Phenology-Based Mapping of an Alien Invasive Species Using Time Series of Multispectral Satellite Data: A Case-Study with Glossy Buckthorn in Québec, Canada

 , ,
, ,  ,
,
Abstract
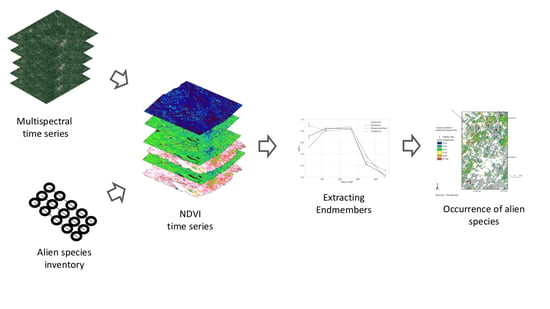
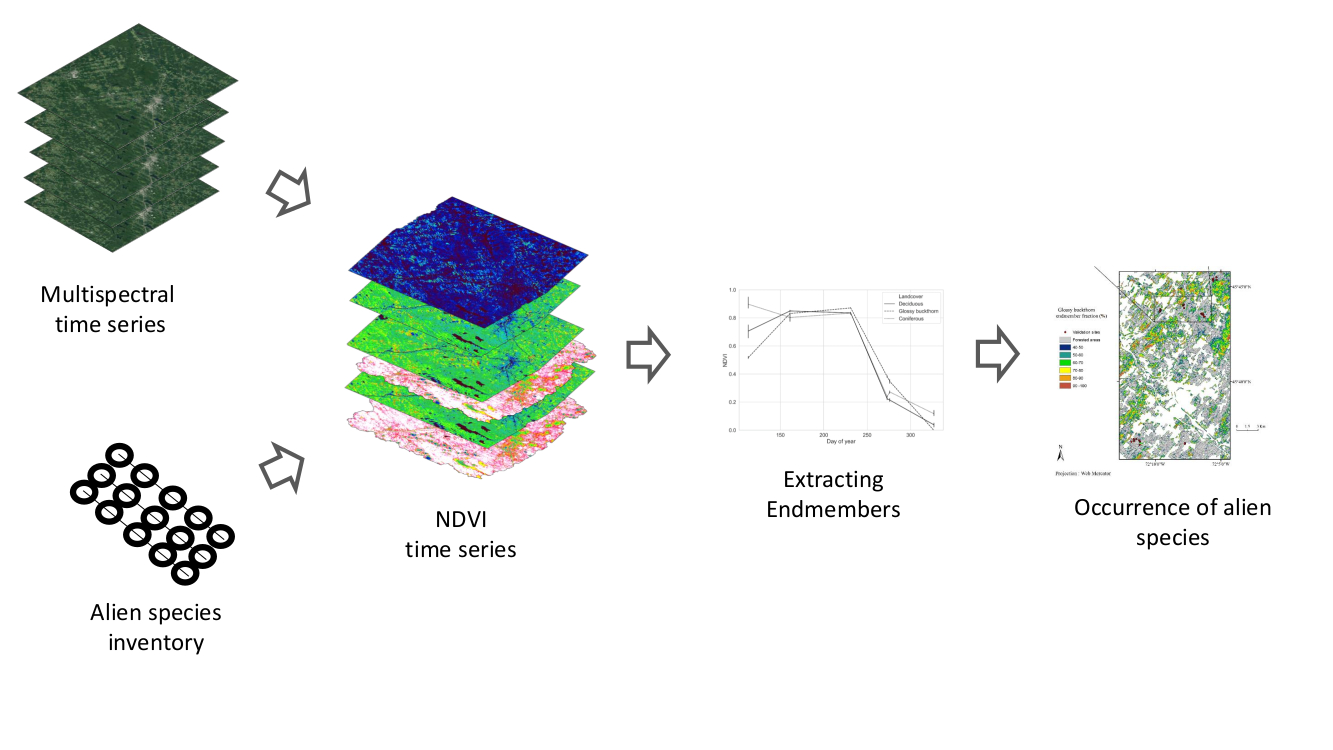
1. Introduction
2. Methods
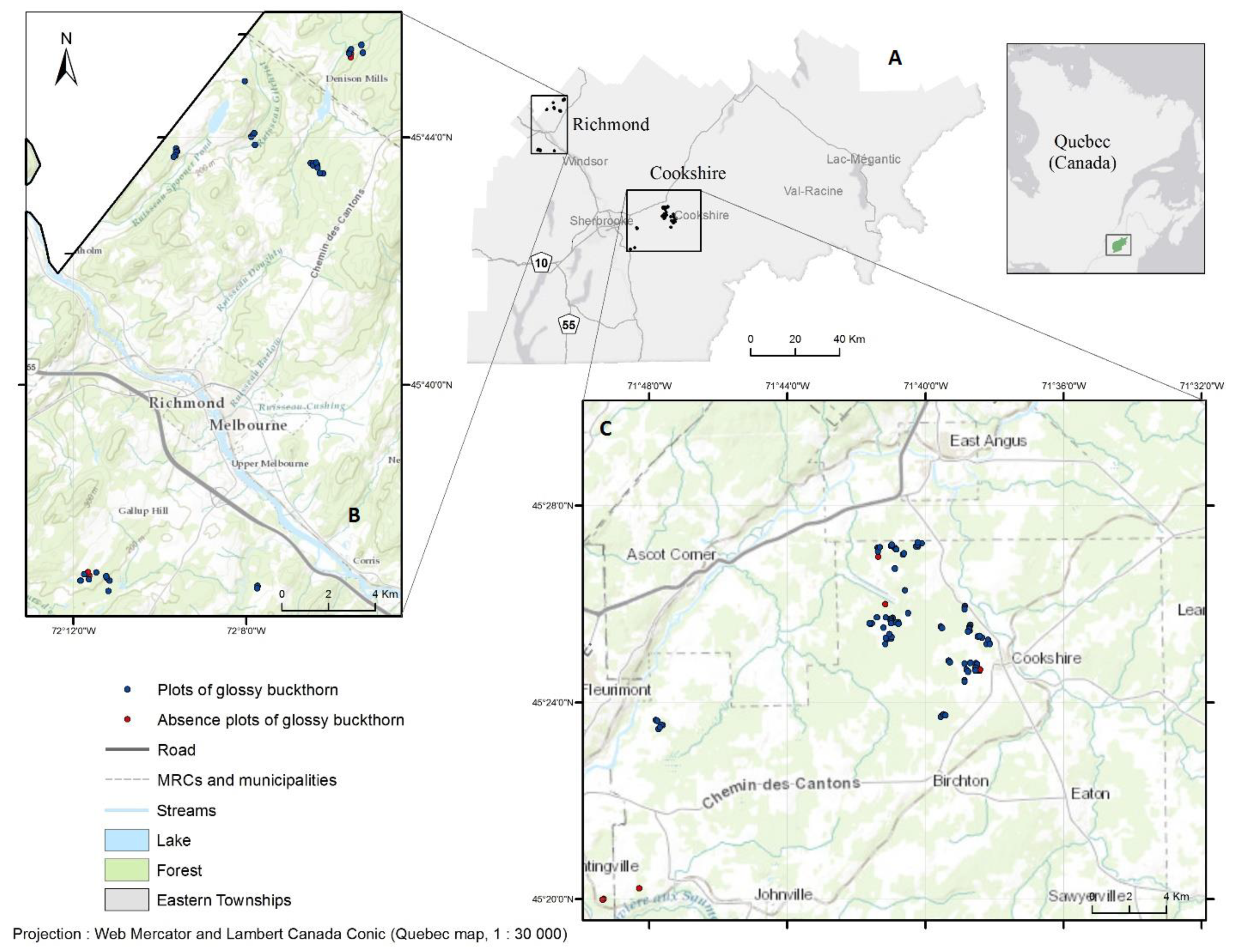
2.1. Study Area
2.2. Field Sampling
2.3. Processing of Landsat-8 OLI Images
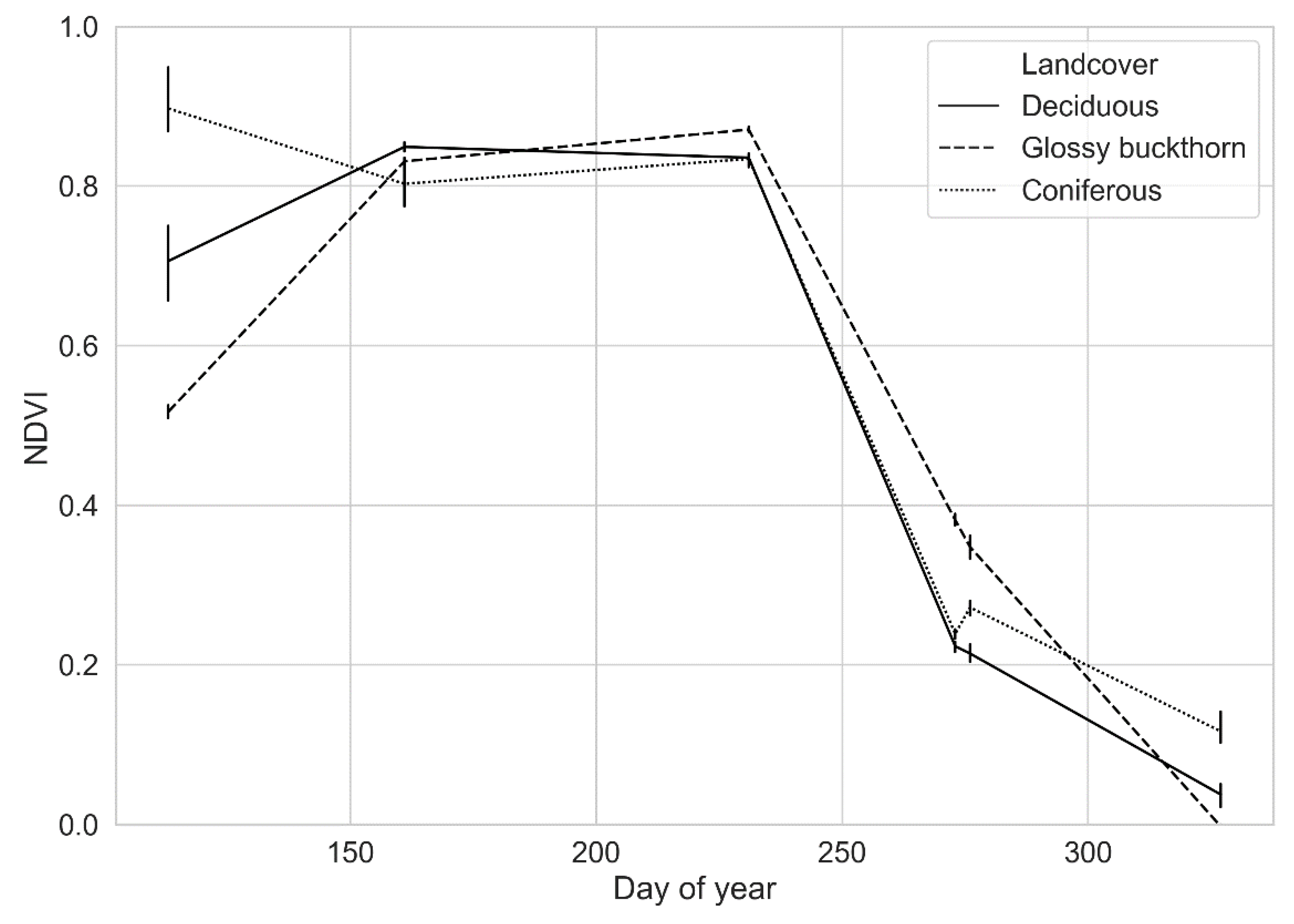
2.4. NDVI Times Series
2.5. Temporal Unmixing
3. Results
3.1. Field-Based Measurements of Glossy Buckthorn Cover
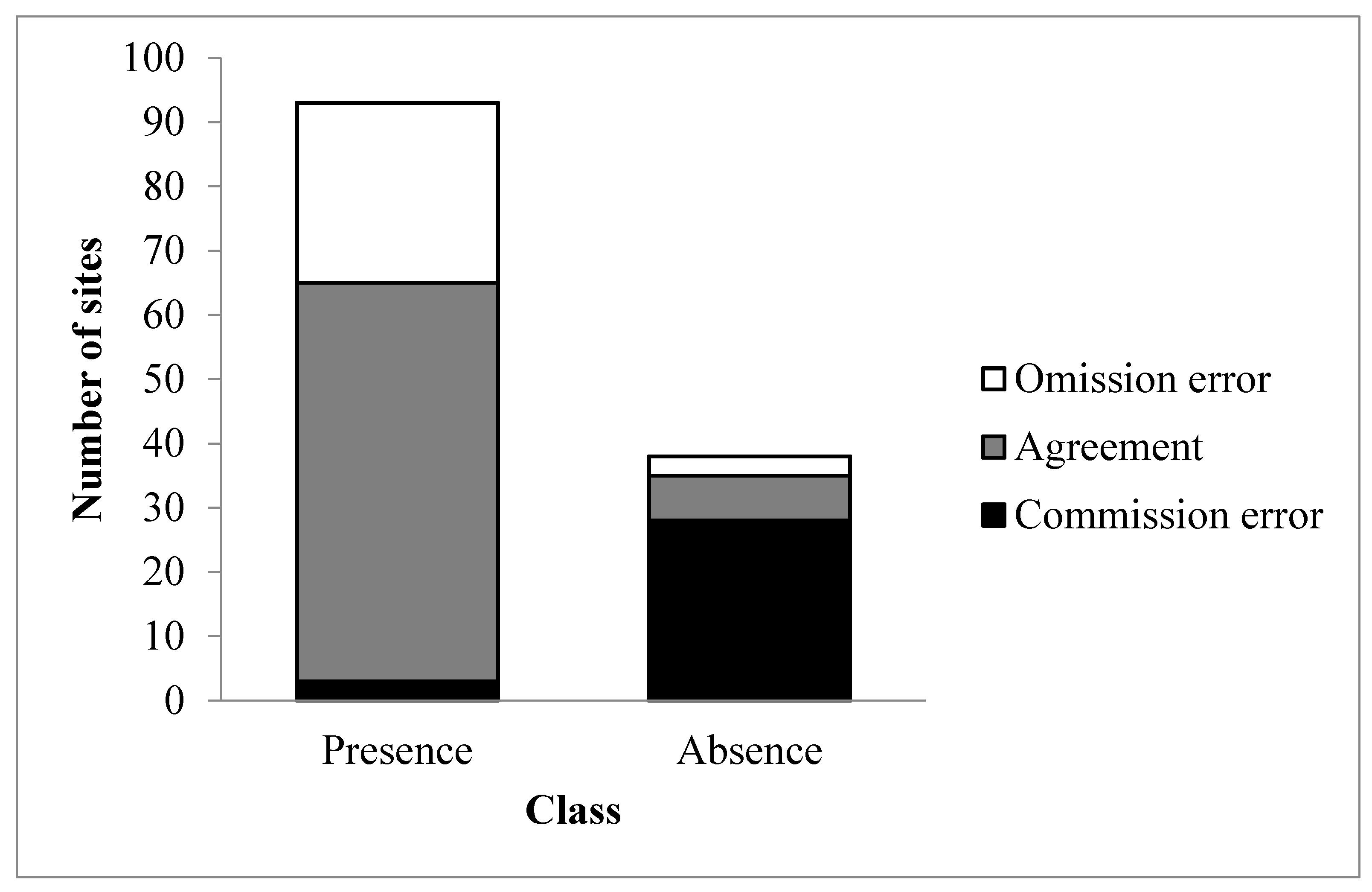
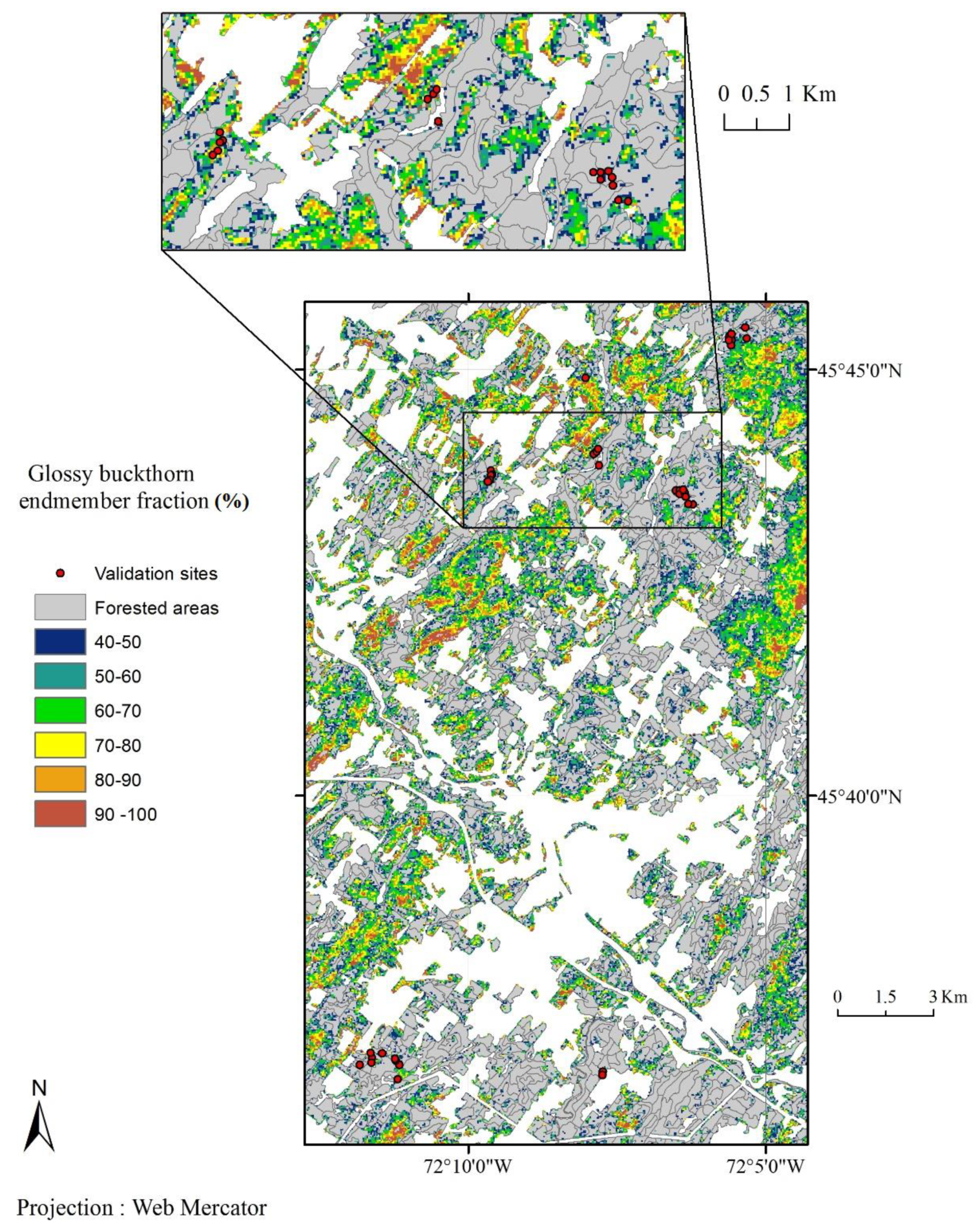
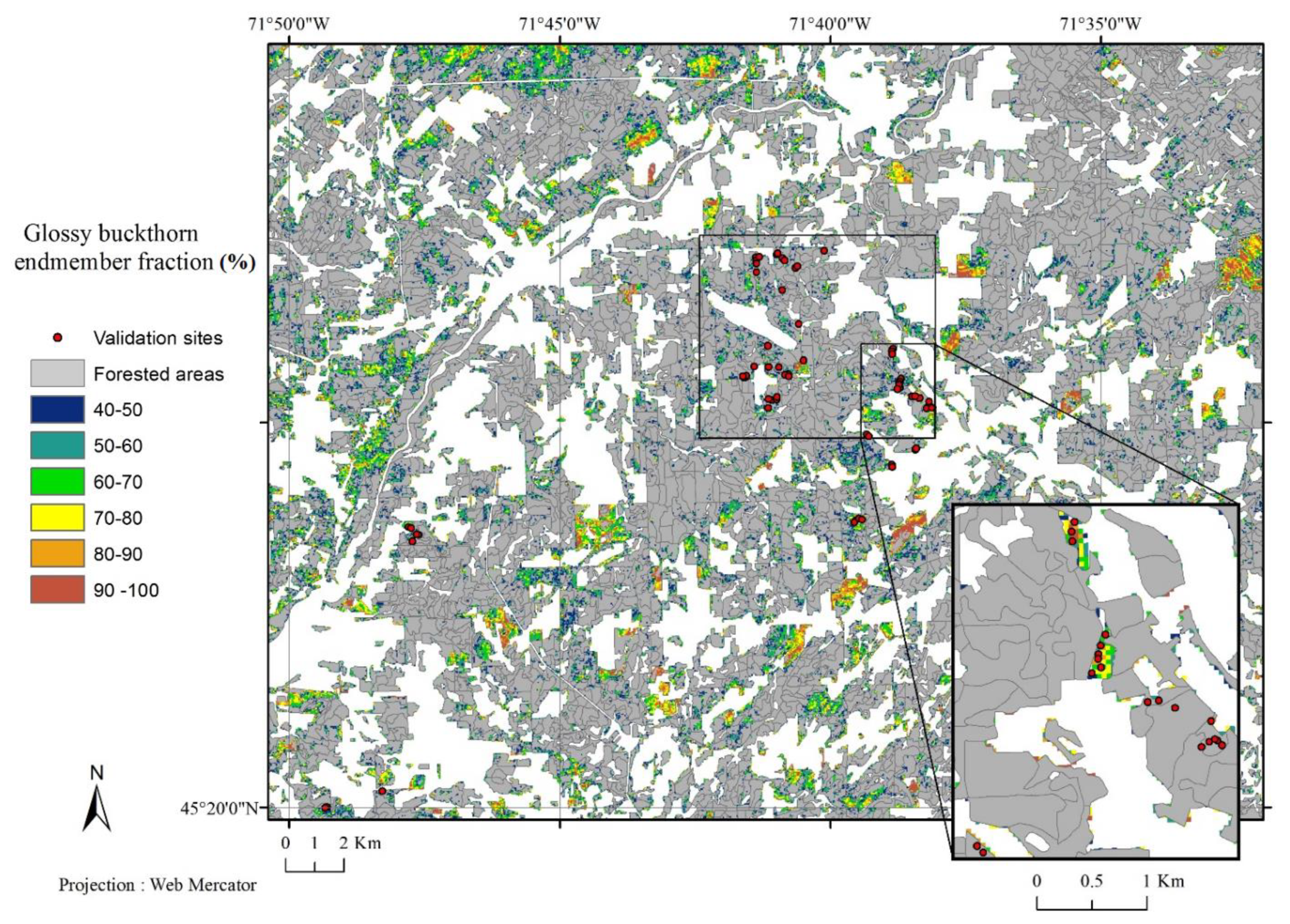
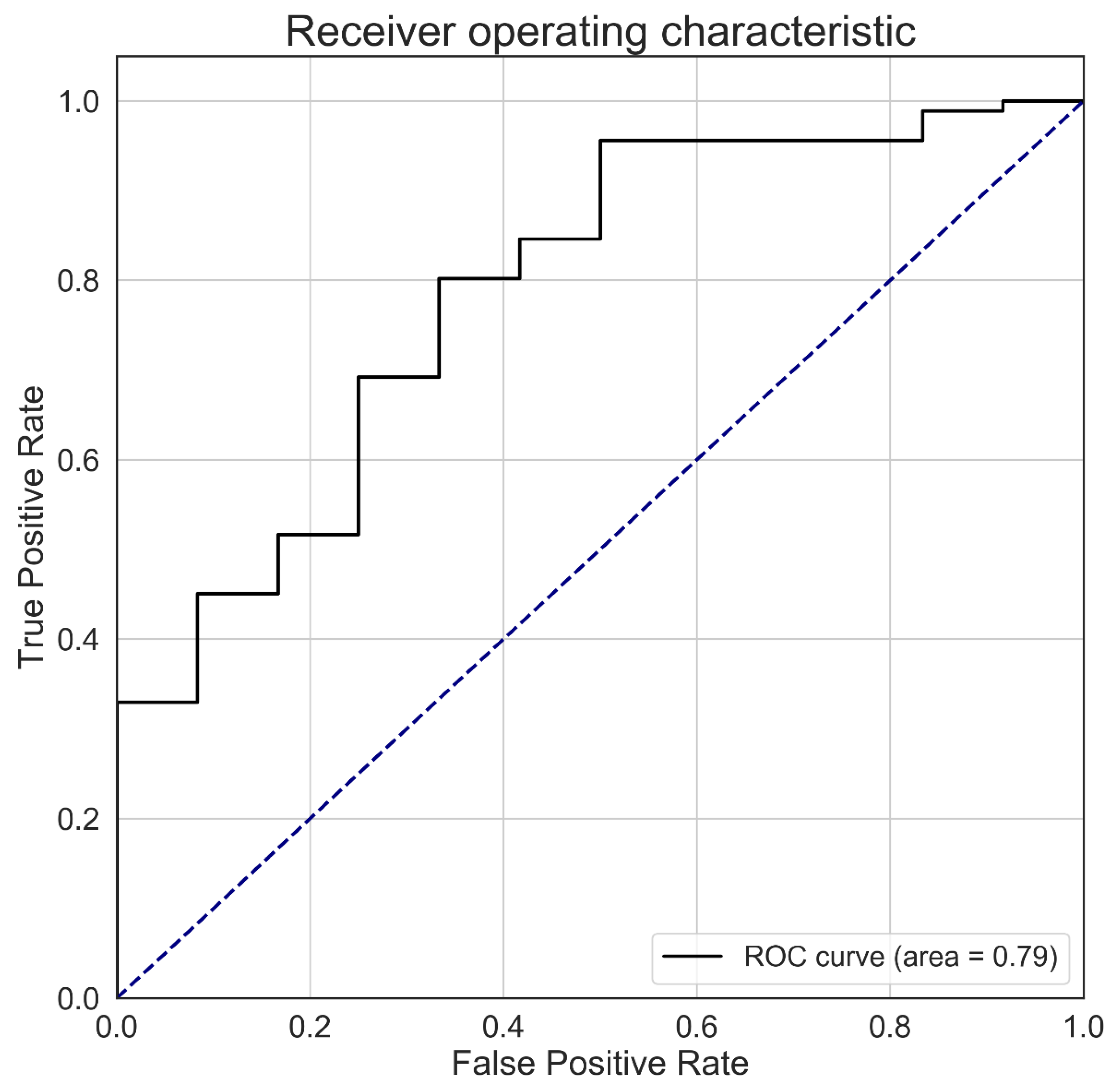
3.2. Validation of the Phenology-Based Map of Glossy Buckthorn Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatement | ||||||
|---|---|---|---|---|---|---|
| Deciduous vs Glossy Buck. | Deciduous vs Coniferous | Coniferous vs Glossy Buck. | ||||
| Day of year | F-Stats | P-Value | F-Stats | P-Value | F-Stats | P-Value |
| 113 | −7.1 | 0.000 | −5.2 | 0.002 | −14.5 | 0.004 |
| 161 | −4.4 | 0.001 | 3.0 | 0.086 | 1.8 | 0.198 |
| 231 | 12.4 | 0.000 | 0.3 | 0.808 | 6.5 | 0.011 |
| 273 | 28.4 | 0.000 | −3.3 | 0.016 | 25.9 | 0.000 |
| 276 | 13.7 | 0.000 | −7.2 | 0.000 | 8.1 | 0.000 |
| 327 | −4.9 | 0.001 | −5.2 | 0.008 | −9.2 | 0.010 |
