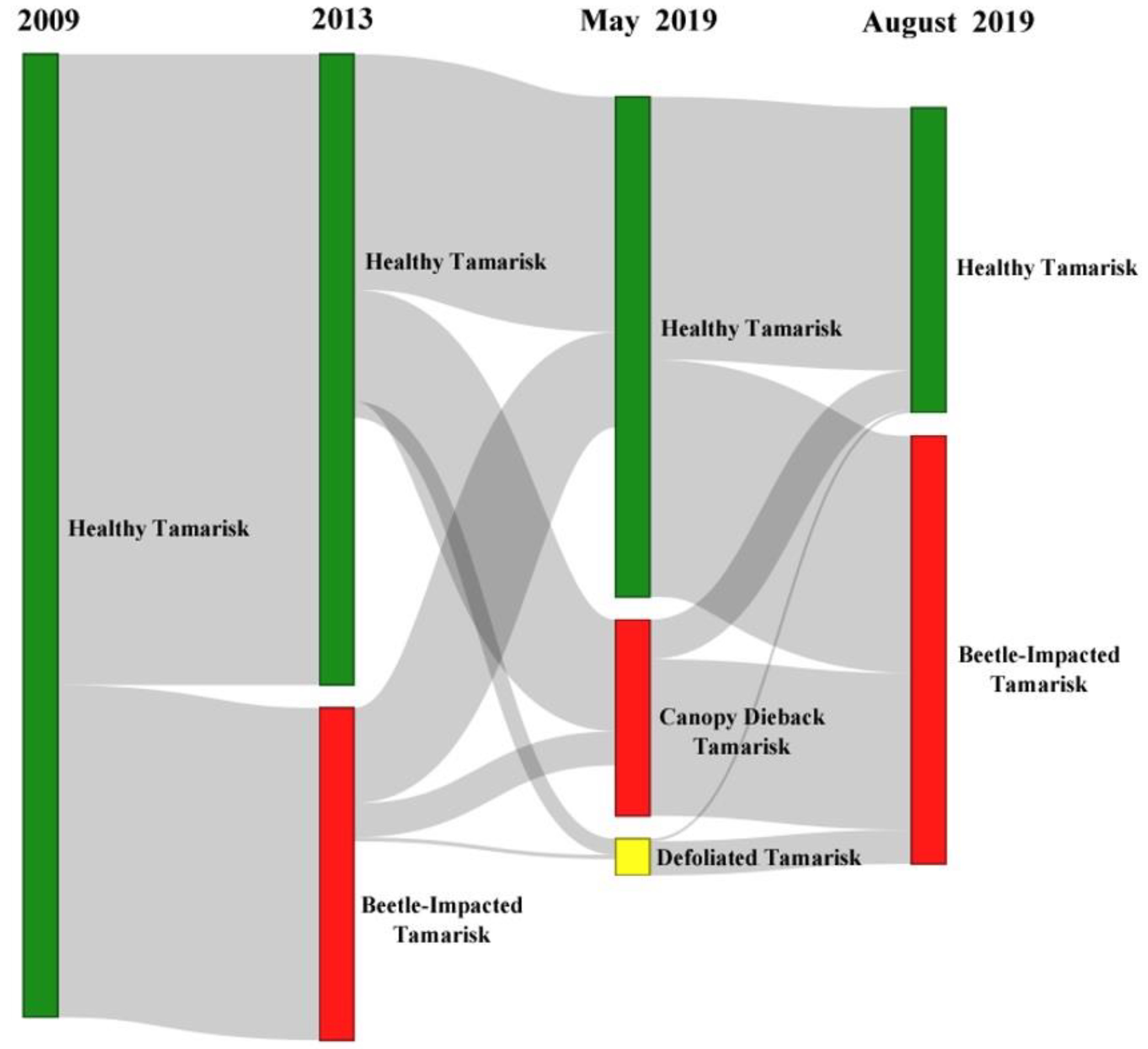
3.2. Change Detection
4. Discussion
4.1. WorldView-2 Image Classification of Tamarisk Distribution, Canopy Dieback, and Defoliation
4.2. Seasonal and Multi-Year Change Detection
4.3. Ecological Implications
4.4. Implications for Future Remote Sensing Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brock, J.H. Tamarix spp. (Salt Cedar), an invasive exotic woody plant in arid and semi-arid riparian habitats of Western USA. In Ecology and Management of Invasive Riverside Plants; de Waal, L.C., Child, L.E., Wade, P.M., Brock, J.H., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1994; pp. 27–43. [Google Scholar]
- Deloach, C.; A Lewis, P.; Herr, J.C.; I Carruthers, R.; Tracy, J.L.; Johnson, J. Host specificity of the leaf beetle, Diorhabda elongata deserticola (Coleoptera: Chrysomelidae) from Asia, a biological control agent for saltcedars (Tamarix: Tamaricaceae) in the Western United States. Biol. Control 2003, 27, 117–147. [Google Scholar] [CrossRef]
- Sueki, S.; Acharya, K.; Huntington, J.; Liebert, R.; Healey, J.; Jasoni, R.; Young, M. Defoliation effects of Diorhabda carinulata on tamarisk evapotranspiration and groundwater levels. Ecohydrology 2015, 8, 1560–1571. [Google Scholar] [CrossRef]
- Zavaleta, E. The Economic Value of Controlling an Invasive Shrub. J. Hum. Environ. 2000, 29, 462–467. [Google Scholar] [CrossRef]
- Liebert, R.; Huntington, J.; Morton, C.; Sueki, S.; Acharya, K. Reduced evapotranspiration from leaf beetle induced tamarisk defoliation in the Lower Virgin River using satellite-based energy balance. Ecohydrology 2015, 9, 179–193. [Google Scholar] [CrossRef]
- Nagler, P.L.; Barreto-Muñoz, A.; Chavoshi Borujeni, S.; Jarchow, C.J.; Gómez-Sapiens, M.M.; Nouri, H.; Herrmann, S.M.; Didan, K. Ecohydrological responses to surface flow across borders: Two decades of changes in vegetation greenness and water use in the riparian corridor of the Colorado River delta. Hydrol. Process. 2020, 34, 4851–4883. [Google Scholar] [CrossRef]
