Tree Line Identification and Dynamics under Climate Change in Wuyishan National Park Based on Landsat Images



Abstract
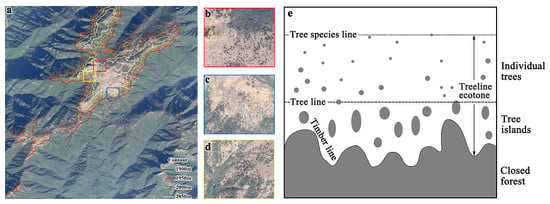
1. Introduction
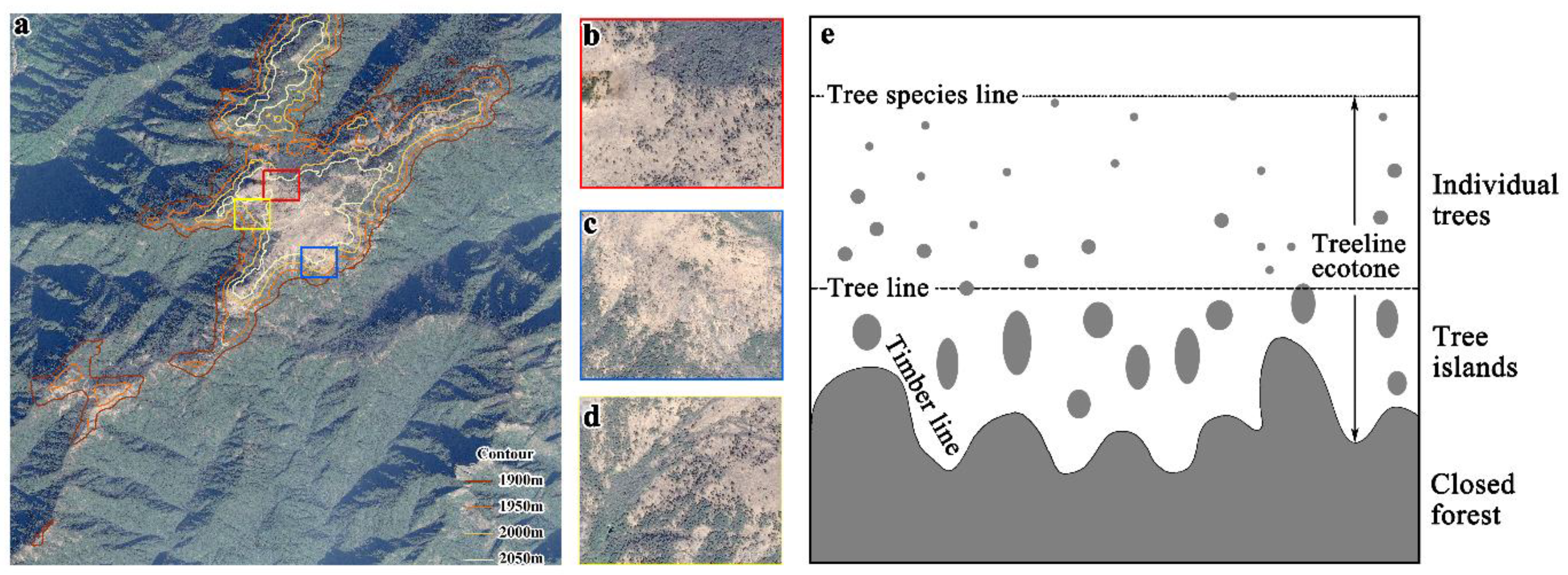
1.1. Alpine Tree Line Ecotone
1.2. Remote Sensing Applications for Tree Line Dynamics
2. Study Area
3. Materials and Methods
3.1. Datasets
3.1.1. Remotely Sensed Imagery
3.1.2. Climate Data
3.2. Methods
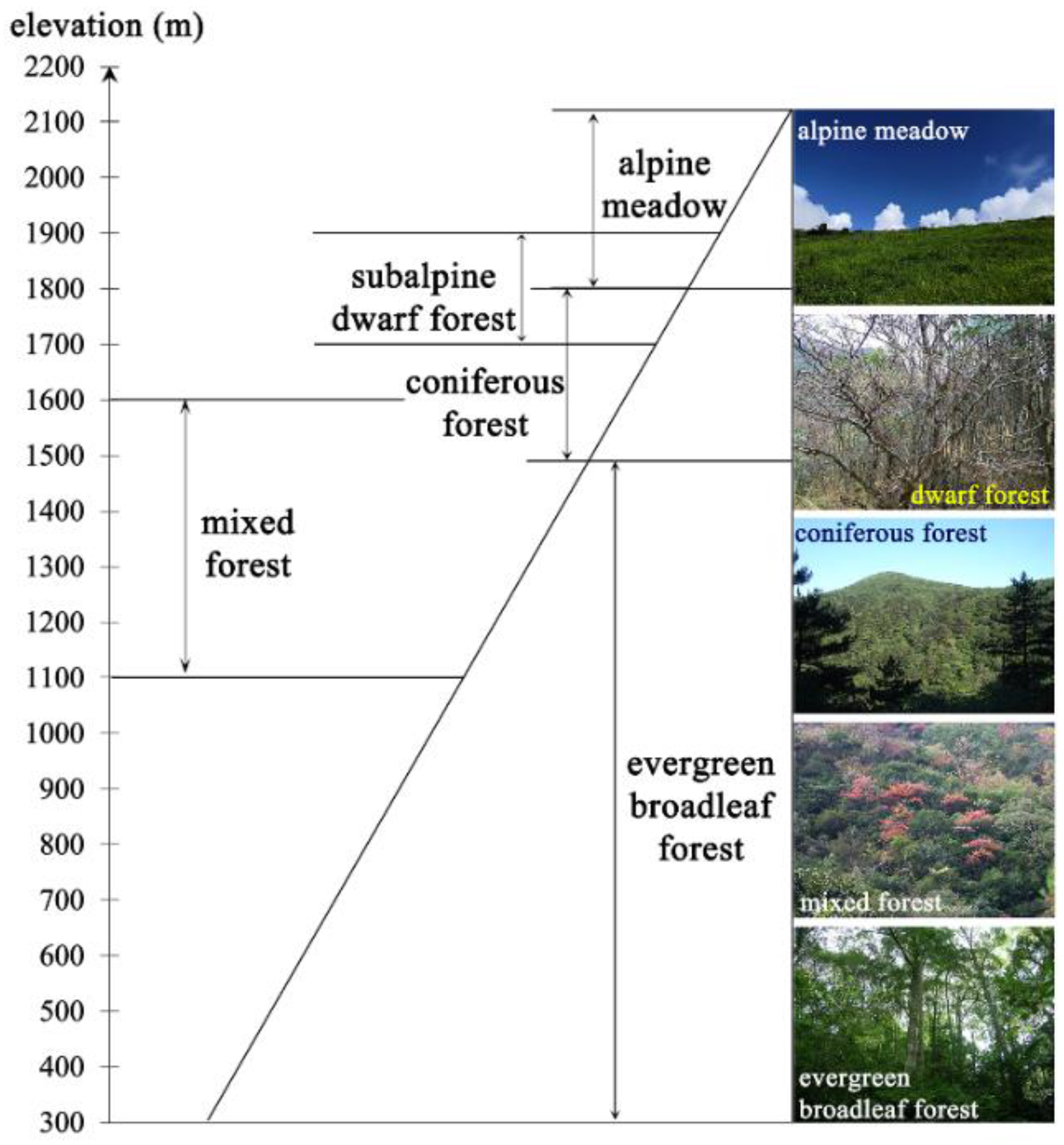
3.2.1. Automatic Tree Line Extraction from Remote-Sensing Imagery
3.2.2. Selecting Optimum Band and Season for Tree Line Extraction
3.2.3. Identification of Type of Extracted Tree Line
3.2.4. Time-Series Analysis and Segmented Linear Regression for Climate Data
3.2.5. Topographic Effects on Local Variations of Upward Tree Line Shift
4. Results
4.1. Automatic Extraction of Tree Line from Landsat Imagery
4.2. Type of Line in Tree Line Ecotone Automatically Extracted from Landsat Imagery
4.3. Climate Impact on Tree Line Upward Shift
4.4. Topographic Effects on Spatial Variations of Tree Line Response to Climate Change
5. Discussions
5.1. Advantages and Disadvantages of Automatic Tree Line Extraction from Landsat Imagery
5.2. Identifying Which Line for the Tree Line Ecotone Was Automatically Extracted from Landsat Imagery
5.3. Climate Impact on Upward Tree Line Shift
5.4. Local Variations in Tree Line Response to Climate Change
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barros, C.; Gueguen, M.; Douzet, R.; Carboni, M.; Boulangeat, I.; Zimmermann, N.E.; Munkemuller, T.; Thuiller, W. Extreme climate events counteract the effects of climate and land-use changes in Alpine tree lines. J. Appl. Ecol. 2017, 54, 39–50. [Google Scholar] [CrossRef]
- Jacob, M.; Annys, S.; Frankl, A.; De Ridder, M.; Beeckman, H.; Guyassa, E.; Nyssen, J. Tree line dynamics in the tropical African highlands—Identifying drivers and dynamics. J. Veg. Sci. 2015, 26, 9–20. [Google Scholar] [CrossRef]
- Balestrini, R.; Arese, C.; Freppaz, M.; Buffagni, A. Catchment features controlling nitrogen dynamics in running waters above the tree line (central Italian Alps). Hydrol. Earth Syst. Sci. 2013, 17, 989–1001. [Google Scholar] [CrossRef]
- Prentice, I.C.; Cramer, W.; Harrison, S.P.; Leemans, R.; Monserud, R.A.; Solomon, A.M. A global biome model based on plant physiology and dominance, soil properties and climate. J. Biogeogr. 1992, 19, 117–134. [Google Scholar] [CrossRef]
- Woodward, F.I. Climate and Plant Distribution; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Dawes, M.A.; Philipson, C.D.; Fonti, P.; Bebi, P.; Haettenschwiler, S.; Hagedorn, F.; Rixen, C. Soil warming and co2 enrichment induce biomass shifts in alpine tree line vegetation. Glob. Chang. Biol. 2015, 21, 2005–2021. [Google Scholar] [CrossRef]
- Hicks, S. The use of annual arboreal pollen deposition values for delimiting tree-lines in the landscape and exploring models of pollen dispersal. Rev. Palaeobot. Palynol. 2001, 117, 1–29. [Google Scholar] [CrossRef]
- Batllori, E.; Gutierrez, E. Regional tree line dynamics in response to global change in the pyrenees. J. Ecol. 2008, 96, 1275–1288. [Google Scholar] [CrossRef]
- Dang, H.; Zhang, K.; Zhang, Y.; Tan, S.; Jiang, M.; Zhang, Q. Tree-line dynamics in relation to climate variability in the Shennongjia Mountains, central China. Can. J. For. Res. 2009, 39, 1848–1858. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. J. Ecol. 2007, 95, 352–363. [Google Scholar] [CrossRef]
