Mapping Mangrove Extent and Change: A Globally Applicable Approach


 ,
,  ,
,  and
and
Abstract

1. Introduction
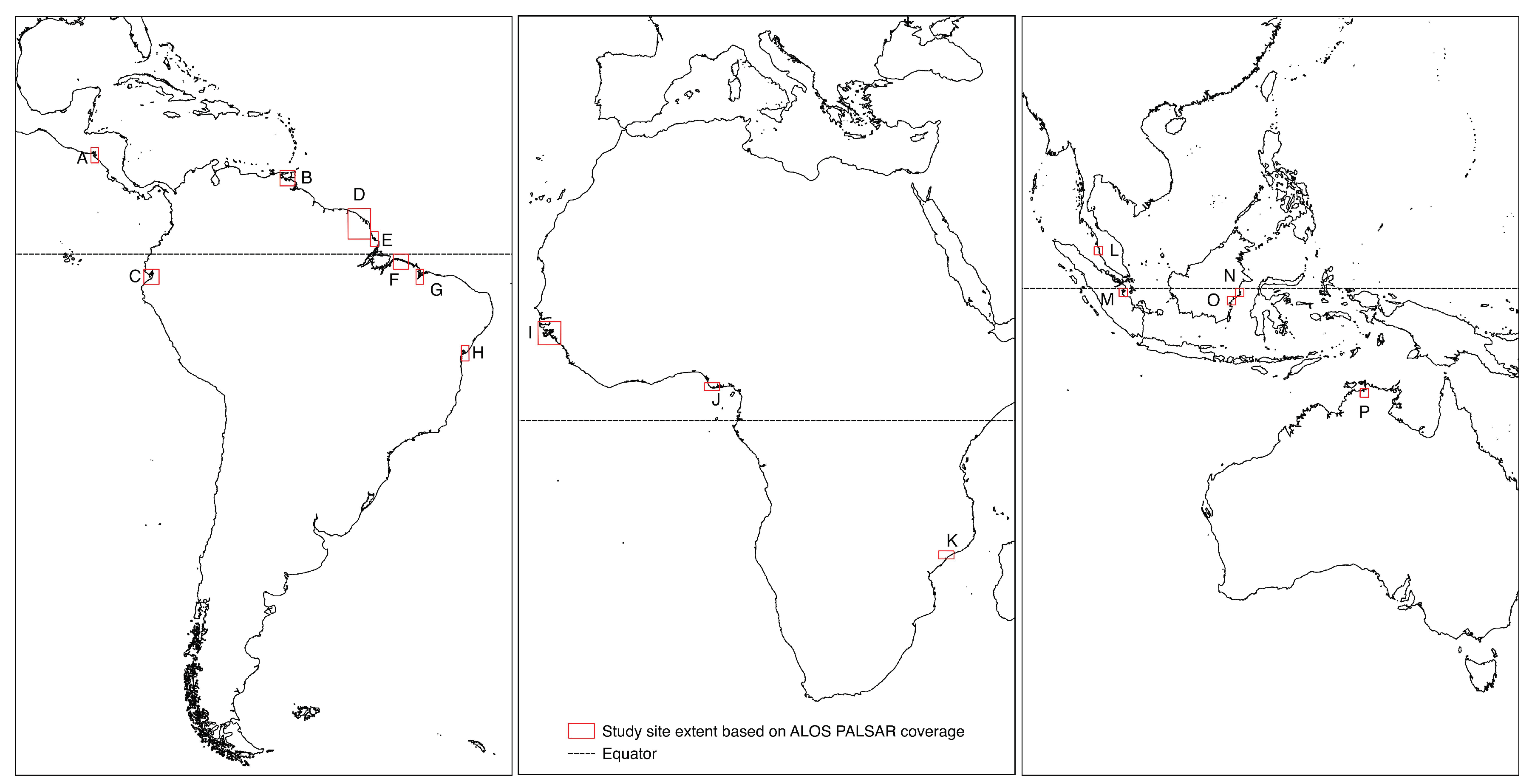
2. Study Sites
2.1. Datasets
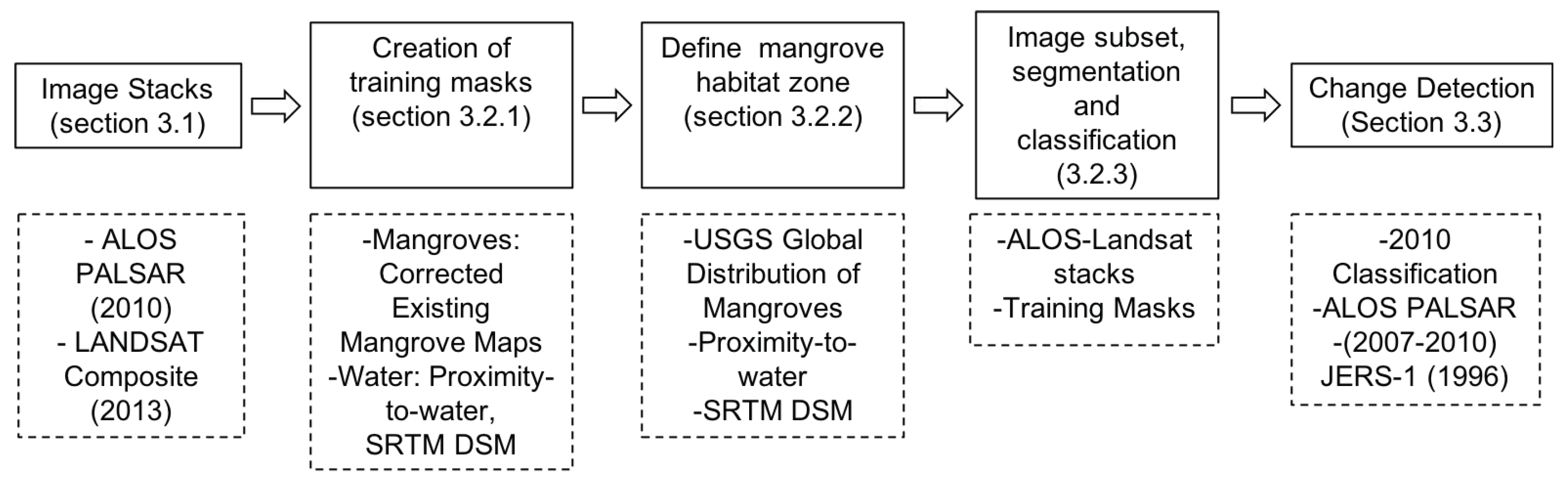
2.2. 2010 Mangrove Baseline Classification
2.2.1. Training Data Mask
2.2.2. Local Mangrove Habitat Regions
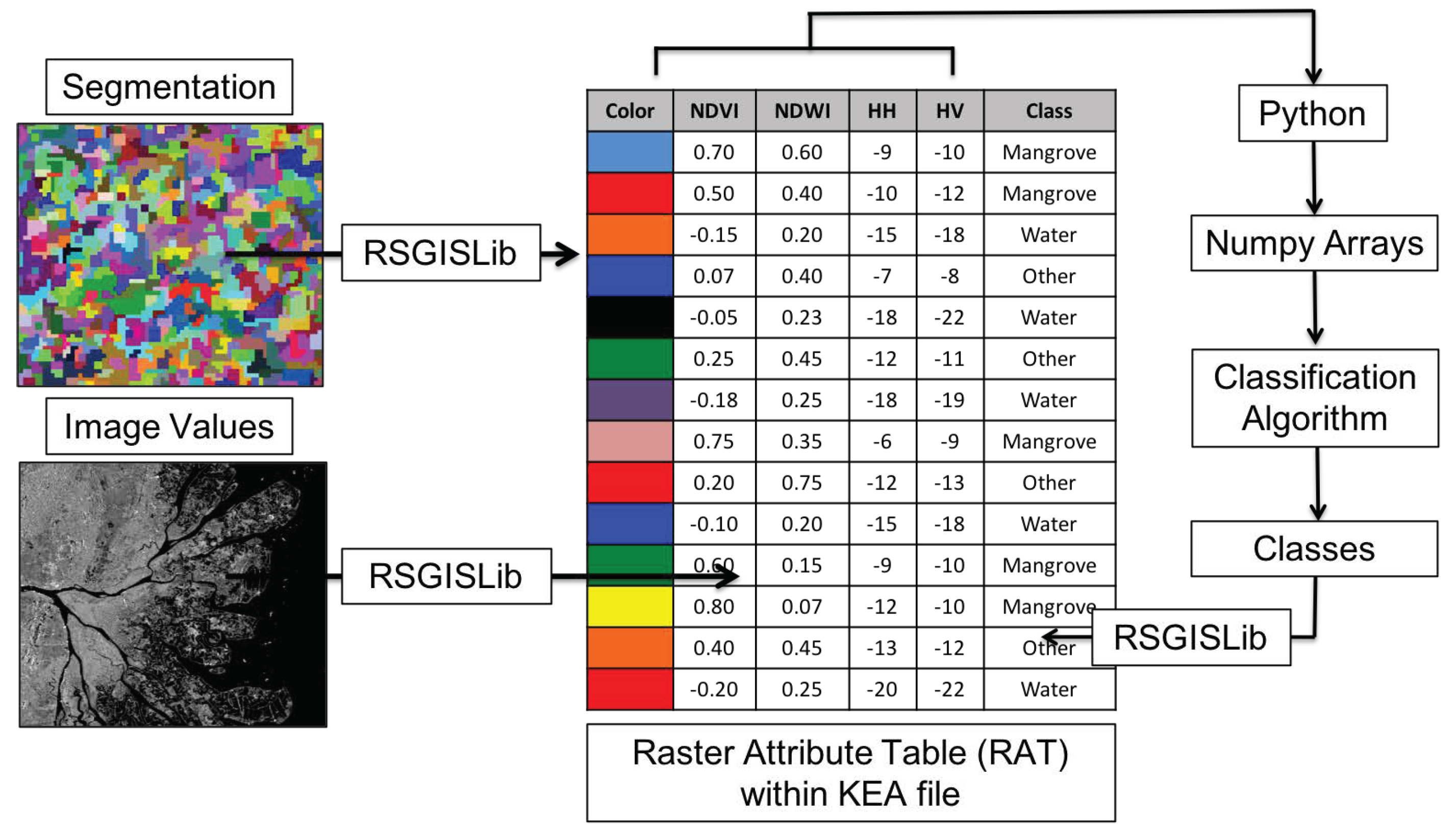
2.2.3. Random Forests Classifier
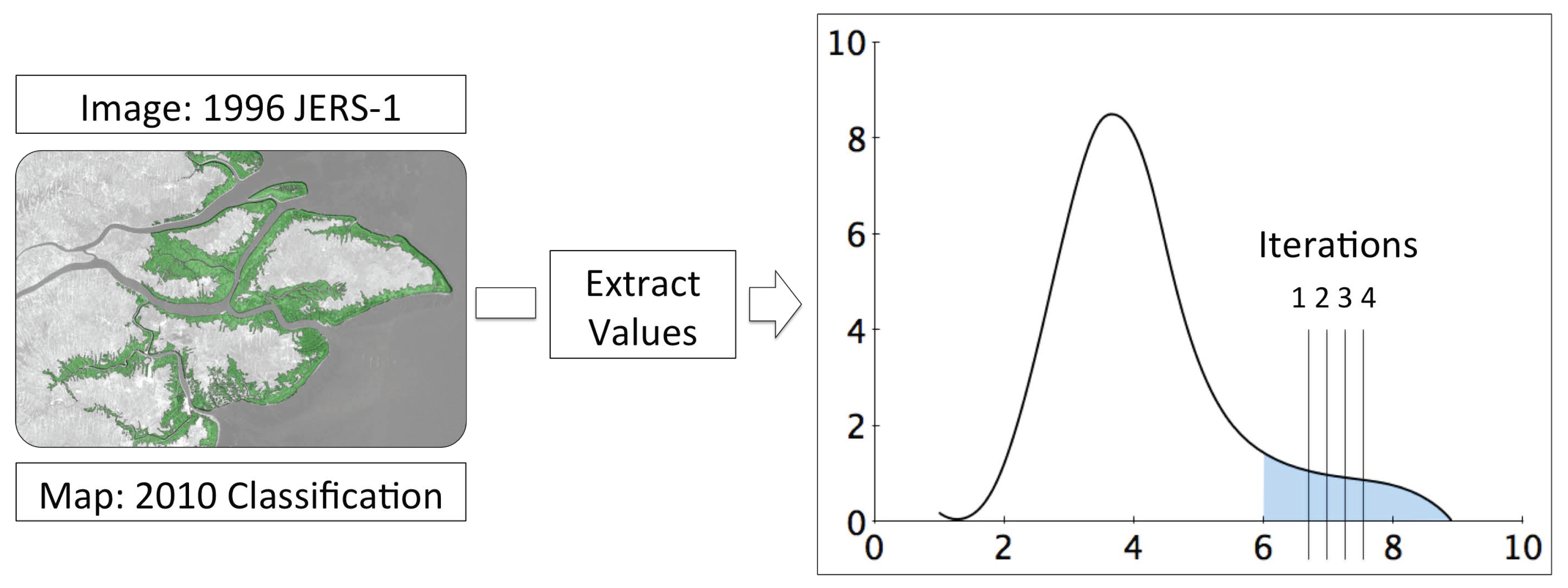
2.3. Change Detection
Annual and Decadal Changes
2.4. Accuracy Assessment
2.4.1. Baseline Accuracy Assessment
2.4.2. Change Detection Accuracy Assessment
- The first approach assessed the accuracy of the output maps, determining their reliability for use within mangrove monitoring and determined whether a detected change was a real-world change in mangrove extent. This is referred to as the “map” accuracy assessment.
