Functional Phenology of a Texas Post Oak Savanna from a CHRIS PROBA Time Series


Abstract
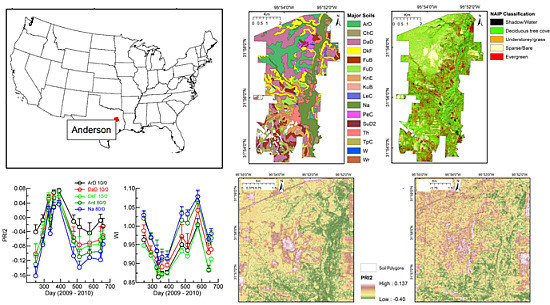
1. Introduction
- (1)
- To characterize the seasonal phenology of deciduous post oak savanna and encroaching evergreen shrubs with functional VIs derived from CHRIS PROBA imagery;
- (2)
- To explore the sensitivity of VIs for distinguishing differences in photosynthetic capacity and water status between deciduous savanna vegetation and evergreen shrubs;
- (3)
- To illustrate the utility and sensitivity of combinations of VIs for describing differences in vegetation function across soil-vegetation associations in a post oak savanna.
2. Materials and Methods
2.1. Study Area
2.2. Remote Sensing Data and Processing
2.2.1. Aerial Photography
2.2.2. CHRIS PROBA Data
2.2.3. Vegetation Indices
2.3. Analysis
2.3.1. Fishnet Polygon Extraction of Tree Cover Density and VI Profiles
2.3.2. Combined Indicators of Vegetation Function
3. Results
3.1. Overall VI Profiles
3.2. Response of VIs to Evergreen Density
3.3. Responses within Tree Density Levels among Soil Types
3.4. Sensitivity of VIs to Spatial Variability in Leaf-on and Leaf-off Periods
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McPherson, G.R. Ecology and Management of North American Savannas; University of Arizona Press: Tuscon, AZ, USA, 1997; 208p. [Google Scholar]
- Srinath, I.; Millington, A.C. Evaluating the potential of the original Texas Land Survey for mapping historical land and vegetation cover. Land 2016, 5, 4. [Google Scholar] [CrossRef]
- Ludeke, K.; German, D.; Scott, J. Texas Vegetation Classification Project: Interpretive Booklet for Phase II. Texas Parks and Wildlife Department and Texas Natural Resources Information System. 2009. Available online: https://morap.missouri.edu/wp-content/uploads/2019/02/Texas_Vegetation_Classification_Phase_2_Interpretive_Booklet.pdf (accessed on 2 December 2009).
- Gould, F.W. Texas Plants. A Checklist and Ecological Summary; Texas Agriculture Experiment Station, Texas A&M University: College Station, TX, USA, 1962. [Google Scholar]
- Singhurst, J.R.; Cathy, J.C.; Prochaska, D.; Haucke, H.; Kroh, G.C.; Holmes, W. The vascular flora of Gus Engeling Wildlife Management Area, Anderson County, Texas. Southeast. Nat. 2009, 2, 347–368. [Google Scholar] [CrossRef]
- Smith, S. Eastern Red-Cedar: Positives, Negatives and Management; Samuel Roberts Noble Foundation: Ardmore, OK, USA, 2011; 8p. [Google Scholar]
- Barnsley, M.J.; Settle, J.J.; Cutter, M.A.; Lobb, D.R.; Teston, F. The PROBA/CHRIS mission: A low-cost smallsat for hyperspectral multiangle observations of the earth surface and atmosphere. IEEE Trans. Geosci. Remote Sens. 2004, 42, 1512–1520. [Google Scholar] [CrossRef]
- Barnsley, M.J.; Lewis, P.; O’Dwyer, S.; Disney, M.I.; Hobson, P.; Cutter, M.; Lobb, D. On the potential of CHRIS/PROBA for estimating vegetation canopy properties from space. Remote Sens. Rev. 2000, 19, 171–189. [Google Scholar] [CrossRef]
- Green, R.O.; Pavri, B.E.; Chrien, T.G. On-orbit radiometric and spectral calibration characteristics of EO-1 Hyperion derived with an underflight of AVIRIS and in situ measurements at Salar de Arizaro, Argentina. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1194–1203. [Google Scholar] [CrossRef]
