A Multi-Temporal Object-Based Image Analysis to Detect Long-Lived Shrub Cover Changes in Drylands

 ,
,  , , and
, , and 
Abstract
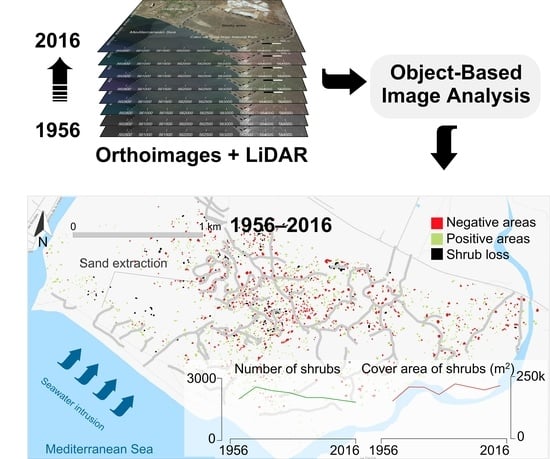
1. Introduction
2. Materials and Methods
2.1. Study Case
2.2. Datasets and Ground Truth
2.3. Object-Based Image Analysis
2.3.1. Image Segmentation
2.3.2. Classification and Validation of Segments
2.4. Sand Extraction Curvature Analysis
2.5. Shrub Area and Shape Dynamics
3. Results
3.1. Segmentation Accuracy
3.2. Classification and Characteristics of Ziziphus lotus Shrubs
3.3. Shrub Number, Area, and Shape Dynamics
3.4. Sand Extraction Mapping and Curvature Analysis
3.5. Spatial Relationships of Shrubs with Sand Extractions, Coastline (Seawater Intrusion), and Protected Area
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Kareiva, P. Spatial Ecology: The Role of Space in Population Dynamics and Interspecific Interactions (MPB-30); Princeton University Press: Princeton, NJ, USA, 2018. [Google Scholar]
- Maestre, F.T.; Escudero, A.; Martinez, I.; Guerrero, C.; Rubio, A. Does spatial pattern matter to ecosystem functioning? Insights from biological soil crusts. Funct. Ecol. 2005, 19, 566–573. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Wilcox, B.P.; Breshears, D.D.; Tongway, D.J.; Imeson, A.C. Vegetation Patches and Runoff–Erosion as Interacting Ecohydrological Processes in Semiarid Landscapes. Ecology 2005, 86, 288–297. [Google Scholar] [CrossRef]
- Thompson, S.E.; Harman, C.J.; Troch, P.A.; Brooks, P.D.; Sivapalan, M. Spatial scale dependence of ecohydrologically mediated water balance partitioning: A synthesis framework for catchment ecohydrology. Water Resour. Res. 2011, 47. [Google Scholar] [CrossRef]
- Aguiar, M.R.; Sala, O.E. Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends Ecol. Evol. 1999, 14, 273–277. [Google Scholar] [CrossRef]
- Puigdefábregas, J. The role of vegetation patterns in structuring runoff and sediment fluxes in drylands. Earth Surf. Process. Landf. 2005, 30, 133–147. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Virginia, R.A.; Kemp, P.R.; De Soyza, A.G.; Tremmel, D.C. Impact of drought on desert shrubs: Effects of seasonality and degree of resource island development. Ecol. Monogr. 1999, 69, 69–106. [Google Scholar] [CrossRef]
- Berdugo, M.; Maestre, F.T.; Kéfi, S.; Gross, N.; Le Bagousse-Pinguet, Y.; Soliveres, S. Aridity preferences alter the relative importance of abiotic and biotic drivers on plant species abundance in global drylands. J. Ecol. 2019, 107, 190–202. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Tsai, Y.; Stow, D.; Chen, H.; Lewison, R.; An, L.; Shi, L. Mapping vegetation and land use types in fanjingshan national nature reserve using Google Earth Engine. Remote Sens. 2018, 10, 927. [Google Scholar] [CrossRef]
- Minasny, B.; Setiawan, B.I.; Saptomo, S.K.; McBratney, A.B. Open digital mapping as a cost-effective method for mapping peat thickness and assessing the carbon stock of tropical peatlands. Geoderma 2018, 313, 25–40. [Google Scholar]
- Schiewe, J.; Tufte, L.; Ehlers, M. Potential and problems of multi-scale segmentation methods in remote sensing. GeoBIT/GIS 2001, 6, 34–39. [Google Scholar]
