
Coverage and Rainfall Response of Biological Soil Crusts Using Multi-Temporal Sentinel-2 Data in a Central European Temperate Dry Acid Grassland



Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensor | Timeframes/Temporal Resolution | Focus Areas | References | |||
|---|---|---|---|---|---|---|
| Platform | Type | Name | Spatial Resolution | |||
| Spaceborne | MS | Kompsat-2 | 1 m | 2 images | Niger | [50] |
| MS | QuickBird, WorldView-2 | 2.5 m | 2 images | Negev | [51] | |
| MS | SPOT | 20 m | 28 images during 2 years | Negev | [45] | |
| MS | Landsat 4–8 | 30–75 m | single images, 31 annual composites | Negev, China, Namibia, South Africa | [22,26,27,28,36,48,52] | |
| MS | Sentinel-2 MSI | 10–20 m | single image, 20 images during 2 years, full time series during 2016 | China, Negev, Idaho | [28,30,53] | |
| MS | Meteosat-9 SEVIRI | 3 km | Every 15 min during 2 months | Negev | [46] | |
| MS | ASTER | 90 m | “several” images * | Negev | [54] | |
| MS | MODIS | 250 m–1 km | Every 16 days during growing season, every 8 days from 2000 to 2014 | Patagonia, China | [55,56] | |
| SAR | SIR-C | 13–30 m | single image | Negev and Sinai | [57] | |
| SAR | ASAR | - * | 15 images during 1.5 years | Negev | [57] | |
| UAV | MS | Ricoh GR II | 0.5 cm | single image | Utah | [58] |
| MS | DJI X5s | 1–3 cm | single images | Hawaii | [59] | |
| MS | - * | - * | single images | China | [28] | |
| Airborne | HS | AMS | 5 m | single images | Australia | [25] |
| HS | CASI 1&2 | 1–1.5 m | single images | Spain, South Africa | [9,10,29,31] | |
| HS | DAIS | 8 m | single images | Negev | [26,36,54] | |
2. Materials and Methods
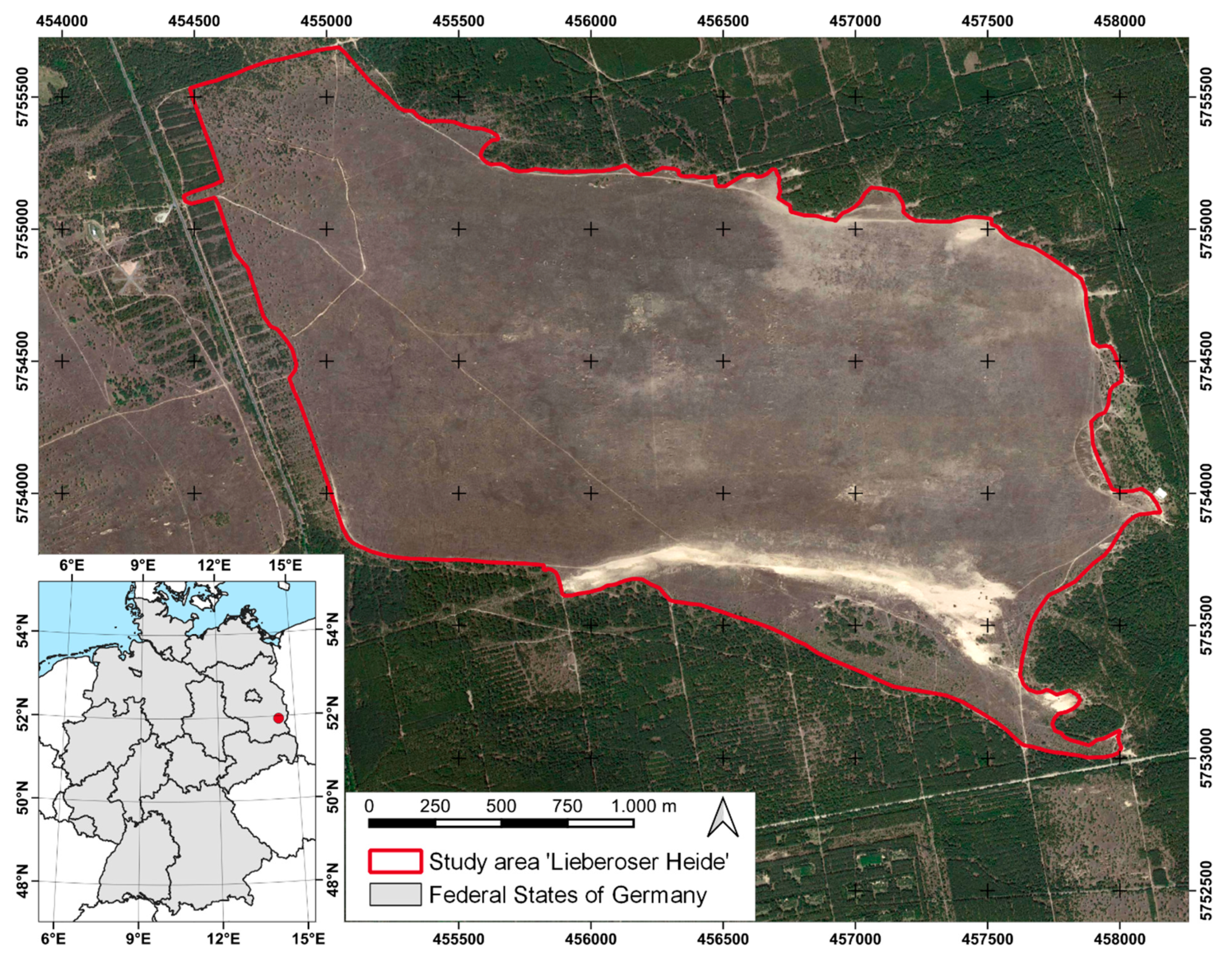
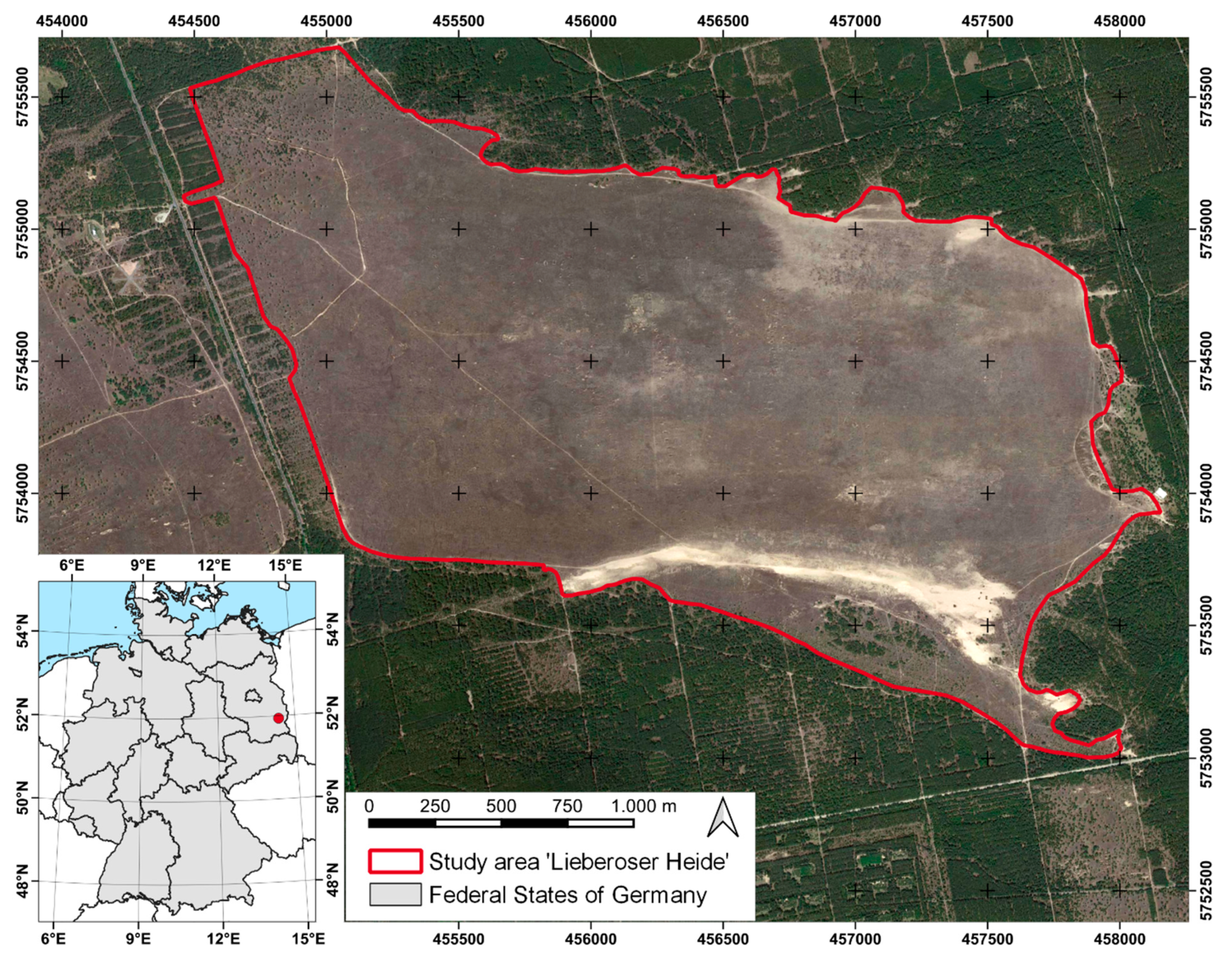
2.1. Study Area
2.2. Dataset Construction and Data Processing
2.2.1. In Situ Data Collection
2.2.2. Meteorological Data
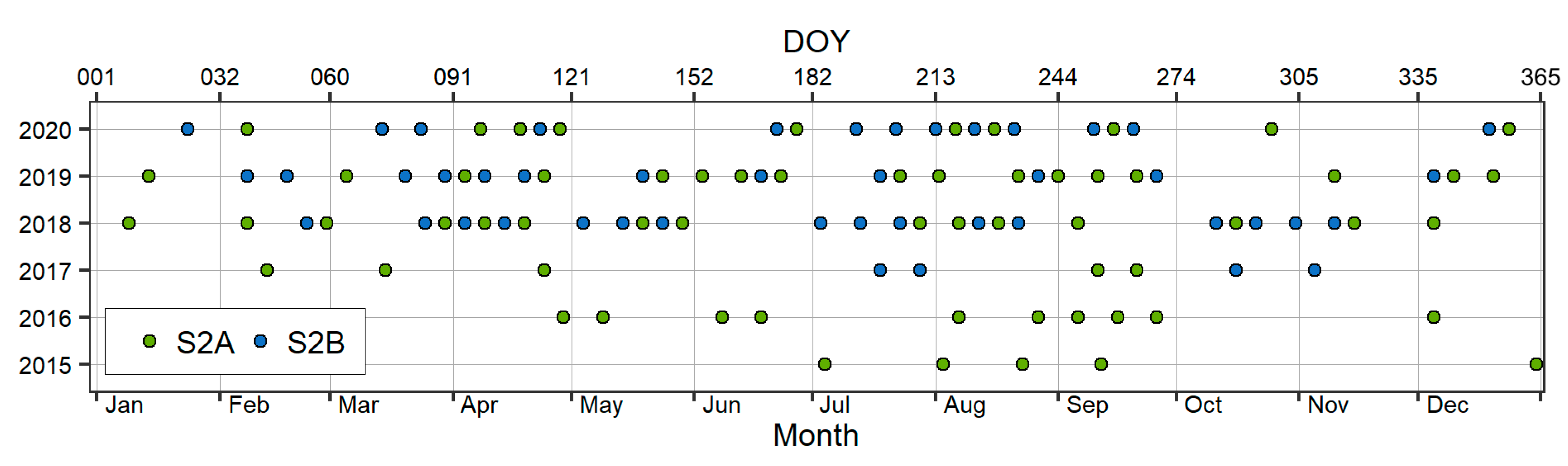
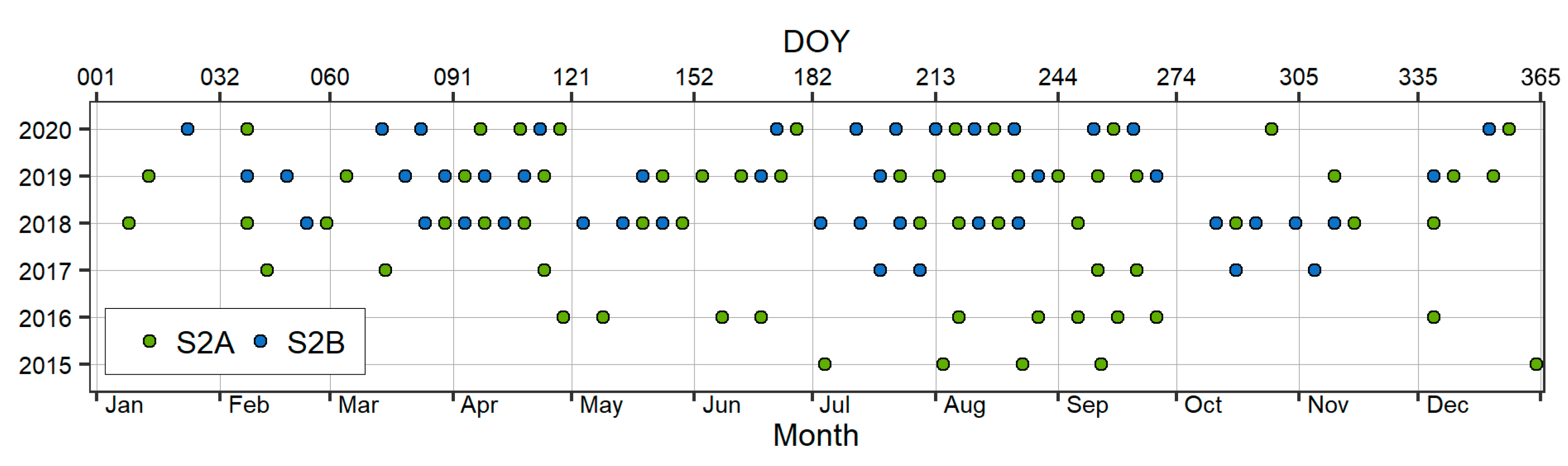
2.2.3. Sentinel-2 Data Preprocessing and Derivation of Multispectral Indices
2.2.4. Topographic Attributes
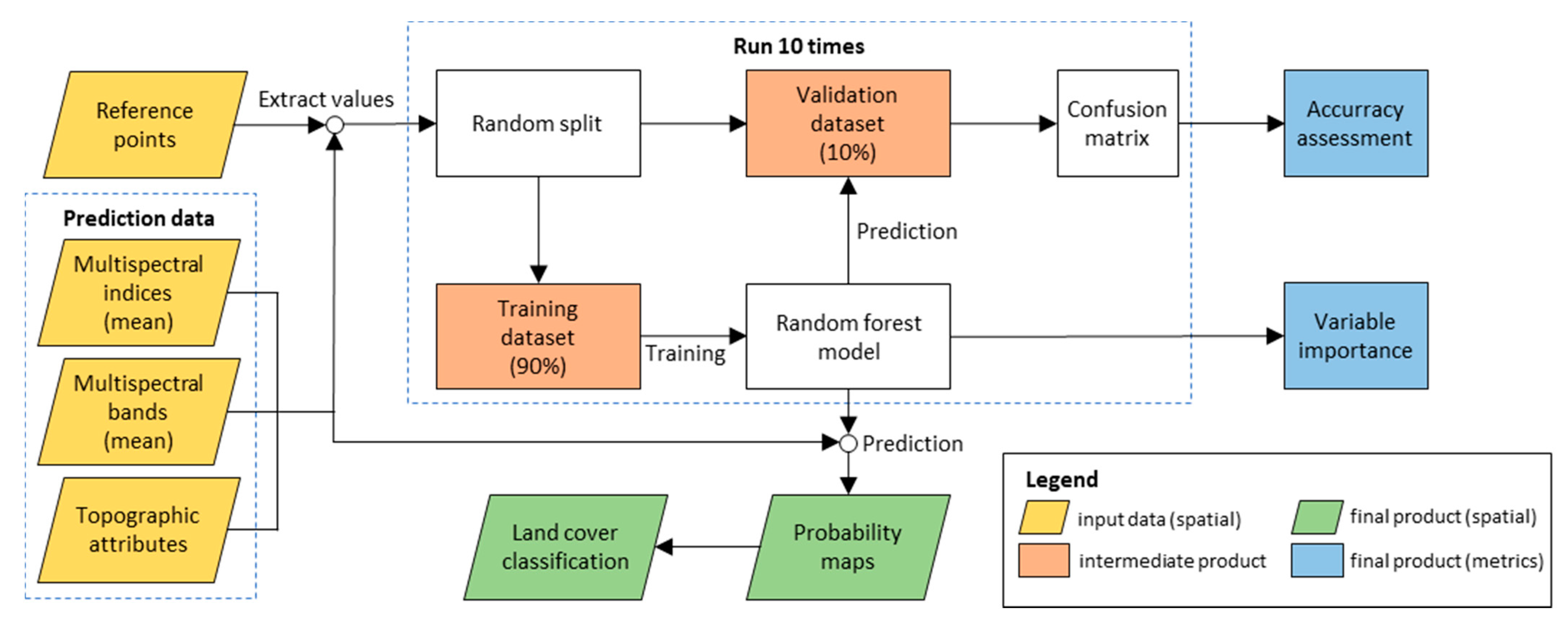
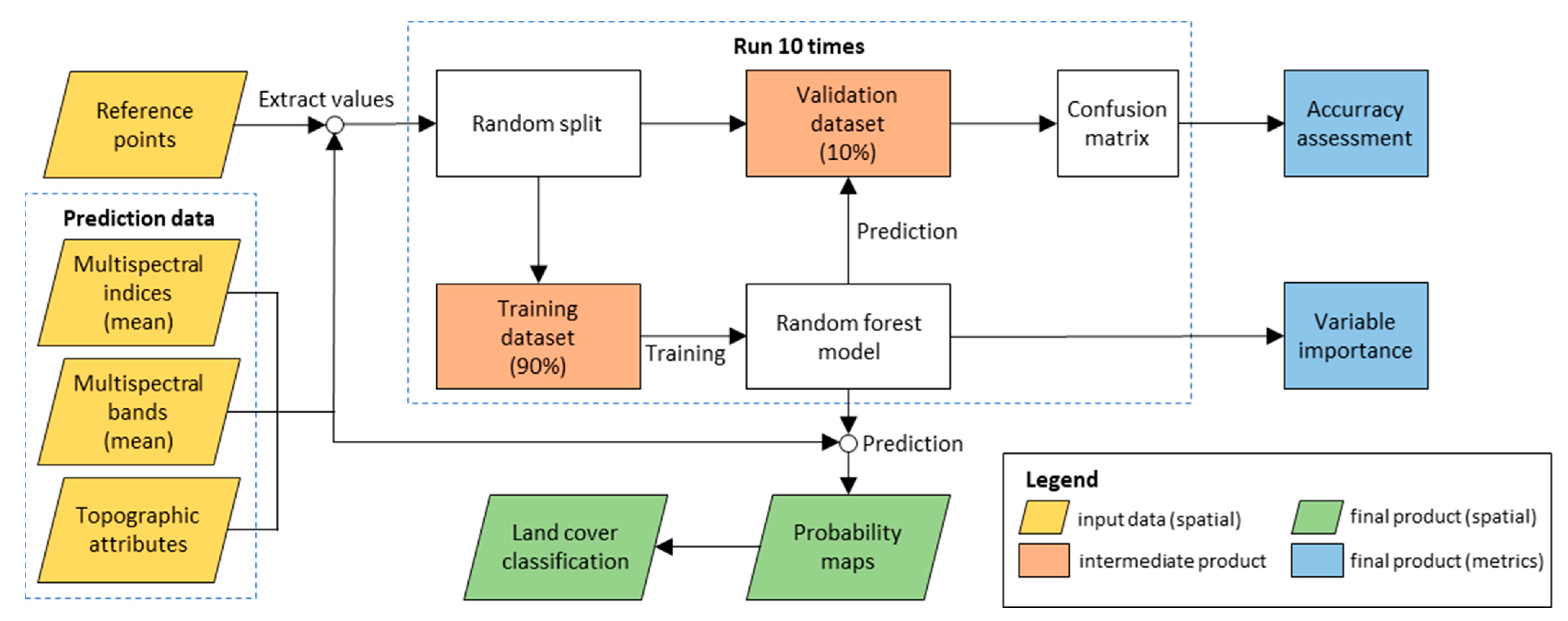
2.3. Biocrust Mapping and Response to Rainfall
2.3.1. Estimating Biological Soil Crust Coverage
2.3.2. Analyzing the Seasonal and Rainfall Event-Based Activity of Biological Soil Crusts
3. Results
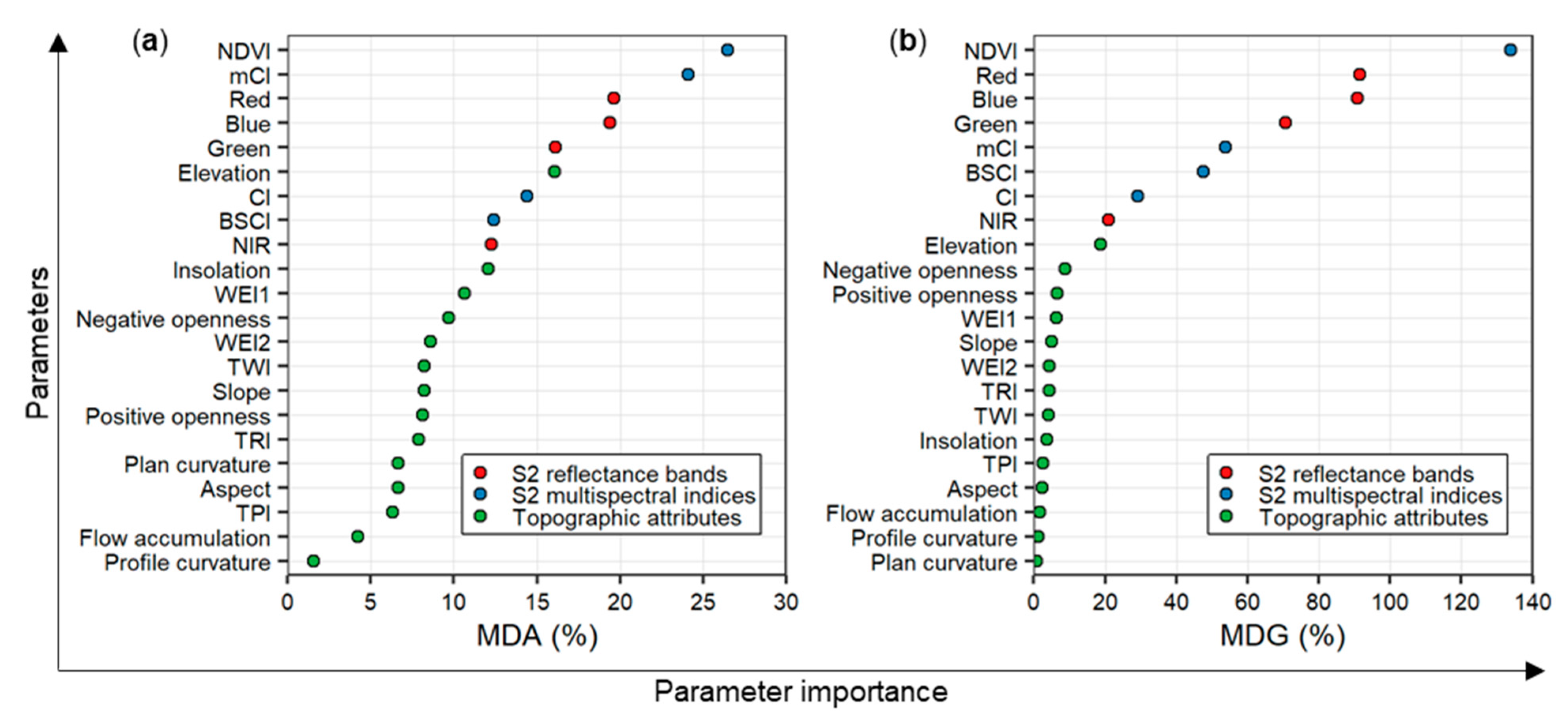
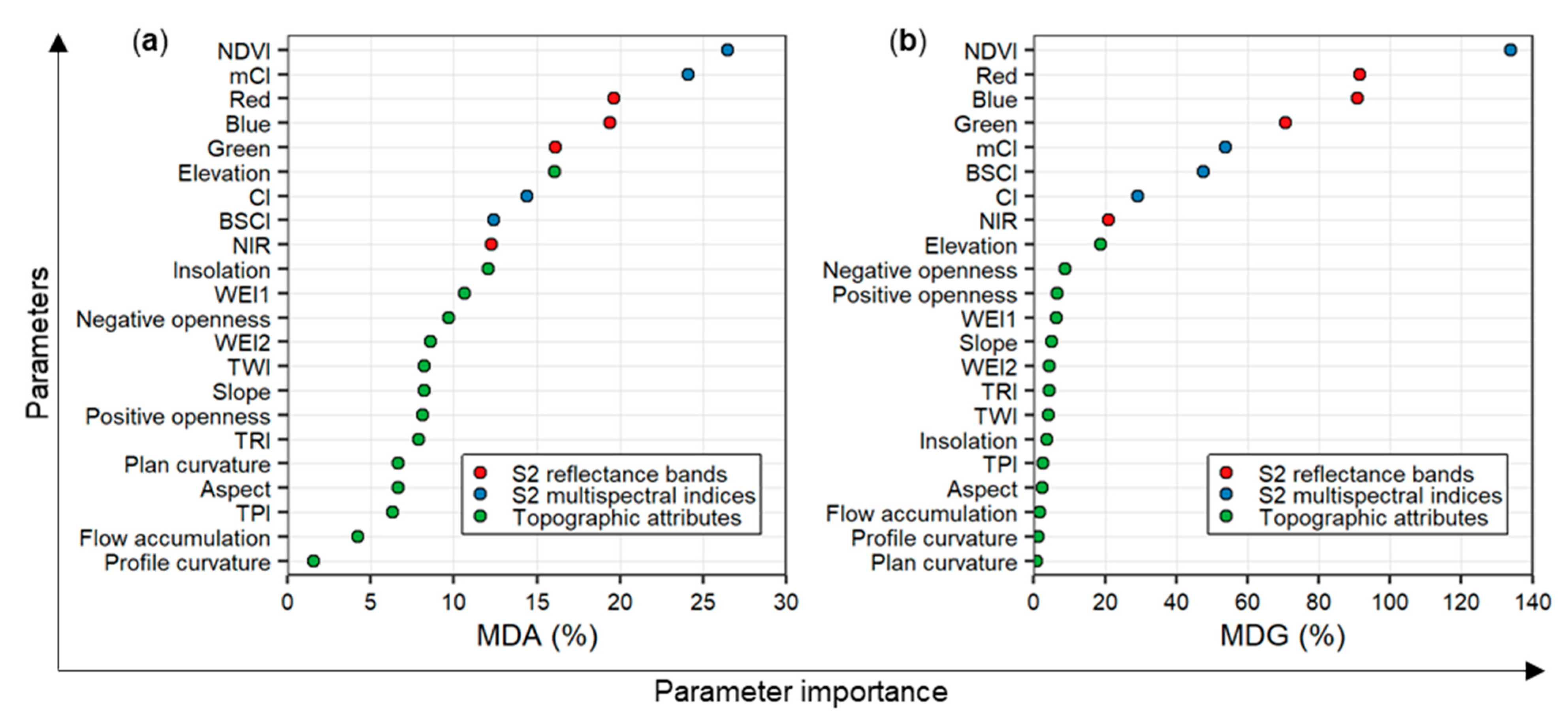
3.1. Performance of the Random Forest Classification Model
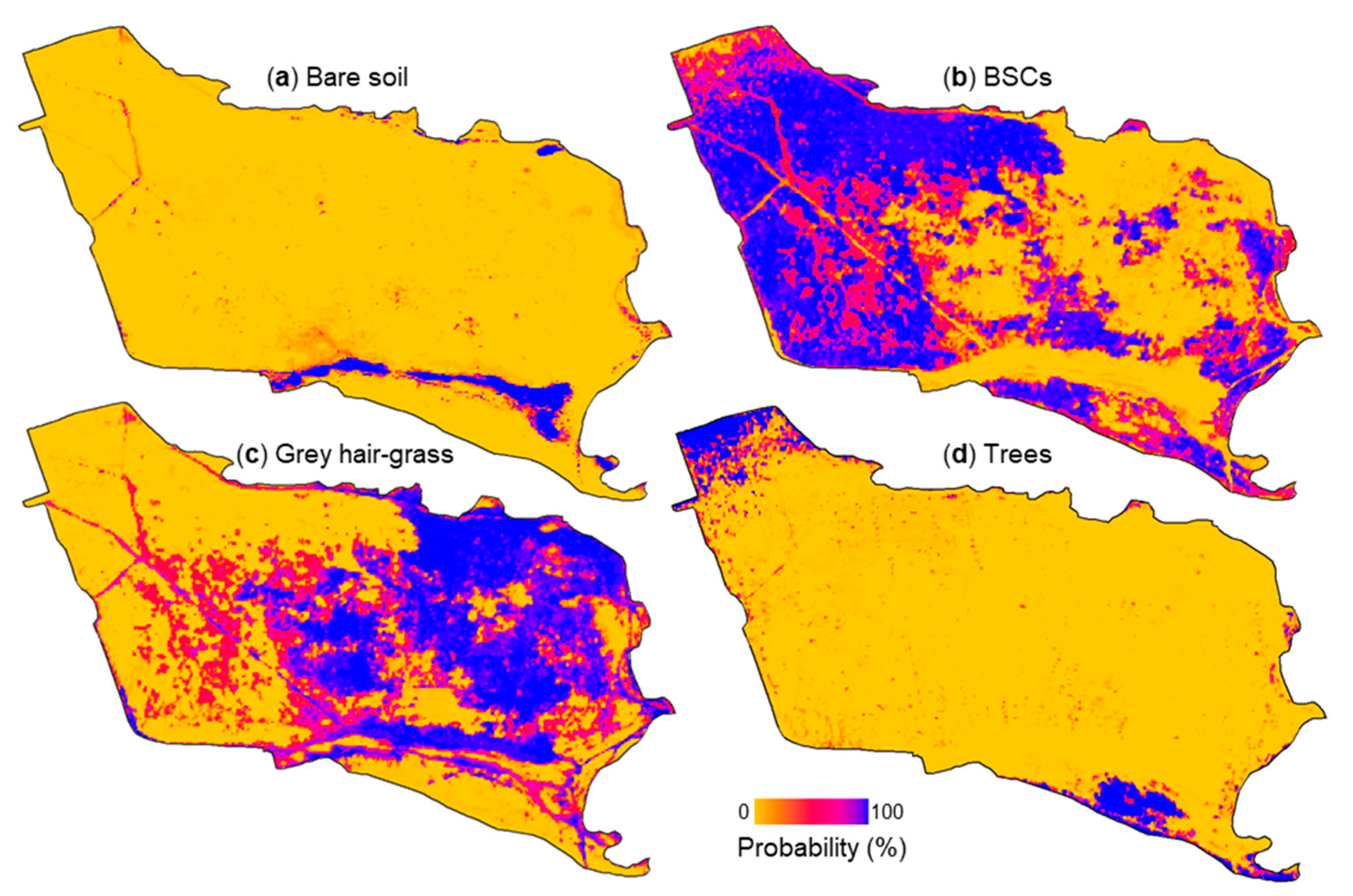
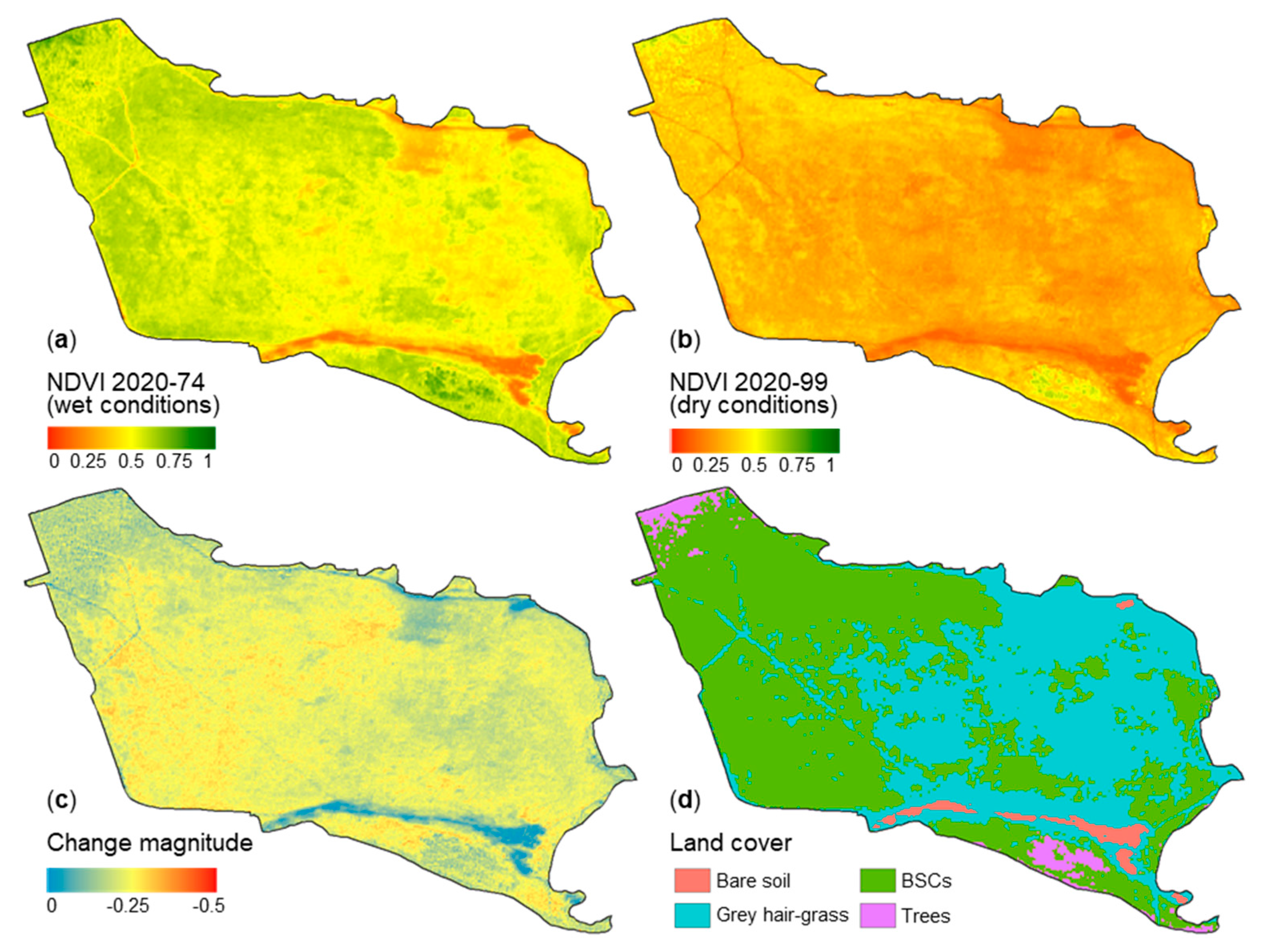
3.2. Estimated Biological Soil Crust Coverage
3.3. Dry and Wet Cycles of Biological Soil Crusts
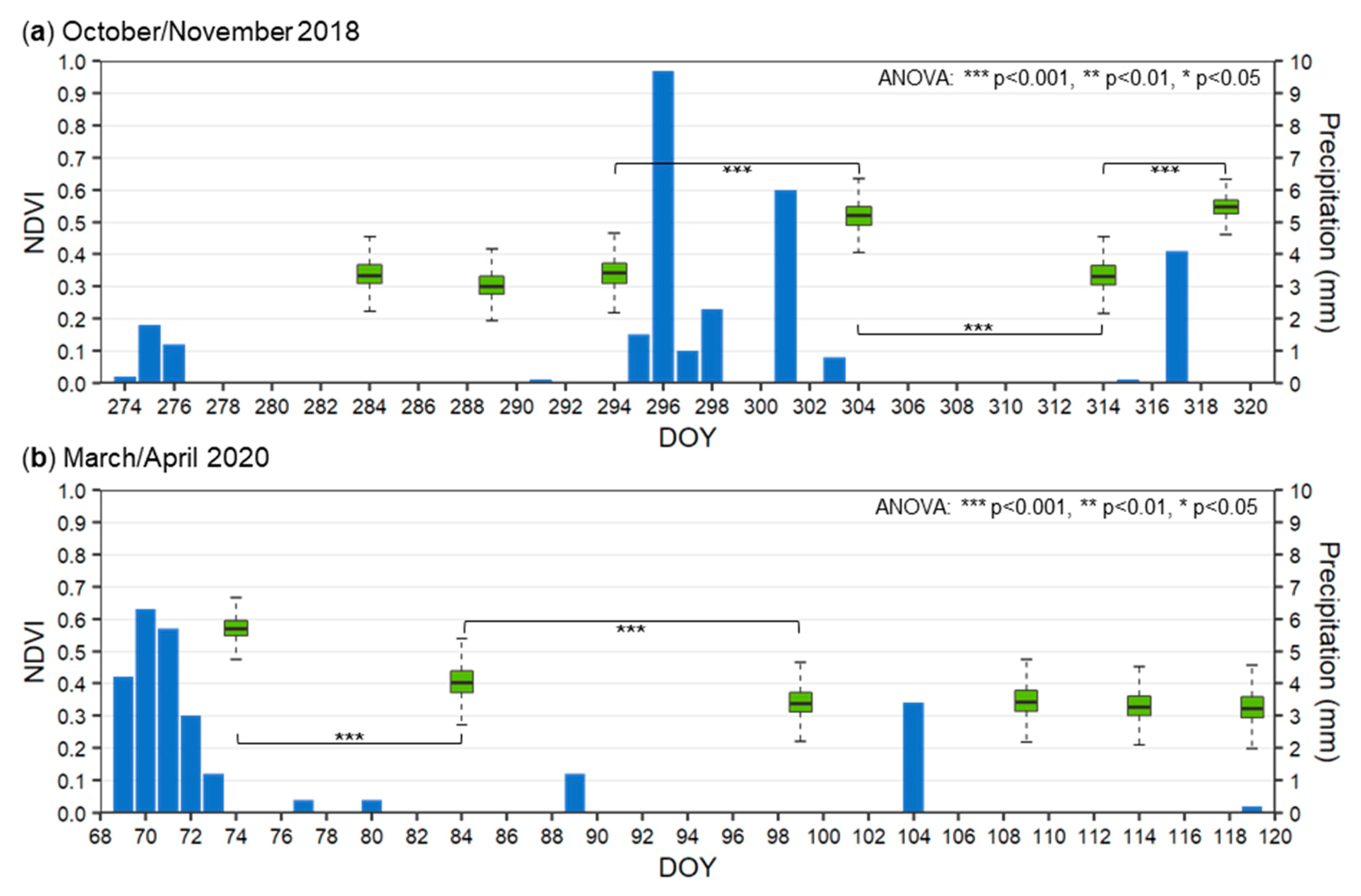
3.3.1. Activity of Biological Soil Crusts in Response to Rainfall Events
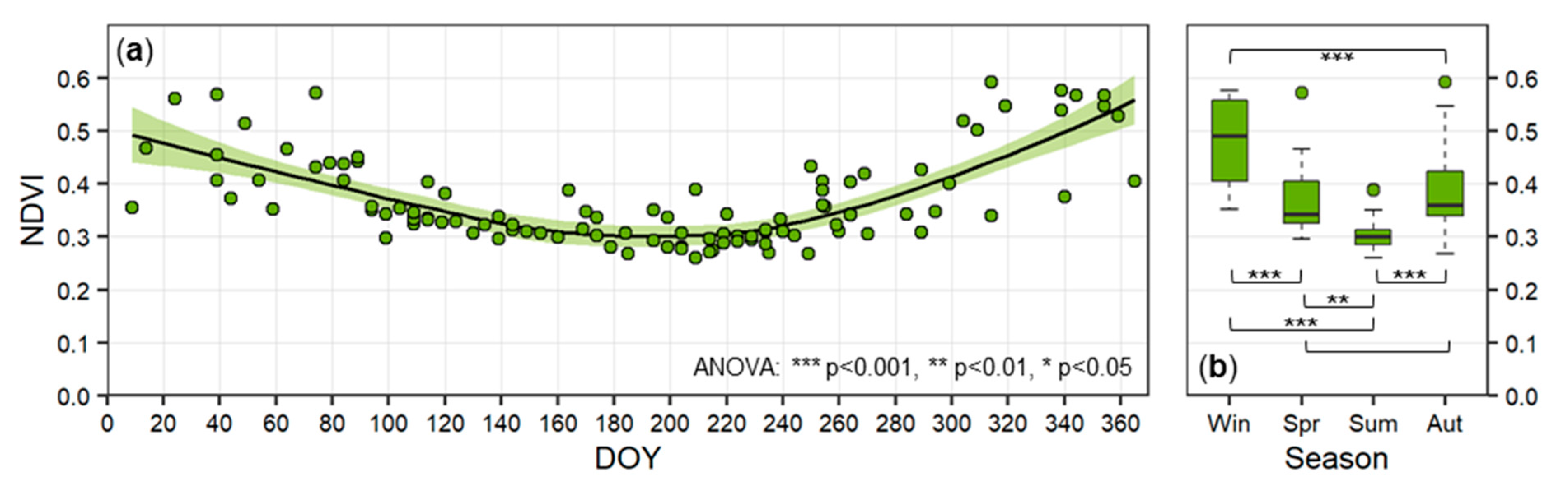
3.3.2. Seasonal Trends in the Activity of Biological Soil Crusts
4. Discussion
4.1. Land Cover Classification and Biological Soil Crust Coverage Estimation
