Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems

 ,
,  , ,
, ,  and
and
Abstract
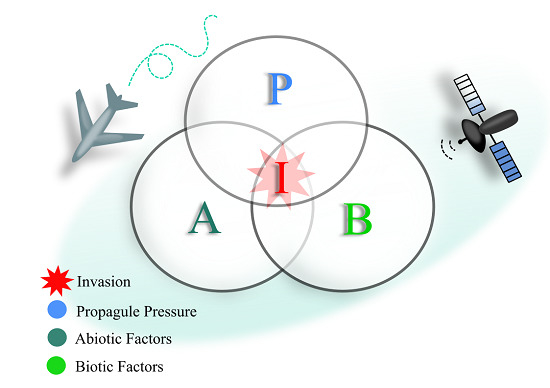
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Invasive Species Data
2.3. Remotely Sensed Data
2.4. Mapping of Propagule Pressure, Abiotic Conditions and Biotic Environment
2.5. Species Distribution Modelling
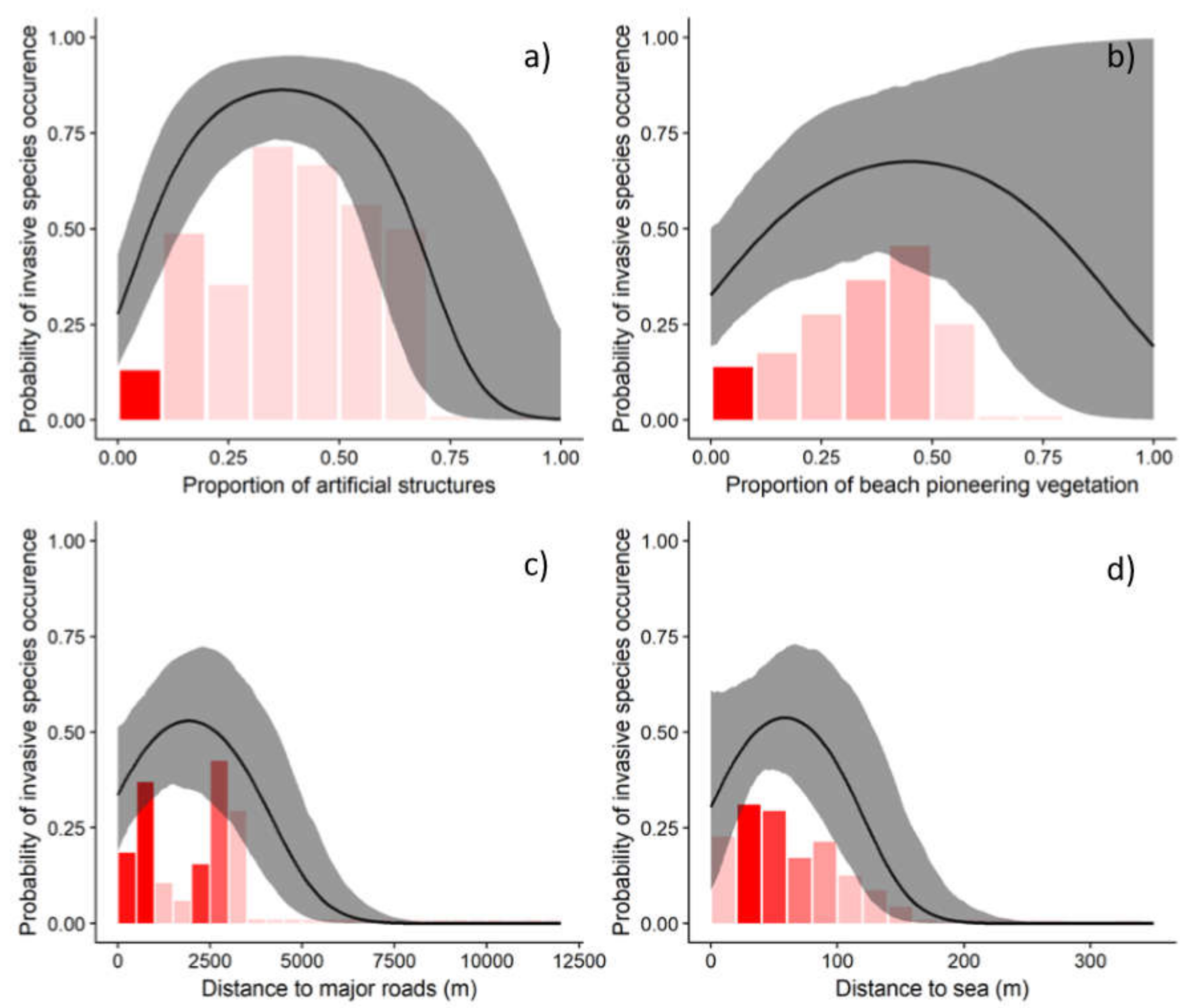
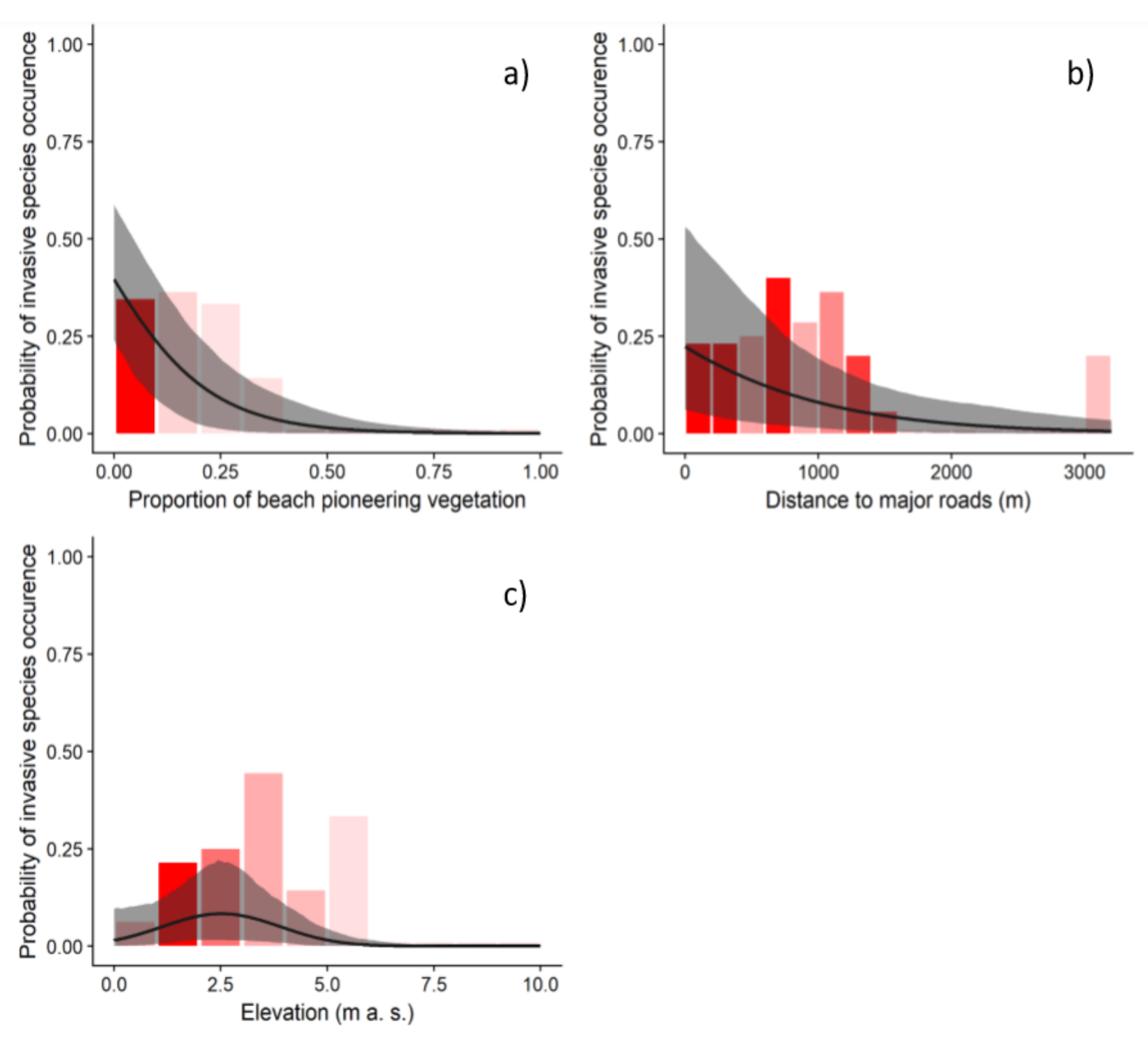
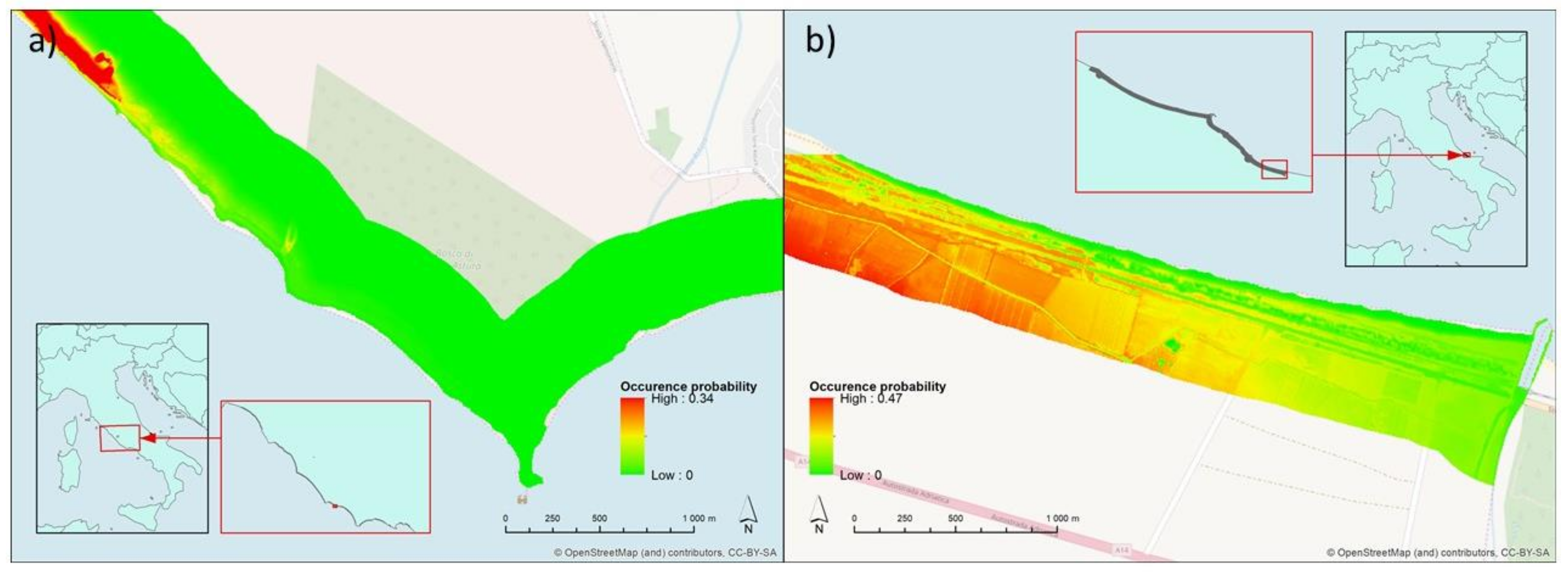
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef]
- Russell, J.C.; Blackburn, T.M. The rise of invasive species denialism. Trends Ecol. Evol. 2017, 32, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D.; Martin, J.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.; Fitter, A. The varying success of invaders. Ecology 1996, 77, 1661–1666. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef]
- Enders, M.; Hütt, M.T.; Jeschke, J.M. Drawing a map of invasion biology based on a network of hypotheses. Ecosphere 2018, 9, e02146. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef]
- Gurevitch, J.; Fox, G.A.; Wardle, G.M.; Taub, D. Emergent insights from the synthesis of conceptual frameworks for biological invasions. Ecol. Lett. 2011, 14, 407–418. [Google Scholar] [CrossRef]
- Bazzichetto, M.; Malavasi, M.; Barták, V.; Acosta, A.T.R.; Moudrý, V.; Carranza, M.L. Modeling plant invasion on Mediterranean coastal landscapes: An integrative approach using remotely sensed data. Landsc. Urban Plan. 2018, 171, 98–106. [Google Scholar] [CrossRef]
- Bellard, C.; Leroy, B.; Thuiller, W.; Rysman, J.F.; Courchamp, F. Major drivers of invasion risks throughout the world. Ecosphere 2016, 7, e01241. [Google Scholar] [CrossRef]
- Leishman, M.R.; Gallagher, R.V. Will there be a shift to alien-dominated vegetation assemblages under climate change? Divers. Distrib. 2015, 21, 848–852. [Google Scholar] [CrossRef]
