Urban Fire Severity and Vegetation Dynamics in Southern California


Abstract
1. Introduction
2. Methods
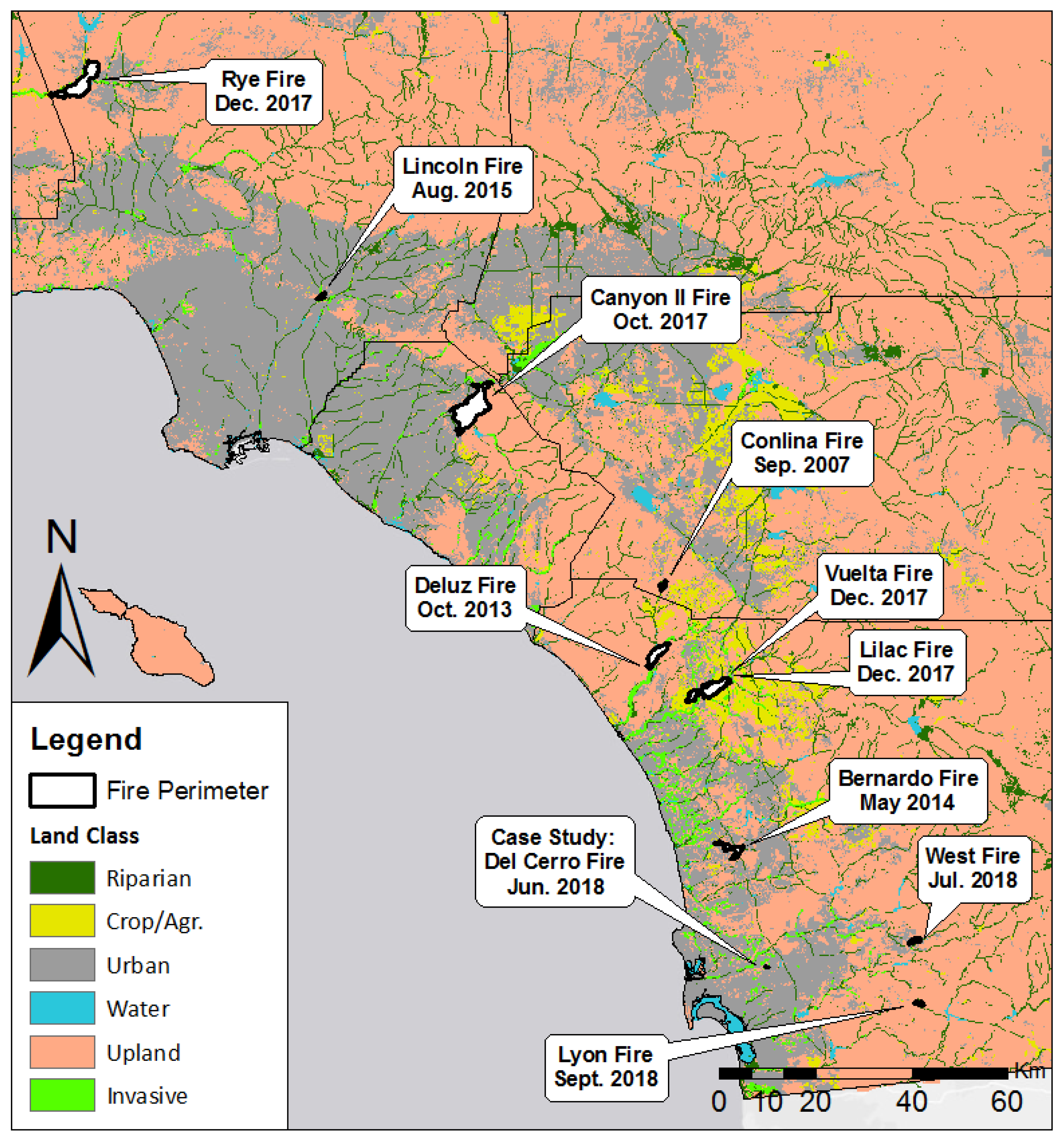
2.1. Study Area
2.2. Vegetation Classification
2.3. Burn Severity and Vegetation Metrics
2.4. Differenced Normalized Burn Ratio (dNBR)
2.5. Differenced Normalized Difference Vegetation Index (dNDVI)
2.6. Statistical Analysis
3. Results
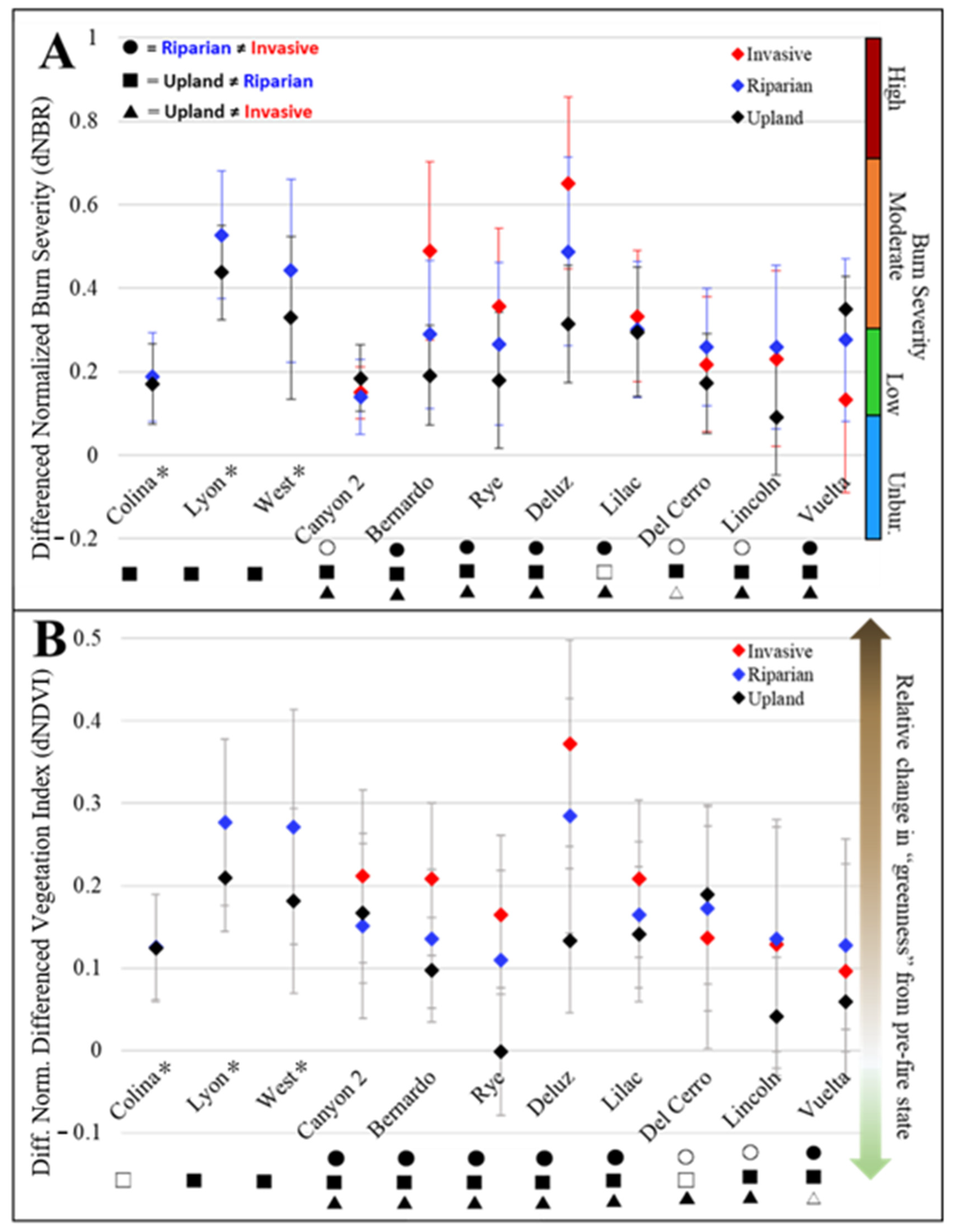
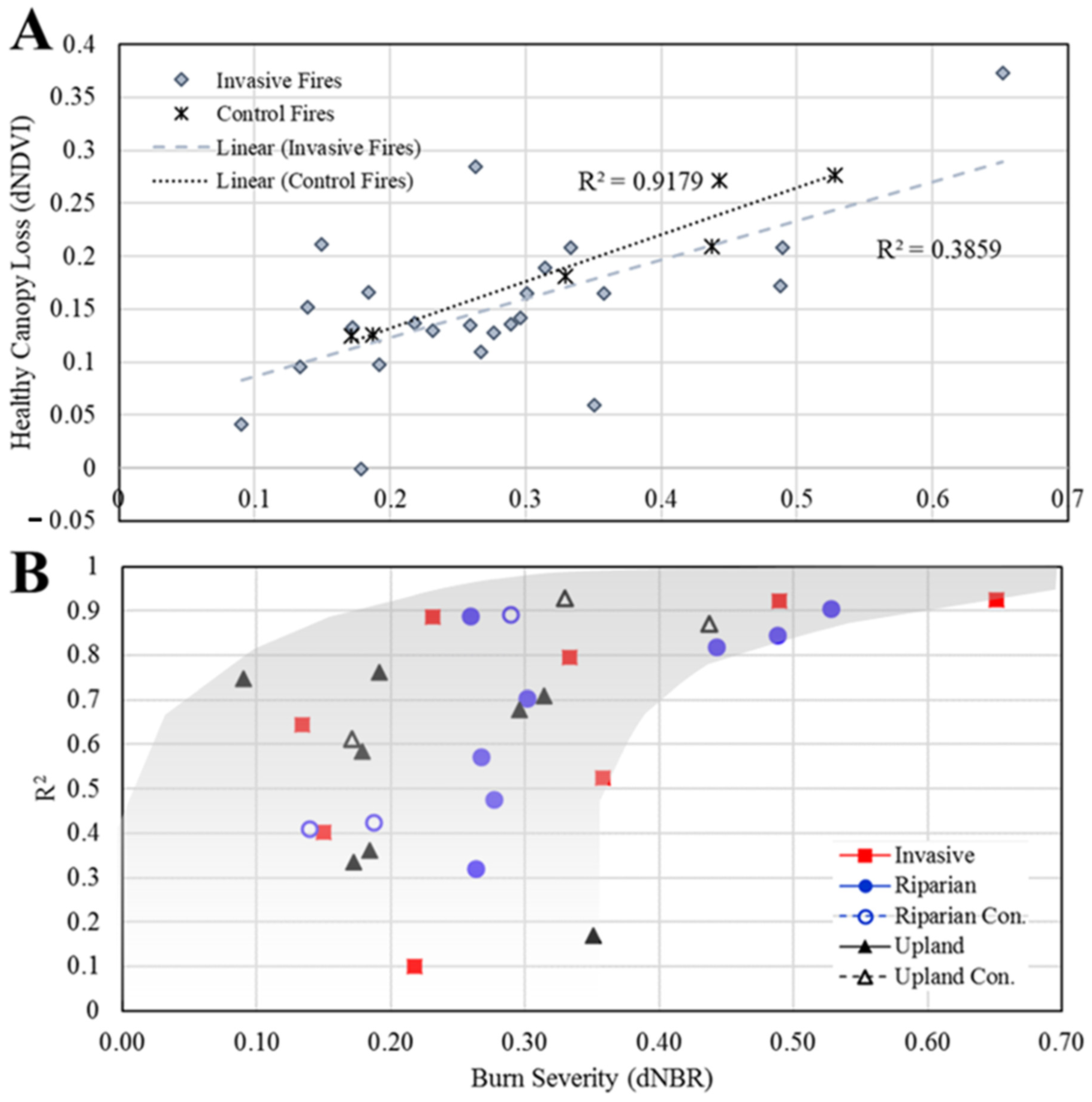
3.1. Immediate Burn Severity and Canopy Loss by Fire
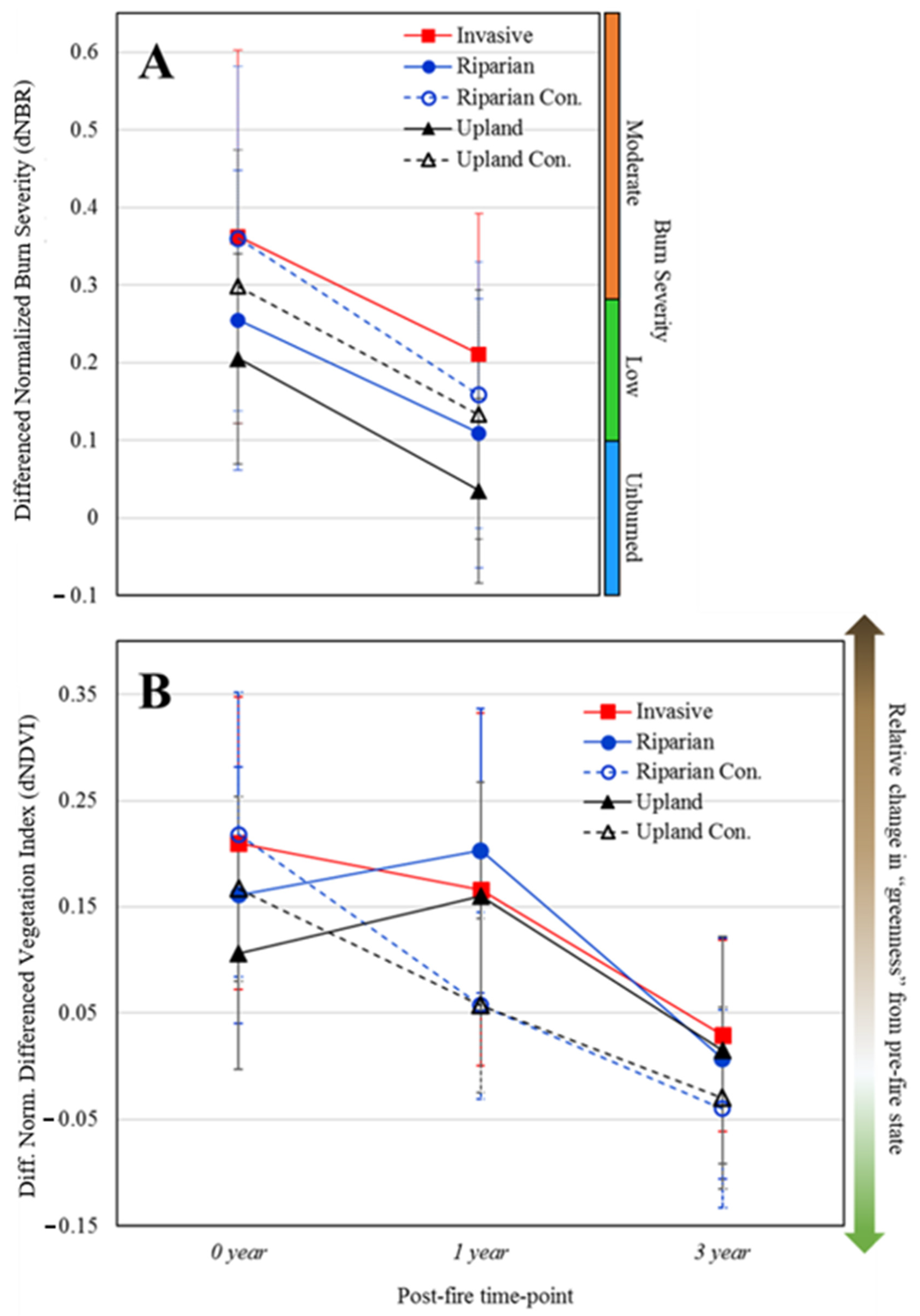
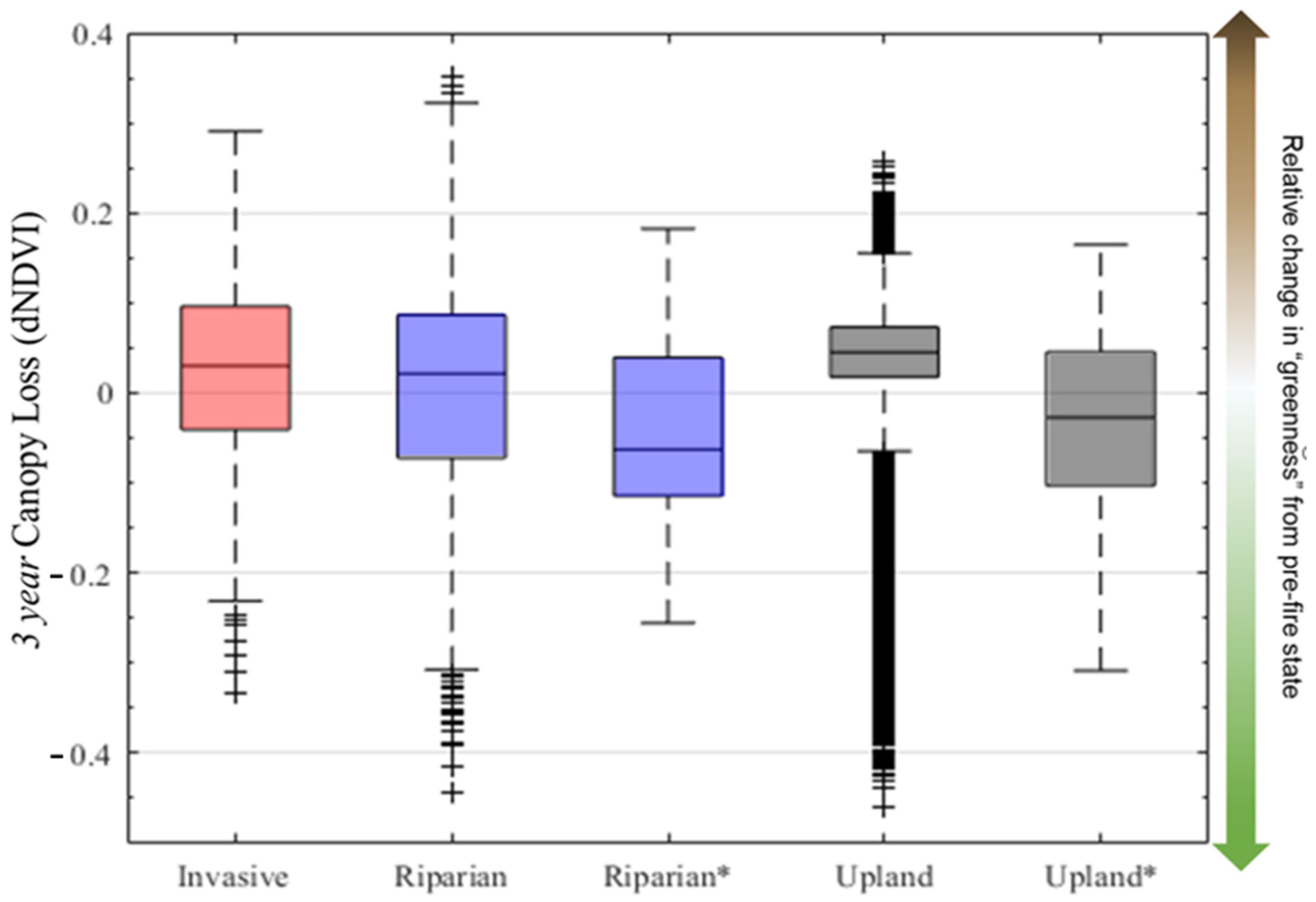
3.2. Average Burn Severity and Canopy Loss Patterns over Time
4. Discussion
4.1. Influence of Invasive Plants on Burn Severity and Green Canopy Loss after Fire
4.2. Evidence of the Grass–Fire Cycle
4.3. Highlighting the Uncertainty of Burn Severity and Canopy Loss Metrics in Urban Riparian Environments