- Zheng, X.; Wu, B.; Weston, M.; Zhang, J.; Gan, M.; Zhu, J.; Deng, J.; Wang, K.; Teng, L. Rural settlement subdivision by using landscape metrics as spatial contextual information. Remote Sens. 2017, 9, 486. [Google Scholar] [CrossRef]
- Hallberg, B.; Smith-Jonforsen, G.; Ulander, L.M. Measurements on individual trees using multiple VHF SAR images. IEEE Trans. Geosci. Remote Sens. 2005, 43, 2261–2269. [Google Scholar] [CrossRef]
- Maksymiuk, O.; Schmitt, M.; Auer, S.; Stilla, U. Single tree detection in millimeterwave SAR data by morphological attribute filters. Proc. Jahrestag. DGPF 2014, 34. [Google Scholar]
- Hamraz, H.; Contreras, M.A.; Zhang, J. Vertical stratification of forest canopy for segmentation of understory trees within small-footprint airborne LiDAR point clouds. ISPRS J. Photogramm. Remote Sens. 2017, 130, 385–392. [Google Scholar] [CrossRef]
- Blaschke, T. Object based image analysis for remote sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- Deblauwe, V.; Barbier, N.; Couteron, P.; Lejeune, O.; Bogaert, J. The global biogeography of semi-arid periodic vegetation patterns. Glob. Ecol. Biogeogr. 2008, 17, 715–723. [Google Scholar] [CrossRef]
- Kéfi, S.; Rietkerk, M.; Alados, C.L.; Pueyo, Y.; Papanastasis, V.P.; ElAich, A.; de Ruiter, P.C. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 2007, 449, 213–217. [Google Scholar] [CrossRef]
- Burnett, C.; Blaschke, T. A multi-scale segmentation/object relationship modelling methodology for landscape analysis. Ecol. Model. 2003, 168, 233–249. [Google Scholar] [CrossRef]
- Hellesen, T.; Matikainen, L. An Object-Based Approach for Mapping Shrub and Tree Cover on Grassland Habitats by Use of LiDAR and CIR Orthoimages. Remote Sens. 2013, 5, 558–583. [Google Scholar] [CrossRef]
- Kallio, T. Protection of spruce stumps against Fomes annosus (Fr.) Cooke by some wood-inhabiting fungi. Acta For. Fenn. 1971, 117, 1–20. [Google Scholar] [CrossRef]
- Eriksson, O. Regional dynamics of plants: A review of evidence for remnant, source-sink and metapopulations. Oikos 1996, 77, 248–258. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- García, D.; Zamora, R. Persistence, multiple demographic strategies and conservation in long-lived Mediterranean plants. J. Veg. Sci. 2003, 14, 921–926. [Google Scholar] [CrossRef]
- Rey, P.J.; Cancio, I.; Manzaneda, A.J.; González-Robles, A.; Valera, F.; Salido, T.; Alcántara, J.M. Regeneration of a keystone semiarid shrub over its range in Spain: Habitat degradation overrides the positive effects of plant–animal mutualisms. Plant Biol. 2018, 20, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Guirado, E.; Alcaraz-Segura, D.; Rigol-Sánchez, J.P.; Gisbert, J.; Martínez-Moreno, F.J.; Galindo-Zaldívar, J.; González-Castillo, L.; Cabello, J. Remote-sensing-derived fractures and shrub patterns to identify groundwater dependence. Ecohydrology 2018, 11, 1933. [Google Scholar] [CrossRef]
- Tirado, R.; Pugnaire, F.I. Shrub spatial aggregation and consequences for reproductive success. Oecologia 2003, 136, 296–301. [Google Scholar] [CrossRef]