- Woods, K.D. Problems with edges: Tree lines as indicators of climate change (or not). Appl. Veg. Sci. 2014, 17, 4–5. [Google Scholar] [CrossRef]
- Aakala, T.; Hari, P.; Dengel, S.; Newberry, S.L.; Mizunuma, T.; Grace, J. A prominent stepwise advance of the tree line in north-east Finland. J. Ecol. 2014, 102, 1582–1591. [Google Scholar] [CrossRef]
- Bader, M.Y.; Ruijten, J.J.A. A topography-based model of forest cover at the alpine tree line in the tropical Andes. J. Biogeogr. 2008, 35, 711–723. [Google Scholar] [CrossRef]
- Batllori, E.; Julio Camarero, J.; Gutierrez, E. Current regeneration patterns at the tree line in the Pyrenees indicate similar recruitment processes irrespective of the past disturbance regime. J. Biogeogr. 2010, 37, 1938–1950. [Google Scholar] [CrossRef]
- Korner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Ameztegui, A.; Coll, L.; Brotons, L.; Ninot, J.M. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Glob. Ecol. Biogeogr. 2016, 25, 263–273. [Google Scholar] [CrossRef]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Im, S.T.; Dvinskaya, M.L.; Ranson, K.J. Climate-induced mountain tree-line evolution in southern Siberia. Scand. J. For. Res. 2010, 25, 446–454. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Hagedorn, F.; Knorre, A.A.; Fedotova, E.V.; Vaganov, E.A.; Naurzbaev, M.M.; Moiseev, P.A.; Rigling, A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas 2012, 41, 56–67. [Google Scholar] [CrossRef]
- Mamet, S.D.; Kershaw, G.P. Subarctic and alpine tree line dynamics during the last 400 years in north-western and central Canada. J. Biogeogr. 2012, 39, 855–868. [Google Scholar] [CrossRef]
- Mathisen, I.E.; Mikheeva, A.; Tutubalina, O.V.; Aune, S.; Hofgaard, A. Fifty years of tree line change in the Khibiny Mountains, Russia: Advantages of combined remote sensing and dendroecological approaches. Appl. Veg. Sci. 2014, 17, 6–16. [Google Scholar] [CrossRef]
- Payette, S. Contrasted dynamics of northern Labrador tree lines caused by climate change and migrational lag. Ecology 2007, 88, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Van Bogaert, R.; Haneca, K.; Hoogesteger, J.; Jonasson, C.; De Dapper, M.; Callaghan, T.V. A century of tree line changes in sub-Arctic Sweden shows local and regional variability and only a minor influence of 20th century climate warming. J. Biogeogr. 2011, 38, 907–921. [Google Scholar] [CrossRef]
- Jacob, M.; Frankl, A.; Beeckman, H.; Mesfin, G.; Hendrickx, M.; Guyassa, E.; Nyssen, J. North Ethiopian Afro-Alpine Tree Line Dynamics and Forest-Cover Change Since the Early 20th Century. Land Degrad. Dev. 2015, 26, 654–664. [Google Scholar] [CrossRef]
- Cullen, L.E.; Stewart, G.H.; Duncan, R.P.; Palmer, J.G. Disturbance and climate warming influences on New Zealand Nothofagus tree-line population dynamics. J. Ecol. 2001, 89, 1061–1071. [Google Scholar] [CrossRef]
- Harsch, M.A.; Buxton, R.; Duncan, R.P.; Hulme, P.E.; Wardle, P.; Wilmshurst, J. Causes of tree line stability: Stem growth, recruitment and mortality rates over 15 years at New Zealand Nothofagus tree lines. J. Biogeogr. 2012, 39, 2061–2071. [Google Scholar] [CrossRef]
- Liang, E.; Lu, X.; Ren, P.; Li, X.; Zhu, L.; Eckstein, D. Annual increments of juniper dwarf shrubs above the tree line on the central Tibetan Plateau: A useful climatic proxy. Ann. Bot. 2012, 109, 721–728. [Google Scholar] [CrossRef]