- Snyder, K.A.; Scott, R.L. Longer term effects of biological control on tamarisk evapotranspiration and carbon dioxide exchange. Hydrol. Process. 2020, 34, 223–236. [Google Scholar] [CrossRef]
- Owens, M.K.; Moore, G.W. Saltcedar Water Use: Realistic and Unrealistic Expectations. Rangel. Ecol. Manag. 2007, 60, 553–557. [Google Scholar] [CrossRef]
- Meinhardt, K.A.; Gehring, C.A. Disrupting mycorrhizal mutualisms: A potential mechanism by which exotic tamarisk outcompetes native cottonwoods. Ecol. Appl. 2012, 22, 532–549. [Google Scholar] [CrossRef]
- Merkel, D.L.; Hopkins, H.H. Life History of Salt Cedar (Tamarix gallica L.). Trans. Kansas Acad. Sci. 1957, 60, 360–369. [Google Scholar] [CrossRef]
- Glenn, E.P.; Nagler, P.L. Comparative ecophysiology of Tamarix ramosissima and native trees in western U.S. riparian zones. J. Arid. Environ. 2005, 61, 419–446. [Google Scholar] [CrossRef]
- Gaskin, J.F.; Schaal, B.A. Hybrid Tamarix widespread in U.S. invasion and undetected in native Asian range. Proc. Natl. Acad. Sci. USA 2002, 99, 11256–11259. [Google Scholar] [CrossRef] [PubMed]
- Bedford, A.; Sankey, T.T.; Sankey, J.B.; Durning, L.; Ralston, B.E. Remote sensing of tamarisk beetle (Diorhabda carinulata) impacts along 412 km of the Colorado River in the Grand Canyon, Arizona, USA. Ecol. Indic. 2018, 89, 365–375. [Google Scholar] [CrossRef]
- Bedford, A.; Sankey, T.T.; Sankey, J.B.; Durning, L.E.; Ralston, B.E. Remote Sensing Derived Maps of Tamarisk (2009) and Beetle Impacts (2013) along 412 km of the Colorado River in the Grand Canyon, Arizona; U.S. Geological Survey data release; U.S. Geological Survey: Flagstaff, AZ, USA, 2018. [CrossRef]
- Nagler, P.L.; Nguyen, U.; Bateman, H.L.; Jarchow, C.J.; Glenn, E.P.; Waugh, W.J.; Van Riper, C., III. Northern tamarisk beetle (Diorhabda carinulata) and tamarisk (Tamarix spp.) interactions in the Colorado River basin. Restor. Ecol. 2018, 26, 348–359. [Google Scholar] [CrossRef]
- Sankey, T.T.; Sankey, J.B.; Horne, R.; Bedford, A. Remote Sensing of Tamarisk Biomass, Insect Herbivory, and Defoliation: Novel Methods in the Grand Canyon Region, Arizona. Photogramm. Eng. Remote. Sens. 2016, 82, 645–652. [Google Scholar] [CrossRef]