- Martínez-Lage, A.V. Las extracciones de áridos en el litoral de almería para su utilización en la agricultura intensiva (1956-1997). In Proceedings of the Actas de Las Jornadas Sobre el Litoral de Almería: Caracterización, Ordenación y Gestión de un Espacio Geográfico Celebradas, Andalusia, Spain, 20–24 May 1997; Instituto de Estudios Almerienses: Andalusia, Spain, 1999; pp. 83–110. [Google Scholar]
- Tirado, R. 5220 Matorrales arborescentes con Ziziphus. VV AA Bases Ecológicas Prelim; Para Conserv. Los Tipos Hábitat Interés Comunitario En España; Dir. Gral. de Medio Natural; Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, Spain, 2009. [Google Scholar]
- Daniele, L.; Sola, F.; Izquierdo, A.V.; Bosch, A.P. Coastal aquifers and desalination plants: Some interpretations to new situations. In Proceedings of the Conference on Water Observation and Information System for Decision Support, Balwois, Ohrid, Republic of Macedonia, 25–29 May 2010. [Google Scholar]
- Cancio, I.; González-Robles, A.; Bastida, J.M.; Manzaneda, A.J.; Salido, T.; Rey, P.J. Habitat loss exacerbates regional extinction risk of the keystone semiarid shrub Ziziphus lotus through collapsing the seed dispersal service by foxes (Vulpes vulpes). Biodivers. Conserv. 2016, 25, 693–709. [Google Scholar] [CrossRef]
- Martín-Rosales, W.; Gisbert, J.; Pulido-Bosch, A.; Vallejos, A.; Fernández-Cortés, A. Estimating groundwater recharge induced by engineering systems in a semiarid area (southeastern Spain). Environ. Geol. 2007, 52, 985–995. [Google Scholar] [CrossRef]
- García García, J.P.; Sánchez Caparós, A.; Castillo, E.; Marín, I.; Padilla, A.; Rosso, J.I. Hidrogeoquímica de las aguas subterráneas en la zona de Cabo de Gata. In Tecnología de la Intrusión de Agua de Mar en Acuíferos Costeros: Países Mediterráneos; IGME: Granada, Spain, 2003. [Google Scholar]
- Mendoza-Fernández, A.J.; MartíNez-Hernández, F.; Pérez-García, F.J.; Garrido-Becerra, J.A.; Benito, B.M.; Salmerón-Sánchez, E.; Guirado, J.; Merlo, M.E.; Mota, J.F. Extreme habitat loss in a Mediterranean habitat: Maytenus senegalensis subsp. europaea. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2015, 149, 503–511. [Google Scholar] [CrossRef]
- Rivas Goday, S.; Bellot, F. Las formaciones de Ziziphus lotus (L.) Lamk. en las dunas del Cabo de Gata. An. Inst. Esp. Edafol. Ecol. Fisiol. Veg. 1944, 3, 109–126. [Google Scholar]
- Pugnaire, F.I.; Armas, C.; Maestre, F.T. Positive plant interactions in the Iberian Southeast: Mechanisms, environmental gradients, and ecosystem function. J. Arid Environ. 2011, 75, 1310–1320. [Google Scholar] [CrossRef]
- Oyonarte, C.; Rey, A.; Raimundo, J.; Miralles, I.; Escribano, P. The use of soil respiration as an ecological indicator in arid ecosystems of the SE of Spain: Spatial variability and controlling factors. Ecol. Indic. 2012, 14, 40–49. [Google Scholar] [CrossRef]
- Goy, J.L.; Zazo, C. Synthesis of the quaternary in the almeria littoral neotectonic activity and its morphologic features, western betics, Spain. Tectonophysics 1986, 130, 259–270. [Google Scholar] [CrossRef]