- The second approach assessed the performance of the change detection method and its ability to detect differences in time-series imagery, irrespective of the cause of the change between the two images. This is referred to as the “algorithm” accuracy assessment. This differs from the “map” accuracy assessment by not punishing the method for detecting a difference between images, even if it was not a change in mangrove extent.
3. Results
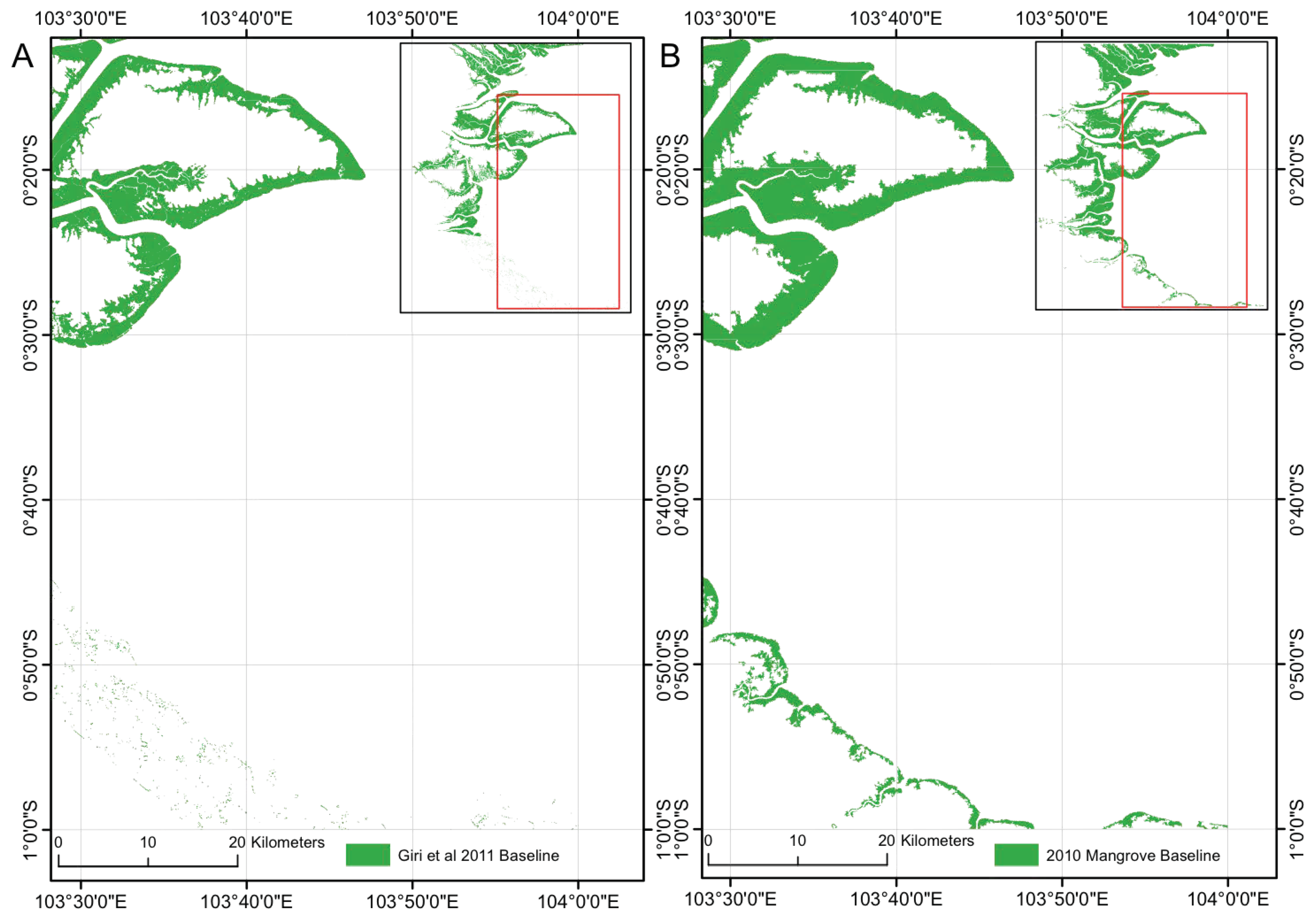
3.1. 2010 Baseline Classification
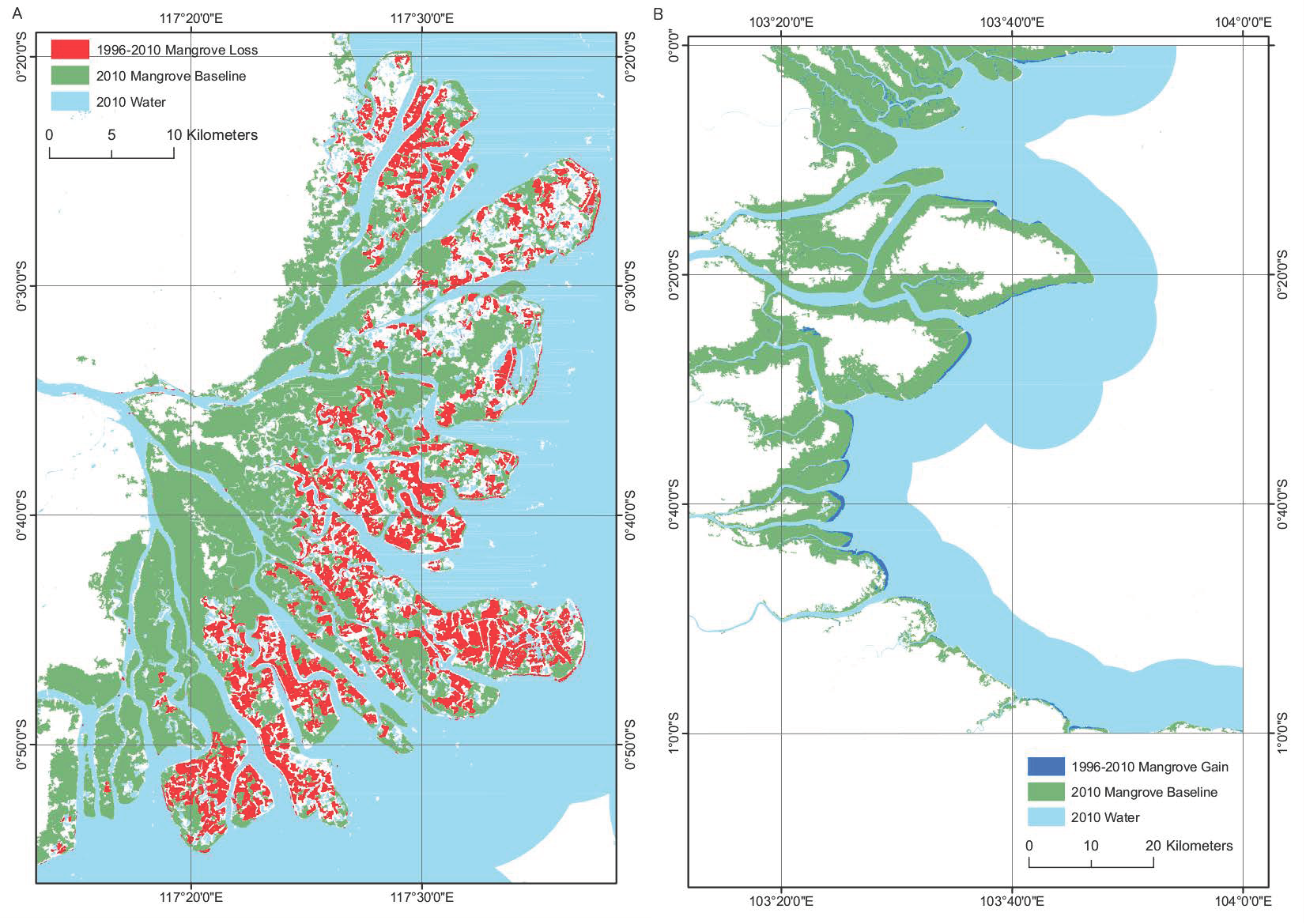
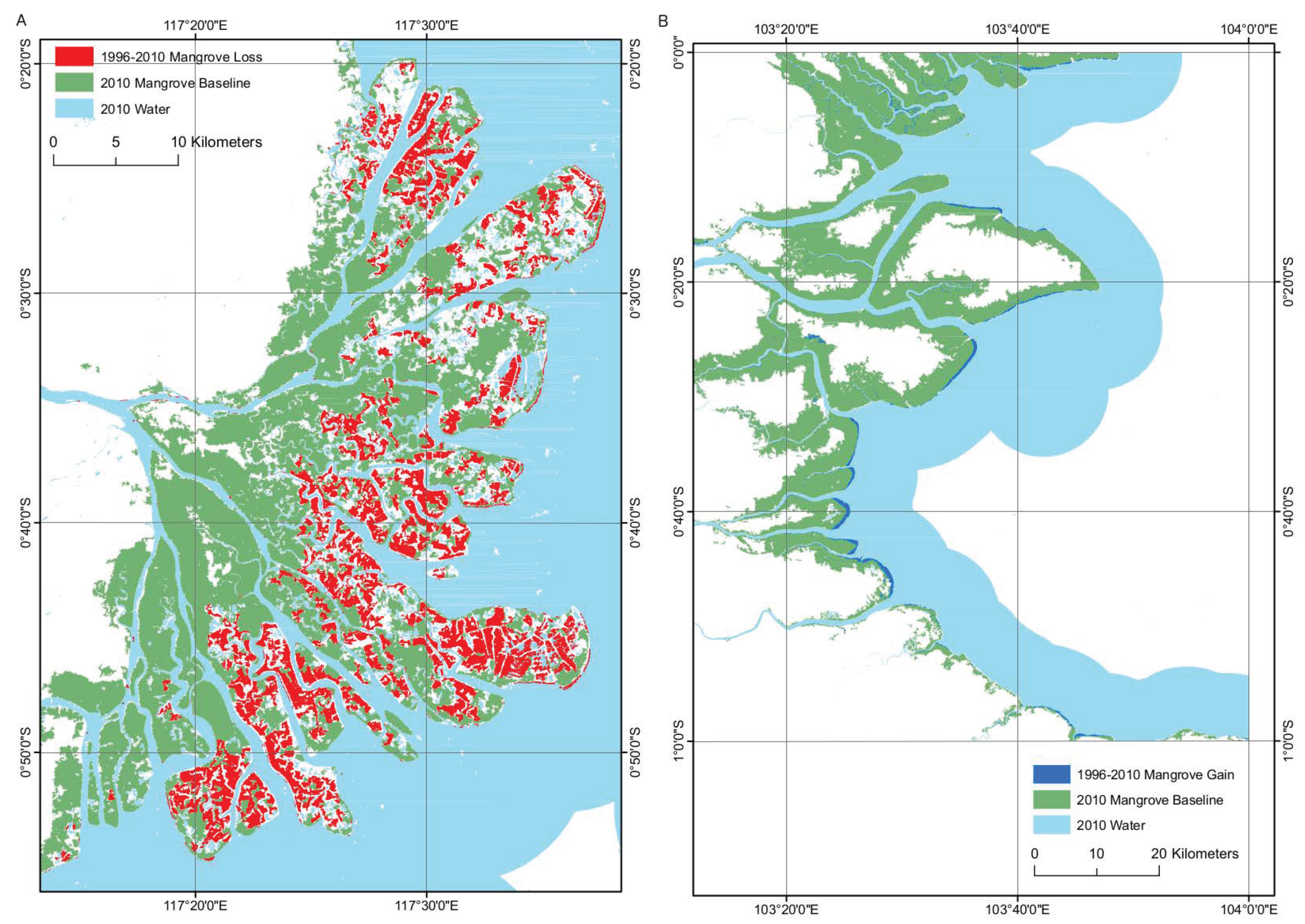
3.2. Change detection
3.2.1. ALOS PALSAR 2007–2010
3.2.2. JERS-1 SAR/ALOS PALSAR 1996–2010
4. Discussion
4.1. Baseline Classification
4.2. Change Detection
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Jennerjahn, T.C.; Ittekkot, V. Relevance of mangroves for the production and deposition of organic matter along tropical continental margins. Naturwissenschaften 2002, 89, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Murray, B.C.; Pendleton, L.; Jenkins, W.A.; Sifleet, S. Green Payments for Blue Carbon: Economic Incentives for Protecting Threatened Coastal Habitats; Nicholas Institute for Environmental Policy Solutions, Duke University: Durham, NC, USA, 2011; NI R 11-04. [Google Scholar]