- Hennessy, M. Colorado River Water Rights: Property Rights in Transition. Univ. Chic. Law Rev. 2004, 71, 1661–1687. [Google Scholar]
- DeLoach, C.J.; Carruthers, R.I.; Dudley, T.L.; Eberts, D.; Kazmer, D.J.; Knutson, A.E.; Bean, D.W.; Knight, J.; Lewis, P.A.; Milbrath, L.R.; et al. First results for control of saltcedar (Tamarix spp.) in the open field in the western United States. In Proceedings of the XI International Symposium on Biological Control of Weeds, Entomology, Canberra, Australia, 27 April–2 May 2003; Cullen, W., Briese, D.T., Kriticos, D.J., Lonsdale, W.M., Morin, L., Scott, J.K., Eds.; 2004; pp. 505–513. [Google Scholar]
- RiversEdge West Yearly Distribution (2007–2019) of Tamarisk Beetle (Diorhabda spp.). Available online: https://riversedgewest.org/sites/default/files/files/2019_Annual_Map_New_Color.pdf (accessed on 5 May 2020).
- Berro, A.M.; Evans, S.; Ferguson, E.M.; Royer, T.A. Acceptability of Diorhabda carinulata, an Introduced Biological Control Agent ofTamarix, as Prey for Selected Resident Native Arthropod Predators. Southwest. Èntomologist 2017, 42, 665–676. [Google Scholar] [CrossRef]
- Moran, P.J.; Deloach, C.J.; Dudley, T.L.; Sanabria, J. Open field host selection and behavior by tamarisk beetles (Diorhabda spp.) (Coleoptera: Chrysomelidae) in biological control of exotic saltcedars (Tamarix spp.) and risks to non-target athel (T. aphylla) and native Frankenia spp. Biol. Control 2009, 50, 243–261. [Google Scholar] [CrossRef]
- Bloodworth, B.; Shafroth, P.; Sher, A.; Manners, R.; Bean, D.; Johnson, M.J.; Hinojosa-Huerta, O. Tamarisk Beetle (Diorhabda spp.) in the Colorado River Basin: Synthesis of an Expert Panel Forum; Scientific and Technical Report no 1; Colorado Mesa University, Ruth Powell Hutchins Water Center: Grand Junction, CO, USA, 2016; 19p, Available online: https://riversedgewest.org/sites/default/files/files/Tamarisk_Beetle_Expert_Panel_Report.pdf (accessed on 5 May 2020).