- Sola, F.; Daniele, L.; Sánchez Martos, F.; Vallejos, A.; Urízar, R.; Pulido Bosch, A. Influencia de la desaladora de Rambla Morales (Almería) sobre las características hidrogeológicas del acuífero del que se abastece. Los Acuíferos Costeros Retos Soluc. 2007, 1, 997–1004. [Google Scholar]
- Tirado, R.; Bråthen, K.A.; Pugnaire, F.I. Mutual positive effects between shrubs in an arid ecosystem. Sci. Rep. 2015, 5, 14710. [Google Scholar] [CrossRef]
- Aksoy, S.; Tilton, J.C.; Tarabalka, Y. Image segmentation algorithms for land categorization. In Remote Sensing Handbook V.1 Remotely Sensed Data Characterization, Classification, and Accuracies; Taylor & Francis: Abingdon, UK, 2015; pp. 317–342. ISBN 978-1-4822-1786-5. [Google Scholar]
- Benz, U.C.; Hofmann, P.; Willhauck, G.; Lingenfelder, I.; Heynen, M. Multi-resolution, object-oriented fuzzy analysis of remote sensing data for GIS-ready information. ISPRS J. Photogramm. Remote Sens. 2004, 58, 239–258. [Google Scholar] [CrossRef]
- Drăguţ, L.; Csillik, O.; Eisank, C.; Tiede, D. Automated parameterisation for multi-scale image segmentation on multiple layers. ISPRS J. Photogramm. Remote Sens. 2014, 88, 119–127. [Google Scholar] [CrossRef]
- Kavzoglu, T.; Tonbul, H. A comparative study of segmentation quality for multi-resolution segmentation and watershed transform. In Proceedings of the 2017 8th International Conference on Recent Advances in Space Technologies (RAST), Istanbul, Turkey, 19–22 June 2017; pp. 113–117. [Google Scholar]
- Zhan, Q.; Molenaar, M.; Tempfli, K.; Shi, W. Quality assessment for geo-spatial objects derived from remotely sensed data. Int. J. Remote Sens. 2005, 26, 2953–2974. [Google Scholar] [CrossRef]
- Tian, J.; Chen, D.-M. Optimization in multi-scale segmentation of high-resolution satellite images for artificial feature recognition. Int. J. Remote Sens. 2007, 28, 4625–4644. [Google Scholar] [CrossRef]
- Kavzoglu, T.; Yildiz, M. Parameter-based performance analysis of object-based image analysis using aerial and Quikbird-2 images. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2014, 2, 31. [Google Scholar] [CrossRef]
- Liu, Y.; Bian, L.; Meng, Y.; Wang, H.; Zhang, S.; Yang, Y.; Shao, X.; Wang, B. Discrepancy measures for selecting optimal combination of parameter values in object-based image analysis. ISPRS J. Photogramm. Remote Sens. 2012, 68, 144–156. [Google Scholar] [CrossRef]
- Nussbaum, S.; Menz, G. SEaTH–A New Tool for Feature Analysis. In Object-Based Image Analysis and Treaty Verification: New Approaches in Remote Sensing–Applied to Nuclear Facilities in Iran; Nussbaum, S., Menz, G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 51–62. ISBN 978-1-4020-6961-1. [Google Scholar]
- Gao, Y.; Mas, J.F.; Kerle, N.; Pacheco, J.A.N. Optimal region growing segmentation and its effect on classification accuracy. Int. J. Remote Sens. 2011, 32, 3747–3763. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Safford, H.D.; Cheng, Y.B.; Ustin, S.L. Mapping mountain vegetation using species distribution modeling, image-based texture analysis, and object-based classification. Appl. Veg. Sci. 2008, 11, 499–508. [Google Scholar] [CrossRef]