4.2. Dry and Wet Cycles of Biological Soil Crusts
4.2.1. Activity of the Biological Soil Crusts in Response to Rainfall Events
4.2.2. Seasonal Trends in the Activity of Biological Soil Crusts
4.3. Future Research Potential to Enhance the Remote Sensing-Based Monitoring of Biological Soil Crusts
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belnap, J. Structure and functioning of biological soil crusts: A synthesis. In Biological Soil Crusts: Structure, Function, and Management, 1st ed.; Belnap, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 471–479. ISBN 3540437576. [Google Scholar]
- Burgheimer, J.; Wilske, B.; Maseyk, K.; Karnieli, A.; Zaady, E.; Yakir, D.; Kesselmeier, J. Relationships between Normalized Difference Vegetation Index (NDVI) and carbon fluxes of biologic soil crusts assessed by ground measurements. J. Arid. Environ. 2006, 64, 651–669. [Google Scholar] [CrossRef]
- Rodriguez-Caballero, E.; Belnap, J.; Büdel, B.; Crutzen, P.J.; Andreae, M.O.; Pöschl, U.; Weber, B. Dryland photoautotrophic soil surface communities endangered by global change. Nat. Geosci. 2018, 11, 185–189. [Google Scholar] [CrossRef]
- Colesie, C.; Felde, V.J.M.N.L.; Büdel, B. Composition and macrostructure of biological soil crusts. In Biological Soil Crusts: An Organizing Principle in Drylands; Weber, B., Büdel, B., Belnap, J., Eds.; Springer: Cham, Germany, 2016; pp. 159–172. ISBN 978-3-319-30212-6. [Google Scholar]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Belnap, J.; Walker, B.J.; Munson, S.M.; Gill, R.A. Controls on sediment production in two U.S. deserts. Aeolian Res. 2014, 14, 15–24. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, E.; Castro, A.J.; Chamizo, S.; Quintas-Soriano, C.; Garcia-Llorente, M.; Cantón, Y.; Weber, B. Ecosystem services provided by biocrusts: From ecosystem functions to social values. J. Arid. Environ. 2018, 159, 45–53. [Google Scholar] [CrossRef]
- Reed, S.C.; Delgado-Baquerizo, M.; Ferrenberg, S. Biocrust science and global change: Editorial. New Phytol. 2019, 223, 1047–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Caballero, E.; Paul, M.; Tamm, A.; Caesar, J.; Büdel, B.; Escribano, P.; Hill, J.; Weber, B. Biomass assessment of microbial surface communities by means of hyperspectral remote sensing data. Sci. Total Environ. 2017, 586, 1287–1297. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, E.; Escribano, P.; Cantón, Y. Advanced image processing methods as a tool to map and quantify different types of biological soil crust. ISPRS J. Photogramm. Remote Sens. 2014, 90, 59–67. [Google Scholar] [CrossRef]
- Büdel, B. Biological soil crusts in european temperate and mediterranean regions. In Biological Soil Crusts: Structure, Function, and Management, 1st ed.; Belnap, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 75–86. ISBN 3540437576. [Google Scholar]
- Jentsch, A.; Beyschlag, W. Vegetation ecology of dry acidic grasslands in the lowland area of Central europe. Flora-Morphol. Distrib. Funct. Ecol. Plants 2003, 198, 3–25. [Google Scholar] [CrossRef]
