Prediction of Competition Indices in a Norway Spruce and Silver Fir-Dominated Forest Using Lidar Data


 , ,
, ,  , and
, and
Abstract
1. Introduction
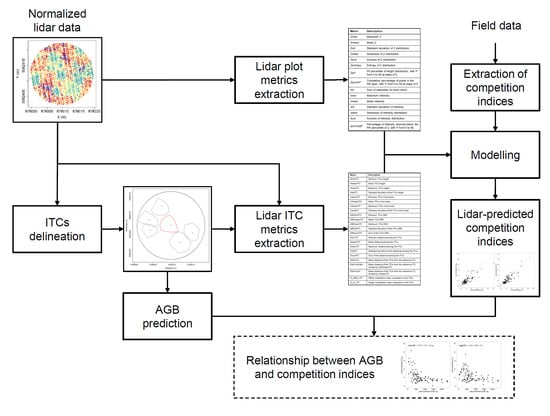
2. Materials and Methods
2.1. Data Set Description
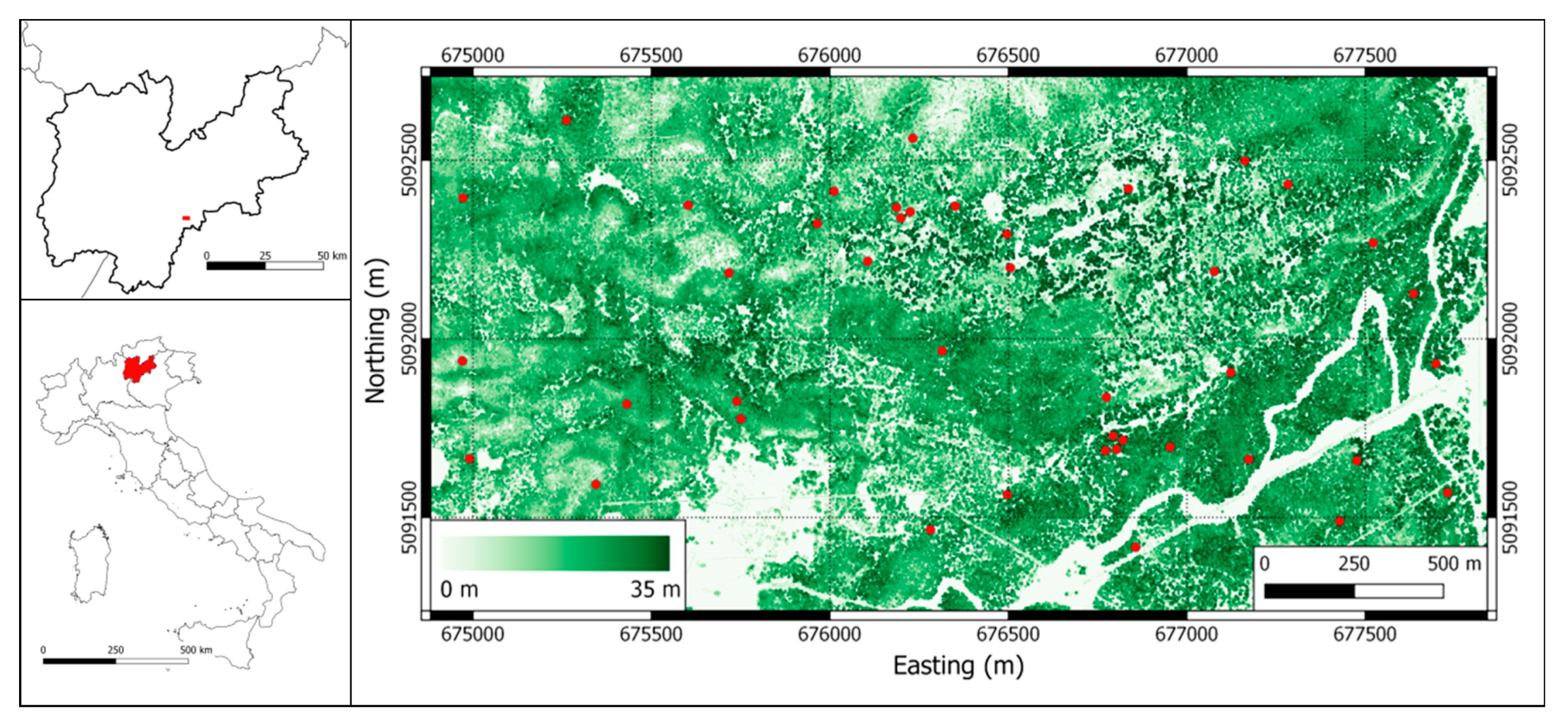
2.1.1. Study Area
2.1.2. Lidar Data
2.1.3. Field Data
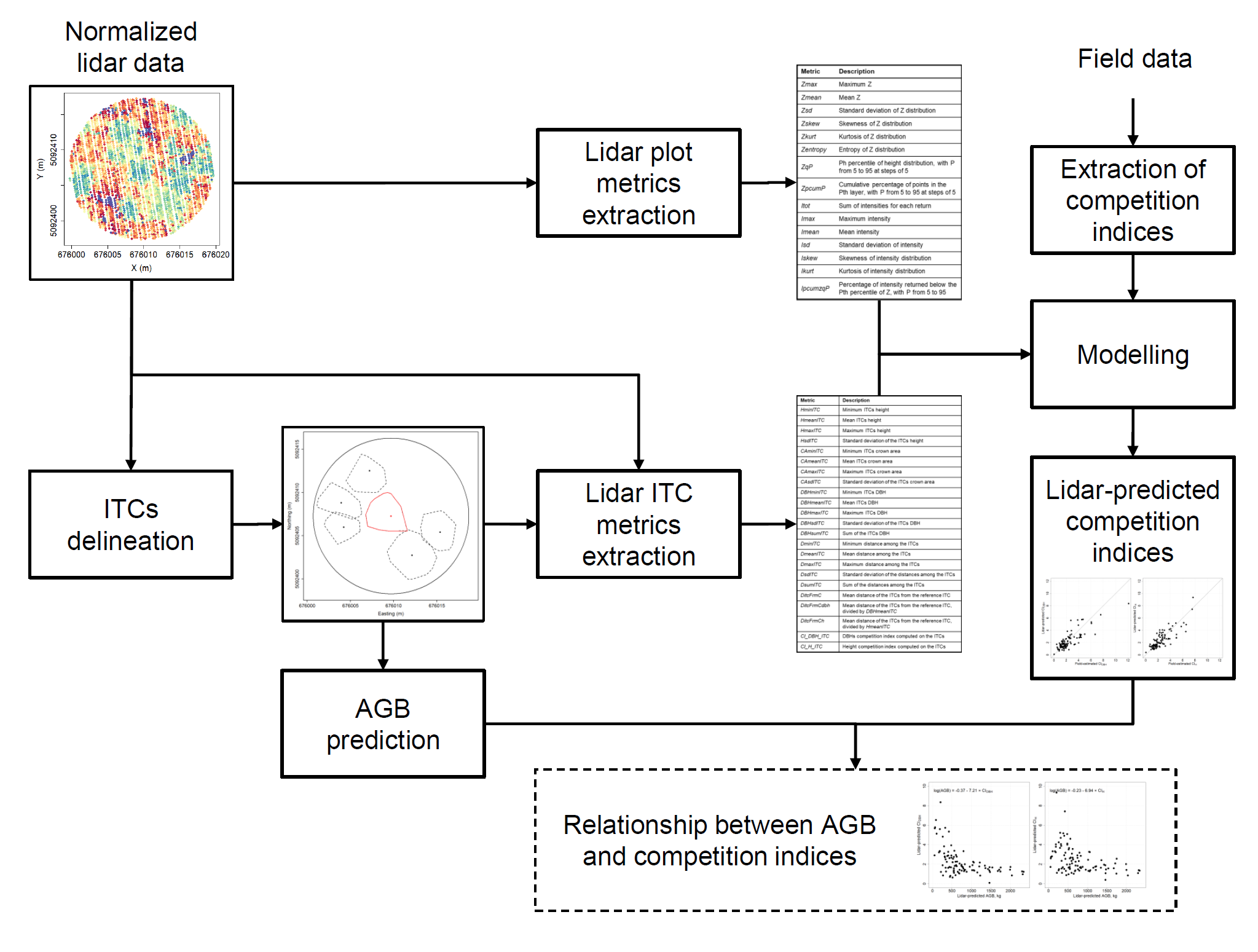
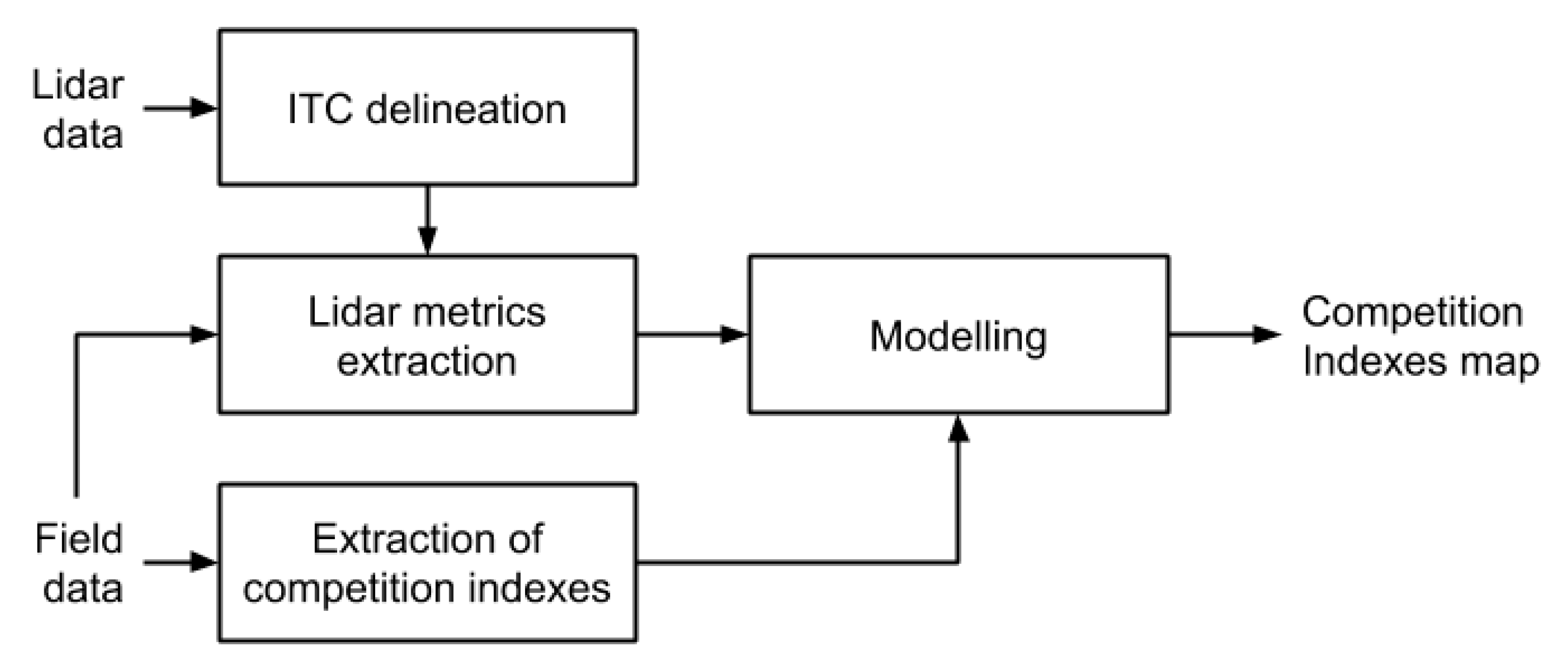
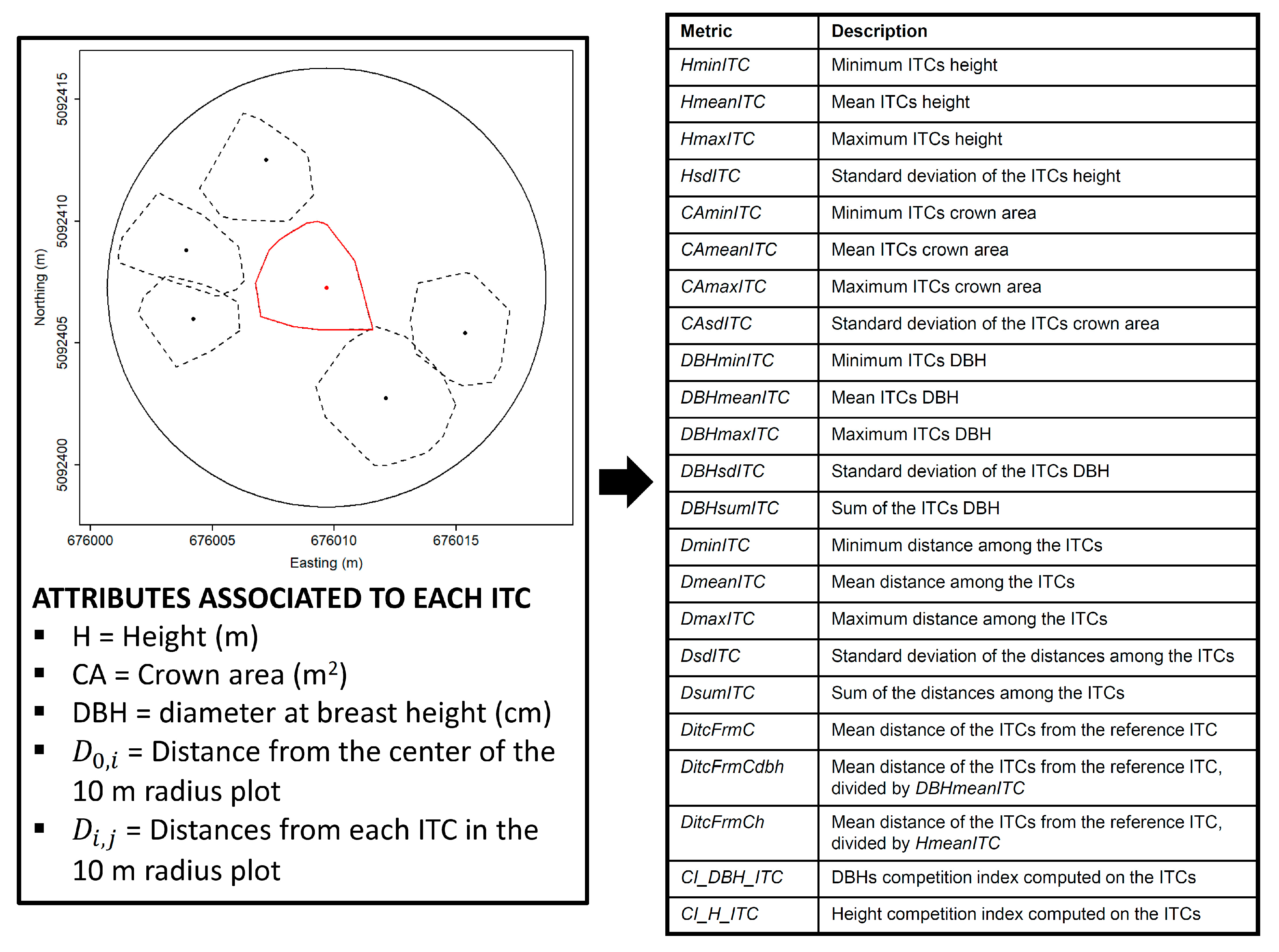
2.2. Extraction of Competition Indices
2.3. ITCs Delineation
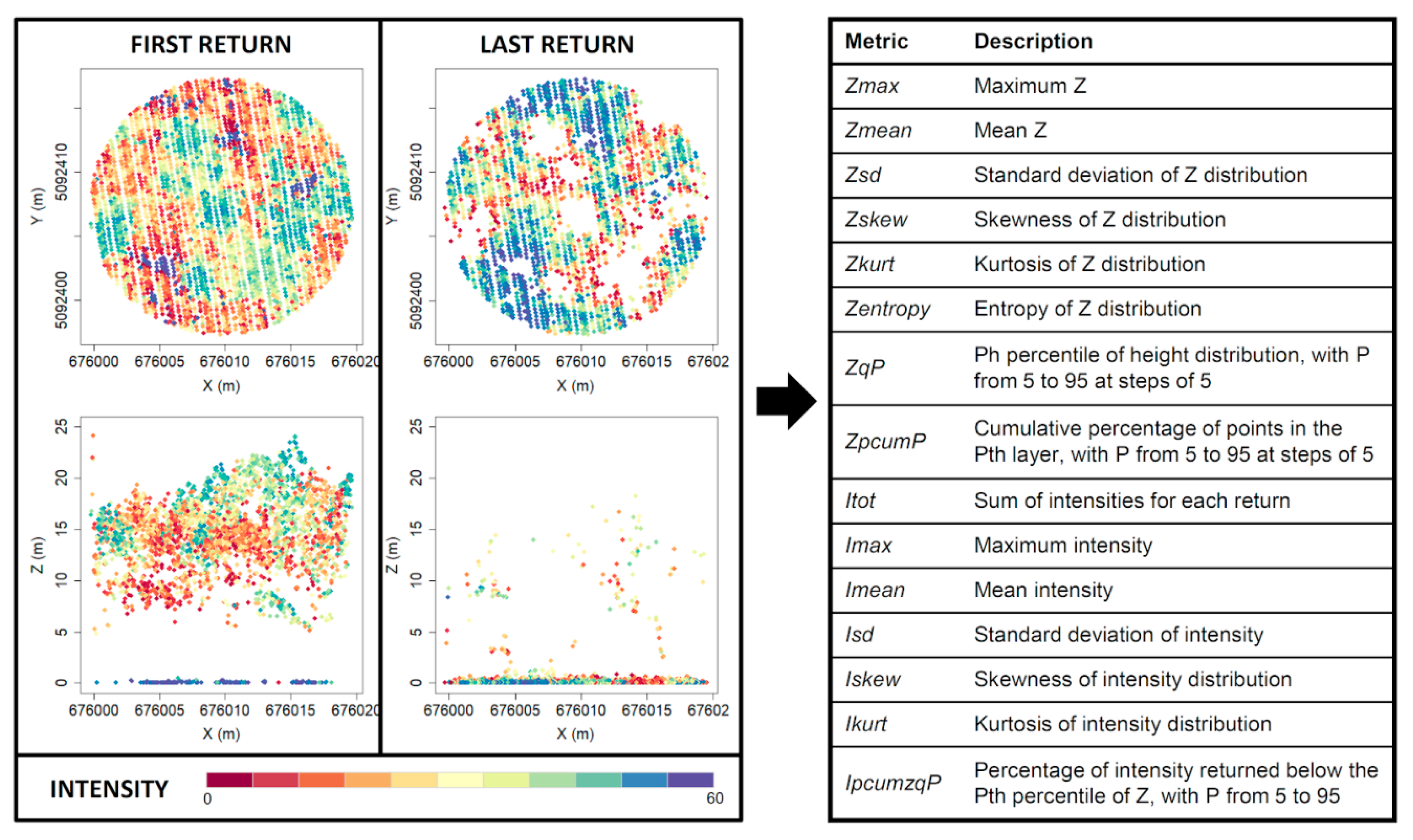
2.4. Lidar Metrics Extraction
2.5. Prediction
2.6. Relationship between AGB and Competition Indices
3. Results
3.1. ITC Crown Delineation
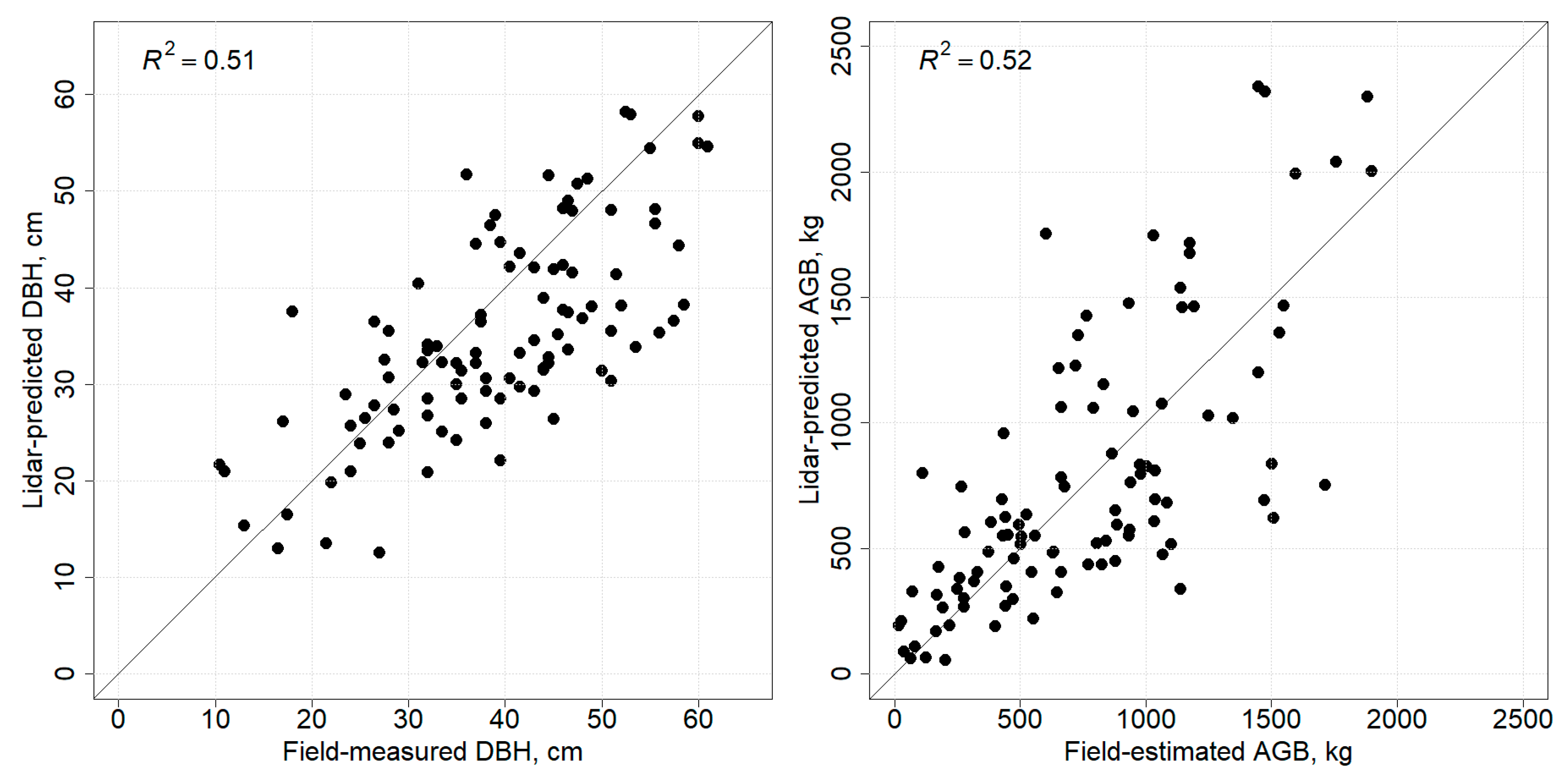
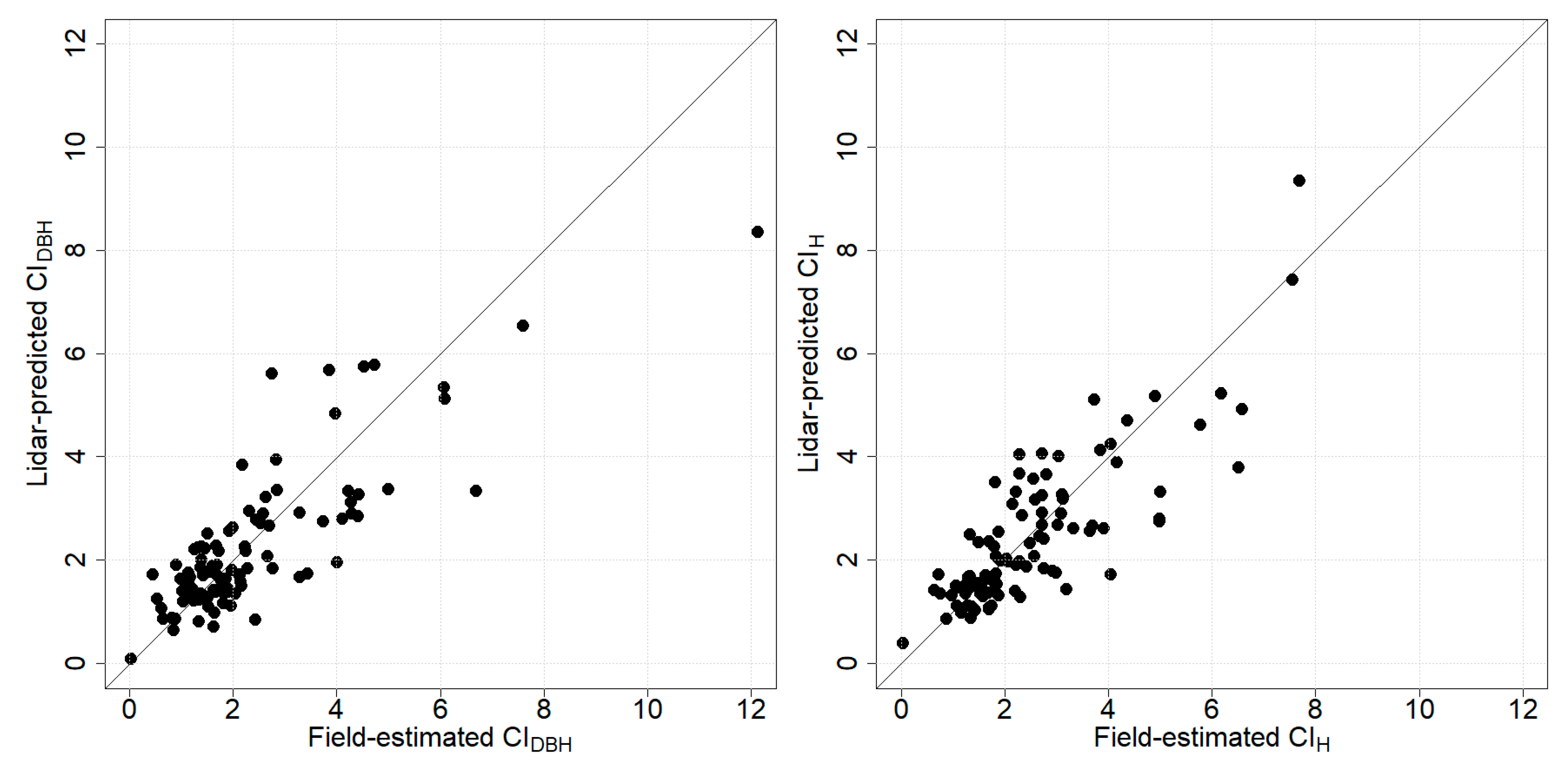
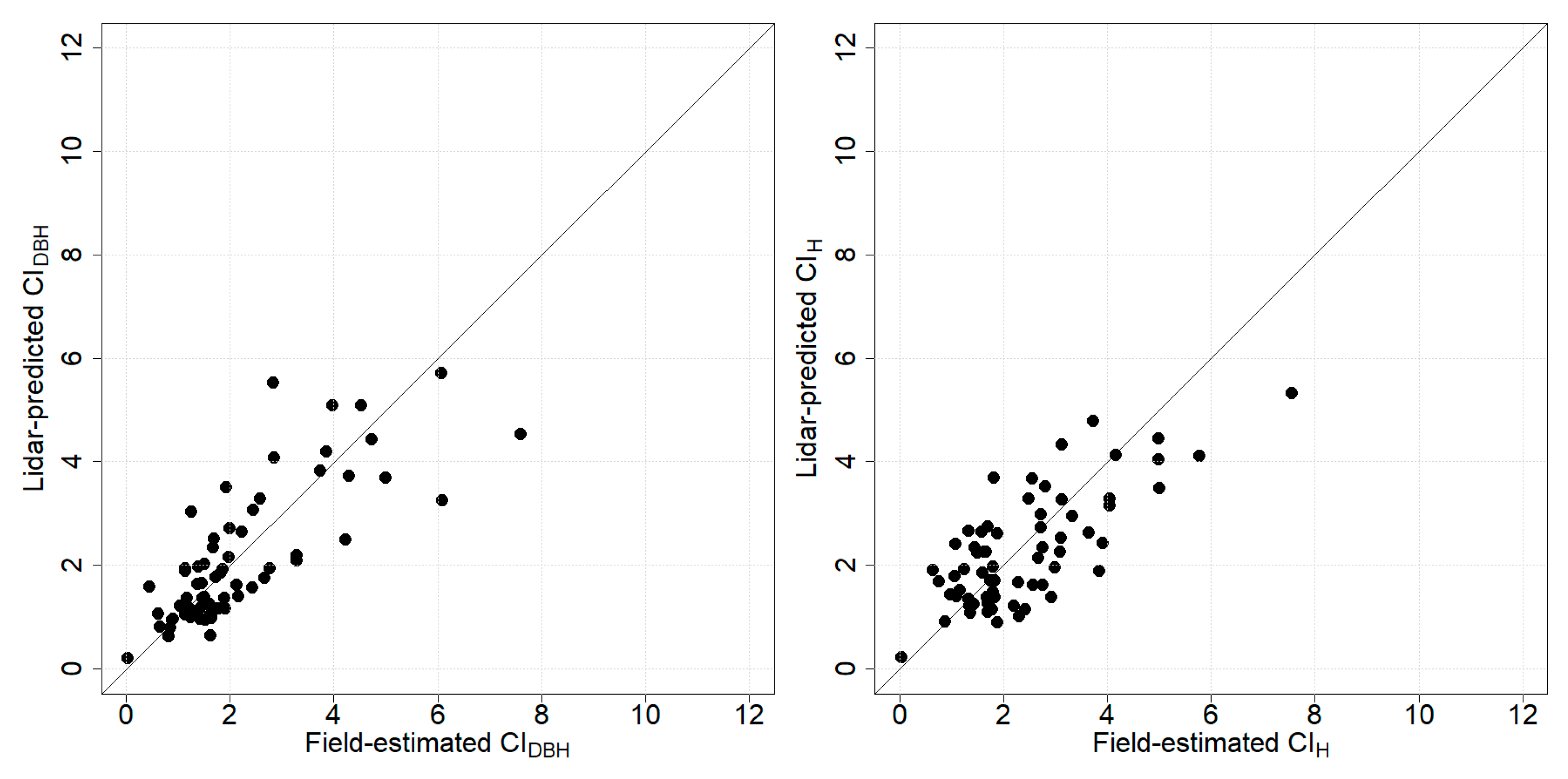
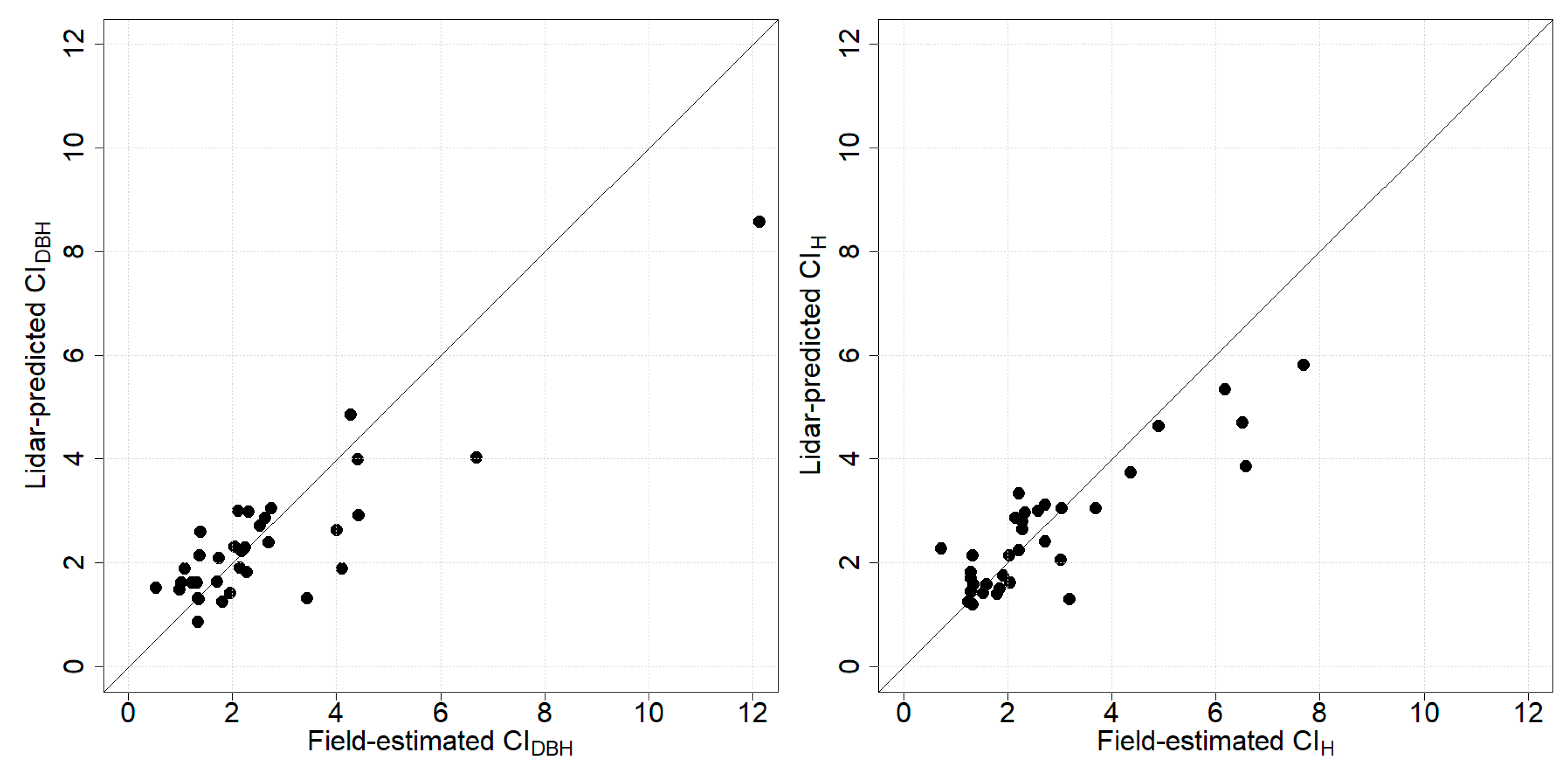
3.2. Prediction of Competition Indices
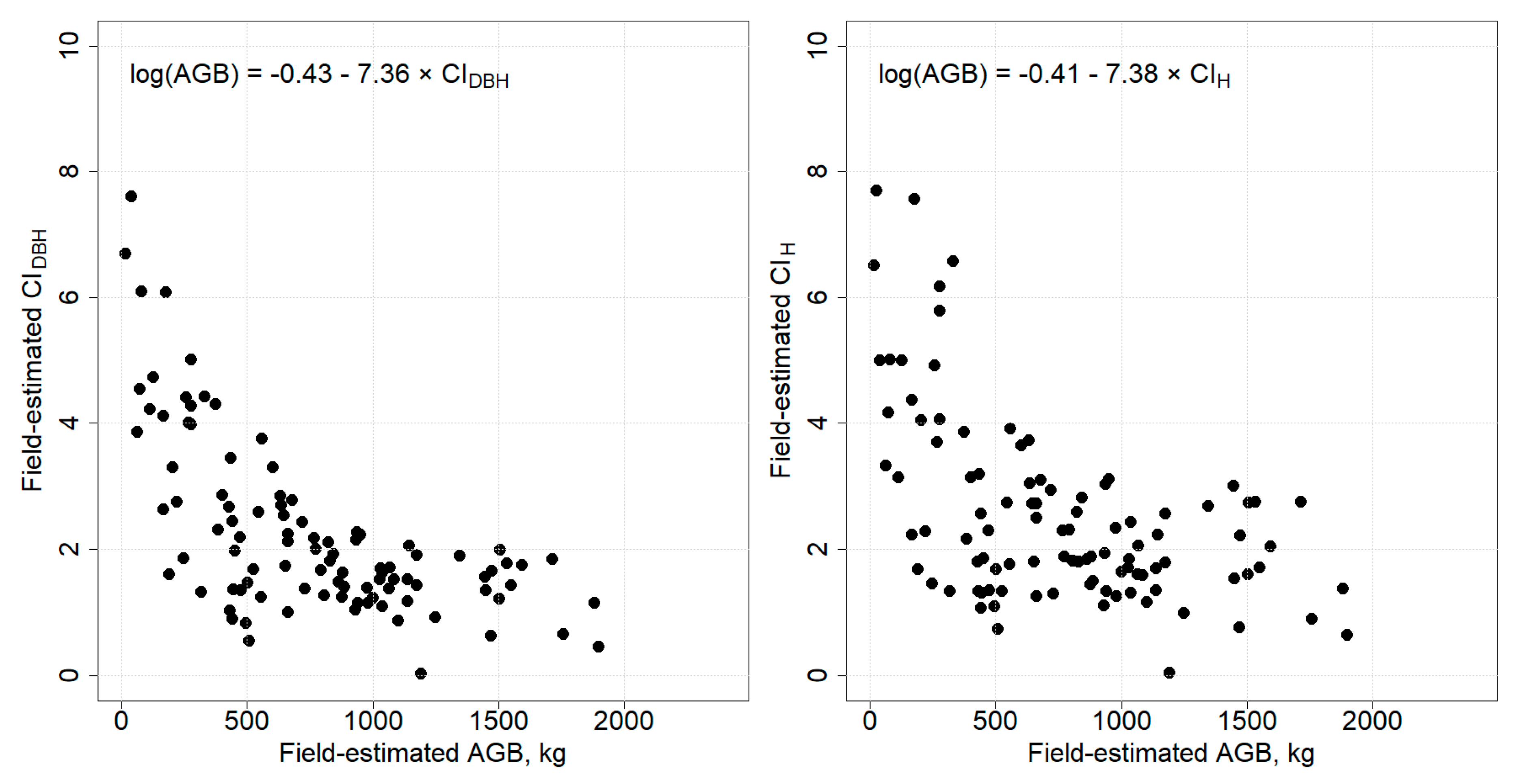
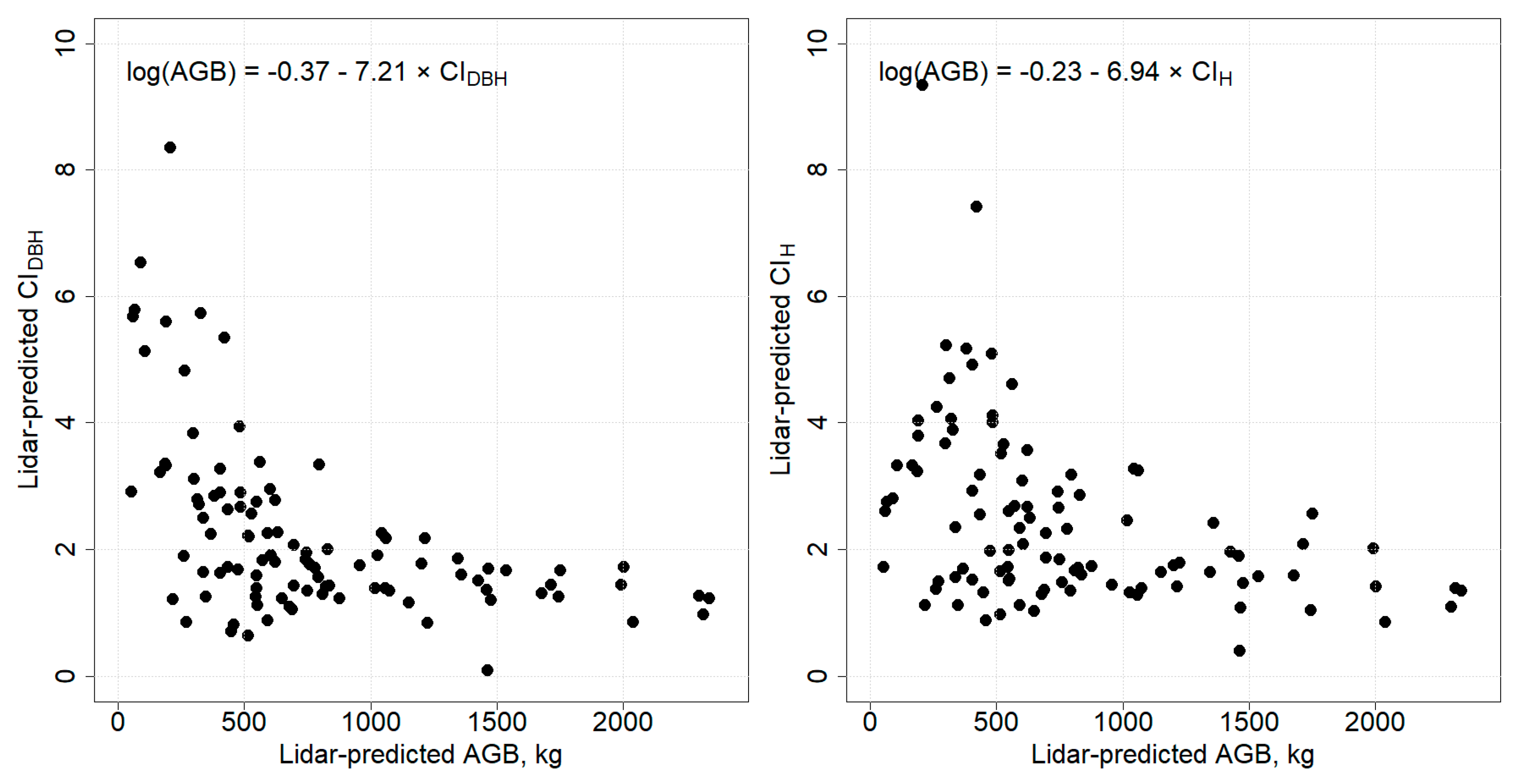
3.3. Relationship between Competition Indices and AGB
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jucker, T.; Avăcăriței, D.; Bărnoaiea, I.; Duduman, G.; Bouriaud, O.; Coomes, D.A. Climate modulates the effects of tree diversity on forest productivity. J. Ecol. 2016, 104, 388–398. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, L. Local Analysis of Tree Competition and Growth. For. Sci. 2003, 49, 938–955. [Google Scholar] [CrossRef]
- Liang, H.; Huang, J.-G.; Ma, Q.; Li, J.; Wang, Z.; Guo, X.; Zhu, H.; Jiang, S.; Zhou, P.; Yu, B.; et al. Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China. Agric. For. Meteorol. 2019, 274, 7–17. [Google Scholar] [CrossRef]