- Hamilton, S.E.; Friess, D.A. Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat. Clim. Chang. 2018, 8, 240. [Google Scholar] [CrossRef]
- Lucas, R.; Robelo, L.; Fatoyinbo, L.; Rosenqvist, A.; Itoh, T.; Shimada, M.; Simard, M.; Souza-Filho, P.; Thomas, N.; Trettin, C.; et al. Contribution of L-band SAR to Systematic Global Mangrove Monitoring. Mar. Freshw. Res. 2014, 65, 589–603. [Google Scholar] [CrossRef]
- Blaber, S.J.M.; Milton, D.A. Species composition, community structure and zoogeography of fishes of mangrove estuaries in the Solomon Islands. Mar. Biol. 1990, 105, 259–267. [Google Scholar] [CrossRef]
- Frusher, S.D.; Giddins, R.L.; Smith, T.J. Distribution and abundance of grapsid crabs (Grapsidae) in a mangrove estuary: Effects of sediment characteristics, salinity tolerances, and osmoregulatory ability. Estuaries 1994, 17, 647–654. [Google Scholar] [CrossRef]
- Rasmussen, A.R.; Murphy, J.C.; Ompi, M.; Gibbons, J.W.; Uetz, P. Marine reptiles. PLoS ONE 2011, 6, e27373. [Google Scholar] [CrossRef] [PubMed]
- Platt, S.G.; Thorbjarnarson, J.B. Population status and conservation of Morelet’s crocodile, Crocodylus moreletii, in northern Belize. Biol. Conserv. 2000, 96, 21–29. [Google Scholar] [CrossRef]
- Angelici, F.M.; Politano, E.; Bogudue, A.J.; Luiselli, L. Distribution and habitat of otters (Aonyx capensis and Lutre maculicollis) in souuthern Nigeria. Ital. J. Zool. 2005, 72, 223–227. [Google Scholar] [CrossRef]
- Nowak, K. Frequent water drinking by zanzibar red colobus (Procolobus kirkii) in a mangrove forest refuge. Am. J. Primatol. 2008, 70, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Dahdouh-Guebas, F.; Jayatissa, L.P.; Di Nitto, D.; Bosire, J.O.; Lo Seen, D.; Koedam, N. How effective were mangroves as a defence against the recent tsunami? Curr. Biol. 2005, 15, R443–R447. [Google Scholar] [CrossRef] [PubMed]