- Büdel, B.; Colesie, C.; Green, T.G.A.; Grube, M.; Lázaro Suau, R.; Loewen-Schneider, K.; Maier, S.; Peer, T.; Pintado, A.; Raggio, J.; et al. Improved appreciation of the functioning and importance of biological soil crusts in Europe: The Soil Crust International Project (SCIN). Biodivers. Conserv. 2014, 23, 1639–1658. [Google Scholar] [CrossRef] [Green Version]
- Spröte, R.; Fischer, T.; Veste, M.; Raab, T.; Wiehe, W.; Lange, P.; Bens, O.; Hüttl, R.F. Biological topsoil crusts at early successional stages on Quaternary substrates dumped by mining in Brandenburg, NE Germany. Geomorphologie 2010, 16, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Gypser, S.; Veste, M.; Fischer, T.; Lange, P. Formation of soil lichen crusts at reclaimed post-mining sites, Lower Lusatia, North-east Germany. Graph. Scr. 2015, 27, 3–14. [Google Scholar]
- Ellenberg, H.; Leuschner, C.; Dierschke, H. Vegetation Mitteleuropas mit den Alpen in Ökologischer, Dynamischer und Historischer Sicht; E. Ulmer: Stuttgart, Germany, 2010; ISBN 978-3-8001-2824-2. [Google Scholar]
- Fischer, T.; Veste, M.; Wiehe, W.; Lange, P. Water repellency and pore clogging at early successional stages of microbiotic crusts on inland dunes, Brandenburg, NE Germany. CATENA 2010, 80, 47–52. [Google Scholar] [CrossRef]
- Pluis, J.L.A. Algal crust formation in the inland dune area, Laarder Wasmeer, the Netherlands. Vegetatio 1994, 113, 41–51. [Google Scholar] [CrossRef]
- Bültmann, H.; Daniels, F.J.A. Biodiversity of terricolous lichen vegetation. Ber. Reinh. Tüxen Ges. 2000, 12, 393–397. [Google Scholar]
- Dümig, A.; Veste, M.; Hagedorn, F.; Fischer, T.; Lange, P.; Spröte, R.; Kögel-Knabner, I. Organic matter from biological soil crusts induces the initial formation of sandy temperate soils. CATENA 2014, 122, 196–208. [Google Scholar] [CrossRef]
- Fischer, T.; Yair, A.; Veste, M.; Geppert, H. Hydraulic properties of biological soil crusts on sand dunes studied by 13C-CP/MAS-NMR: A comparison between an arid and a temperate site. CATENA 2013, 110, 155–160. [Google Scholar] [CrossRef]
- Wessels, D.C.J.; van Vuuren, D.R.J. Landsat imagery-its possible use in mapping the distribution of major lichen communities in the Namib Desert, South West Africa. Madoqa 1986, 14, 369–373. [Google Scholar]
- Smith, W.K.; Dannenberg, M.P.; Yan, D.; Herrmann, S.; Barnes, M.L.; Barron-Gafford, G.A.; Biederman, J.A.; Ferrenberg, S.; Fox, A.M.; Hudson, A.; et al. Remote sensing of dryland ecosystem structure and function: Progress, challenges, and opportunities. Remote Sens. Environ. 2019, 233, 111401. [Google Scholar] [CrossRef]
- Rozenstein, O.; Adamowski, J. A review of progress in identifying and characterizing biocrusts using proximal and remote sensing. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 245–255. [Google Scholar] [CrossRef]