- Fajardo, A.; McIntire, E.J.B. Reversal of multicentury tree growth improvements and loss of synchrony at mountain tree lines point to changes in key drivers. J. Ecol. 2012, 100, 782–794. [Google Scholar] [CrossRef]
- Lara, A.; Villalba, R.; Wolodarsky-Franke, A.; Aravena, J.C.; Luckman, B.H.; Cuq, E. Spatial and temporal variation in Nothofagus pumilio growth at tree line along its latitudinal range (35 degrees 40′–55 degrees S) in the Chilean Andes. J. Biogeogr. 2005, 32, 879–893. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Lloyd, A.H. Ecological histories from Alaskan tree lines provide insight into future change. Ecology 2005, 86, 1687–1695. [Google Scholar] [CrossRef]
- IPCC. IPCC fifth assessment report. Weather 2013, 68, 310. [Google Scholar]
- Schrag, A.M.; Bunn, A.G.; Graumlich, L.J. Influence of bioclimatic variables on tree-line conifer distribution in the Greater Yellowstone Ecosystem: Implications for species of conservation concern. J. Biogeogr. 2008, 35, 698–710. [Google Scholar] [CrossRef]
- Schworer, C.; Gavin, D.G.; Walker, I.R.; Hu, F.S. Holocene tree line changes in the Canadian Cordillera are controlled by climate and topography. J. Biogeogr. 2017, 44, 1148–1159. [Google Scholar] [CrossRef]
- Trant, A.J.; Lewis, K.; Cranston, B.H.; Wheeler, J.A.; Jameson, R.G.; Jacobs, J.D.; Hermanutz, L.; Starzomski, B.M. Complex Changes in Plant Communities across a Subarctic Alpine Tree Line in Labrador, Canada. Arctic 2015, 68, 500–512. [Google Scholar] [CrossRef]
- Hertel, D.; Schoeling, D. Norway Spruce Shows Contrasting Changes in Below- Versus Above-Ground Carbon Partitioning towards the Alpine Tree line: Evidence from a Central European Case Study. Arct. Antarct. Alp. Res. 2011, 43, 46–55. [Google Scholar] [CrossRef]
- Mazepa, V.S. Stand density in the last millennium at the upper tree-line ecotone in the Polar Ural Mountains. Can. J. For. Res. Rev. Can. Rech. For. 2005, 35, 2082–2091. [Google Scholar] [CrossRef]
- Batllori, E.; Camarero, J.J.; Ninot, J.M.; Gutierrez, E. Seedling recruitment, survival and facilitation in alpine Pinus uncinata tree line ecotones. Implications and potential responses to climate warming. Glob. Ecol. Biogeogr. 2009, 18, 460–472. [Google Scholar] [CrossRef]
- Elliott, G.P. Influences of 20th-century warming at the upper tree line contingent on local-scale interactions: Evidence from a latitudinal gradient in the Rocky Mountains, USA. Glob. Ecol. Biogeogr. 2011, 20, 46–57. [Google Scholar] [CrossRef]
- Kullman, L. Tree line population monitoring of Pinus sylvestris in the Swedish Scandes, 1973-2005: Implications for tree line theory and climate change ecology. J. Ecol. 2007, 95, 41–52. [Google Scholar] [CrossRef]
- Astudillo-Sanchez, C.C.; Villanueva-Diaz, J.; Endara-Agramont, A.R.; Nava-Bernal, G.E.; Gomez-Albores, M.A. The influence of climate on Pinus hartwegii Lindl. recruitment at the alpine tree line ecotone in Monte Tlaloc, Mexico. Agrociencia 2017, 51, 105–118. [Google Scholar]
- Trant, A.J.; Hermanutz, L. Advancing towards novel tree lines? A multispecies approach to recent tree line dynamics in subarctic alpine Labrador, northern Canada. J. Biogeogr. 2014, 41, 1115–1125. [Google Scholar] [CrossRef]
- Asselin, H.; Payette, S. Origin and long-term dynamics of a subarctic tree line. Ecoscience 2006, 13, 135–142. [Google Scholar] [CrossRef]