Appendix B

References
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Knight, K.S.; Kurylo, J.S.; Endress, A.G.; Stewart, J.R.; Reich, P.B. Ecology and ecosystem impacts of common buckthorn (Rhamnus cathartica): A review. Biol. Invasions 2007, 9, 925–937. [Google Scholar] [CrossRef]
- Krumm, F.; Vítková, L. Introduced Tree Species in European Forests: Opportunities and Challenges; European Forest Institute: Berlin, Germany, 2016. [Google Scholar]
- Medan, D. Reproductive biology ofFrangula alnus (Rhamnaceae) in southern Spain. Plant Syst. Evol. 1994, 193, 173–186. [Google Scholar] [CrossRef]
- Converse, C.K. Element Stewardship Asbstrat for Rhamnus cathartica, Rhamnus frangula (syn. Frangula alnus); The Nature Conservancy: Arlington, VA, USA, 1984. [Google Scholar]
- Howell, J.A.; Blackwell, W.H., Jr. The history of Rhamnus frangula (glossy buckthorn) in the Ohio flora. Castanea 1977, 42, 111–115. [Google Scholar]
- Catling, P.M.; Porebski, Z.S. The History of Invasion and Current Status of Glossy Buckthorn, Rhamnus-Frangula, in Southern Ontario. Can. Field-Nat. 1994, 108, 305–310. [Google Scholar]
- Haber, E. Spread and impact of alien plants across Canadian landscapes. In Alien Invaders in Canada’s Waters, Wetlands and Forests; Canadian Forest Service: Ottawa, ON, Canada, 2002; pp. 43–57. [Google Scholar]