- Lewis, M.; Jooste, V.; De Gasparis, A. Discrimination of arid vegetation with airborne multispectral scanner hyperspectral imagery. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1471–1479. [Google Scholar] [CrossRef]
- Karnieli, A. Development and implementation of spectral crust index over dune sands. Int. J. Remote Sens. 1997, 18, 1207–1220. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, M.Y.; Wang, L.; Shimazaki, H.; Tamura, M. A new index for mapping lichen-dominated biological soil crusts in desert areas. Remote Sens. Environ. 2005, 96, 165–175. [Google Scholar] [CrossRef]
- Chen, X.; Wang, T.; Liu, S.; Peng, F.; Tsunekawa, A.; Kang, W.; Guo, Z.; Feng, K. A new application of random forest algorithm to estimate coverage of moss-dominated biological soil crusts in semi-arid mu us sandy land, China. Remote Sens. 2019, 11, 1286. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Caballero, E.; Escribano, P.; Olehowski, C.; Chamizo, S.; Hill, J.; Cantón, Y.; Weber, B. Transferability of multi- and hyperspectral optical biocrust indices. ISPRS J. Photogramm. Remote Sens. 2017, 126, 94–107. [Google Scholar] [CrossRef]
- Panigada, C.; Tagliabue, G.; Zaady, E.; Rozenstein, O.; Garzonio, R.; Di Mauro, B.; De Amicis, M.; Colombo, R.; Cogliati, S.; Miglietta, F.; et al. A new approach for biocrust and vegetation monitoring in drylands using multi-temporal Sentinel-2 images. Prog. Phys. Geogr. Earth Environ. 2019, 43, 496–520. [Google Scholar] [CrossRef]
- Weber, B.; Olehowski, C.; Knerr, T.; Hill, J.; Deutschewitz, K.; Wessels, D.C.J.; Eitel, B.; Büdel, B. A new approach for mapping of biological soil crusts in semidesert areas with hyperspectral imagery. Remote Sens. Environ. 2008, 112, 2187–2201. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, E.; Román, J.R.; Chamizo, S.; Roncero Ramos, B.; Cantón, Y. Biocrust landscape-scale spatial distribution is strongly controlled by terrain attributes: Topographic thresholds for colonization in a semiarid badland system. Earth Surf. Process. Landf. 2019, 44, 2771–2779. [Google Scholar] [CrossRef]
- Weber, B.; Hill, J. Remote sensing of biological soil crusts at different scales. In Biological Soil Crusts: An Organizing Principle in Drylands; Weber, B., Büdel, B., Belnap, J., Eds.; Springer: Cham, Switzerland, 2016; pp. 215–236. ISBN 978-3-319-30212-6. [Google Scholar]
- O’Neill, A.L. Reflectance spectra of microphytic soil crusts in semi-arid Australia. Int. J. Remote Sens. 1994, 15, 675–681. [Google Scholar] [CrossRef]
- Karnieli, A.; Sarafis, V. Reflectance spectrophotometry of cyanobacteria within soil crusts—A diagnostic tool. Int. J. Remote Sens. 1996, 17, 1609–1615. [Google Scholar] [CrossRef]
- Karnieli, A.; Tsoar, H. Spectral reflectance of biogenic crust developed on desert dune sand along the Israel-Egypt border. Int. J. Remote Sens. 1995, 16, 369–374. [Google Scholar] [CrossRef]
- Karnieli, A.; Kidron, G.J.; Glaesser, C.; Ben-Dor, E. Spectral characteristics of cyanobacteria soil crust in semiarid environments. Remote Sens. Environ. 1999, 69, 67–75. [Google Scholar] [CrossRef]