- A Lewis, P.; Deloach, C.; E Knutson, A.; Tracy, J.L.; O Robbins, T. Biology of Diorhabda elongata deserticola (Coleoptera: Chrysomelidae), an Asian leaf beetle for biological control of saltcedars (Tamarix spp.) in the United States. Biol. Control 2003, 27. [Google Scholar] [CrossRef]
- Jamison, L.R.; Johnson, M.J.; Bean, D.W.; Van Riper, C. Phenology and Abundance of Northern Tamarisk Beetle, Diorhabda carinulata Affecting Defoliation of Tamarix. Southwest. Èntomologist 2018, 43, 571–584. [Google Scholar] [CrossRef]
- Nagler, P.L.; Brown, T.; Hultine, K.R.; Van Riper, C.; Bean, D.W.; Dennison, P.E.; Murray, R.S.; Glenn, E.P. Regional scale impacts of Tamarix leaf beetles (Diorhabda carinulata) on the water availability of western U.S. rivers as determined by multi-scale remote sensing methods. Remote. Sens. Environ. 2012, 118, 227–240. [Google Scholar] [CrossRef]
- Dudley, T.L.; Bean, D.W. Tamarisk biocontrol, endangered species effects and resolution of conflict through riparian res-toration. Biol. Control 2012, 57, 331–347. [Google Scholar]
- Kennard, D.; Louden, N.; Gemoets, D.; Ortega, S.; González, E.; Bean, D.; Cunningham, P.; Johnson, T.; Rosen, K.; Stahlke, A. Tamarix dieback and vegetation patterns following release of the northern tamarisk beetle (Diorhabda carinulata) in western Colorado. Biol. Control 2016, 101, 114–122. [Google Scholar] [CrossRef]
- Jarchow, C.J.; Waugh, W.J.; Didan, K.; Barreto-Muñoz, A.; Herrmann, S.; Nagler, P.L. Vegetation-groundwater dynamics at a former uranium mill site following invasion of a biocontrol agent: A time series analysis of Landsat normalized difference vegetation index data. Hydrol. Process. 2020, 34, 2739–2749. [Google Scholar] [CrossRef]
- Eckberg, J.R.; Rice, N.A. Northern tamarisk beetle (Diorhabda Carinulata) effects on established tamarisk-feeding inverte-brate populations along the Las Vegas Wash, Clark County, Nevada. Southwest. Nat. 2016, 61, 101–107. [Google Scholar] [CrossRef]
- Bateman, H.L.; Merritt, D.M.; Glenn, E.P.; Nagler, P.L. Indirect effects of biocontrol of an invasive riparian plant (Tamarix) alters habitat and reduces herpetofauna abundance. Biol. Invasions 2015, 17, 87–97. [Google Scholar] [CrossRef]
- Hatten, J. A Satellite Model of Southwestern Willow Flycatcher (Empidonax traillii extimus) Breeding Habitat and a Simulation of Potential Effects of Tamarisk Leaf Beetles (Diorhabda spp.), Southwestern United States; U.S. Geological Survey Open-File Report 2016—1120; U.S. Geological Survey: Reston, VA, USA, 2016; 99p. [CrossRef]
- Paxton, E.H.; Theimer, T.C.; Sogge, M.K. Tamarisk Biocontrol Using Tamarisk Beetles: Potential Consequences for Riparian Birds in the Southwestern United States. Condor 2011, 113, 255–265. [Google Scholar] [CrossRef]
- Carter, G.A.; Lucas, K.L.; Blossom, G.A.; Lassitter, C.L.; Holiday, D.M.; Mooneyhan, D.S.; Fastring, D.R.; Holcombe, T.R.; Griffith, J.A. Remote Sensing and Mapping of Tamarisk along the Colorado River, USA: A Comparative Use of Summer-Acquired Hyperion, Thematic Mapper and QuickBird Data. Remote. Sens. 2009, 1, 318–329. [Google Scholar] [CrossRef]
- Groeneveld, D.P.; Watson, R.P. Near-infrared discrimination of leafless saltcedar in wintertime Landsat TM. Int. J. Remote Sens. 2008, 29, 3577–3588. [Google Scholar] [CrossRef]