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kinoshita, A.M.; Chin, A.; Simon, G.L.; Briles, C.; Hogue, T.S.; O’Dowd, A.P.; Gerlak, A.K.; Albornoz, A.U. Wildfire, water, and society: Toward integrative research in the “Anthropocene”. Anthropocene 2016, 16, 16–27. [Google Scholar] [CrossRef]
- Syphard, A.D.; Radeloff, V.C.; Keeley, J.E.; Hawbaker, T.J.; Clayton, M.K.; Stewart, S.I.; Hammer, R.B. Human Influence on California Fire Regimes. Ecol. Appl. 2007, 17, 1388–1402. [Google Scholar] [CrossRef] [PubMed]
- Syphard, A.D.; Brennan, T.J.; Keeley, J.E. Drivers of chaparral type conversion to herbaceous vegetation in coastal Southern California. Divers. Distrib. 2019, 25, 90–101. [Google Scholar] [CrossRef]
- Dwire, K.A.; Kauffman, J.B. Fire and riparian ecosystems in landscapes of the western USA. For. Ecol. Manag. 2003, 178, 61–74. [Google Scholar] [CrossRef]
- Verkaik, I.; Rieradevall, M.; Cooper, S.D.; Melack, J.M.; Dudley, T.L.; Prat, N. Fire as a disturbance in mediterranean climate streams. Hydrobiologia 2013, 719, 353–382. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C.J. Historic Fire Regime in Southern California Shrublands. Conserv. Biol. 2001, 15, 1536–1548. [Google Scholar] [CrossRef]
- White, M.D.; Greer, K.A. The effects of watershed urbanization on the stream hydrology and riparian vegetation of Los Peñasquitos Creek, California. Landsc. Urban Plan. 2006, 74, 125–138. [Google Scholar] [CrossRef]
- D’Antonio, C.M. Fire, plant invasions, and global changes. In Invasive Species in a Changing World, 2nd ed.; Island Press: Washington, DC, USA, 2000; pp. 65–93. ISBN 78-1559637824. [Google Scholar]