- Aakala, T.; Berninger, F.; Starr, M. The roles of competition and climate in tree growth variation in northern boreal old-growth forests. J. Veg. Sci. 2018, 29, 1040–1051. [Google Scholar] [CrossRef]
- Contreras, M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Ford, K.R.; Breckheimer, I.K.; Franklin, J.F.; Freund, J.A.; Kroiss, S.J.; Larson, A.J.; Theobald, E.J.; HilleRisLambers, J. Competition alters tree growth responses to climate at individual and stand scales. Can. J. For. Res. 2017, 47, 53–62. [Google Scholar] [CrossRef]
- Ledermann, T. Evaluating the performance of semi-distance-independent competition indices in predicting the basal area growth of individual trees. Can. J. For. Res. 2010, 40, 796–805. [Google Scholar] [CrossRef]
- Radtke, P.J.; Westfall, J.A.; Burkhart, H.E. Conditioning a distance-dependent competition index to indicate the onset of inter-tree competition. For. Ecol. Manag. 2003, 175, 17–30. [Google Scholar] [CrossRef]
- Larocque, G.R. Examining Different Concepts for the Development of a Distance-Dependent Competition Model for Red Pine Diameter Growth Using Long-Term Stand Data Differing in Initial Stand Density. For. Sci. 2002, 48, 24–34. [Google Scholar] [CrossRef]
- Kahriman, A.; Şahin, A.; Sönmez, T.; Yavuz, M. A novel approach to selecting a competition index: The effect of competition on individual-tree diameter growth of Calabrian pine. Can. J. For. Res. 2018, 48, 1217–1226. [Google Scholar] [CrossRef]
- McTague, J.; Weiskittel, A. Individual-Tree Competition Indices and Improved Compatibility with Stand-Level Estimates of Stem Density and Long-Term Production. Forests 2016, 7, 238. [Google Scholar] [CrossRef]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.-D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Sun, S.; Cao, Q.V.; Cao, T. Evaluation of distance-independent competition indices in predicting tree survival and diameter growth. Can. J. For. Res. 2019, 49, 440–446. [Google Scholar] [CrossRef]