- Ji, W.; Wang, L. Phenology-guided saltcedar (Tamarix spp.) mapping using Landsat TM images in western U.S. Remote Sens. Environ. 2016, 173, 29–38. [Google Scholar] [CrossRef]
- Narumalani, S.; Mishra, D.R.; Burkholder, J.; Merani, P.B.; Willson, G. A Comparative Evaluation of ISODATA and Spectral Angle Mapping for the Detection of Saltcedar Using Airborne Hyperspectral Imagery. Geocarto Int. 2006, 21, 59–66. [Google Scholar] [CrossRef]
- Ge, S.; Carruthers, R.; Gong, P.; Herrera, A. Texture Analysis for Mapping Tamarix parviflora Using Aerial Photographs along the Cache Creek, California. Environ. Monit. Assess. 2004, 114, 65–83. [Google Scholar] [CrossRef]
- Nagler, P.L.; Pearlstein, S.; Glenn, E.P.; Brown, T.B.; Bateman, H.L.; Bean, D.W.; Hultine, K.R. Rapid dispersal of saltcedar (Tamarix spp.) biocontrol beetles (Diorhabda carinulata) on a desert river detected by phenocams, MODIS imagery and ground observations. Remote. Sens. Environ. 2014, 140, 206–219. [Google Scholar] [CrossRef]
- Snyder, K.A.; Scott, R.L.; McGwire, K. Multiple year effects of a biological control agent (Diorhabda carinulata) on Tamarix (saltcedar) ecosystem exchanges of carbon dioxide and water. Agric. For. Meteorol. 2012, 146, 161–169. [Google Scholar] [CrossRef]
- Dennison, P.E.; Nagler, P.L.; Hultine, K.R.; Glenn, E.P.; Ehleringer, J.R. Remote monitoring of tamarisk defoliation and evapo-transpiration following saltcedar leaf beetle attack. Remote Sens. Environ. 2009, 113, 1462–1472. [Google Scholar] [CrossRef]
- Ji, W.; Wang, L.; Knutson, A.E. Detection of the spatiotemporal patterns of beetle-induced tamarisk ( Tamarix spp.) defoliation along the Lower Rio Grande using Landsat TM images. Remote Sens. Environ. 2017, 193, 76–85. [Google Scholar] [CrossRef]
- Durning, L.E.; Sankey, J.B.; Davis, P.A.; Sankey, T.T. Four-Band Image Mosaic of the Colorado River Corridor Downstream of Glen Canyon Dam in Arizona, Derived from the May 2013 Airborne Image Acquisition; U.S. Geological Survey Data Series 1027; U.S. Geological Survey: Reston, VA, USA, 2016. [CrossRef]
- Durning, L.E.; Sankey, J.B.; Davis, P.A.; Sankey, T.T. Four Band Image Mosaic of the Colorado River Corridor in Arizona—2013, including Accuracy Assessment Data; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2016. [CrossRef]
- Kruse, F.A.; Lefkoff, A.B.; Boardman, J.B.; Heidebrecht, K.B.; Shapiro, A.T.; Barloon, P.J.; Goetz, A.F.H. The Spectral Image Processing System (SIPS)—Interactive Visualization and Analysis of Imaging spectrometer Data. Remote Sens. Environ. 1993, 44, 145–163. [Google Scholar] [CrossRef]
- Durning, L.E.; Sankey, J.B.; Chain, G.R.; Sankey, T.T. Riparian Vegetation Classification of the Colorado River Corridor, Grand Canyon, Arizona, 2013—Data; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2017. [CrossRef]
- Elkind, K.; Sankey, T.T.; Munson, S.M.; Aslan, C.E. Invasive buffelgrass detection using high-resolution satellite and UAV imagery on Google Earth Engine. Remote Sens. Ecol. Conserv. 2019. [Google Scholar] [CrossRef]
- Sankey, T.; Dickson, B.; Sesnie, S.; Wang, O.; Olsson, A.; Zachmann, L. WorldView-2 High Spatial Resolution Improves Desert Invasive Plant Detection. Photogramm. Eng. Remote Sens. 2014, 80, 885–893. [Google Scholar] [CrossRef]
- Durning, L.E.; Sankey, J.B.; Bedford, A.; Sankey, T.T. Riparian Species Vegetation Classification Data for the Colorado River within Grand Canyon Derived from 2013 Airborne Imagery; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2018. [CrossRef]