- Malavasi, M.; Acosta, A.T.R.; Carranza, M.L.; Bartolozzi, L.; Basset, A.; Bassignana, M.; Campanaro, A.; Canullo, R.; Carruggio, F.; Cavallaro, V.; et al. Plant invasions in Italy: An integrative approach using the European LifeWatch infrastructure database. Ecol. Ind. 2018, 91, 182–188. [Google Scholar]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 8, 306–314. [Google Scholar] [CrossRef]
- Vaz, A.S.; Alcaraz-Segura, D.; Campos, J.C.; Vicente, J.R.; Honrado, J.P. Managing plant invasions through the lens of remote sensing: A review of progress and the way forward. Sci. Total Environ. 2018, 642, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.E.; Ustin, S.L. Habitat suitability modelling of an invasive plant with advanced remote sensing data. Divers. Distrib. 2009, 15, 627–640. [Google Scholar] [CrossRef]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.N.; Schmidtlein, S.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef]
- Cord, A.F.; Meentemeyer, R.K.; Leitão, P.J.; Václavík, T. Modelling species distributions with remote sensing data: Bridging disciplinary perspectives. J. Biogeogr. 2013, 40, 2226–2227. [Google Scholar] [CrossRef]
- Bruno, J.F.; Kennedy, C.W.; Rand, T.A.; Grant, M.B. Landscape- scale patterns of biological invasions in shoreline plant communities. Oikos 2004, 107, 531–540. [Google Scholar] [CrossRef]
- Chytrý, M.; Maskell, L.C.; Pino, J.; Pyšek, P.; Vilà, M.; Font, X.; Smart, S.M. Habitat invasions by alien plants: A quantitative comparison among Mediterranean, subcontinental and oceanic regions of Europe. J. Appl. Ecol. 2008, 45, 448–458. [Google Scholar] [CrossRef]
- Defeo, O.; McLachlan, A.; Schoeman, D.S.; Schlacher, T.A.; Dugan, J.; Jones, A.; Lastra, M.; Scapini, F. Threats to sandy beach ecosystems: A review. Estuar. Coast. Shelf Sci. 2009, 81, 1–12. [Google Scholar] [CrossRef]
- Drius, M.; Carranza, M.L.; Stanisci, A.; Jones, L. The role of Italian coastal dunes as carbon sinks and diversity sources. A multi-service perspective. Appl. Geogr. 2016, 75, 127–136. [Google Scholar] [CrossRef]