- Kuehne, C.; Weiskittel, A.R.; Waskiewicz, J. Comparing performance of contrasting distance-independent and distance-dependent competition metrics in predicting individual tree diameter increment and survival within structurally-heterogeneous, mixed-species forests of Northeastern United States. For. Ecol. Manag. 2019, 433, 205–216. [Google Scholar] [CrossRef]
- Hui, G.; Wang, Y.; Zhang, G.; Zhao, Z.; Bai, C.; Liu, W. A novel approach for assessing the neighborhood competition in two different aged forests. For. Ecol. Manag. 2018, 422, 49–58. [Google Scholar] [CrossRef]
- Stadt, K.J.; Huston, C.; Coates, K.D.; Feng, Z.; Dale, M.R.T.; Lieffers, V.J. Evaluation of competition and light estimation indices for predicting diameter growth in mature boreal mixed forests. Ann. For. Sci. 2007, 64, 477–490. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. Evaluation of Competition Indices in Individual Tree Growth Models. For. Sci. 1995, 41, 360–377. [Google Scholar] [CrossRef]
- Tomé, M.; Burkhart, H. Distance-Dependent Competition Measures for Predicting Growth of Individual Trees. For. Sci. 1989, 35, 816–831. [Google Scholar]
- Dale, V.H.; Doyle, T.W.; Shugart, H.H. A comparison of tree growth models. Ecol. Model. 1985, 29, 145–169. [Google Scholar] [CrossRef]
- Thorpe, H.C.; Astrup, R.; Trowbridge, A.; Coates, K.D. Competition and tree crowns: A neighborhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Lang, A.C.; Härdtle, W.; Bruelheide, H.; Geißler, C.; Nadrowski, K.; Schuldt, A.; Yu, M.; von Oheimb, G. Tree morphology responds to neighbourhood competition and slope in species-rich forests of subtropical China. For. Ecol. Manag. 2010, 260, 1708–1715. [Google Scholar] [CrossRef]