- Kasprak, A.; Sankey, J.B.; Butterfield, B.J. Future regulated flows of the Colorado River in Grand Canyon foretell decreased areal extent of sediment and increases in riparian vegetation. Environ. Res. Lett. 2021, 16, 014029. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Palmquist, E.; Ralston, B. Hydrological regime and climate interactively shape riparian vegetation composition along the Colorado River, Grand Canyon. Appl. Veg. Sci. 2018, 21, 572–583. [Google Scholar] [CrossRef]
- Darrah, A.J.; van Riper, C. Riparian bird density decline in response to biocontrol of Tamarix from riparian ecosystems along the Dolores River in SW Colorado, USA. Biol. Invasions 2018, 20, 709–720. [Google Scholar] [CrossRef]
- Harms, R.S.; Hiebert, R.D. Vegetation Response Following Invasive Tamarisk (Tamarix spp.) Removal and Implications for Riparian Restoration. Restor. Ecol. 2006, 14, 461–472. [Google Scholar] [CrossRef]
- Ostoja, S.M.; Brooks, M.L.; Dudley, T.; Lee, S.R. Short-Term Vegetation Response Following Mechanical Control of Saltcedar (Tamarix spp.) on the Virgin River, Nevada, USA. Invasive Plant Sci. Manag. 2014, 7, 310–319. [Google Scholar] [CrossRef]
- González, E.; Sher, A.A.; Anderson, R.M.; Bay, R.F.; Bean, D.W.; Bissonnete, G.J.; Cooper, D.J.; Dohrenwend, K.; Eichhorst, K.D.; El Waer, H.; et al. Secondary invasions of noxious weeds associated with control of invasive Tamarix are frequent, idiosyncratic and persistent. Biol. Conserv. 2017, 213, 106–114. [Google Scholar] [CrossRef]
- González, E.; Shafroth, P.B.; Lee, S.R.; Ostoja, S.M.; Brooks, M.L. Combined effects of biological control of an invasive shrub and fluvial processes on riparian vegetation dynamics. Biol. Invasions 2020, 22, 2339–2356. [Google Scholar] [CrossRef]
- O’Neill, M.; Ustin, S.L.; Hager, S.; Root, R. Mapping the distribution of leafy spurge at Theodore Roosevelt National Park using AVIRIS. In Proceedings of the 9th JPL Airborne Earth Science Workshop, Pasadena, CA, USA, 23–25 February 2000. [Google Scholar]
- Root, R.R.; Anderson, G.L.; Hager, S.N.; Brown, K.E.; Dudek, K.B.; Kokaly, R.F. Application of advanced remote sensing and modeling techniques for the detection and management of leafy spurge: Challenges and opportunities. In Proceedings of the 57th Annual Meeting of the Society for Range Management, Salt Lake City, UT, USA, 24–30 January 2004; Society for Range Management: Wichita, KS, USA, 2004. [Google Scholar]
- Dudek, K.B.; Root, R.R.; Kokaly, R.F.; Anderson, G.L. Increased spatial and temporal consistency of leafy spurge maps from multidate AVIRIS imagery: A hybrid linear spectral mixture analysis/mixture-tuned matched filtering approach. In Proceedings of the 13th JPL Airborne Earth Science Workshop, Pasadena, CA, USA, 31 March–2 April 2004. [Google Scholar]
- Williams, A.P.; Hunt, E. Estimation of leafy spurge cover from hyperspectral imagery using mixture tuned matched filtering. Remote. Sens. Environ. 2002, 82, 446–456. [Google Scholar] [CrossRef]
- Glenn, N.F.; Mundt, J.T.; Weber, K.T.; Prather, T.S.; Lass, L.W.; Pettingill, J. Hyperspectral data processing for repeat detection of small infestations of leafy spurge. Remote. Sens. Environ. 2005, 95, 3548–3564. [Google Scholar] [CrossRef]