- Berninger, F.; Hari, P.; Nikinmaa, E.; Lindholm, M.; Merilainen, J. Use of modeled photosynthesis and decomposition to describe tree growth at the northern tree line. Tree Physiol. 2004, 24, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Cullen, L.E.; Palmer, J.G.; Duncan, R.P.; Stewart, G.H. Climate change and tree-ring relationships of Nothofagus menziesii tree-line forests. Can. J. For. Res. 2001, 31, 1981–1991. [Google Scholar] [CrossRef]
- Trant, A.J.; Jameson, R.G.; Hermanutz, L. Persistence at the Tree Line: Old Trees as Opportunists. Arctic 2011, 64, 367–370. [Google Scholar] [CrossRef]
- Franke, A.K.; Braeuning, A.; Timonen, M.; Rautio, P. Growth response of Scots pines in polar-alpine tree-line to a warming climate. For. Ecol. Manag. 2017, 399, 94–107. [Google Scholar] [CrossRef]
- Gou, X.; Zhang, F.; Deng, Y.; Ettl, G.J.; Yang, M.; Gao, L.; Fang, K. Patterns and dynamics of tree-line response to climate change in the eastern Qilian Mountains, northwestern China. Dendrochronologia 2012, 30, 121–126. [Google Scholar] [CrossRef]
- Fang, K.; Gou, X.; Chen, F.; Peng, J.; D’Arrigo, R.; Wright, W.; Li, M.-H. Response of regional tree-line forests to climate change: Evidence from the northeastern Tibetan Plateau. Trees Struct. Funct. 2009, 23, 1321–1329. [Google Scholar] [CrossRef]
- Rundqvist, S.; Hedenas, H.; Sandstrom, A.; Emanuelsson, U.; Eriksson, H.; Jonasson, C.; Callaghan, T.V. Tree and Shrub Expansion Over the Past 34 Years at the Tree-Line Near Abisko, Sweden. Ambio 2011, 40, 683–692. [Google Scholar] [CrossRef]
- Bello-Rodriguez, V.; Cubas, J.; Del Arco, M.J.; Martin, J.L.; Maria Gonzalez-Mancebo, J. Elevational and structural shifts in the treeline of an oceanic island (Tenerife, Canary Islands) in the context of global warming. Int. J. Appl. Earth Obs. Geoinf. 2019, 82. [Google Scholar] [CrossRef]
- Alftine, K.J.; Malanson, G.P. Directional positive feedback and pattern at an alpine tree line. J. Veg. Sci. 2004, 15, 3–12. [Google Scholar] [CrossRef]
- Pennisi, E. Tree Line Shifts. Science 2013, 341, 484. [Google Scholar] [CrossRef] [PubMed]
- Binney, H.A.; Gething, P.W.; Nield, J.M.; Sugita, S.; Edwards, M.E. Tree line identification from pollen data: Beyond the limit? J. Biogeogr. 2011, 38, 1792–1806. [Google Scholar] [CrossRef]
- Birks, H.H.; Bjune, A.E. Can we detect a west Norwegian tree line from modern samples of plant remains and pollen? Results from the DOORMAT project. Veg. Hist. Archaeobot. 2010, 19, 325–340. [Google Scholar] [CrossRef]
- Bjune, A.E. Holocene vegetation history and tree-line changes on a north-south transect crossing major climate gradients in southern Norway—Evidence from pollen and plant macrofossils in lake sediments. Rev. Palaeobot. Palynol. 2005, 133, 249–275. [Google Scholar] [CrossRef]
- Bjune, A.E. After 8 years of annual pollen trapping across the tree line in western Norway: Are the data still anomalous? Veg. Hist. Archaeobot. 2014, 23, 299–308. [Google Scholar] [CrossRef]
- Colombaroli, D.; Henne, P.D.; Kaltenrieder, P.; Gobet, E.; Tinner, W. Species responses to fire, climate and human impact at tree line in the Alps as evidenced by palaeo-environmental records and a dynamic simulation model. J. Ecol. 2010, 98, 1346–1357. [Google Scholar] [CrossRef]
- Wilmking, M.; Harden, J.; Tape, K. Effect of tree line advance on carbon storage in NW Alaska. J. Geophys. Res. Biogeosci. 2006, 111. [Google Scholar] [CrossRef]
- Danby, R. A multiscale study of tree-line dynamics in-southwestern Yukon. Arctic 2003, 56, 427–429. [Google Scholar] [CrossRef][Green Version]