- Purves, D.W.; Lichstein, J.W.; Pacala, S.W. Crown Plasticity and Competition for Canopy Space: A New Spatially Implicit Model Parameterized for 250 North American Tree Species. PLoS ONE 2007, 2, e870. [Google Scholar] [CrossRef]
- King, D.A.; Davies, S.J.; Supardi, M.N.N.; Tan, S. Tree growth is related to light interception and wood density in two mixed dipterocarp forests of Malaysia. Funct. Ecol. 2005, 19, 445–453. [Google Scholar] [CrossRef]
- Wyckoff, P.H.; Clark, J.S. Tree growth prediction using size and exposed crown area. Can. J. For. Res. 2005, 35, 13–20. [Google Scholar] [CrossRef]
- Lu, H.; Mohren, G.M.J.; den Ouden, J.; Goudiaby, V.; Sterck, F.J. Overyielding of temperate mixed forests occurs in evergreen-deciduous but not in deciduous-deciduous species mixtures over time in the Netherlands. For. Ecol. Manag. 2016, 376, 321–332. [Google Scholar] [CrossRef]
- Forrester, D.I.; Theiveyanathan, S.; Collopy, J.J.; Marcar, N.E. Enhanced water use efficiency in a mixed Eucalyptus globulus and Acacia mearnsii plantation. For. Ecol. Manag. 2010, 259, 1761–1770. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Laganière, J.; Reich, P.B.; Bergeron, Y.; Paré, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Bosela, M.; Tobin, B.; Šebeň, V.; Petráš, R.; Larocque, G.R. Different mixtures of Norway spruce, silver fir, and European beech modify competitive interactions in central European mature mixed forests. Can. J. For. Res. 2015, 45, 1577–1586. [Google Scholar] [CrossRef]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The Development of Pine Plantation Silviculture in the Southern United States. J. For. 2007, 105, 337–347. [Google Scholar] [CrossRef]