- Fagan, M.E.; Peart, D.R. Impact of the invasive shrub glossy buckthorn (Rhamnus frangula L.) on juvenile recruitment by canopy trees. For. Ecol. Manag. 2004, 194, 95–107. [Google Scholar] [CrossRef]
- Hamelin, C.; Truax, B.; Gagnon, D. Invasive glossy buckthorn impedes growth of red oak and sugar maple under-planted in a mature hybrid poplar plantation. New For. 2016, 47, 897–911. [Google Scholar] [CrossRef]
- Frappier, B.; Eckert, R.T.; Lee, T.D. Experimental removal of the non-indigenous shrub Rhamnus frangula (glossy buckthorn): Effects on native herbs and woody seedlings. Northeast. Nat. 2004, 11, 333–342. [Google Scholar] [CrossRef]
- Beaudet, M.; Cauboue, M.; Thiffault, N.; Cartier, P.; Martineau, P.; Boulet, B. L’autécologie des especes concurrentes. In Le Guide Sylvicole du Québec; Les publications du Québec: Québec, QC, Canada, 2013; Volume 1, pp. 180–279. [Google Scholar]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Madden, M. Remote Sensing and Geographic Information System Operations for Vegetation Mapping of Invasive Exotics. Weed Technol. 2004, 18, 1457–1463. [Google Scholar] [CrossRef]
- Wang, N. Application of Remote Sensing in Detecting and Monitoring Forest Regeneration Process in a Disturbed Environment. M.Sc. Thesis, Carleton University, Ottawa, ON, Canada, 1995. [Google Scholar]