- Carranza, M.L.; Acosta, A.T.R.; Stanisci, A.; Pirone, G.; Ciaschetti, G. Ecosystem classification for EU habitat distribution assessment in sandy coastal environments: An application in central Italy. Environ. Monit. Assess. 2008, 140, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.; Blasi, C.; Carranza, M.L.; Ricotta, C.; Stanisci, A. Quantifying ecological mosaic connectivity and with a new topoecological index. Phytocoenologia 2003, 33, 623–631. [Google Scholar] [CrossRef]
- Bazzichetto, M.; Malavasi, M.; Acosta, A.T.R.; Carranza, M.L. How does dune morphology shape coastal EC habitats occurrence? A remote sensing approach using airborne LiDAR on the Mediterranean coast. Ecol. Indic. 2016, 71, 618–626. [Google Scholar] [CrossRef]
- Carranza, M.L.; Drius, M.; Malavasi, M.; Frate, L.; Stanisci, A.; Acosta, A.T.R. Assessing land take and its effects on dune carbon pools. An insight into the Mediterranean coastline. Ecol. Indic. 2018, 85, 951–955. [Google Scholar] [CrossRef]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. The impact of human pressure on landscape patterns and plant species richness in Mediterranean coastal dunes. Plant Biosyst. 2016, 150, 73–82. [Google Scholar] [CrossRef]
- Sperandii, M.G.; Prisco, I.; Stanisci, A.; Acosta, A.T.R. RanVegDunes—A random plot database of Italian coastal dunes. Phytocoenologia 2017, 47, 231–232. [Google Scholar] [CrossRef]
- Acosta, A.; Carranza, M.L.; Ciaschetti, G.; Conti, F.L.; Di Martino, L.; D’orazio, G.; Frattaroli, A.R.; Izzi, C.F.; Pirone, G.; Stanisci, A. Alien species growing in costal dunes of Central Italy. Webbia 2007, 6, 77–84. [Google Scholar] [CrossRef]