- Pearlman, J.S.; Barry, P.S.; Segal, C.C.; Shepanski, J.; Beiso, D.; Carman, S.L. Hyperion, a space-based imaging spectrometer. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1160–1173. [Google Scholar] [CrossRef]
- Middleton, E.M.; Ungar, S.G.; Mandl, D.; Ong, L.; Frye, S.; Campbell, P.E.; Landis, D.R.; Young, J.P.; Pollack, N.H. The Earth Observing One (EO-1) satellite mission: Over a decade in space. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. (JSTARS) 2013, 6, 243–256. [Google Scholar] [CrossRef]
- Duca, R.; Del Frate, F. Hyperspectral and multiangle CHRIS-PROBA images for the generation of land cover maps. IEEE Trans. Geosci. Remote Sens. 2008, 46, 2857–2866. [Google Scholar] [CrossRef]
- Stagakis, S.; Markos, N.; Sykioti, O.; Kyparissis, A. Monitoring canopy biophysical and biochemical parameters in ecosystem scale using satellite hyperspectral imagery: An application on a Phlomis fruticosa Mediterranean ecosystem using multiangular CHRIS/PROBA observations. Remote Sens. Environ. 2010, 114, 977–994. [Google Scholar] [CrossRef]
- Mousivand, A.; Menenti, M.; Gorte, B.; Verhoef, W. Multi-temporal, multi-sensor retrieval of terrestrial vegetation properties from spectral-directional radiometric data. Remote Sens. Environ. 2015, 158, 311–330. [Google Scholar] [CrossRef]
- Verrelst, J.; Romijn, E.; Kooistra, L. Mapping vegetation density in a heterogeneous river floodplain ecosystem using pointable CHRIS/PROBA data. Remote Sens. 2012, 4, 2866–2889. [Google Scholar] [CrossRef]
- García Millán, V.E.; Sánchez-Azofeifa, A.; Málvarez García, G.C.; Rivard, B. Quantifying tropical dry forest succession in the Americas using CHRIS/PROBA. Remote Sens. Environ. 2014, 144, 120–136. [Google Scholar] [CrossRef]
- Torbick, N.; Corbiere, M.A. Multiscale mapping assessment of Lake Champlain cyanobacterial harmful algal blooms. Int. J. Environ. Res. Public Health 2015, 12, 11560–11578. [Google Scholar] [CrossRef] [PubMed]
- Gürsoy, Ö.; Birdal, A.C.; Özyonar, F.; Kasaka, E. Determining and monitoring the water quality of Kizilirmak River of Turkey: First results. Int. Arch. Photogram. Remote Sens. Spat. Inf. Sci. 2015, 40, 1469–1474. [Google Scholar] [CrossRef]
- Tian, Y.; Guo, Z.Q.; Qiao, Y.C.; Lei, X.; Xie, F. Remote sensing of water quality monitoring in Guanting Reservoir. Shengtai Xuebao 2015, 35, 2217–2226. [Google Scholar] [CrossRef][Green Version]
- Latorre-Carmona, P.; Knyazikhin, Y.; Alonso, L.; Moreno, J.F.; Pla, F.; Yan, Y. On hyperspectral remote sensing of leaf biophysical constituents: Decoupling vegetation structure and leaf optics using CHRIS-PROBA data over crops in barrax. IEEE Geosci. Remote Sens. Lett. 2014, 11, 1579–1583. [Google Scholar] [CrossRef]
- Laurent, V.C.E.; Verhoef, W.; Clevers, J.G.P.W.; Schaepman, M.E. Inversion of a coupled canopy-atmosphere model using multi-angular top-of-atmosphere radiance data: A forest case study. Remote Sens. Environ. 2011, 115, 2603–2612. [Google Scholar] [CrossRef]
- Hilker, T.; Coops, N.C.; Hall, F.G.; Nichol, C.J.; Lyapustin, A.; Black, T.A.; Wulder, M.A.; Leuning, R.; Barr, A.; Hollinger, D.Y.; et al. Inferring terrestrial photosynthetic light use efficiency of temperate ecosystems from space. J. Geophys. Res. Biogeosci. 2011, 116, G03014. [Google Scholar] [CrossRef]