- Zweig, C.L.; Newman, S. Using landscape context to map invasive species with medium-resolution satellite imagery. Restor. Ecol. 2015, 23, 524–530. [Google Scholar] [CrossRef]
- Frazier, A.E.; Wang, L. Characterizing spatial patterns of invasive species using sub-pixel classifications. Remote Sens. Environ. 2011, 115, 1997–2007. [Google Scholar] [CrossRef]
- Van Lier, O.R.; Fournier, R.A.; Bradley, R.L.; Thiffault, N. A multi-resolution satellite imagery approach for large area mapping of ericaceous shrubs in Northern Quebec, Canada. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 334–343. [Google Scholar] [CrossRef]
- Miao, X.; Gong, P.; Swope, S.; Pu, R.; Carruthers, R.; Anderson, G.L. Detection of yellow starthistle through band selection and feature extraction from hyperspectral imagery. Photogramm. Eng. Remote Sens. 2007, 73, 1005. [Google Scholar]
- Andrew, M.E.; Ustin, S.L. The role of environmental context in mapping invasive plants with hyperspectral image data. Remote Sens. Environ. 2008, 112, 4301–4317. [Google Scholar] [CrossRef]
- Tuanmu, M.-N.; Viña, A.; Bearer, S.; Xu, W.; Ouyang, Z.; Zhang, H.; Liu, J. Mapping understory vegetation using phenological characteristics derived from remotely sensed data. Remote Sens. Environ. 2010, 114, 1833–1844. [Google Scholar] [CrossRef]