- Chen, Y.; Lu, D.; Luo, G.; Huang, J. Detection of vegetation abundance change in the alpine tree line using multitemporal Landsat Thematic Mapper imagery. Int. J. Remote Sens. 2015, 36, 4683–4701. [Google Scholar] [CrossRef]
- Virtanen, R.; Luoto, M.; Rama, T.; Mikkola, K.; Hjort, J.; Grytnes, J.-A.; Birks, H.J.B. Recent vegetation changes at the high-latitude tree line ecotone are controlled by geomorphological disturbance, productivity and diversity. Glob. Ecol. Biogeogr. 2010, 19, 810–821. [Google Scholar] [CrossRef]
- Coops, N.C.; Morsdorf, F.; Schaepman, M.E.; Zimmermann, N.E. Characterization of an alpine tree line using airborne LiDAR data and physiological modeling. Glob. Chang. Biol. 2013, 19, 3808–3821. [Google Scholar] [CrossRef]
- Cervena, L.; Kupkova, L.; Sucha, R. Field Spectroscopy for Vegetation Evaluation along the Nutrient and Elevation Gradient above the Tree Line in the Krkonose Mountains National Park. In XXIII ISPRS Congress, Commission VI; Halounova, L., Safar, V., Gong, J., Hanzl, V., Wu, H., Vyas, A., Wang, L., Musikhin, I., Tsai, F., Gruen, A., et al., Eds.; ISPRS: Prague, Czech Republic, 2016; Volume 41, pp. 211–214. [Google Scholar]
- D’Arrigo, R.D.; Kaufmann, R.K.; Davi, N.; Jacoby, G.C.; Laskowski, C.; Myneni, R.B.; Cherubini, P. Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory, Canada. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, M.; Adams, J.; Wang, X. Can Landsat imagery detect tree line dynamics? Int. J. Remote Sens. 2009, 30, 1327–1340. [Google Scholar] [CrossRef]
- Smith, E.K.; Resler, L.M.; Vance, E.A.; Carstensen, L.W., Jr.; Kolivras, K.N. Blister Rust Incidence in Tree line Whitebark Pine, Glacier National Park, USA: Environmental and Topographic Influences. Arct. Antarct. Alp. Res. 2011, 43, 107–117. [Google Scholar] [CrossRef]
- Carlson, B.Z.; Georges, D.; Rabatel, A.; Randin, C.F.; Renaud, J.; Delestrade, A.; Zimmermann, N.E.; Choler, P.; Thuiller, W. Accounting for tree line shift, glacier retreat and primary succession in mountain plant distribution models. Divers. Distrib. 2014, 20, 1379–1391. [Google Scholar] [CrossRef]
- Rosen, P.; Persson, P. Fourier-transform infrared spectroscopy (FTIRS), a new method to infer past changes in tree-line position and TOC using lake sediment. J. Paleolimnol. 2006, 35, 913–923. [Google Scholar] [CrossRef]
- Paulsen, J.; Korner, C. GIS-analysis of tree-line elevation in the Swiss Alps suggests no exposure effect. J. Veg. Sci. 2001, 12, 817–824. [Google Scholar] [CrossRef]
- Groen, T.A.; Fanta, H.G.; Hinkov, G.; Velichkov, I.; Van Duren, I.; Zlatanov, T. Tree Line Change Detection Using Historical Hexagon Mapping Camera Imagery and Google Earth Data. Gisci. Remote Sens. 2012, 49, 933–943. [Google Scholar] [CrossRef]
- Zong, S.; Wu, Z.; Xu, J.; Li, M.; Gao, X.; He, H.; Du, H.; Wang, L. Current and Potential Tree Locations in Tree Line Ecotone of Changbai Mountains, Northeast China: The Controlling Effects of Topography. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Wallentin, G.; Tappeiner, U.; Strobl, J.; Tasser, E. Understanding alpine tree line dynamics: An individual-based model. Ecol. Model. 2008, 218, 235–246. [Google Scholar] [CrossRef]
- Luo, G.; Dai, L. Detection of alpine tree line change with high spatial resolution remotely sensed data. J. Appl. Remote Sens. 2013, 7. [Google Scholar] [CrossRef]