- Pinker, R.T.; Karnieli, A. Characteristic spectral reflectance of a semi-arid environment. Int. J. Remote Sens. 1995, 16, 1341–1363. [Google Scholar] [CrossRef]
- Chen, X.; Wang, T.; Liu, S.; Peng, F.; Kang, W.; Guo, Z.; Feng, K.; Liu, J.; Tsunekawa, A. Spectral response assessment of moss-dominated biological soil Crust coverage under dry and wet conditions. Remote Sens. 2020, 12, 1158. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Caballero, E.; Knerr, T.; Weber, B. Importance of biocrusts in dryland monitoring using spectral indices. Remote Sens. Environ. 2015, 170, 32–39. [Google Scholar] [CrossRef]
- Chamizo, S.; Stevens, A.; Cantón, Y.; Miralles, I.; Domingo, F.; van Wesemael, B. Discriminating soil crust type, development stage and degree of disturbance in semiarid environments from their spectral characteristics. Eur. J. Soil Sci. 2012, 63, 42–53. [Google Scholar] [CrossRef]
- Blanco-Sacristán, J.; Panigada, C.; Tagliabue, G.; Gentili, R.; Colombo, R.; Ladrón de Guevara, M.; Maestre, F.T.; Rossini, M. Spectral diversity successfully estimates the α-diversity of biocrust-forming lichens. Remote Sens. 2019, 11, 2942. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, L.; Jung, P.; Obermeier, W.; Büdel, B.; Bendix, J. Estimating net photosynthesis of biological soil crusts in the atacama using hyperspectral remote sensing. Remote Sens. 2018, 10, 891. [Google Scholar] [CrossRef] [Green Version]
- Román, J.R.; Rodríguez-Caballero, E.; Rodríguez-Lozano, B.; Roncero-Ramos, B.; Chamizo, S.; Águila-Carricondo, P.; Cantón, Y. Spectral response analysis: An indirect and non-destructive methodology for the chlorophyll quantification of biocrusts. Remote Sens. 2019, 11, 1350. [Google Scholar] [CrossRef] [Green Version]
- Burgheimer, J.; Wilske, B.; Maseyk, K.; Karnieli, A.; Zaady, E.; Yakir, D.; Kesselmeier, J. Ground and space spectral measurements for assessing the semi-arid ecosystem phenology related to CO2 fluxes of biological soil crusts. Remote Sens. Environ. 2006, 101, 1–12. [Google Scholar] [CrossRef]
- Rozenstein, O.; Agam, N.; Serio, C.; Masiello, G.; Venafra, S.; Achal, S.; Puckrin, E.; Karnieli, A. Diurnal emissivity dynamics in bare versus biocrusted sand dunes. Sci. Total Environ. 2015, 506–507, 422–429. [Google Scholar] [CrossRef]
- Potter, C.; Weigand, J. Imaging analysis of biological soil crusts to understand surface heating properties in the mojave desert of California. CATENA 2018, 170, 1–9. [Google Scholar] [CrossRef]
- Noy, K.; Ohana-Levi, N.; Panov, N.; Silver, M.; Karnieli, A. A long-term spatiotemporal analysis of biocrusts across a diverse arid environment: The case of the Israeli-Egyptian sandfield. Sci. Total Environ. 2021, 774, 145154. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A. Monitoring Vegetation Systems in the Great Plains with ERTS; NASA Special Publication: Washington, DC, USA, 10–14 December 1974.