- Xu, C.-Y.; Griffin, K.L.; Schuster, W. Leaf phenology and seasonal variation of photosynthesis of invasive Berberis thunbergii (Japanese barberry) and two co-occurring native understory shrubs in a northeastern United States deciduous forest. Oecologia 2007, 154, 11–21. [Google Scholar] [CrossRef]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359. [Google Scholar] [CrossRef]
- Polgar, C.; Gallinat, A.; Primack, R.B. Drivers of leaf-out phenology and their implications for species invasions: Insights from Thoreau’s Concord. New Phytol. 2014, 202, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E. Estimating cover of an invasive grass (Bromus tectorum) using tobit regression and phenology derived from two dates of Landsat ETM + data. Int. J. Remote Sens. 2005, 26, 2491–2507. [Google Scholar] [CrossRef]
- Resasco, J.; Hale, A.; Henry, M.; Gorchov, D. Detecting an invasive shrub in a deciduous forest understory using late-fall Landsat sensor imagery. Int. J. Remote Sens. 2007, 28, 3739–3745. [Google Scholar] [CrossRef]
- Shiferaw, H.; Schaffner, U.; Bewket, W.; Alamirew, T.; Zeleke, G.; Teketay, D.; Eckert, S. Modelling the current fractional cover of an invasive alien plant and drivers of its invasion in a dryland ecosystem. Sci. Rep. 2019, 1. [Google Scholar] [CrossRef] [PubMed]