- Wang, X.Z.; Ashfaq, R.A.R.; Fu, A.M. Fuzziness based sample categorization for classifier performance improvement. J. Intell. Fuzzy Syst. 2015, 29, 1185–1196. [Google Scholar] [CrossRef]
- Cohen, J. Weighted kappa: Nominal scale agreement provision for scaled disagreement or partial credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef]
- Jordan, G. Morphometric analysis and tectonic interpretation of digital terrain data: A case study. Earth Surf. Process. Landf. 2003, 28, 807–822. [Google Scholar] [CrossRef]
- Rigol-Sanchez, J.P.; Stuart, N.; Pulido-Bosch, A. ArcGeomorphometry: A toolbox for geomorphometric characterisation of DEMs in the ArcGIS environment. Comput. Geosci. 2015, 85, 155–163. [Google Scholar] [CrossRef]
- Evans, I.S. General geomorphology, derivatives of altitude and descriptive statistics. In Spatial Analysis in Geomorphology; Chorley, R.J., Ed.; Harper and Row: Manhattan, NY, USA, 1972; pp. 17–90. [Google Scholar]
- Collinge, S.K.; Palmer, T.M. The influences of patch shape and boundary contrast on insect response to fragmentation in California grasslands. Landsc. Ecol. 2002, 17, 647–656. [Google Scholar] [CrossRef]
- Schumaker, N.H. Using Landscape Indices to Predict Habitat Connectivity. Ecology 1996, 77, 1210–1225. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern Analysis, Spatial Statistics and Geographic Exegesis. Version 2. Methods Ecol. Evol. 2011, 2, 229–232. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Rango, A.; Havstad, K.M.; Paris, J.F.; Beck, R.F.; McNeely, R.; Gonzalez, A.L. Object-oriented image analysis for mapping shrub encroachment from 1937 to 2003 in southern New Mexico. Remote Sens. Environ. 2004, 93, 198–210. [Google Scholar] [CrossRef]
- Fernández-Buces, N.; Siebe, C.; Cram, S.; Palacio, J.L. Mapping soil salinity using a combined spectral response index for bare soil and vegetation: A case study in the former lake Texcoco, Mexico. J. Arid Environ. 2006, 65, 644–667. [Google Scholar] [CrossRef]
- Yu, Q.; Gong, P.; Clinton, N.; Biging, G.; Kelly, M.; Schirokauer, D. Object-based Detailed Vegetation Classification with Airborne High Spatial Resolution Remote Sensing Imagery. Photogramm. Eng. Remote Sens. 2006, 7, 799–811. [Google Scholar] [CrossRef]
- Murray, H.; Lucieer, A.; Williams, R. Texture-based classification of sub-Antarctic vegetation communities on Heard Island. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 138–149. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Browning, D.M.; Rango, A. A comparison of three feature selection methods for object-based classification of sub-decimeter resolution UltraCam-L imagery. Int. J. Appl. Earth Obs. Geoinf. 2012, 15, 70–78. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Rango, A. Texture and Scale in Object-Based Analysis of Subdecimeter Resolution Unmanned Aerial Vehicle (UAV) Imagery. IEEE Trans. Geosci. Remote Sens. 2009, 47, 761–770. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Kennedy-Bowdoin, T.; Jones, M.O.; Martin, R.E.; Boardman, J.; Hughes, R.F. Invasive species detection in Hawaiian rainforests using airborne imaging spectroscopy and LiDAR. Remote Sens. Environ. 2008, 112, 1942–1955. [Google Scholar] [CrossRef]