- Beaugendre, N.; Malam Issa, O.; Choné, A.; Cerdan, O.; Desprats, J.-F.; Rajot, J.L.; Sannier, C.; Valentin, C. Developing a predictive environment-based model for mapping biological soil crust patterns at the local scale in the Sahel. CATENA 2017, 158, 250–265. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Panov, N.; Shachak, M.; Zaady, E.; Karnieli, A. structural changes of desertified and managed shrubland landscapes in response to drought: Spectral, spatial and temporal analyses. Remote Sens. 2014, 6, 8134–8164. [Google Scholar] [CrossRef] [Green Version]
- Green, G.M. Use of SIR-A and Landsat MSS data in mapping shrub and intershrub vegetation at Koonamore, South Australia. Photogramm. Eng. Remote Sens. 1986, 52, 659–670. [Google Scholar]
- Enterkine, J. Remote Sensing Time-Series Analysis, Machine Learning, and K-Means Clustering Improves Dryland Vegetation and Biological Soil Crust Classification. Master’s Thesis, Boise State University, Boise, ID, USA, 2019. [Google Scholar]
- Rozenstein, O.; Karnieli, A. Identification and characterization of biological soil crusts in a sand dune desert environment across Israel–Egypt border using LWIR emittance spectroscopy. J. Arid. Environ. 2015, 112, 75–86. [Google Scholar] [CrossRef]
- Gaitán, J.J.; Bran, D.; Oliva, G.; Ciari, G.; Nakamatsu, V.; Salomone, J.; Ferrante, D.; Buono, G.; Massara, V.; Humano, G.; et al. Evaluating the performance of multiple remote sensing indices to predict the spatial variability of ecosystem structure and functioning in Patagonian steppes. Ecol. Indic. 2013, 34, 181–191. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Qin, S.; Wu, B.; Ding, G.; Wu, X.; Gao, Y.; Zhu, Y. Carrying capacity for vegetation across northern China drylands. Sci. Total Environ. 2020, 710, 136391. [Google Scholar] [CrossRef]
- Rozenstein, O.; Siegal, Z.; Blumberg, D.G.; Adamowski, J. Investigating the backscatter contrast anomaly in synthetic aperture radar (SAR) imagery of the dunes along the Israel–Egypt border. Int. J. Appl. Earth Obs. Geoinf. 2016, 46, 13–21. [Google Scholar] [CrossRef]
- Havrilla, C.A.; Villarreal, M.L.; DiBiase, J.L.; Duniway, M.C.; Barger, N.N. Ultra-high-resolution mapping of biocrusts with Unmanned Aerial Systems. Remote Sens. Ecol. 2020, 6, 441–456. [Google Scholar] [CrossRef]
- Collier, E.A. Mapping Biological Soil Crust Cover in the Kawaihae Watershed. Master’s Thesis, University of Hawai’I, Hilo, HI, USA, 2019. [Google Scholar]
- Qin, Z.; Berliner, P.; Karnieli, A. Micrometeorological modeling to understand the thermal anomaly in the sand dunes across the Israel–Egypt border. J. Arid. Environ. 2002, 51, 281–318. [Google Scholar] [CrossRef] [Green Version]
- Band, L.E. Spatial aggregation of complex terrain. Geogr. Anal. 1989, 21, 279–293. [Google Scholar] [CrossRef]
- Fels, J.E. Modeling and Mapping Potential Vegetation Using Digital Terrain Data: Applications in the Ellicott Rock Wilderness of North Carolina, South Carolina and Georgia. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 1994. [Google Scholar]
- Burrough, P.A.; Wilson, J.P.; van Gaans, P.F.M.; Hansen, A.J. Fuzzy k-means classification of topo-climatic data as an aid to forest mapping in the Greater Yellowstone Area, USA. Landsc. Ecol. 2001, 16, 523–546. [Google Scholar] [CrossRef]
- Pfeffer, K.; Pebesma, E.J.; Burrough, P.A. Mapping alpine vegetation using vegetation observations and topographic attributes. Landsc. Ecol. 2003, 18, 759–776. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Meave, J.A.; Zermeño-Hernández, I.; Ibarra-Manríquez, G. Effects of slope aspect and topographic position on environmental variables, disturbance regime and tree community attributes in a seasonal tropical dry forest. J. Veg. Sci. 2016, 27, 1094–1103. [Google Scholar] [CrossRef]
- Alexander, R.W.; Millington, A.C. Vegetation Mapping: From Patch to Planet; Wiley: Chichester, UK, 2000; ISBN 978-0-471-96592-3. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Pal, M. Random forest classifier for remote sensing classification. Int. J. Remote Sens. 2005, 26, 217–222. [Google Scholar] [CrossRef]
- Mandanici, E.; Bitelli, G. Preliminary Comparison of Sentinel-2 and Landsat 8 Imagery for a Combined Use. Remote Sens. 2016, 8, 1014. [Google Scholar] [CrossRef] [Green Version]
- Gypser, S.; Herppich, W.B.; Fischer, T.; Lange, P.; Veste, M. Photosynthetic characteristics and their spatial variance on biological soil crusts covering initial soils of post-mining sites in Lower Lusatia, NE Germany. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 220, 103–116. [Google Scholar] [CrossRef]
- Kleefeld, A.; Gypser, S.; Herppich, W.B.; Bader, G.; Veste, M. Identification of spatial pattern of photosynthesis hotspots in moss- and lichen-dominated biological soil crusts by combining chlorophyll fluorescence imaging and multispectral BNDVI images. Pedobiologia 2018, 68, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.; Veste, M.; Eisele, A.; Bens, O.; Spyra, W.; Hüttl, R.F. Small scale spatial heterogeneity of Normalized Difference Vegetation Indices (NDVIs) and hot spots of photosynthesis in biological soil crusts. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 159–167. [Google Scholar] [CrossRef]
- Schumacher, H. Faszination Lieberoser Heide. In Faszination Lieberoser Heide: Landschaft Zwischen Wald, Wasser und Weite; Brandenburg, S.N., Ed.; Regia-Verl.: Cottbus, Germany, 2010; pp. 14–33. ISBN 978-3-86929-180-2. [Google Scholar]
- Brunk, I.; Anders, K.; Mähnert, P.; Mrzljak, J.; Nocker, U.; Saure, C.; Vorwald, J.; Borries, J.; Wiegleb, G. Der ehemalige truppenübungsplatz lieberose. In Handbuch Offenlandmanagement; Anders, K., Mrzljak, J., Wallschläger, D., Wiegleb, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 227–242. ISBN 978-3-642-62218-2. [Google Scholar]