- Radoux, J.; Chomé, G.; Jacques, D.C.; Waldner, F.; Bellemans, N.; Matton, N.; Lamarche, C.; D’Andrimont, R.; Defourny, P. Sentinel-2’s Potential for Sub-Pixel Landscape Feature Detection. Remote Sens. 2016, 8, 488. [Google Scholar] [CrossRef]
- Müllerová, J.; Brůna, J.; Bartaloš, T.; Dvořák, P.; Vítková, M.; Pyšek, P. Timing is important: Unmanned aircraft vs. satellite imagery in plant invasion monitoring. Front. Plant Sci. 2017, 8, 887. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Saucier, J.-P.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Écologie Forestière—Manuel de Foresterie, 2nd ed.; Doucet, R., Côté, M., Eds.; Ordre des ingénieurs forestiers du Québec: Québec, QC, Canada, 2009; pp. 186–205. [Google Scholar]
- Environnement Canada. Normales et Moyennes Climatiques de la Région de l’Estrie; Environnement Canada: Ottawa, ON, Canada, 2016.
- Laliberté, F.; Gauthier, J.; Boileau, J.F.; Chauvette, B. Portrait de la Forêt Naturelle et des Enjeux Écologiques de l’Estrie. Master’s Thesis, Université de Montréal, Montreal, QC, Canada, 2015; 114p. [Google Scholar]
- Ministère des Forêts, de la Faune et des Parcs (MFFP). Norme de Stratification Écoforestière du 4e Inventaire Écoforestier du Québec Méridional; Gouvernement du Québec: Québec, QC, Canada, 2015; 111p. Available online: http://www.mffp.gouv.qc.ca/forets/inventaire/pdf/norme-stratification.pdf (accessed on 3 February 2020).