- Davidson, R.L. Effect of Root/Leaf Temperature Differentials on Root/Shoot Ratios in Some Pasture Grasses and Clover. Ann. Bot. 1969, 33, 561–569. [Google Scholar] [CrossRef]
- Sebastià, M.-T. Plant guilds drive biomass response to global warming and water availability in subalpine grassland. J. Appl. Ecol. 2006, 44, 158–167. [Google Scholar] [CrossRef]
- Zhou, W.; Cheng, X.; Wu, R.; Han, H.; Kang, F.; Zhu, J.; Tian, P. Effect of intraspecific competition on biomass partitioning of Larix principis-rupprechtii. J. Plant Interact. 2018, 13, 1–8. [Google Scholar] [CrossRef]
- Lin, Y.; Huth, F.; Berger, U.; Grimm, V. The role of belowground competition and plastic biomass allocation in altering plant mass-density relationships. Oikos 2014, 123. [Google Scholar] [CrossRef]
- Cahill, J.F., Jr.; Casper, B.B. Investigating the relationship between neighbor root biomass and belowground competition: Field evidence for symmetric competition belowground. Oikos 2000, 90, 311–320. [Google Scholar] [CrossRef]
- Petersen, K.S.; Ares, A.; Terry, T.A.; Harrison, R.B. Vegetation competition effects on aboveground biomass and macronutrients, leaf area, and crown structure in 5-year old Douglas-fir. New For. 2008, 35, 299–311. [Google Scholar] [CrossRef]
- Badreldin, N.; Sanchez-Azofeifa, A. Estimating Forest Biomass Dynamics by Integrating Multi-Temporal Landsat Satellite Images with Ground and Airborne LiDAR Data in the Coal Valley Mine, Alberta, Canada. Remote Sens. 2015, 7, 2832–2849. [Google Scholar] [CrossRef]
- Hansen, E.; Gobakken, T.; Bollandsås, O.; Zahabu, E.; Næsset, E. Modeling Aboveground Biomass in Dense Tropical Submontane Rainforest Using Airborne Laser Scanner Data. Remote Sens. 2015, 7, 788–807. [Google Scholar] [CrossRef]