- Godefroid, S.; Koedam, N. Urban plant species patterns are highly driven by density and function of built-up areas. Landsc. Ecol. 2007, 22, 1227–1239. [Google Scholar] [CrossRef]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Coffman, G.C. Factors Influencing Invasion of Giant Reed (Arundo donax) in Riparian Ecosystems of Mediterranean-Type Climate Regions. Ph.D. Thesis, University of California, Los Angeles, CA, USA, 2007. [Google Scholar]
- Franco, M.G.; Mundo, I.A.; Veblen, T.T. Field-Validated Burn-Severity Mapping in North Patagonian Forests. Remote Sens. 2020, 12, 214. [Google Scholar] [CrossRef]
- Collins, K.M.; Penman, T.D.; Price, O.F. Some Wildfire Ignition Causes Pose More Risk of Destroying Houses than Others. PLoS ONE 2016, 11, e0162083. [Google Scholar] [CrossRef] [PubMed]
- Giessow, J.; Casanova, J.; Leclerc, R.; MacArthur, R.; Fleming, G. Arundo Donax (Giant Reed): Distribution and Impact Report; State Water Resources Control Board California Invasive Plant Council (Cal-IPC): Berkeley, CA, USA, 2011; pp. 1–240. [Google Scholar]
- Coffman, G.C.; Ambrose, R.F.; Rundel, P.W. Wildfire promotes dominance of invasive giant reed (Arundo donax) in riparian ecosystems. Biol. Invasions 2010, 12, 2723–2734. [Google Scholar] [CrossRef]
- Cushman, J.H.; Gaffney, K.A. Community-level consequences of invasion: Impacts of exotic clonal plants on riparian vegetation. Biol. Invasions 2010, 12, 2765–2776. [Google Scholar] [CrossRef]
- Pettit, N.E.; Naiman, R.J. Fire in the Riparian Zone: Characteristics and Ecological Consequences. Ecosystems 2007, 10, 673–687. [Google Scholar] [CrossRef]
- Lutes, D.C.; Keane, R.E.; Caratti, J.F.; Key, C.H.; Benson, N.C.; Sutherland, S.; Gangi, L.J. FIREMON: Fire Effects Monitoring and Inventory System; RMRS-GTR-164; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ft. Collins, CO, USA, 2006.