- Carmona, F.; Rivas, R.; Fonnegra, D.C. Vegetation index to estimate chlorophyll content from multispectral remote sensing data. Eur. J. Remote Sens. 2015, 48, 319–326. [Google Scholar] [CrossRef]
- Agapiou, A.; Hadjimitsis, D.G.; Alexakis, D.D. Evaluation of Broadband and Narrowband Vegetation Indices for the Identification of Archaeological Crop Marks. Remote Sens. 2012, 4, 3892–3919. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of chlorophyll a, chlorophyll b, chlorophyll a+b, and total carotenoid content in eucalyptus leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical Properties and Nondestructive Estimation of Anthocyanin Content in Plant Leaves. Photochem. Photobiol. 2001, 71, 38–45. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing Carotenoid Content in Plant Leaves with Reflectance Spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Filella, I.; Amaro, T.; Araus, J.L.; Peñuelas, J. Relationship between photosynthetic radiation-use efficiency of barley canopies and the photochemical reflectance index (PRI). Physiol. Plant. 1996, 96, 211–216. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Nondestructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Guerschman, J.-P.; Hill, M.J.; Barrett, D.J.; Renzullo, L.; Marks, A.; Botha, E. Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and soil in mixed tree-grass vegetation using the EO-1 and MODIS sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
- Hill, M.J.; Renzullo, L.J.; Guerschman, J.P.; Marks, A.S.; Barrett, D.J. Use of vegetation index “fingerprints” from Hyperion data to characterize vegetation states within land cover/land use types in an Australian tropical savanna. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. (JSTARS) 2013, 6, 309–319. [Google Scholar] [CrossRef]
- Hill, M.J. Vegetation index suites as indicators of vegetation state in grassland and savanna: An analysis with simulated SENTINEL 2 data for a North American transect. Remote Sens. Environ. 2013, 137, 94–111. [Google Scholar] [CrossRef]
- Haucke, H.; Prochaska, D. Management Plan for Gus Engeling Research and Demonstration Area; Texas Parks and Wildlife Department: Austin, TX, USA, 1998; 22p.
- Alonso, L.; Gómez-Chova, L.; Moreno, J.; Guanter, L.; Brockmann, C.; Fomferra, N.; Quast, R.; Regner, P. CHRIS/PROBA Toolbox for hyperspectral and multiangular data exploitations. In Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium, University of Cape Town, Cape Town, South Africa, 12–17 July 2009; Volume 2, p. II-202. [Google Scholar] [CrossRef]
- Fomferra, N.; Brockmann, C. BEAM—The ENVISAT MERIS and AATSR toolbox. In Proceedings of the MERIS (A)ATSR Workshop 2005, Frascati, Italy, 26–30 September 2005; 3p. [Google Scholar]
- Guanter, L.; Alonso, L.; Gomez-Chova, L.; Moreno, J. PROBA/CHRIS Atmospheric Correction Module Algorithm Theoretical Basis Document; Contract No. 20442/07/I-LG; ESA ESRIN: Frascati, Italy, 2008; 27p. [Google Scholar]
- Berk, A.; Anderson, G.P.; Acharya, P.K.; Hoke, M.L.; Chetwynd, J.H.; Bernstein, L.S.; Shettle, E.P.; Matthew, M.W.; Adler-Golden, S.M. MODTRAN4 Version 3 Revision 1 User’s Manual; Technical Report; Air Force Research Laboratory: Hanscom Air Force Base, MA, USA, 2003; 91p.