- Walsh, S.J.; McCleary, A.L.; Mena, C.F.; Shao, Y.; Tuttle, J.P.; González, A.; Atkinson, R. QuickBird and Hyperion data analysis of an invasive plant species in the Galapagos Islands of Ecuador: Implications for control and land use management. Remote Sens. Environ. 2008, 112, 1927–1941. [Google Scholar] [CrossRef]
- Partridge, T.R. Vegetation recovery following sand mining on coastal dunes at Kaitorete Spit, Canterbury, New Zealand. Biol. Conserv. 1992, 61, 59–71. [Google Scholar] [CrossRef]
- Tengberg, A.; Chen, D. A comparative analysis of nebkhas in central Tunisia and northern Burkina Faso. Geomorphology 1998, 22, 181–192. [Google Scholar] [CrossRef]
- Wang, X.; Yang, F.; Yang, D.; Chen, X. Relationship between the growth of Tamarix ramosissima and morphology of nebkhas in oasis-desert ecotones. In Global Climate Change and Its Impact on Food & Energy Security in the Drylands, Proceedings of the Eleventh International Dryland Development Conference, Beijing, China, 18–21 March 2013; International Dryland Development Commission (IDDC): Beijing, China, 2014; pp. 616–628. [Google Scholar]
- Sánchez, F.J.T. El uso del agua en Nijar: Implicaciones ambientales del modelo actual de gestión. Rev. Estud. Reg. 2008, 83, 145–176. [Google Scholar]
- Houérou, H.N.L. Agroforestry and sylvopastoralism: The role of trees and shrubs (Trubs) in range rehabilitation and development. Sci. Chang. Planétaires Sécher. 2006, 17, 343–348. [Google Scholar]
- Howard, J.; Merrifield, M. Mapping Groundwater Dependent Ecosystems in California. PLoS ONE 2010, 5, e11249. [Google Scholar] [CrossRef] [PubMed]
- Ponce, V.M. Effect of groundwater pumping on the health of arid vegetative ecosystems. Online Report, December. 2014. Available online: http://ponce.sdsu.edu/effect_of_groundwater_pumping.html (accessed on 10 November 2019).
- Roques, K.G.; O’connor, T.G.; Watkinson, A.R. Dynamics of shrub encroachment in an African savanna: Relative influences of fire, herbivory, rainfall and density dependence. J. Appl. Ecol. 2001, 38, 268–280. [Google Scholar] [CrossRef]
- Sturm, M.; Racine, C.; Tape, K. Increasing shrub abundance in the Arctic. Nature 2001, 411, 546–547. [Google Scholar] [CrossRef]
- Leonard, J.A.; Yeary, R.A. Exposure of Workers Using Hand-Held Equipment During Urban Application of Pesticides to Trees and Ornamental Shrubs. Am. Ind. Hyg. Assoc. J. 1990, 51, 605–609. [Google Scholar] [CrossRef]
- Bear, J.; Cheng, A.H.-D.; Sorek, S.; Ouazar, D.; Herrera, I. Seawater Intrusion in Coastal Aquifers: Concepts, Methods and Practices; Springer Science & Business Media: Berlin, Germany, 1999; Volume 14. [Google Scholar]
- Mahlknecht, J.; Merchán, D.; Rosner, M.; Meixner, A.; Ledesma-Ruiz, R. Assessing seawater intrusion in an arid coastal aquifer under high anthropogenic influence using major constituents, Sr and B isotopes in groundwater. Sci. Total Environ. 2017, 587, 282–295. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Nègre, R. Recherches Phytogéographiques Sur L’étage de Végétation Méditerranéen Aride (Sous-Étage Chaud) au Maroc Occidental; Société des Sciences Naturelles et Physiques du Maroc: Rabat, Morocco, 1959. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Source | Spatial Resolution | Band Number | Year |