- Hoppert, M.; Reimer, R.; Kemmling, A.; Schröder, A.; Günzl, B.; Heinken, T. Structure and reactivity of a biological soil crust from a xeric sandy soil in central europe. Geomicrobiol. J. 2004, 21, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Brankatschk, R.; Fischer, T.; Veste, M.; Zeyer, J. Succession of N cycling processes in biological soil crusts on a central European inland dune. FEMS Microbiol. Ecol. 2013, 83, 149–160. [Google Scholar] [CrossRef]
- Zaady, E.; Kuhn, U.; Wilske, B.; Sandoval-Soto, L.; Kesselmeier, J. Patterns of CO2 exchange in biological soil crusts of successional age. Soil Biol. Biochem. 2000, 32, 959–966. [Google Scholar] [CrossRef]
- R core team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P.; et al. Sentinel-2: ESA’s Optical High-Resolution Mission for GMES Operational Services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Gascon, F.; Bouzinac, C.; Thépaut, O.; Jung, M.; Francesconi, B.; Louis, J.; Lonjou, V.; Lafrance, B.; Massera, S.; Gaudel-Vacaresse, A.; et al. Copernicus sentinel-2A calibration and products validation status. Remote Sens. 2017, 9, 584. [Google Scholar] [CrossRef] [Green Version]
- Ranghetti, L.; Boschetti, M.; Nutini, F.; Busetto, L. “sen2r”: An R toolbox for automatically downloading and preprocessing Sentinel-2 satellite data. Comput. Geosci. 2020, 139, 104473. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, S.; Woodcock, C.E. Improvement and expansion of the Fmask algorithm: Cloud, cloud shadow, and snow detection for Landsats 4–7, 8, and Sentinel 2 images. Remote Sens. Environ. 2015, 159, 269–277. [Google Scholar] [CrossRef]
- Baetens, L.; Desjardins, C.; Hagolle, O. Validation of copernicus sentinel-2 cloud masks obtained from MAJA, Sen2Cor, and FMask processors using reference cloud masks generated with a supervised active learning procedure. Remote Sens. 2019, 11, 433. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Tang, G.; Zhang, T. A systematic classification research of topographic descriptive attribute in digital terrain analysis. In Proceedings of the XXIst ISPRS Congress Beijing 2008, Beijing, China, 3–11 July 2008. [Google Scholar]
- Beaudette, D.E.; Dahlgren, R.A.; O’Geen, A.T. Terrain-shape indices for modeling soil moisture dynamics. Soil Sci. Soc. Am. J. 2013, 77, 1696–1710. [Google Scholar] [CrossRef] [Green Version]
- Polidori, L.; El Hage, M. Digital elevation model quality assessment methods: A critical review. Remote Sens. 2020, 12, 3522. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef] [Green Version]
- Olaya, V. Chapter 6 basic land-surface parameters. In Geomorphometry-Concepts, Software, Applications; Hengl, T., Reuter, H.I., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 141–169. ISBN 9780123743459. [Google Scholar]
- Qin, C.-Z.; Zhu, A.-X.; Pei, T.; Li, B.-L.; Scholten, T.; Behrens, T.; Zhou, C.-H. An approach to computing topographic wetness index based on maximum downslope gradient. Precis. Agric. 2011, 12, 32–43. [Google Scholar] [CrossRef]
- Hofierka, J.; Súri, M. The solar radiation model for Open source GIS: Implementation and applications. In Proceedings of the Open Source GIS-GRASS Users Conference, Trento, Italy, 11–13 September 2002. [Google Scholar]
- Yokohama, R.; Shirasawa, M.; Pike, R.J. Visualizing topography by openness. A new application of image processing to digital elevation models. Photogramm. Eng. Remote Sens. 2002, 68, 257–265. [Google Scholar]
- Guisan, A.; Weiss, S.B.; Weiss, A.D. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999, 143, 107–122. [Google Scholar] [CrossRef]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. A terrain ruggedness index that quantifies topographic heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Moore, I.D.; Grayson, R.B.; Ladson, A.R. Digital terrain modelling: A review of hydrological, geomorphological, and biological applications. Hydrol. Process. 1991, 5, 3–30. [Google Scholar] [CrossRef]
- Böhner, J.; Antonić, O. Chapter 8 land-surface parameters specific to Topo-CLIMATOLOGY. In Geomorphometry-Concepts, Software, Applications; Hengl, T., Reuter, H.I., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 195–226. ISBN 9780123743459. [Google Scholar]
- Liaw, A.; Wiener, M.; Breiman, L.; Cutler, A. Package RandomForest: Breiman and Cutler’s Rendom Forests for Classification and Regression. Version 4.6-14. Available online: https://cran.r-project.org/web/packages/randomForest/randomForest.pdf (accessed on 9 December 2020).
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Maxwell, A.E.; Warner, T.A.; Fang, F. Implementation of machine-learning classification in remote sensing: An applied review. Int. J. Remote Sens. 2018, 39, 2784–2817. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Guo, X.; Yu, H. Variable selection using mean decrease accuracy and mean decrease gini based on random forest. In Proceedings of the 2016 7th IEEE International Conference on Software Engineering and Service Science (ICSESS), Beijing, China, 8 August 2016; pp. 219–224, ISBN 978-1-4673-9904-3. [Google Scholar]
- Congalton, R.G.; Green, K. Assessing the Accuracy of Remotely Sensed Data: Principles and Practices, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009; ISBN 9780367656676. [Google Scholar]
- McColl, R.W. Encyclopedia of World Geography; Checkmark Books/Facts on File: New York, NY, USA, 2005; ISBN 0816057869. [Google Scholar]
- DWD. Aus Extrem Wurde Normal: Sommer in Deutschland, der Schweiz und Österreich Immer Heißer. Available online: https://www.dwd.de/DE/presse/pressemitteilungen/DE/2020/20200702_dach_news.html (accessed on 20 May 2020).