- Halofsky, J.E.; Hibbs, D.E. Controls on early post-fire woody plant colonization in riparian areas. For. Ecol. Manag. 2009, 258, 1350–1358. [Google Scholar] [CrossRef]
- Swain, D.L.; Langenbrunner, B.; Neelin, J.D.; Hall, A. Increasing precipitation volatility in twenty-first-century California. Nat. Clim. Chang. 2018, 8, 427–433. [Google Scholar] [CrossRef]
- US Geological Survey and US Department of Agriculture, Natural Resources Conservation Service. National Hydrography Dataset. Available online: https://www.usgs.gov/core-science-systems/ngp/national-hydrography/nhdplus-high-resolution (accessed on 1 January 2020).
- California Department of Forestry and Fire Protection. Fire Perimeters Data. In Fire and Resources Assessment Program (FRAP); California Department of Forestry and Fire Protection’s Fire and Resource Assessment Program (FRAP): Sacramento, CA, USA, 2018. [Google Scholar]
- California Department of Forestry and Fire Protection. Vegetation (FVEG) Database. In Fire and Resources Assessment Program (FRAP); California Department of Forestry and Fire Protection’s Fire and Resource Assessment Program (FRAP): Sacramento, CA, USA, 2015. [Google Scholar]
- US Geological Survey. Landsat 8 (L8) Data Users Handbook. Available online: https://prd-wret.s3.us-west-2.amazonaws.com/assets/palladium/production/atoms/files/LSDS-1574_L8_Data_Users_Handbook-v5.0.pdf (accessed on 1 January 2020).
- Lentile, L.B.; Morgan, P.; Hudak, A.T.; Bobbitt, M.J.; Lewis, S.A.; Smith, A.M.S.; Robichaud, P.R. Post-Fire Burn Severity and Vegetation Response Following Eight Large Wildfires Across the Western United States. Fire Ecol. 2007, 3, 91–108. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.C. Landscape Assessment (LA). In FIREMON: Fire Effects Monitoring and Inventory System; Lutes, D.C., Keane, R.E., Caratti, J.F., Key, C.H., Benson, N.C., Sutherland, S., Gangi, L.J., Eds.; General Technical Report RMRS-GTR-164-CD; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; pp. LA-1–LA-55. [Google Scholar]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Peñuelas, J.; Valentini, R. Relationships Between NDVI, Canopy Structure, and Photosynthesis in Three Californian Vegetation Types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef]
- Hope, A.; Tague, C.; Clark, R. Characterizing post-fire vegetation recovery of California chaparral using TM/ETM+ time-series data. Int. J. Remote Sens. 2007, 28, 1339–1354. [Google Scholar] [CrossRef]
- Tran, B.N.; Tanase, M.A.; Bennett, L.T.; Aponte, C. Evaluation of Spectral Indices for Assessing Fire Severity in Australian Temperate Forests. Remote Sens. 2018, 10, 1680. [Google Scholar] [CrossRef]
- Keeley, J.E.; Keeley, S.C. Post-Fire Regeneration of Southern California Chaparral. Am. J. Bot. 1981, 68, 524–530. [Google Scholar] [CrossRef]
- Sparks, A.M.; Kolden, C.A.; Talhelm, A.F.; Smith, A.M.S.; Apostol, K.G.; Johnson, D.M.; Boschetti, L. Spectral Indices Accurately Quantify Changes in Seedling Physiology Following Fire: Towards Mechanistic Assessments of Post-Fire Carbon Cycling. Remote Sens. 2016, 8, 572. [Google Scholar] [CrossRef]
- Holstein, G. Deciduous Islands in an Evergreen Sea. In California Riparian Forests: Ecology, Conservation and Productive Management; University of California Press: Berkeley, CA, USA, 1984. [Google Scholar]
- Davis, F.W.; Keller, E.A.; Parikh, A.; Florsheim, J. Recovery of the Chaparral Riparian Zone After Wildfire. In Proceedings of the California Riparian Systems Conference: Protection, Management, and Restoration for the 1990s, Davis, CA, USA, 22–24 September 1988; Gen. Tech. Rep. PSW-GTR-110. Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1989; pp. 194–203. [Google Scholar]