- Celesti-Grapow, L.; Alessandrini, A.; Arrigoni, P.V.; Banfi, E.; Bernardo, L.; Bovio, M.; Brundu, G.; Cagiotti, M.R.; Camarda, I.; et al. Inventory of the nonnative flora of Italy. Plant Biosyst. 2009, 143, 386–430. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardsond, D.V.; Rejmànekm, M.; Webster, G.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Toward better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Santoro, R.; Jucker, T.; Carranza, M.; Acosta, A. Assessing the effects of Carpobrotus invasion on coastal dune soils. Does the nature of the invaded habitat matter? Community Ecol. 2011, 12, 234–240. [Google Scholar] [CrossRef]
- Santoro, R.; Carboni, M.; Carranza, M.L.; Acosta, A.T.R. Focal species diversity patterns can provide diagnostic information on plant invasions. J. Nat. Conserv. 2012, 20, 85–91. [Google Scholar] [CrossRef]
- Mihulka, S.; Pysek, P. Invasion history of Oenothera congeners in Europe: A comparative study of spreading rates in the last 200 years. J. Biogeogr. 2001, 28, 597–609. [Google Scholar] [CrossRef]
- Weaver, S.E. The biology of Canadian weeds. 115. Conyza canadensis. Can. J. Plant Sci. 2001, 81, 867–875. [Google Scholar] [CrossRef]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. What has happened to coastal dunes in the last half century? A multitemporal coastal landscape analysis in Central Italy. Landsc. Urban Plan. 2013, 119, 54–63. [Google Scholar] [CrossRef]
- Malavasi, M.; Conti, L.; Carboni, M.; Cutini, M.; Acosta, A.T.R. Multifaceted analysis of patch-level plant diversity in response to landscape spatial pattern and history on Mediterranean dunes. Ecosystems 2016, 19, 850–864. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Congalton, R.; Green, K. Assessing the Accuracy of Remotely Sensed Data: Principles and Practices; Lewis Publisher: New York, NY, USA, 2008. [Google Scholar]
- Carboni, M.; Thuiller, W.; Izzi, F.; Acosta, A.T.R. Disentangling the relative effects of environmental versus human factors on the abundance of native and alien plant species in Mediterranean sandy shores. Divers. Distrib. 2010, 16, 537–546. [Google Scholar] [CrossRef]