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Cutter, M. CHRIS Data Format; Surrey Satellite Technology: Guildford, UK, 2008; 37p. [Google Scholar]
- Moy, A.; Le, S.; Verhoeven, A. Different strategies for photoprotection during autumn senescence in maple and oak. Physiol. Plant. 2015, 155, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; O’Keefe, J.; Holbrook, N.M.; Field, T.S. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA. Ecol. Res. 2003, 18, 677–694. [Google Scholar] [CrossRef]
- Keskitalo, J.; Bergquist, G.; Gardeström, P.; Jansson, S. A cellular timetable of autumn senescence. Plant Phys. 2005, 139, 1635–1648. [Google Scholar] [CrossRef] [PubMed]
- García-Plazaola, J.I.; Hernández, A.; Becerril, J.M. Antioxidant and pigment composition during autumnal leaf senescence in woody deciduous species differing in their ecological traits. Plant Biol. 2003, 5, 557–566. [Google Scholar] [CrossRef]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Phys. 1990, 7, 227–238. [Google Scholar] [CrossRef]
- Donovan, L.A.; West, J.B.; McLeod, K.W. Quercus species differ in water and nutrient characteristics in a resource-limited fall-line sandhill habitat. Tree Phys. 2000, 20, 929–936. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Collins, S.L. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology 2012, 93, 697–703. [Google Scholar] [CrossRef]
- Hinckley, T.M.; Dougherty, P.M.; Lassoie, J.P.; Roberts, J.E.; Teskey, R.O. A severe drought: Impact on tree growth, phenology, net photosynthetic rate and water relations. Am. Midland Nat. 1979, 102, 307–316. Available online: https://www.jstor.org/stable/2424658 (accessed on 15 August 2019). [CrossRef]
- Ginter-Whitehouse, D.L.; Hinckley, T.M.; Pallardy, S.G. Spatial and temporal aspects of water relations of three tree species with different vascular anatomy. For. Sci. 1983, 29, 317–329. [Google Scholar]
- Caterina, G.L.; Will, R.E.; Turton, D.J.; Wilson, D.S.; Zou, C.B. Water use of Juniperus virginiana trees encroached into mesic prairies in Oklahoma, USA. Ecohydrology 2014, 7, 1124–1134. [Google Scholar] [CrossRef]
- Eggemeyer, K.D.; Awada, T.; Harvey, F.E.; Wedin, D.A.; Zhou, X.; Zanner, C.W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C4 grasses in a semiarid grassland. Tree Phys. 2009, 29, 157–169. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, R.D.; Hallgren, S.W.; Lynch, T.B.; Burton, J.A.; Palmer, M.W. Long-term directional changes in upland Quercus forests throughout Oklahoma, USA. J. Veg. Sci. 2010, 21, 606–615. [Google Scholar] [CrossRef]
- Chastain, R.A., Jr.; Struckhoff, M.A.; He, H.S.; Larsen, D.R. Mapping Vegetation Communities Using Statistical Data Fusion in the Ozark National Scenic Riverways, Missouri, USA. Photogramm. Eng. Remote Sens. 2008, 74, 247–264. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Brudvig, L.A.; Mabry, C.M.; Evans, C.W.; Karnitz, H.M. Defining reference information for restoring ecologically rare tallgrass oak savannas in the Midwestern United States. J. For. 2005, 103, 345–350. [Google Scholar] [CrossRef]
- Grundel, R.; Pavlovic, N.B. Using conservation value to assess land restoration and management alternatives across a degraded oak savanna landscape. J. Appl. Ecol. 2008, 45, 315–324. [Google Scholar] [CrossRef]
- Chen, Q.; Baldocchi, D.; Gong, P.; Kelly, M. Isolating individual trees in a savanna woodland using small footprint LiDAR data. Photogram. Eng. Remote Sens. 2006, 72, 923–932. [Google Scholar] [CrossRef]
- Asner, G.P.; Jones, M.O.; Martin, R.E.; Knapp, D.E.; Hughes, R.F. Remote sensing of native and invasive species in Hawaiian forests. Remote Sens. Environ. 2008, 112, 1912–1926. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar] [CrossRef]