- Bendix, J.; Cowell, C.M. Impacts of Wildfire on the Composition and Structure of Riparian Forests in Southern California. Ecosystems 2010, 13, 99–107. [Google Scholar] [CrossRef]
- Minnich, R.A. Fire Mosaics in Southern California and Northern Baja California. Science 1983, 219, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Veraverbeke, S.; Lhermitte, S.; Verstraeten, W.W.; Goossens, R. The temporal dimension of differenced Normalized Burn Ratio (dNBR) fire/burn severity studies: The case of the large 2007 Peloponnese wildfires in Greece. Remote Sens. Environ. 2010, 114, 2548–2563. [Google Scholar] [CrossRef]
- Park, I.W.; Hooper, J.; Flegal, J.M.; Jenerette, G.D. Impacts of climate, disturbance and topography on distribution of herbaceous cover in Southern California chaparral: Insights from a remote-sensing method. Divers. Distrib. 2018, 24, 497–508. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire Management Impacts on Invasive Plants in the Western United States. Conserv. Biol. 2006, 20, 375–384. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; MacDermott, H.J.; Nichols, S.C.; Murphy, B.P. A grass–fire cycle eliminates an obligate-seeding tree in a tropical savanna. Ecol. Evol. 2014, 4, 4185–4194. [Google Scholar] [CrossRef]
- Perdue, R.E. Arundo donax—Source of musical reeds and industrial cellulose. Econ. Bot. 1958, 12, 368–404. [Google Scholar] [CrossRef]
- Bell, G.P. Ecology and Management of Arundo donax, and Approaches to Riparian Habitat Restoration in Southern California. Available online: https://ic.arc.losrios.edu/~veiszep/05spr2001/stelmok/g26files/attach5.html (accessed on 13 May 2020).
- Iverson, M.E.; Jackson, N.E.; Frandsen, P.; Douthit, S. The impact of Arundo donax on water resources. In Arundo Donax Workshop Proceedings, California Exotic Pest Plant Council, Riverside; Jackson, N.E., Frandsen, P., Duthoit, S., Eds.; Team Arundo Riverside: Ontario, CA, USA, 1994; pp. 19–26. [Google Scholar]
- D’Antonio, C.M.; Vitousek, P.M. Biological Invasions by Exotic Grasses, the Grass/Fire Cycle, and Global Change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Keeley, J.E.; Baer-Keeley, M.; Fotheringham, C.J. Alien Plant Dynamics Following Fire in Mediterranean-Climate California Shrublands. Ecol. Appl. 2005, 15, 2109–2125. [Google Scholar] [CrossRef]
- Keeley, J.E. Chaparral. In North American Terrestrial Vegetation; Cambridge University Press: Cambridge, UK, 2000; pp. 203–254. [Google Scholar]
- Keeley, J.E.; Brennan, T.; Pfaff, A.H. Fire severity and ecosytem responses following crown fires in California shrublands. Ecol. Appl. 2008, 18, 1530–1546. [Google Scholar] [CrossRef]
- Kobziar, L.N.; McBride, J.R. Wildfire burn patterns and riparian vegetation response along two northern Sierra Nevada streams. For. Ecol. Manag. 2006, 222, 254–265. [Google Scholar] [CrossRef]
- Park, I.W.; Jenerette, G.D. Causes and feedbacks to widespread grass invasion into chaparral shrub dominated landscapes. Landsc. Ecol. 2019, 34, 459–471. [Google Scholar] [CrossRef]
- Barro, S.C.; Conard, S.G. Fire effects on California chaparral systems: An overview. Environ. Int. 1991, 17, 135–149. [Google Scholar] [CrossRef]
- Coppoletta, M.; Merriam, K.E.; Collins, B.M. Post-fire vegetation and fuel development influences fire severity patterns in reburns. Ecol. Appl. 2016, 26, 686–699. [Google Scholar] [CrossRef]
- Morgan, P.; Keane, R.E.; Dillon, G.K.; Jain, T.B.; Hudak, A.T.; Karau, E.C.; Sikkink, P.G.; Holden, Z.A.; Strand, E.K. Challenges of assessing fire and burn severity using field measures, remote sensing and modelling. Int. J. Wildland Fire 2014, 23, 1045. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Ignition Date (m/d/y) | Containment Date (m/d/y) | County | Impacted River | Ignition type | Total Area Burned (km2) | Percent Invasive Cover | Percent Riparian Cover | Percent Upland Cover |