- Malavasi, M.; Bartak, V.; Carranza, M.L.; Simova, P.; Acosta, A.T.R. Landscape pattern and plant biodiversity in Mediterranean coastal dune ecosystems: Do habitat loss and fragmentation really matter? J. Biogeogr. 2018, 45, 1367–1377. [Google Scholar] [CrossRef]
- Bazzichetto, M.; Malavasi, M.; Barták, V.; Acosta, A.T.R.; Rocchini, D.; Carranza, M.L. Plant invasion risk: A quest for invasive species distribution modelling in managing protected areas. Ecol. Indic. 2018, 95, 311–319. [Google Scholar] [CrossRef]
- Carranza, M.L.; Ricotta, C.; Carboni, M.; Acosta, A.T.R. Habitat selection by invasive alien plants: A bootstrap approach. Preslia 2011, 83, 529–536. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Zhang, D. A Coefficient of Determination for Generalized Linear Models. Am. Stat. 2017, 71, 310–316. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E. Spatio-temporal dynamics of plant invasions: Linking pattern to process. Ecoscience 2005, 12, 302–315. [Google Scholar] [CrossRef]
- Chytrý, M.; Pyšek, P.; Wild, J.; Pino, J.; Maskell, L.C.; Vilà, M. European map of alien plant invasions based on the quantitative assessment across habitats. Divers. Distrib. 2009, 15, 98–107. [Google Scholar] [CrossRef]
- Alpert, P. The advantages and disadvantages of being introduced. Biol. Invasions 2006, 8, 1523–1534. [Google Scholar] [CrossRef]
- Carranza, M.L.; Carboni, M.; Feola, S.; Acosta, A.T.R. Landscape-scale patterns of alien plant species on coastal dunes. The case of iceplant in central Italy. Appl. Veg. Sci. 2010, 13, 135–145. [Google Scholar] [CrossRef]
- EEC. Council Directive 1143/2014/EC of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species. Off. J. L 2014, 317/35, 35–55. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor (Abbreviation; Units) | Data Source | Use in the Full Model | |
|---|---|---|---|
| Lazio | Molise | ||
| Propagule pressure (P): | |||