- Sheridan, R.; Popescu, S.; Gatziolis, D.; Morgan, C.; Ku, N.-W. Modeling Forest Aboveground Biomass and Volume Using Airborne LiDAR Metrics and Forest Inventory and Analysis Data in the Pacific Northwest. Remote Sens. 2014, 7, 229–255. [Google Scholar] [CrossRef]
- Popescu, S.C.; Wynne, R.H.; Scrivani, J.A. Fusion of Small-Footprint Lidar and Multispectral Data to Estimate Plot-Level Volume and Biomass in Deciduous and Pine Forests in Virginia, USA. For. Sci. 2004, 50, 551–565. [Google Scholar] [CrossRef]
- Simonson, W.; Ruiz-Benito, P.; Valladares, F.; Coomes, D. Modelling above-ground carbon dynamics using multi-temporal airborne lidar: Insights from a Mediterranean woodland. Biogeosciences 2016, 13, 961–973. [Google Scholar] [CrossRef]
- Lo, C.-S.; Lin, C. Growth-Competition-Based Stem Diameter and Volume Modeling for Tree-Level Forest Inventory Using Airborne LiDAR Data. IEEE Trans. Geosci. Remote Sens. 2013, 51, 2216–2226. [Google Scholar] [CrossRef]
- Lin, C.; Thomson, G.; Popescu, S. An IPCC-Compliant Technique for Forest Carbon Stock Assessment Using Airborne LiDAR-Derived Tree Metrics and Competition Index. Remote Sens. 2016, 8, 528. [Google Scholar] [CrossRef]
- Ma, Q.; Su, Y.; Tao, S.; Guo, Q. Quantifying individual tree growth and tree competition using bi-temporal airborne laser scanning data: A case study in the Sierra Nevada Mountains, California. Int. J. Digit. Earth 2018, 11, 485–503. [Google Scholar] [CrossRef]
- Yu, X.; Hyyppä, J.; Litkey, P.; Kaartinen, H.; Vastaranta, M.; Holopainen, M. Single-Sensor Solution to Tree Species Classification Using Multispectral Airborne Laser Scanning. Remote Sens. 2017, 9, 108. [Google Scholar] [CrossRef]