- Imbery, F.; Friedrich, K.; Koppe, C.; Janssen, W.; Pfeifroth, U.; Daßler, J.; Bissolli, P. 2018 Wärmster Sommer im Norden und Osten Deutschlands. Available online: https://www.dwd.de/DE/leistungen/besondereereignisse/temperatur/20180906_waermstersommer_nordenosten2018.pdf?__blob=publicationFile&v=6 (accessed on 20 May 2021).
- Reyer, C.; Bachinger, J.; Bloch, R.; Hattermann, F.F.; Ibisch, P.L.; Kreft, S.; Lasch, P.; Lucht, W.; Nowicki, C.; Spathelf, P.; et al. Climate change adaptation and sustainable regional development: A case study for the Federal State of Brandenburg, Germany. Reg. Environ. Chang. 2012, 12, 523–542. [Google Scholar] [CrossRef] [Green Version]
- Ghassemian, H. A review of remote sensing image fusion methods. Inf. Fusion 2016, 32, 75–89. [Google Scholar] [CrossRef]
- Schönbrodt-Stitt, S.; Ahmadian, N.; Kurtenbach, M.; Conrad, C.; Romano, N.; Bogena, H.R.; Vereecken, H.; Nasta, P. Statistical exploration of Sentinel-1 data, terrain parameters, and in-situ data for estimating the near-surface soil moisture in a mediterranean agroecosystem. Front. Water 2021, 3, 75. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, G.; Zhu, L.; Wang, H. Biocrust wetting induced change in soil surface roughness as influenced by biocrust type, coverage and wetting patterns. Geoderma 2017, 306, 1–9. [Google Scholar] [CrossRef]

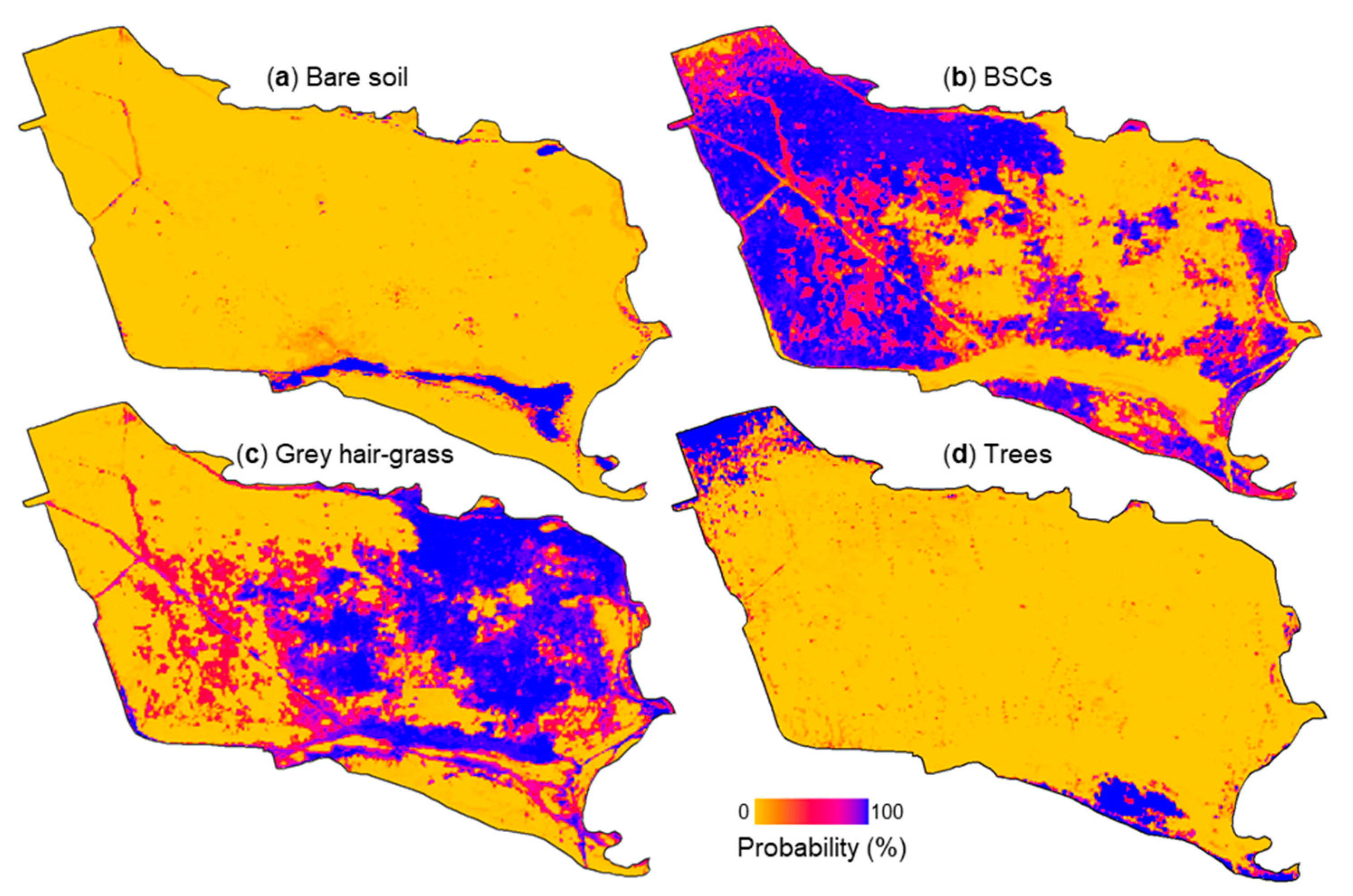

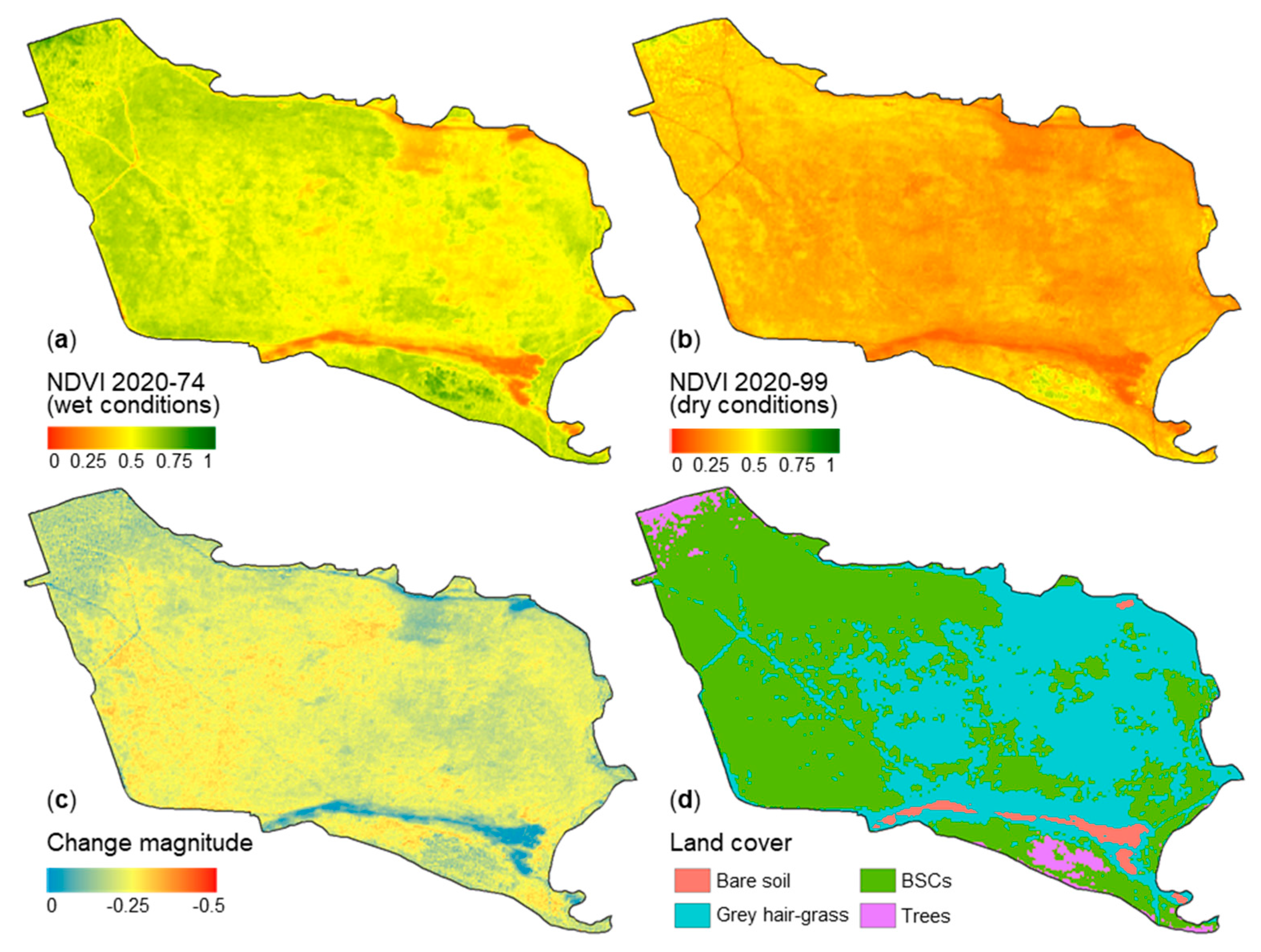
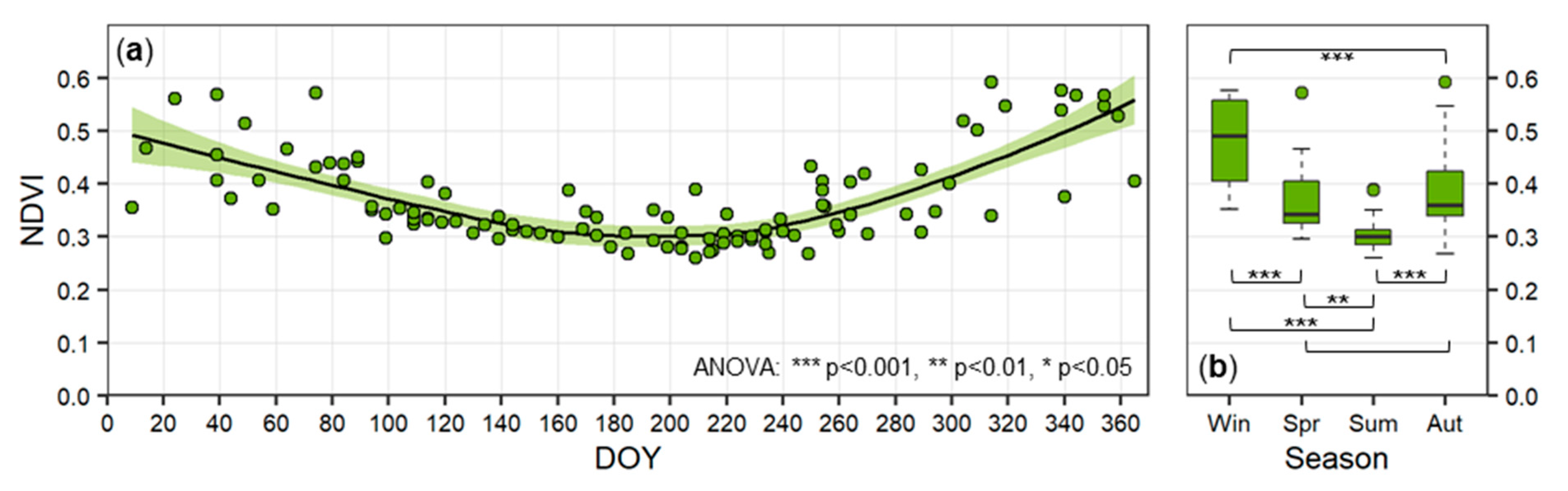
| Attribute (unit) | Relevance |
|---|---|
| Aspect (degree) | the landscape’s spatial heterogeneity affecting the surface energy balance and through this, the soil water retention capacity and water availability [88] |
| Elevation (m.a.s.l.) | the physical landscape properties and their spatial patterns, and key attributes for the further derivation of terrain-shape indices |
| Flow accumulation (-) | the effects of the depth and velocity of flow (here by integrating the multiple flow direction based on the maximum downslope gradient; top-down processing) [89] |
| Insolation (W m−2) | the annual sum of direct and diffuse potential incoming solar radiation calculated according to times of sunrise and sunset [90] |
| Topographic openness (positive and negative; rad m−1) | the dominance (positive) or enclosure (negative) of a landscape location [91] |
| Plan curvature (rad m−1) | the contour line formed by intersecting a horizontal plane with the surface, thus a proxy on the convergence or divergence of water during downslope flow [88] |
| Profile curvature (rad m−1) | the surface in the direction of the steepest slope, thus a proxy on the acceleration/deceleration of surface water [88] |
| Slope angle (degree) | the landscape’s spatial heterogeneity and catchment-related hydrological processes (e.g., flow direction, water runoff velocity and accumulation) [88] |
| Topographic Position Index (TPI; -) | the topographic slope positions and for landform classifications [92] |
| Terrain Ruggedness Index (TRI; -) | the surface heterogeneity of the landscape; averaged from the absolute differences in elevation between a focal cell and its eight neighboring DEM cells [93] |
| Topographic Wetness Index (TWI; -) | the topography’s spatial scale effect on hydrological processes and proxy on the terrain-related soil moisture potential [94] |
| Wind Exposition Index (WEI1; -) | the wind shadowed pixels/areas (values < 1) and pixels/area that are exposed to wind (values > 1) [95] |
| Wind Effect Index (WEI2; -) | The direction in which the wind is going (windward/leeward index) [95] |
| Statistics | Total | Biological Soil Crusts | Bare Soil | Grey Hair-Grass | Trees | ||||
|---|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Dry | Wet | Dry | Wet | ||
| Nr. of observations (i.e., pixels) | 535,875 | 208,967 | 86,190 | 6860 | 3021 | 149,595 | 65,775 | 11,036 | 4431 |
| Minimum | 0.073 | 0.138 | 0.248 | 0.073 | 0.078 | 0.090 | 0.099 | 0.278 | 0.402 |
| 1st quartile | 0.273 | 0.301 | 0.520 | 0.108 | 0.124 | 0.231 | 0.392 | 0.464 | 0.636 |
| Median | 0.329 | 0.329 | 0.548 | 0.123 | 0.159 | 0.255 | 0.451 | 0.514 | 0.681 |
| 3rd quartile | 0.450 | 0.364 | 0.575 | 0.141 | 0.199 | 0.275 | 0.494 | 0.575 | 0.736 |
| Maximum | 0.894 | 0.645 | 0.759 | 0.439 | 0.517 | 0.544 | 0.680 | 0.805 | 0.894 |
| Average | 0.361 | 0.335 | 0.546 | 0.126 | 0.168 | 0.251 | 0.438 | 0.522 | 0.686 |
| Standard deviation (SD) | 0.121 | 0.047 | 0.045 | 0.026 | 0.054 | 0.037 | 0.076 | 0.077 | 0.070 |
| Coefficient of variation (%) | 33.2 | 14.1 | 8.3 | 20.8 | 32.2 | 14.6 | 17.3 | 14.8 | 10.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rieser, J.; Veste, M.; Thiel, M.; Schönbrodt-Stitt, S. Coverage and Rainfall Response of Biological Soil Crusts Using Multi-Temporal Sentinel-2 Data in a Central European Temperate Dry Acid Grassland. Remote Sens. 2021, 13, 3093. https://doi.org/10.3390/rs13163093
Rieser J, Veste M, Thiel M, Schönbrodt-Stitt S. Coverage and Rainfall Response of Biological Soil Crusts Using Multi-Temporal Sentinel-2 Data in a Central European Temperate Dry Acid Grassland. Remote Sensing. 2021; 13(16):3093. https://doi.org/10.3390/rs13163093
Chicago/Turabian StyleRieser, Jakob, Maik Veste, Michael Thiel, and Sarah Schönbrodt-Stitt. 2021. "Coverage and Rainfall Response of Biological Soil Crusts Using Multi-Temporal Sentinel-2 Data in a Central European Temperate Dry Acid Grassland" Remote Sensing 13, no. 16: 3093. https://doi.org/10.3390/rs13163093
APA StyleRieser, J., Veste, M., Thiel, M., & Schönbrodt-Stitt, S. (2021). Coverage and Rainfall Response of Biological Soil Crusts Using Multi-Temporal Sentinel-2 Data in a Central European Temperate Dry Acid Grassland. Remote Sensing, 13(16), 3093. https://doi.org/10.3390/rs13163093