- Liu, S.; Su, H.; Cao, G.; Wang, S.; Guan, Q. Learning from data: A post classification method for annual land cover analysis in urban areas. ISPRS J. Photogramm. Remote Sens. 2019, 154, 202–215. [Google Scholar] [CrossRef]
- Claverie, M.; Vermote, E.F.; Franch, B.; Masek, J.G. Evaluation of the Landsat-5 TM and Landsat-7 ETM + surface reflectance products. Remote Sens. Environ. 2015, 169, 390–403. [Google Scholar] [CrossRef]
- Wang, L.; Shi, C.; Diao, C.; Ji, W.; Yin, D. A survey of methods incorporating spatial information in image classification and spectral unmixing. Int. J. Remote Sens. 2016, 37, 3870–3910. [Google Scholar] [CrossRef]
- Holden, Z.A.; Evans, J.S. Using fuzzy C-means and local autocorrelation to cluster satellite-inferred burn severity classes. Int. J. Wildland Fire 2010, 19, 853–860. [Google Scholar] [CrossRef]
- Kowe, P.; Mutanga, O.; Odindi, J.; Dube, T. Exploring the spatial patterns of vegetation fragmentation using local spatial autocorrelation indices. J. Appl. Remote Sens. 2019, 13. [Google Scholar] [CrossRef]
- Gong, W.; Fang, S.; Yang, G.; Ge, M. Using a Hidden Markov Model for Improving the Spatial-Temporal Consistency of Time Series Land Cover Classification. ISPRS Int. J. Geo Inf. 2017, 6, 292. [Google Scholar] [CrossRef]
- Guo, L.; Xue, P.; Li, M.; Shao, X. Seed bank and regeneration dynamics of Emmenopterys henryi population on the western side of Wuyi Mountain, South China. J. For. Res. 2017, 28, 943–952. [Google Scholar] [CrossRef]
- You, W.; Lin, L.; Wu, L.; Ji, Z.; Yu, J.; Zhu, J.; Fan, Y.; He, D. Geographical information system-based forest fire risk assessment integrating national forest inventory data and analysis of its spatiotemporal variability. Ecol. Indic. 2017, 77, 176–184. [Google Scholar] [CrossRef]
- He, S.; Gallagher, L.; Su, Y.; Wang, L.; Cheng, H. Identification and assessment of ecosystem services for protected area planning: A case in rural communities of Wuyishan national park pilot. Ecosyst. Serv. 2018, 31, 169–180. [Google Scholar] [CrossRef]
- Ye, H.; Li, G.; Yuan, X.; Zheng, M. Fractionation and Bioavailability of Trace Elements in Wuyi Rock Tea Garden Soil. Pol. J. Environ. Stud. 2018, 27, 421–430. [Google Scholar] [CrossRef]
- Huang, S.; Ye, G.; Lin, J.; Chen, K.; Xu, X.; Ruan, H.; Tan, F.; Chen, H.Y.H. Autotrophic and heterotrophic soil respiration responds asymmetrically to drought in a subtropical forest in the Southeast China. Soil Biol. Biochem. 2018, 123, 242–249. [Google Scholar] [CrossRef]
- Li, M.; Zheng, Y.; Fan, R.; Zhong, Q.; Cheng, D. Scaling relationships of twig biomass allocation in Pinus hwangshanensis along an altitudinal gradient. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Hu, H.-W.; Wang, J.; He, J.-Z.; Zheng, C.; Wan, X.; Huang, Z. Niche separation of comammox Nitrospira and canonical ammonia oxidizers in an acidic subtropical forest soil under long-term nitrogen deposition. Soil Biol. Biochem. 2018, 126, 114–122. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, X.; Luo, Y.; Xu, Z.; Xu, L.; Ruan, H.; Xu, X. Consistent temperature sensitivity of labile soil organic carbon mineralization along an elevation gradient in the Wuyi Mountains, China. Appl. Soil Ecol. 2017, 117, 32–37. [Google Scholar] [CrossRef]
- Sun, J.; Fan, R.; Niklas, K.J.; Zhong, Q.; Yang, F.; Li, M.; Chen, X.; Sun, M.; Cheng, D. “Diminishing returns” in the scaling of leaf area vs. dry mass in Wuyi Mountain bamboos, Southeast China. Am. J. Bot. 2017, 104, 993–998. [Google Scholar] [CrossRef]