- Scrinzi, G.; Galvagni, D.; Marzullo, L. I Nuovi Modelli Dendrometrici Per la Stima Delle Masse Assestamentali in Provincia di Trento. 2010. Available online: http://sito.entecra.it/portale/public/documenti/sff_modelli_dendrometrici.pdf?lingua=EN (accessed on 15 September 2019).
- Dalponte, M.; Coomes, D.A. Tree-centric mapping of forest carbon density from airborne laser scanning and hyperspectral data. Methods Ecol. Evol. 2016, 7, 1236–1245. [Google Scholar] [CrossRef]
- IPCC Guidelines for National Greenhouse Gas Inventories. Agriculture, Forestry and Other Land Use. 2006. Available online: https://www.ipcc-nggip.iges.or.jp/public/2006gl/ (accessed on 15 September 2019).
- Wang, D.; Tang, S.-C.; Hsieh, H.-C.; Chung, C.-H.; Lin, C.-Y. Distance-dependent competition measures for individual tree growth on a Taiwania plantation in the Liuguei area. Taiwan J. For. Sci. 2012, 27, 215–227. [Google Scholar]
- Hegyi, F. A simulation model for managing jack-pine stands. In Growth Models for Tree and Stand Simulation; Fries, J.R., Ed.; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–76. [Google Scholar]
- Papaik, M.J.; Canham, C.D. Multi-model analysis of tree competition along environmental gradients in southern New England forests. Ecol. Appl. 2006, 16, 1880–1892. [Google Scholar] [CrossRef]
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Szewczyk, J.; Maciejewski, Z. Shade-tolerant tree species from temperate forests differ in their competitive abilities: A case study from Roztocze, south-eastern Poland. For. Ecol. Manag. 2012, 282, 28–35. [Google Scholar] [CrossRef]
- Dalponte, M. Package ‘itcSegment’; 2018; pp. 1–12. Available online: https://cran.r-project.org/web/packages/itcSegment/index.html (accessed on 15 September 2019).