- Feilhauer, H.; Asner, G.P.; Martin, R.E. Multi-method ensemble selection of spectral bands related to leaf biochemistry. Remote Sens. Environ. 2015, 164, 57–65. [Google Scholar] [CrossRef]
- Chadwick, K.; Asner, G. Organismic-scale remote sensing of canopy foliar traits in lowland tropical forests. Remote Sens. 2016, 8, 87. [Google Scholar] [CrossRef]
- McManus, K.; Asner, G.; Martin, R.; Dexter, K.; Kress, W.; Field, C. Phylogenetic structure of foliar spectral traits in tropical forest canopies. Remote Sens. 2016, 8, 196. [Google Scholar] [CrossRef]
- Mitchell, R.; Cathey, J.C.; Drabbert, B.; Prochaska, D.; Dupree, S.; Sosebee, R. Managing Yaupon with fire and herbicodes in the Texas post oak savannah. Rangelands 2005, 27, 17–19. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Dominant ESD Description | Soil Description (Code) | % Area | ASW 100 cm (%) | 2 Land Cover (%) POW; SG; HF; SFHF; JM |
|---|---|---|---|---|
| Clayey Upland | Kirvin–Sacul association, sloping; (KnE) Sacul fine sandy loam, 1 to 5 percent slopes (SaC) | 4.8 0.4 | 12.3 9.8 | 94; 0; 0; 0; 0; 0 |
| Loamy Upland | Elrose fine sandy loam, 1 to 3 percent slopes (EiB); Kullit fine sandy loam, 1 to 3 percent slopes (KuB) | 0.2 0.6 | 13.1 13.0 | 71; 22; 0; 0; 4 |
| Northern Sandy Loamy Upland | Rentzel fine sand, 0 to 5 percent slopes (ChC); Lilbert loamy fine sand, 0 to 3 percent slopes (FuB); Lilbert loamy fine sand, 3 to 8 percent slopes (FuD); Larue loamy fine sand, 1 to 3 percent slopes (LaB); Rentzel loamy fine sand, 0 to 5 percent slopes (LeC); Trep loamy fine sand, 1 to 5 percent slopes (TpC); | 2.7 7.3 2.6 0.3 0.3 4.8 | 8.6 9.6 9.2 10.0 8.6 10.0 | 85; 8; 0; 0; 3 |
| Northern Deep Sandy Upland | Darco fine sand, 1 to 8 percent slopes (DaD); Darco, Kirvin, and Tenaha soils, sloping (DkF) | 28.5 8.6 | 7.8 9.8 | 89; 7; 0; 0; 2 |
| Very Deep Sandy Upland | Tonkawa fine sand, 1 to 8 percent slopes (ArD) | 7.3 | 6.0 | 66; 26; 0; 0; 3 |
| Terrace | Annona fine sandy loam, 1 to 5 percent slopes (SsC); Annona soils, 3 to 10 percent slopes, eroded (SuD2) | 0.3 0.4 | 14.6 14.6 | 95; 0; 3; 0; 0 |
| Stream Bottomland | Thenas fine sandy loam (Th) | 4.4 | 13.8 | 9; 0; 76; 15; 0 |
| Sandy Bottomland | Naconiche loamy fine sand, 0 to 5 percent slopes (PeC) | 1.1 | 9.7 | 99; 0; 0; 0; 0 |
| Loamy Bottomland | Nahatche and Pluck soils (Na) | 21.9 | 16.1 | 3; 0; 43; 53; 0 |
| Land Cover Type | Species |
|---|---|
| Post oak motte and woodland (POW) | Overstory: Quercus stellata, Q. marilandica, Q. nigra, Q. falcata, Q. incana, Q. fusiformis, Ulmus crassifolia, Celtis laevigata, Carya texana, Mid-story: Prosopis spp., Diospyros virginiana, Ilex vomitoria, I. decidua, U. alata, Sideroxylon lanuginosum, Callicarpa americana, Juniperus virginiana |
| Savanna grassland (SG) | Herbaceous: Shizachyrium scoparium, Sorghastrum nutans, Bothriochloa saccharoides, Stipa leucotricha, Sporobolus asper var. asper, Paspalum plicatulum Scattered: Q. stellata, Q. marilandica, Q. falcata, I. vomitoria, J. virginiana |
| Floodplain hardwood forest (HF) | Overstory: Fraxinus americana, Q. stellata, F. pennsylvanica, U. crassifolia, U. americana Mid-story: C. laevigata, Salix spp. |
| Seasonally flooded hardwood forest (SHHF) | Overstory: Q. marilandica, U. americana, Carya illinoinensis, F. pennsylvanica, Q. phellos, Q. lyrate, Liquidambar styraciflua |
| Juniper or mesquite shrubland/woodland (JM) | J. virginiana, Prosopis spp., Q. marilandica, Q. virginiana, U. crassifolia, U. alata, C. laevigata, D. texana |
| Successional shrublands (Evergreens) | J. virginiana, I. vomitoria, C. laevigata, U. crassifolia |