- Xu, D.D.; Guo, X.L.; Li, Z.Q.; Yang, X.H.; Yin, H. Measuring the dead component of mixed grassland with Landsat imagery. Remote Sens Environ. 2014, 142, 33–43. [Google Scholar] [CrossRef]
- Syariz, M.A.; Lin, B.-Y.; Denaro, L.G.; Jaelani, L.M.; Math Van, N.; Lin, C.-H. Spectral-consistent relative radiometric normalization for multitemporal Landsat 8 imagery. ISPRS J. Photogramm. Remote Sens. 2019, 147, 56–64. [Google Scholar] [CrossRef]
- Fan, X.; Liu, Y. A global study of NDVI difference among moderate-resolution satellite sensors. ISPRS J. Photogramm. Remote Sens. 2016, 121, 177–191. [Google Scholar] [CrossRef]
- Rousi, M.; Possen, B.J.M.H.; Ruotsalainen, S.; Silfver, T.; Mikola, J. Temperature and soil fertility as regulators of tree line Scots pine growth and survival-implications for the acclimation capacity of northern populations. Glob. Chang. Biol. 2018, 24, E545–E559. [Google Scholar] [CrossRef] [PubMed]
- Peringer, A.; Rosenthal, G. Establishment patterns in a secondary tree line ecotone. Ecol. Model. 2011, 222, 3120–3131. [Google Scholar] [CrossRef]
- Wagemann, J.; Ties, B.; Rollenbeck, R.; Peters, T.; Bendix, J. Regionalization of wind-speed data to analyse tree-line wind conditions in the eastern andes of southern Ecuador. Erdkunde 2015, 69, 3–19. [Google Scholar] [CrossRef]
- Wang, L.; Godbold, D.L. Soil N mineralization profiles of co-existing woody vegetation islands at the alpine tree line. Eur. J. For. Res. 2017, 136, 881–892. [Google Scholar] [CrossRef]
- McIntire, E.J.B.; Piper, F.I.; Fajardo, A. Wind exposure and light exposure, more than elevation-related temperature, limit tree line seedling abundance on three continents. J. Ecol. 2016, 104, 1379–1390. [Google Scholar] [CrossRef]
- Tingstad, L.; Olsen, S.L.; Klanderud, K.; Vandvik, V.; Ohlson, M. Temperature, precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone. Oecologia 2015, 179, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Maher, E.L.; Germino, M.J.; Hasselquist, N.J. Interactive effects of tree and herb cover on survivorship, physiology, and microclimate of conifer seedlings at the alpine tree-line ecotone. Can. J. For. Res. Rev. Can. Rech. For. 2005, 35, 567–574. [Google Scholar] [CrossRef]
- Gavilan, R.G.; Callaway, R.M. Effects of foundation species above and below tree line. Plant. Biosyst. 2017, 151, 665–672. [Google Scholar] [CrossRef]
- Gervais, B.R.; MacDonald, G.M. Tree-ring and summer-temperature response to volcanic aerosol forcing at the northern tree-line, Kola Peninsula, Russia. Holocene 2001, 11, 499–505. [Google Scholar] [CrossRef]
- Brown, C.D. Tree-line Dynamics Adding Fire to Climate Change Prediction. Arctic 2010, 63, 488–492. [Google Scholar] [CrossRef]
- Cierjacks, A.; Iglesias, J.E.; Wesche, K.; Hensen, I. Impact of sowing, canopy cover and litter on seedling dynamics of two Polylepis species at upper tree lines in central Ecuador. J. Trop. Ecol. 2007, 23, 309–318. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Wilson, A.E.; Fastie, C.L.; Landis, R.M. Population dynamics of black spruce and white spruce near the arctic tree line in the southern Brooks Range, Alaska. Can. J. For. Res. 2005, 35, 2073–2081. [Google Scholar] [CrossRef]
- Cairns, D.M.; Moen, J. Herbivory influences tree lines. J. Ecol. 2004, 92, 1019–1024. [Google Scholar] [CrossRef]
- Ducic, V.; Milovanovic, B.; Durdic, S. Identification of recent factors that affect the formation of the upper tree line in eastern Serbia. Arch. Biol. Sci. 2011, 63, 825–830. [Google Scholar] [CrossRef]