- Noujdina, N.V.; Ustin, S.L. Mapping downy brome (Bromis tectorum) using multidate AVIRIS data. Weed Sci. 2008, 56, 173–179. [Google Scholar] [CrossRef]
- Mbaabu, P.R.; Ng, W.-T.; Schaffner, U.; Gichaba, M.; Olago, D.; Choge, S.; Oriaso, S.; Eckert, S. Spatial Evolution of Prosopis Invasion and its Effects on LULC and Livelihoods in Baringo, Kenya. Remote. Sens. 2019, 11, 1217. [Google Scholar] [CrossRef]
- Malavasi, M.; Barták, V.; Jucker, T.; Acosta, A.T.R.; Carranza, M.L.; Bazzichetto, M. Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems. Remote. Sens. 2019, 11, 275. [Google Scholar] [CrossRef]
- Martin, F.-M.; Müllerová, J.; Borgniet, L.; Dommanget, F.; Breton, V.; Evette, A. Using Single- and Multi-Date UAV and Satellite Imagery to Accurately Monitor Invasive Knotweed Species. Remote. Sens. 2018, 10, 1662. [Google Scholar] [CrossRef]
- Bransky, N.D.; Sankey, T.T.; Sankey, J.B. Northern Tamarisk Beetle Impact and Classification Maps, Grand Canyon National Park, Arizona; U.S. Geological Survey data release; U.S. Geological Survey: Reston, VA, USA, 2021. [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAM Classification Angles | ||||
|---|---|---|---|---|
| May | August | |||
| Class | Radians | Degrees | Radians | Degrees |
| Bare Ground | 0.204 | 11.698 | 0.213 | 12.175 |
| Healthy Tamarisk | 0.291 | 11.555 | 0.233 | 13.321 |
| Mixed Healthy Leaf and Canopy Dieback Tamarisk | 0.091 | 5.204 | 0.207 | 6.112 |
| Canopy Dieback | 0.954 | 3.104 | 0.050 | 2.865 |
| Defoliated Tamarisk | 0.069 | 3.940 | 0.100 | 5.730 |
| Observed Healthy Tamarisk | Observed Canopy Dieback | Observed Defoliated Tamarisk | Total | User’s Accuracy | |
|---|---|---|---|---|---|
| Classified Healthy Tamarisk | 50 | 13 | 1 | 64 | 78.1% |
| Classified Canopy Dieback | 7 | 32 | 0 | 39 | 82.1% |
| Classified Defoliated Tamarisk | 0 | 0 | 2 | 2 | 100.0% |
| Total | 57 | 45 | 3 | 105 | |
| Producer’s Accuracy | 87.7% | 71.1% | 66.7% | ||
| Overall Accuracy | 80.0% |
| Observed Healthy Tamarisk | Observed Defoliated Tamarisk | Total | User’s Accuracy | |
|---|---|---|---|---|
| Classified Healthy Tamarisk | 13 | 7 | 20 | 65.0% |
| Classified Defoliated Tamarisk | 4 | 41 | 45 | 91.1% |
| Total | 17 | 48 | 65 | |
| Producer’s Accuracy | 76.5% | 85.4% | ||
| Overall Accuracy | 83.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bransky, N.; Sankey, T.; B. Sankey, J.; Johnson, M.; Jamison, L. Monitoring Tamarix Changes Using WorldView-2 Satellite Imagery in Grand Canyon National Park, Arizona. Remote Sens. 2021, 13, 958. https://doi.org/10.3390/rs13050958
Bransky N, Sankey T, B. Sankey J, Johnson M, Jamison L. Monitoring Tamarix Changes Using WorldView-2 Satellite Imagery in Grand Canyon National Park, Arizona. Remote Sensing. 2021; 13(5):958. https://doi.org/10.3390/rs13050958
Chicago/Turabian StyleBransky, Nathaniel, Temuulen Sankey, Joel B. Sankey, Matthew Johnson, and Levi Jamison. 2021. "Monitoring Tamarix Changes Using WorldView-2 Satellite Imagery in Grand Canyon National Park, Arizona" Remote Sensing 13, no. 5: 958. https://doi.org/10.3390/rs13050958
APA StyleBransky, N., Sankey, T., B. Sankey, J., Johnson, M., & Jamison, L. (2021). Monitoring Tamarix Changes Using WorldView-2 Satellite Imagery in Grand Canyon National Park, Arizona. Remote Sensing, 13(5), 958. https://doi.org/10.3390/rs13050958