- Gopal, B.; Chauhan, M. Biodiversity and its conservation in the Sundarban mangrove ecosystem. Aquatic Sci. 2006, 68, 338–354. [Google Scholar] [CrossRef]
- Field, C.D. Impact of expected climate change on mangroves. Hydrobiologia 1995, 295, 75–81. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Traditional and medicinal uses of mangroves. Mangroves Salt Marshes 1998, 2, 133–148. [Google Scholar] [CrossRef]
- Nfotabong-Atheull, A.; Din, N.; Koum, L.G.E.; Satyanarayana, B.; Koedam, N.; Dahdouh-Guebas, F. Assessing forest products usage and local residents’ perception of environmental changes in peri-urban and rural mangroves of Cameroon, Central Africa. J. Ethnobiol. Ethnomed. 2011, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Ewel, K.C.; Twilley, R.R.; Eong, J.I.N.; Usda, O.N.G.; Service, F. Different kinds of mangrove forests provide different goods and services AMONG among mangrove forests. Glob. Ecol. Biogeogr. Lett. 1998, 7, 83–94. [Google Scholar] [CrossRef]
- Walton, M.; Samonte-Tan, G.; Primavera, J.; Edwards-Jones, G.; Le Vay, L. Are mangroves worth replanting? The direct economic benefits of a community-based reforestation project. Environ. Conserv. 2006, 33, 335–343. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Danielsen, F.; Sørensen, M.K.; Olwig, M.F.; Selvam, V.; Parish, F.; Burgess, N.D.; Hiraishi, T.; Karunagaran, V.M.; Rasmussen, M.S.; Hansen, L.B.; et al. The Asian tsunami: A protective role for coastal vegetation. Science 2005, 310, 643. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Barbier, E.B. Valuing the storm protection service of estuarine and coastal ecosystems. Ecosyst. Serv. 2015, 11, 32–38. [Google Scholar] [CrossRef]
- FAO. The world’s mangroves 1980–2005; Technical Report; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007. [Google Scholar]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove Forests: One of the World’s Threatened Major Tropical Environments. BioScience 2001, 51, 807–815. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2016. Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016; 200p. [Google Scholar]