- Grace, J.; Berninger, F.; Nagy, L. Impacts of climate change on the tree line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef]
- Sarmiento, F.O.; Frolich, L.M. Andean cloud forest tree lines—Naturalness, agriculture and the human dimension. Mt. Res. Dev. 2002, 22, 278–287. [Google Scholar] [CrossRef]
- Milligan, A.L.; Putwain, P.D.; Cox, E.S.; Ghorbani, J.; Le Duc, M.G.; Marrs, R.H. Developing an integrated land management strategy for the restoration of moorland vegetation on Molinia caerulea-dominated vegetation for conservation purposes in upland Britain. Biol. Conserv. 2004, 119, 371–385. [Google Scholar] [CrossRef]
- Yamazaki, J.Y.; Tsuchiya, S.; Nagano, S.; Maruta, E. Photoprotective mechanisms against winter stresses in the needles of Abies mariesii grown at the tree line on Mt. Norikura in Central Japan. Photosynthetica 2007, 45, 547–554. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | January | February | March | April | May | June | July | August | September | October | November | December |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1987 | 1 TM | |||||||||||
| 1988 | 1 TM | 2 TM | ||||||||||
| 1989 | 1 TM | 1 TM | ||||||||||
| 1990 | 1 TM | |||||||||||
| 1991 | 2 TM | |||||||||||
| 1992 | 1 TM | 1 TM | 1 TM | |||||||||
| 1993 | 1 TM | |||||||||||
| 1994 | 1 TM | 1 TM | ||||||||||
| 1995 | 1 TM | |||||||||||
| 1996 | 1 TM | 1 TM | 1 TM | 1 TM | ||||||||
| 1997 | 1 TM | |||||||||||
| 1998 | 1 TM | 1 TM | 1 TM | 1 TM | ||||||||
| 1999 | 1 TM | 2 TM | ||||||||||
| 2000 | 1 ETM+ | 1 TM | 1 TM 1 ETM+ | 1 TM | 1 ETM+ | |||||||
| 2001 | 1 TM | 1 TM | 1 ETM+ | 1 ETM+ | ||||||||
| 2002 | 1 TM | 1 TM | 1 ETM+ | |||||||||
| 2003 | 2 ETM+ | 1 TM | 1 ETM+ | 1 TM | 1 TM, 1 ETM+, 1 ETM+ | |||||||
| 2004 | 1 ETM+ | 1 TM | 1 ETM+ | 1 ETM+ | 2 TM | 1 TM | 1 TM, 1 ETM+ | |||||
| 2005 | 1 ETM+ | 1 ETM+ | 1 ETM+ | |||||||||
| 2006 | 1 TM, 1 ETM+ | 1 ETM+ | ||||||||||
| 2007 | 1 TM, 1 ETM+ | 1 TM | 2 ETM+ | |||||||||
| 2008 | 1 ETM+ | 1 TM, 1 ETM+ | ||||||||||
| 2009 | 1 ETM+ | 1 ETM+ | 1 ETM+ | 1 TM | 1 TM | |||||||
| 2010 | 1 TM | 1 TM | 1 TM | 1 TM | ||||||||
| 2011 | 1 TM | 1 ETM+ | ||||||||||
| 2012 | 1 ETM+ | 1 ETM+ | 1 ETM+ | 1 ETM+ | ||||||||
| 2013 | 1 OLI | 1 OLI | ||||||||||
| 2014 | 1 OLI | 1 OLI | 1 OLI | |||||||||
| 2015 | 1 OLI | 1 OLI | 1 OLI | |||||||||
| 2016 | 1 OLI | 1 OLI | ||||||||||
| 2017 | 1 OLI | 1 OLI | ||||||||||
| 2018 | 1 OLI | 1 OLI |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Geng, Q.; Jin, C.; Xu, Z.; Xu, X. Tree Line Identification and Dynamics under Climate Change in Wuyishan National Park Based on Landsat Images. Remote Sens. 2020, 12, 2890. https://doi.org/10.3390/rs12182890
Xu D, Geng Q, Jin C, Xu Z, Xu X. Tree Line Identification and Dynamics under Climate Change in Wuyishan National Park Based on Landsat Images. Remote Sensing. 2020; 12(18):2890. https://doi.org/10.3390/rs12182890
Chicago/Turabian StyleXu, Dandan, Qinghong Geng, Changshan Jin, Zikun Xu, and Xia Xu. 2020. "Tree Line Identification and Dynamics under Climate Change in Wuyishan National Park Based on Landsat Images" Remote Sensing 12, no. 18: 2890. https://doi.org/10.3390/rs12182890
APA StyleXu, D., Geng, Q., Jin, C., Xu, Z., & Xu, X. (2020). Tree Line Identification and Dynamics under Climate Change in Wuyishan National Park Based on Landsat Images. Remote Sensing, 12(18), 2890. https://doi.org/10.3390/rs12182890