|---|---|---|---|
| Andalusian Environmental Information Network (REDIAM) | 1 m/pixel | 1 | 1956 |
| 1 m/pixel | 1 | 1977 | |
| 0.5 m/pixel | 3 | 1984 | |
| 0.5 m/pixel | 3 | 1997 | |
| 0.5 m/pixel | 3 | 2004 | |
| 0.5 m/pixel | 3 | 2008 | |
| Google EarthTM | 0.5 m/pixel | 3 | 2013 |
| 0.5 m/pixel | 3 | 2016 | |
| Airborne laser scanning | 1 m point spacing | - | 2011 |
| Year | 1956 | 1977 | 1984 | 1997 | 2004 | 2008 | 2013 | 2016 |
|---|---|---|---|---|---|---|---|---|
| Scale parameter | 30 | 20 | 35 | 30 | 15 | 20 | 20 | 20 |
| RMSE (m2) | 112.91 | 69.96 | 110.04 | 120.64 | 80.57 | 84.26 | 97.02 | 46.38 |
| MBE (m2) | 14.07 | 15.07 | −6.59 | 0.78 | 32.78 | 15.74 | −9.96 | −6.37 |
| ED2 | 0.59 | 0.35 | 0.43 | 0.49 | 0.51 | 0.47 | 0.42 | 0.35 |
| Year | 1956 | 1977 | 1984 | 1997 | 2004 | 2008 | 2013 | 2016 |
|---|---|---|---|---|---|---|---|---|
| Separability (J) | ||||||||
| Brightness | 0.92 | 1.24 | 0.88 | 0.97 | 1.04 | 1.13 | 0.53 | 0.86 |
| GLCM Homogeneity | 0.03 | 0.87 | 0.47 | 0.29 | 0.34 | 0.56 | 0.45 | 0.8 |
| GLCM Contrast | 0.25 | 0.77 | 0.14 | 0.29 | 0.94 | 0.26 | 1.01 | 0.79 |
| GLCM Entropy | 0.06 | 0.38 | 0.38 | 0.24 | 0 | 0.41 | 1.24 | 0.56 |
| GLCM Mean | 0.91 | 1.32 | 0.89 | 1 | 1.03 | 1.14 | 0.57 | 0.86 |
| GLCM SD | 0.06 | 0.35 | 0.2 | 0.3 | 0.06 | 0.45 | 0.84 | 0.26 |
| GLCM Correlation | 0.01 | 0.06 | 0.06 | 0.2 | 0.35 | 1.93 | 0.94 | 0.59 |
| Area | 0.55 | 0.15 | 0.92 | 0.04 | 0.38 | 0.7 | 1.08 | 0.19 |
| Border Length | 0.7 | 0.54 | 0.27 | 0.35 | 0.06 | 1.37 | 0.79 | 0.07 |
| Border Index | 0.79 | 0.69 | 0.51 | 0.5 | 0.17 | 1.7 | 0.28 | 0.32 |
| Compactness | 0.73 | 0.63 | 0.46 | 0.51 | 0.5 | 1.38 | 0.29 | 0.46 |
| Density | 0.32 | 1.11 | 0.52 | 0.82 | 0.86 | 0.64 | 1.42 | 0.85 |
| Roundness | 0.63 | 0.35 | 0.25 | 0.36 | 0.22 | 1.44 | 0.29 | 0.2 |
| Shape Index | 0.82 | 0.7 | 0.52 | 0.57 | 0.25 | 1.74 | 0.22 | 0.4 |
| Year | 1956 | 1977 | 1984 | 1997 | ||||||||
| Class | Z | S | sum | Z | S | sum | Z | S | sum | Z | S | sum |
| Z | 52 | 5 | 57 | 47 | 0 | 47 | 59 | 2 | 61 | 57 | 1 | 58 |
| S | 7 | 55 | 62 | 13 | 60 | 73 | 0 | 58 | 58 | 3 | 59 | 62 |
| Uncl. | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Sum | 60 | 60 | 60 | 60 | 60 | 60 | 60 | 60 | ||||
| Prod. | 0.86 | 0.92 | 0.78 | 1 | 0.98 | 0.97 | 0.95 | 0.98 | ||||
| User | 0.92 | 0.88 | 1 | 0.82 | 0.97 | 1 | 0.98 | 0.95 | ||||
| Total | ||||||||||||
| KIA-c | 0.74 | 0.84 | 0.64 | 1 | 0.95 | 0.94 | 0.9 | 0.96 | ||||
| OA | 0.89 | 0.89 | 0.89 | 0.89 | 0.97 | 0.97 | 0.97 | 0.97 | ||||
| KIA | 0.79 | 0.79 | 0.78 | 0.78 | 0.95 | 0.95 | 0.93 | 0.93 | ||||
| Year | 2004 | 2008 | 2013 | 2016 | ||||||||
| Class | Z | S | sum | Z | S | sum | Z | S | sum | Z | S | sum |
| Z | 59 | 1 | 60 | 55 | 0 | 55 | 57 | 1 | 58 | 58 | 1 | 59 |
| S | 1 | 59 | 60 | 5 | 60 | 65 | 3 | 59 | 62 | 2 | 59 | 61 |