| Proportion of artificial structures (ART; %) | 2008 color orthophotos | Yes | Yes |
| Distance to major roads (dist_roads; m) | OpenStreetMap | Yes | Yes |
| Presence of agricultural land (AGR-F; True/False) | 2008 color orthophotos | Yes | Yes |
| Abiotic conditions (A): | |||
| Elevation (elev; m a.s.l.) | LiDAR | Yes | Yes |
| Terrain slope (slope; degree) | LiDAR | Yes | Yes |
| Distance to sea (dist_sea; m) | 2008 color orthophotos | Yes | Yes |
| Biotic environment (B): | |||
| Proportion of beach pioneer vegetation (BPV; %) | 2008 color orthophotos | Yes | Yes |
| Presence of semi-natural vegetation (SN-F; True/False) | 2008 color orthophotos | Yes | Yes |
| Proportion of herbaceous dune vegetation (HDV; %) | 2008 color orthophotos | Yes | Yes |
| Proportion of forests (AFF; %) | 2008 color orthophotos | No | No |
| Presence of forests (AFF-F; True/False) | 2008 color orthophotos | Yes | No |
| Presence of wetlands (WET-F; True/False) | 2008 color orthophotos | Yes | Yes |
| Proportion of woody dune vegetation (WDV; %) | 2008 color orthophotos | No | No |
| Presence of woody dune vegetation (WDV-F; True/False) | 2008 color orthophotos | No | Yes |
| Species richness (ric; number of native species recorded in the vegetation plots) | Vegetation plots | Yes | Yes |
| Model | Formula | Res. Deviance | Res. DF | AIC | R2 | Disp. |
|---|---|---|---|---|---|---|
| Lazio: | ||||||
| Full | IASocc ~ AFF−-F + AGR-F + ART2 + BPV2 + HDV2 + SN-F + WET-F + dist_roads2 + dist_sea2 + elev2 + slope2 + ric2 | 354.5 | 484 | 396.5 | 0.339 | 0.846 |
| Most parsim. | IASocc ~ ART2 + BPV2 + WET-F + dist_roads2 + dist_sea2 + elev2 + ric2 | 360.2 | 491 | 388.2 | 0.331 | 0.875 |
| Molise: | ||||||
| Full | IASocc ~ AGR-F + ART + BPV + HDV2 + SN-F + WDV-F + WET-F + dist_roads + dist_sea2 + elev2 + slope + ric2 | 107.8 | 146 | 141.8 | 0.343 | 0.641 |
| Most parsim. | IASocc ~ BPV + dist_roads + dist_sea2 + elev2 + ric2 | 114.7 | 154 | 132.7 | 0.301 | 0.675 |