- Campbell, J.B.; Wynne, R.H. Introduction to Remote Sensing, 5th ed.; The Guilford Press: New York, NY, USA, 2011; 667p. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974; 547p. [Google Scholar]
- Vicente-Serrano, S.M.; Pérez-Cabello, F.; Lasanta, T. Assessment of radiometric correction techniques in analyzing vegetation variability and change using time series of Landsat images. Remote Sens. Environ. 2008, 112, 3916–3934. [Google Scholar] [CrossRef]
- Song, C.; Woodcock, C.E.; Seto, K.C.; Lenney, M.P.; Macomber, S.A. Classification and change detection using Landsat TM data: When and how to correct atmospheric effects? Remote Sens. Environ. 2001, 75, 230–244. [Google Scholar] [CrossRef]
- Geomatica, version 10; PCI Geomatics: Markham, ON, Canada, 2015.
- Natural Resources Canada. Canadian Digital Elevation Model Product Specifications, Edition 1.1; Government of Canada: Québec, QC, Canada, 2013. Available online: http://ftp.maps.canada.ca/pub/nrcan_rncan/elevation/cdem_mnec/doc/CDEM_product_specs.pdf (accessed on 3 February 2020).
- Adams, J.B.; Gillespie, A.R. Remote Sensing of Landscapes with Spectral Images: A Physical Modeling Approach, 1st ed.; Cambridge University Press: Cambridge, UK, 2006; 378p. [Google Scholar]
- ENVI Image Analysis Software, version 10.1.; Harris Geospatial: Broomfield, CO, USA, 2011.