| Spatial Resolution | Image Size | Nominal View Angles | Spectral Bands | Spectral Range |
|---|---|---|---|---|
| 34 m pixels at 556 km altitude | 372 × 374 pixels (12.65 × 12.72 km) | +55°, +36°, 0° (nadir), −36°, −55° | 62 bands 9–12 nm width except 20 nm at 930 and 950 nm | Min 406–992 nm Max 415–1003 nm Mid 411–997 nm |
| Date | Day (2009–2010) | Observation Zenith (°), Azimuth (°) | Solar Zenith (°), Azimuth (°) | Local Fly-by Time GMT -5/-6 (hr) | Cloud (%) | Image Used | 1 Leaf Stage |
|---|---|---|---|---|---|---|---|
| 18 June 2009 | 169 | 5.73, 222.82 | 28.0, 100.55 | 11.25 | 32.0 | N (site cloud) | On |
| 9 August 2009 | 221 | 19.60, 136.99 | 35.0, 109.43 | 11.11 | 13.1 | N (site cloud) | On |
| 18 August 2009 | 230 | 15.84, 137.97 | 36.0, 113.95 | 11.12 | 17.4 | N (site cloud) | On |
| 6 September 2009 | 249 | 21.07, 315.08 | 38.0, 127.26 | 11.25 | 0 | Y | On |
| 19 October 2009 | 292 | 8.83, 139.26 | 51.0, 140.89 | 11.09 | 0 | Y | Sen |
| 6 November 2009 | 310 | 5.00, 140.00 | 56.0, 145.64 | 10.11 | 6.6 | N | Sen |
| 24 November 2009 | 328 | 5.00, 140.00 | 60.0, 147.91 | 10.12 | 0 | Y | Off |
| 3 December 2009 | 337 | 5.00, 140.00 | 61.0, 148.18 | 10.13 | 0 | Y | Off |
| 20 December 2009 | 358 | 5.00, 140.00 | 65.0, 145.06 | 10.04 | 0 | N (off site) | Off |
| 21 December 2009 | 359 | 5.00, 140.00 | 63.0, 147.30 | 10.15 | 0 | Y | Off |
| 25 January 2010 | 390 | 9.35, 224.82 | 62.0, 139.70 | 10.07 | 0 | Y | Off |
| 14 April 2010 | 469 | 12.29, 138.53 | 42.0, 113.72 | 10.50 | 0 | Y | On |
| 20 May 2010 | 505 | 2.99, 198.03 | 22.0, 100.50 | 10.50 | 0 | N (missing data) | On |
| 29 May 2010 | 514 | 3.57, 23.84 | 35.0, 97.84 | 10.51 | 0 | Y | On |
| 16 June 2010 | 532 | 9.21, 224.65 | 35.0, 94.62 | 10.51 | 19.1 | N (site cloud) | On |
| 30 July 2010 | 576 | 8.24, 141.47 | 40.0, 99.11 | 10.40 | 10.3 | Y | On |
| 4 September 2010 | 642 | 3.24, 205.52 | 45.0, 114.4 | 10.39 | 0 | Y | On |
| 19 October 2010 | 657 | 10.45, 224.75 | 56.0, 132.99 | 10.37 | 3.2 | Y (site clear) | Sen |
| Narrow Band Vegetation Index 1 | Narrow Band Formula 1 | CHRIS PROBA Equivalent 2 | Context |
|---|---|---|---|
| ARI1 (Anthocyanin Reflectance Index) | Stress, senescence [26] | ||
| ARI2 (Anthocyanin Reflectance Index 2) | Stress, senescence [26] | ||
| CRI (Carotenoid Reflectance Index) | Senescence [27] | ||
| NDVI (Normalized Difference Vegetation Index) | Chlorophyll, photosynthetic capacity [40] | ||
| PRI (Photochemical Reflectance Index) | Photosynthetic efficiency [29] | ||
| PRI2 (Photochemical Reflectance Index 2) | Photosynthetic efficiency [28] | ||
| PSRI (Plant Senescence Reflectance Index) | Senescence, brownness [30] | ||
| WI (plant Water Index) | Canopy water content [31] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, M.J.; Millington, A.; Lemons, R.; New, C. Functional Phenology of a Texas Post Oak Savanna from a CHRIS PROBA Time Series. Remote Sens. 2019, 11, 2388. https://doi.org/10.3390/rs11202388
Hill MJ, Millington A, Lemons R, New C. Functional Phenology of a Texas Post Oak Savanna from a CHRIS PROBA Time Series. Remote Sensing. 2019; 11(20):2388. https://doi.org/10.3390/rs11202388
Chicago/Turabian StyleHill, Michael J., Andrew Millington, Rebecca Lemons, and Cherie New. 2019. "Functional Phenology of a Texas Post Oak Savanna from a CHRIS PROBA Time Series" Remote Sensing 11, no. 20: 2388. https://doi.org/10.3390/rs11202388
APA StyleHill, M. J., Millington, A., Lemons, R., & New, C. (2019). Functional Phenology of a Texas Post Oak Savanna from a CHRIS PROBA Time Series. Remote Sensing, 11(20), 2388. https://doi.org/10.3390/rs11202388