| Uncl. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sum | 60 | 60 | 60 | 60 | 60 | 60 | 60 | 60 | ||||
| Prod. | 0.98 | 0.99 | 0.92 | 1 | 0.95 | 0.99 | 0.96 | 0.99 | ||||
| User | 0.99 | 0.98 | 0.99 | 0.92 | 0.99 | 0.95 | 0.99 | 0.96 | ||||
| Total | ||||||||||||
| KIA-c | 0.95 | 0.98 | 0.84 | 0.99 | 0.9 | 0.98 | 0.92 | 0.99 | ||||
| OA | 0.98 | 0.98 | 0.96 | 0.96 | 0.97 | 0.97 | 0.98 | 0.96 | ||||
| KIA | 0.97 | 0.97 | 0.91 | 0.91 | 0.94 | 0.94 | 0.96 | 0.99 | ||||
| Year | 1956 | 1977 | 1984 | 1997 | 2004 | 2008 | 2013 | 2016 |
|---|---|---|---|---|---|---|---|---|
| Number of shrubs | 2055 | 2625 | 2434 | 2345 | 2071 | 2078 | 1999 | 1883 |
| Average area (m2) | 82.62 | 78.32 | 87.98 | 76.51 | 93.78 | 100.57 | 99.5 | 111.31 |
| SD area (m2) | 67.65 | 67.23 | 79.45 | 66.91 | 76.77 | 82.28 | 77.84 | 83.22 |
| Minimum area (m2) | 8 | 5 | 7 | 4 | 6 | 7 | 8 | 6 |
| Maximum area (m2) | 525 | 570 | 643 | 701 | 586 | 742 | 678 | 658 |
| Total cover area (m2) | 152,932 | 208,702 | 209,763 | 177,706 | 223,322 | 208,889 | 198,708 | 212,394 |
| Round shape index | 1.32 | 1.52 | 1.47 | 1.53 | 1.71 | 1.76 | 2.04 | 1.94 |
| SD round shape index | 0.16 | 0.26 | 0.26 | 0.27 | 0.36 | 0.38 | 0.47 | 0.46 |
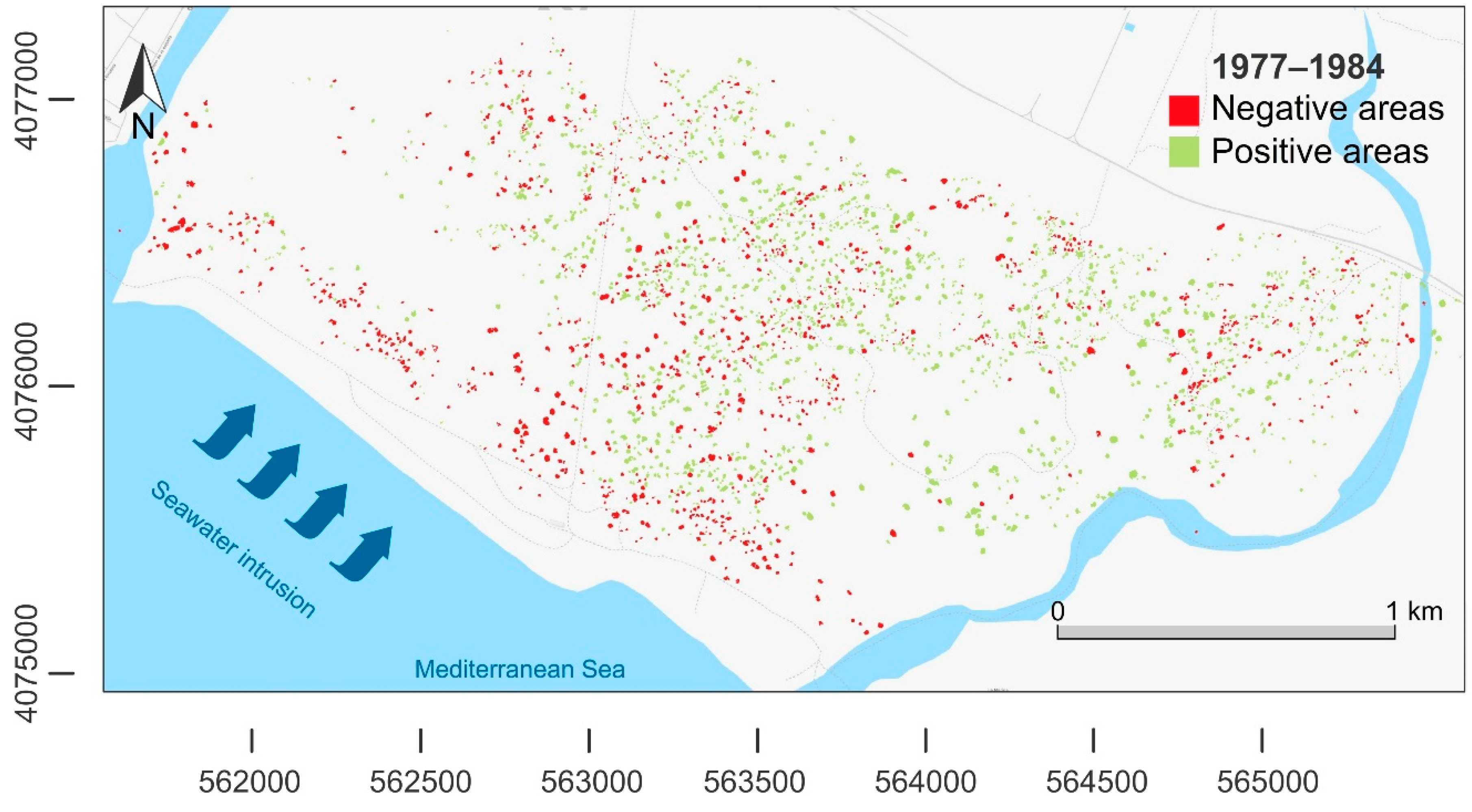
| Difference of Area Between 1956–1977 | Difference of area Between 1977–1984 | Difference of Area Between 1984–1997 | Difference of area Between 1997–2004 | Difference of area Between 2004–2008 | Difference of area Between 2008–2013 | Difference of area Between 2013–2016 | |
|---|---|---|---|---|---|---|---|
| Negative area (m2) | −27,797 | −18,917 | −56,838 | −25,968 | −32,426 | −34,902 | −29,684 |
| Positive area (m2) | 42,753 | 69,207 | 31,594 | 84,898 | 60,870 | 44,147 | 45,101 |
| Balance of areas (m2) | 14,956 | 50,290 | −25,244 | 58,930 | 28,444 | 9245 | 15,417 |
| Negative frequency (n) | 752 | 903 | 1423 | 824 | 726 | 861 | 893 |
| Positive frequency (n) | 1158 | 1650 | 871 | 1405 | 1319 | 1150 | 978 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guirado, E.; Blanco-Sacristán, J.; Rigol-Sánchez, J.P.; Alcaraz-Segura, D.; Cabello, J. A Multi-Temporal Object-Based Image Analysis to Detect Long-Lived Shrub Cover Changes in Drylands. Remote Sens. 2019, 11, 2649. https://doi.org/10.3390/rs11222649
Guirado E, Blanco-Sacristán J, Rigol-Sánchez JP, Alcaraz-Segura D, Cabello J. A Multi-Temporal Object-Based Image Analysis to Detect Long-Lived Shrub Cover Changes in Drylands. Remote Sensing. 2019; 11(22):2649. https://doi.org/10.3390/rs11222649
Chicago/Turabian StyleGuirado, Emilio, Javier Blanco-Sacristán, Juan Pedro Rigol-Sánchez, Domingo Alcaraz-Segura, and Javier Cabello. 2019. "A Multi-Temporal Object-Based Image Analysis to Detect Long-Lived Shrub Cover Changes in Drylands" Remote Sensing 11, no. 22: 2649. https://doi.org/10.3390/rs11222649
APA StyleGuirado, E., Blanco-Sacristán, J., Rigol-Sánchez, J. P., Alcaraz-Segura, D., & Cabello, J. (2019). A Multi-Temporal Object-Based Image Analysis to Detect Long-Lived Shrub Cover Changes in Drylands. Remote Sensing, 11(22), 2649. https://doi.org/10.3390/rs11222649