- Becker, R.H.; Zmijewski, K.A.; Crail, T. Seeing the forest for the invasives: Mapping buckthorn in the Oak Openings. Biol. Invasions 2012, 15, 315–326. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: A review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Eriksson, H.M.; Eklundh, L.; Kuusk, A.; Nilson, T. Impact of understory vegetation on forest canopy reflectance and remotely sensed LAI estimates. Remote Sens. Environ. 2006, 103, 408–418. [Google Scholar] [CrossRef]
- Kuusk, A.; Lang, M.; Nilson, T. Simulation of the reflectance of ground vegetation in sub-boreal forests. Agric. For. Meteorol. 2004, 126, 33–46. [Google Scholar] [CrossRef]
- Peltoniemi, J.I.; Kaasalainen, S.; Näränen, J.; Rautiainen, M.; Stenberg, P.; Smolander, H.; Smolander, S.; Voipio, P. BRDF measurement of understory vegetation in pine forests: Dwarf shrubs, lichen, and moss. Remote Sens. Environ. 2005, 94, 343–354. [Google Scholar] [CrossRef]
- Rautiainen, M.; Suomalainen, J.; Mõttus, M.; Stenberg, P.; Voipio, P.; Peltoniemi, J.; Manninen, T. Coupling forest canopy and understory reflectance in the Arctic latitudes of Finland. Remote Sens. Environ. 2007, 110, 332–343. [Google Scholar] [CrossRef]
- Dobigeon, N.; Altmann, Y.; Brun, N.; Moussaoui, S. Linear and nonlinear unmixing in hyperspectral imaging. In Resolving Spectral Mixtures—With Application from Ultrafast Spectroscopy to Super-Resolution Imaging; Elsevier: Amsterdam, The Netherlands, 2016; p. 45. [Google Scholar]
- Royimani, L.; Mutanga, O.; Odindi, J.; Dube, T.; Matongera, T.N. Advancements in satellite remote sensing for mapping and monitoring of alien invasive plant species (AIPs). Phys. Chem. Earth 2019, 112, 237–245. [Google Scholar] [CrossRef]
- Andrew, M.E.; Ustin, S.L. Habitat suitability modelling of an invasive plant with advanced remote sensing data. Divers. Distrib. 2009, 15, 627–640. [Google Scholar] [CrossRef]
- Kim, S.; Hinckley, T.; Briggs, D. Classifying tree species using structure and spectral data from LIDAR. In Proceedings of the ASPRS/MAPPS 2009 Specialty Conference, San Antonio, TX, USA, 16–19 November 2009. [Google Scholar]
- Reitberger, J.; Krzystek, P.; Stilla, U. Analysis of full waveform LIDAR data for the classification of deciduous and coniferous trees. Int. J. Remote Sens. 2008, 29, 1407–1431. [Google Scholar] [CrossRef]
- Stenberg, P.; Mottus, M.; Rautiainen, M. Modeling the spectral signature of forests: Application of remote sensing models to coniferous canopies. In Advances in Land Remote Sensing; Springer: Berlin, Germany, 2008; pp. 147–171. [Google Scholar]
- Asner, G.P.; Hughes, R.F.; Vitousek, P.M.; Knapp, D.E.; Kennedy-Bowdoin, T.; Boardman, J.; Martin, R.E.; Eastwood, M.; Green, R.O. Invasive plants transform the three-dimensional structure of rain forests. Proc. Natl. Acad. Sci. USA 2008, 105, 4519–4523. [Google Scholar] [CrossRef]




| Date | Day of Year | Cloud Cover (%) |
|---|---|---|
| 23 April 2013 | 113 | 3 |
| 10 June 2013 | 161 | 1 |
| 19 August 2015 | 231 | 7 |
| 30 September 2013 | 273 | 2 |
| 3 October 2014 | 276 | 2 |
| 23 November 2015 | 327 | 3 |
| Variables | Richmond and Cookshire | Richmond | Cookshire | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | S.D. | Min. | Max. | Mean | S.D. | Min. | Max. | Mean | S.D. | Min. | Max. | |
| Buckthorn cover (%) | 62.6 | 21.8 | 0 | 89 | 57.9 | 22.6 | 0 | 85 | 65.3 | 21.1 | 0 | 89 |
| Tree cover (%) | 62.3 | 18.5 | 33 | 90 | 69.6 | 13 | 33 | 90 | 58.2 | 19.9 | 33 | 90 |
| Observed | ||||
| Presence | Absence | Total | ||
| Predicted | Presence | 62 | 3 | 65 |
| Absence | 28 | 7 | 35 | |
| Total | 90 | 10 | 100 | |
| Glossy Buckthorn Cover (%) | |||
|---|---|---|---|
| ≥25% | ≥65% | ≥75% | |
| Agreement (%) | 69 | 74 | 80 |
| Disagreement (%) | 31 | 26 | 20 |
| Total (%) | 100 | 100 | 100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labonté, J.; Drolet, G.; Sylvain, J.-D.; Thiffault, N.; Hébert, F.; Girard, F. Phenology-Based Mapping of an Alien Invasive Species Using Time Series of Multispectral Satellite Data: A Case-Study with Glossy Buckthorn in Québec, Canada. Remote Sens. 2020, 12, 922. https://doi.org/10.3390/rs12060922
Labonté J, Drolet G, Sylvain J-D, Thiffault N, Hébert F, Girard F. Phenology-Based Mapping of an Alien Invasive Species Using Time Series of Multispectral Satellite Data: A Case-Study with Glossy Buckthorn in Québec, Canada. Remote Sensing. 2020; 12(6):922. https://doi.org/10.3390/rs12060922
Chicago/Turabian StyleLabonté, Joanie, Guillaume Drolet, Jean-Daniel Sylvain, Nelson Thiffault, Francois Hébert, and Francois Girard. 2020. "Phenology-Based Mapping of an Alien Invasive Species Using Time Series of Multispectral Satellite Data: A Case-Study with Glossy Buckthorn in Québec, Canada" Remote Sensing 12, no. 6: 922. https://doi.org/10.3390/rs12060922
APA StyleLabonté, J., Drolet, G., Sylvain, J.-D., Thiffault, N., Hébert, F., & Girard, F. (2020). Phenology-Based Mapping of an Alien Invasive Species Using Time Series of Multispectral Satellite Data: A Case-Study with Glossy Buckthorn in Québec, Canada. Remote Sensing, 12(6), 922. https://doi.org/10.3390/rs12060922