| Domain | Predictor | Lazio | Molise | ||||
|---|---|---|---|---|---|---|---|
| LR Statistic | DF | p-Value | LR Statistic | DF | p-Value | ||
| P | ART2 | 66.888 | 2 | 3 × 10−15 *** | - | - | - |
| dist_roads | - | - | - | 9.827 | 1 | 0.0017 ** | |
| dist_roads2 | 18.773 | 2 | 8 × 10−5 *** | - | - | - | |
| A | dist_sea2 | 17.156 | 2 | 0.0002 *** | - | - | - |
| elev2 | 5.630 | 2 | 0.0600 | 9.435 | 2 | 0.0089 ** | |
| B | BPV | - | - | - | 31.219 | 1 | 2 × 10−8 *** |
| BPV2 | 8.159 | 2 | 0.0169 *** | - | - | - | |
| WET−F | 2.324 | 1 | 0.1274 | 2.389 | 1 | 0.1222 | |
| SN−F | - | - | - | 2.523 | 1 | 0.1122 | |
| ric2 | 4.298 | 2 | 0.1170 | 5.792 | 2 | 0.0553 | |
| Domain | Lazio | Molise | ||
|---|---|---|---|---|
| Predictors | ΔDev. | Predictors | ΔDev. | |
| Propagule pressure (P) | ART2, dist_roads2 | 99.45 | dist_roads | 9.83 |
| Abiotic conditions (A) | dist_sea2, elev2 | 23.16 | elev2 | 9.44 |
| Biotic environment (B) | BPV2, WET−F, ric2 | 14.14 | BPV, WET−F,SN−F, ric2 | 41.10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malavasi, M.; Barták, V.; Jucker, T.; Acosta, A.T.R.; Carranza, M.L.; Bazzichetto, M. Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems. Remote Sens. 2019, 11, 275. https://doi.org/10.3390/rs11030275
Malavasi M, Barták V, Jucker T, Acosta ATR, Carranza ML, Bazzichetto M. Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems. Remote Sensing. 2019; 11(3):275. https://doi.org/10.3390/rs11030275
Chicago/Turabian StyleMalavasi, Marco, Vojtěch Barták, Tommaso Jucker, Alicia Teresa Rosario Acosta, Maria Laura Carranza, and Manuele Bazzichetto. 2019. "Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems" Remote Sensing 11, no. 3: 275. https://doi.org/10.3390/rs11030275
APA StyleMalavasi, M., Barták, V., Jucker, T., Acosta, A. T. R., Carranza, M. L., & Bazzichetto, M. (2019). Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems. Remote Sensing, 11(3), 275. https://doi.org/10.3390/rs11030275