|---|---|---|---|---|---|---|---|---|---|
| Del Cerro | 6/3/2018 | 6/4/2018 | San Diego | Alvarado Creek | Juveniles | 0.15 | 8.02% | 37.04% | 53.09% |
| Colina Fire | 9/10/2007 | 9/10/2007 | Riverside | Deluz Creek | Vehicle Accident | 0.87 | 0.00% | 17.93% | 81.29% |
| Lincoln Fire | 8/16/2015 | Unknown | Los Angeles | Rio Hondo | Transient Camp | 0.95 | 14.23% | 41.17% | 37.25% |
| Lyon Fire | 9/9/2013 | 9/12/2013 | San Diego | Wilson Creek | Accidental | 1.06 | 0.00% | 9.87% | 90.13% |
| West | 7/6/2018 | 7/10/2018 | San Diego | Viejas Creek | Unknown | 2.04 | 0.00% | 17.65% | 33.98% |
| Bernardo | 5/13/2014 | 5/18/2014 | San Diego | Lusardi Creek | Sparks from power equipment | 6.26 | 1.25% | 29.43% | 64.99% |
| Vuelta Fire | 6/16/2007 | 6/18/2007 | San Diego | San Luis Rey River | Unknown- transient camp suspected | 9.02 | 37.64% | 58.01% | 4.03% |
| Deluz Fire | 10/5/2013 | 10/14/2013 | San Diego | Santa Margarita River | Unknown | 9.05 | 4.85% | 15.73% | 76.91% |
| Lilac Fire | 12/7/2017 | 12/16/2017 | San Diego | San Luis Rey River | Unknown | 16.5 | 6.23% | 12.19% | 30.25% |
| Rye Fire | 12/5/2017 | 12/13/2017 | Los Angeles | Santa Clara | Unknown | 24.5 | 1.89% | 6.51% | 78.44% |
| Canyon 2 | 10/9/2017 | 10/17/2017 | Orange | Santa Anna River and Santiago Creek | Embers from Canyon 1 Fire | 37.3 | 0.14% | 11.86% | 78.53% |
| Fire Name | Immediately Pre-Fire (1) | Immediately Post-Fire (2) | One-Year Post-Fire (3) | Three-Years Post-Fire (4) |
|---|---|---|---|---|
| Del Cerro | 16 May 2018 | 21 June 2018 | 19 June 2019 | N/A |
| Colina | 25 July 2007 | 11 September 2007 | 27 July 2008 | 17 July 2010 |
| Lincoln | 7 August 2015 | 10 October 2015 | 9 August 2016 | 15 August 2018 |
| Lyon | 10 August 2013 | 27 September 2013 | 29 August 2014 | 3 September 2016 |
| West | 21 June 2018 | 13 July 2018 | 8 June 2019 | N/A |
| Bernardo | 19 May 2014 | 29 May 2014 | 12 May 2015 | 1 May 2017 |
| Vuelta | 6 May 2007 | 25 June 2007 | 25 June 2008 | 20 May 2010 |
| Deluz | 11 September 2013 | 14 November 2013 | 30 September 2014 | 2 September 2016 |
| Lilac | 25 November 2017 | 27 December 2017 | 12 November 2018 | N/A |
| Rye | 2 December 2017 | 4 February 2018 | 26 November 2018 | N/A |
| Canyon 2 | 8 October 2017 | 24 October 2017 | 27 October 2018 | N/A |
| 0-Year Post-Fire | 1-Year Post-Fire | 3-Years Post-Fire | ||||
|---|---|---|---|---|---|---|
| Veg. Class | dNBR | dNDVI | dNBR | dNDVI | dNBR | dNDVI |
| Invasive | 0.362 ± 0.240 | 0.210 ± 0.138 | 0.211 ± 0.181 | 0.166 ± 0.166 | N/A | 0.029 ± 0.090 |
| Riparian | 0.255 ± 0.193 | 0.161 ± 0.121 | 0.109 ± 0.173 | 0.203 ± 0.134 | N/A | 0.007 ± 0.113 |
| Riparian * | 0.360 ± 0.222 | 0.218 ± 0.134 | 0.158 ± 0.172 | 0.057 ± 0.088 | N/A | −0.041 ± 0.093 |
| Upland | 0.205 ± 0.135 | 0.106 ± 0.109 | 0.035 ± 0.119 | 0.160 ± 0.107 | N/A | 0.015 ± 0.107 |
| Upland * | 0.298 ± 0.176 | 0.167 ± 0.087 | 0.133 ± 0.161 | 0.057 ± 0.082 | N/A | −0.033 ± 0.085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathews, L.E.H.; Kinoshita, A.M. Urban Fire Severity and Vegetation Dynamics in Southern California. Remote Sens. 2021, 13, 19. https://doi.org/10.3390/rs13010019
Mathews LEH, Kinoshita AM. Urban Fire Severity and Vegetation Dynamics in Southern California. Remote Sensing. 2021; 13(1):19. https://doi.org/10.3390/rs13010019
Chicago/Turabian StyleMathews, Lauren E. H., and Alicia M. Kinoshita. 2021. "Urban Fire Severity and Vegetation Dynamics in Southern California" Remote Sensing 13, no. 1: 19. https://doi.org/10.3390/rs13010019
APA StyleMathews, L. E. H., & Kinoshita, A. M. (2021). Urban Fire Severity and Vegetation Dynamics in Southern California. Remote Sensing, 13(1), 19. https://doi.org/10.3390/rs13010019