- Dalponte, M.; Frizzera, L.; Ørka, H.O.; Gobakken, T.; Næsset, E.; Gianelle, D. Predicting stem diameters and aboveground biomass of individual trees using remote sensing data. Ecol. Indic. 2018, 85. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23. [Google Scholar] [CrossRef]
- Zhao, K.; Suarez, J.C.; Garcia, M.; Hu, T.; Wang, C.; Londo, A. Utility of multitemporal lidar for forest and carbon monitoring: Tree growth, biomass dynamics, and carbon flux. Remote Sens. Environ. 2018, 204, 883–897. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28. [Google Scholar] [CrossRef]
- Valbuena, R.; Hernando, A.; Manzanera, J.A.; Görgens, E.B.; Almeida, D.R.A.; Mauro, F.; García-Abril, A.; Coomes, D.A. Enhancing of accuracy assessment for forest above-ground biomass estimates obtained from remote sensing via hypothesis testing and overfitting evaluation. Ecol. Model. 2017, 366, 15–26. [Google Scholar] [CrossRef]
- Lipovetsky, S. How Good is Best? Multivariate Case of Ehrenberg-Weisberg Analysis of Residual Errors in Competing Regressions. J. Mod. Appl. Stat. Methods 2013, 12, 242–255. [Google Scholar] [CrossRef]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Temesgen, H.; LeMay, V.; Mitchell, S.J. Tree crown ratio models for multi-species and multi-layered stands of southeastern British Columbia. For. Chron. 2005, 81, 133–141. [Google Scholar] [CrossRef]
- Toma, B. Prediction equations for estimating tree height, crown diameter, crown height and crown ratio of Parkia biglobosa in the Nigerian guinea savanna. Afr. J. Agric. Res. 2012, 7, 6541–6543. [Google Scholar] [CrossRef]
- Waring, R.; Thies, W.; Muscato, D. Stem Growth per Unit of Leaf Area: A Measure of Tree Vigor. For. Sci. 1980, 26, 112–117. [Google Scholar]
- Rodríguez, R.; Espinosa, M.; Hofmann, G.; Marchant, M. Needle mass, fine root and stem wood production in response to silvicultural treatment, tree size and competitive status in radiata pine stands. For. Ecol. Manag. 2003, 186, 287–296. [Google Scholar] [CrossRef]
- Rottmann, M. Waldbauliche Konsequenzen aus Schneebruchkatastrophen. Schweiz. Z. Forstwes. 1985, 136, 167–184. [Google Scholar]
- Vospernik, S.; Monserud, R.A.; Sterba, H. Do individual-tree growth models correctly represent height:diameter ratios of Norway spruce and Scots pine? For. Ecol. Manag. 2010, 260, 1735–1753. [Google Scholar] [CrossRef]
- Zhen, Z.; Quackenbush, L.; Zhang, L. Trends in Automatic Individual Tree Crown Detection and Delineation—Evolution of LiDAR Data. Remote Sens. 2016, 8, 333. [Google Scholar] [CrossRef]
- Eysn, L.; Hollaus, M.; Lindberg, E.; Berger, F.; Monnet, J.-M.; Dalponte, M.; Kobal, M.; Pellegrini, M.; Lingua, E.; Mongus, D.; et al. A benchmark of lidar-based single tree detection methods using heterogeneous forest data from the Alpine Space. Forests 2015, 6, 1721–1747. [Google Scholar] [CrossRef]
- Rosette, J.; North, P.R.J.; Rubio-Gil, J.; Cook, B.; Los, S.; Suarez, J.; Sun, G.; Ranson, J.; Blair, J.B. Evaluating Prospects for Improved Forest Parameter Retrieval From Satellite LiDAR Using a Physically-Based Radiative Transfer Model. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 45–53. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Trees | Norway Spruce | Silver Fir | |
|---|---|---|---|
| Height (m) | 16.9 (40.1, 2.1) | 20.5 (40.1, 3.1) | 18.1 (40.0, 3.2) |
| DBH (cm) | 22.3 (81.0, 7.0) | 28.6 (79.5, 7.0) | 25.4 (81.0, 7.0) |
| AGB (kg) | 284.7 (3468.5, 1.2) | 453.5 (3215.0, 1.6) | 353.6 (2917.7, 1.6) |
| 5.1 (19.3, 0.04) | 5.0 (18.3, 0.7) | 4.2 (19.3, 0.04) | |
| 5.7 (22.8, 0.02) | 5.1 (17.5, 0.5) | 4.3 (15.8, 0.02) |
| Lidar Metric | Estimate | Std. Error | p | Significance |
|---|---|---|---|---|
| (Intercept) | −2.77 | 0.52 | 0.00 | *** |
| CI_H_ITC | 1.82 | 0.21 | 0.00 | *** |
| Zpcum1_F | −0.02 | 0.00 | 0.00 | *** |
| Iskew_L | −0.28 | 0.11 | 0.01 | * |
| DBHsumITC | −0.01 | 0.00 | 0.00 | *** |
| DsdITC | 0.30 | 0.08 | 0.00 | *** |
| CAmeanITC | 0.03 | 0.01 | 0.00 | *** |
| Lidar Metric | Estimate | Std. Error | p | Significance |
|---|---|---|---|---|
| (Intercept) | 2.13 | 0.58 | 0.00 | *** |
| CI_H_ITC | 0.59 | 0.11 | 0.00 | *** |
| Iskew_F | 0.37 | 0.11 | 0.00 | ** |
| Zpcum2_F | −0.02 | 0.00 | 0.00 | *** |
| Isd_L | −0.15 | 0.04 | 0.00 | *** |
| Zq95_L | −0.03 | 0.01 | 0.00 | ** |
| Lidar metric | Estimate | Significance | Lidar metric | Estimate | Significance |
|---|---|---|---|---|---|
| (Intercept) | −1.07 | *** | (Intercept) | −1.25 | ** |
| CI_H_ITC | 1.42 | *** | CI_DBH_ITC | 0.59 | *** |
| Zpcum2_F | −0.03 | *** | DsdITC | 0.43 | *** |
| Zq20_F | −0.05 | *** | Zq95_L | −0.05 | *** |
| Lidar metric | Estimate | Significance | Lidar metric | Estimate | Significance |
|---|---|---|---|---|---|
| (Intercept) | 17.39 | ** | (Intercept) | 3.71 | *** |
| Ipcumzq90_F | −0.13 | * | Imean_F | −0.10 | *** |
| Imean_F | −0.09 | ** | Isd_L | −0.19 | *** |
| Ipcumzq50_L | −0.05 | ** | Zpcum2_L | 0.02 | * |
| All Trees | Silver Fir | Norway Spruce | ||||
|---|---|---|---|---|---|---|
| Number of samples | 100 | 100 | 66 | 66 | 34 | 34 |
| Metrics selected | 6 | 5 | 3 | 3 | 3 | 3 |
| MD | −0.11 | −0.09 | −0.09 | −0.11 | −0.30 | −0.20 |
| MD% | −4.8 | −3.4 | −4.1 | −4.5 | −11.4 | −7.2 |
| MAD | 0.75 | 0.67 | 0.70 | 0.83 | 0.89 | 0.71 |
| MAD% | 31.9 | 26.7 | 32.0 | 34.3 | 33.9 | 25.7 |
| RMSD | 1.03 | 0.92 | 0.98 | 0.99 | 1.37 | 1.00 |
| RMSD% | 43.9 | 36.3 | 44.7 | 41.2 | 52.0 | 36.3 |
| 0.68 | 0.66 | 0.60 | 0.53 | 0.72 | 0.73 | |
| 0.64 | 0.62 | 0.54 | 0.44 | 0.57 | 0.66 | |
| R2R | 1.08 | 1.07 | 1.11 | 1.19 | 1.27 | 1.10 |
| SSR | 1.07 | 1.07 | 1.07 | 1.08 | 1.25 | 1.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Versace, S.; Gianelle, D.; Frizzera, L.; Tognetti, R.; Garfì, V.; Dalponte, M. Prediction of Competition Indices in a Norway Spruce and Silver Fir-Dominated Forest Using Lidar Data. Remote Sens. 2019, 11, 2734. https://doi.org/10.3390/rs11232734
Versace S, Gianelle D, Frizzera L, Tognetti R, Garfì V, Dalponte M. Prediction of Competition Indices in a Norway Spruce and Silver Fir-Dominated Forest Using Lidar Data. Remote Sensing. 2019; 11(23):2734. https://doi.org/10.3390/rs11232734
Chicago/Turabian StyleVersace, Soraya, Damiano Gianelle, Lorenzo Frizzera, Roberto Tognetti, Vittorio Garfì, and Michele Dalponte. 2019. "Prediction of Competition Indices in a Norway Spruce and Silver Fir-Dominated Forest Using Lidar Data" Remote Sensing 11, no. 23: 2734. https://doi.org/10.3390/rs11232734
APA StyleVersace, S., Gianelle, D., Frizzera, L., Tognetti, R., Garfì, V., & Dalponte, M. (2019). Prediction of Competition Indices in a Norway Spruce and Silver Fir-Dominated Forest Using Lidar Data. Remote Sensing, 11(23), 2734. https://doi.org/10.3390/rs11232734