- Thomas, N.; Lucas, R.; Bunting, P.; Hardy, A.; Rosenqvist, A.; Simard, M. Distribution and drivers of global mangrove forest change, 1996–2010. PLoS ONE 2017, 12, e0179302. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proc. Natl. Acad. Sci. USA 2016, 113, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Souza Filho, P.W.M.; Farias Martins, E.D.S.; da Costa, F.R. Using mangroves as a geological indicator of coastal changes in the Bragança macrotidal flat, Brazilian Amazon: A remote sensing data approach. Ocean Coast. Manag. 2006, 49, 462–475. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Lucas, R.M.; Donnelly, B.E.; Pfitzner, K.; Milne, A.K.; Finlayson, M. A new map of mangroves for Kakadu National Park, Northern Australia, based on stereo aerial photography. Aquatic Conserv. Mar. Freshw. Ecosyst. 2007, 17, 446–467. [Google Scholar] [CrossRef]
- Spalding, M.; Blasco, F.; Field, C. World Mangrove Atlas; International Society for Mangrove Ecosystems: Okinawa, Japan, 1997. [Google Scholar]
- Spalding, M. World Atlas of Mangroves; Routledge: Abingdon, Oxon, UK, 2010. [Google Scholar]
- Friess, D.A.; Webb, E.L. Variability in mangrove change estimates and implications for the assessment of ecosystem service provision. Glob. Ecol. Biogeogr. 2014, 23, 715–725. [Google Scholar] [CrossRef]
- Ruiz-Luna, A.; Acosta-Velázquez, J.; Berlanga-Robles, C.A. On the reliability of the data of the extent of mangroves: A case study in Mexico. Ocean Coast. Manag. 2008, 51, 342–351. [Google Scholar] [CrossRef]
- Mejía-Rentería, J.C.; Castellanos-Galindo, G.A.; Cantera-Kintz, J.R.; Hamilton, S.E. A comparison of Colombian Pacific mangrove extent estimations: Implications for the conservation of a unique Neotropical tidal forest. Estuar. Coast. Shelf Sci. 2018, 212, 233–240. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial Ecoregions of the World: A New Map of Life on Earth A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Bunting, P.; Rosenqvist, A.; Lucas, R.; Rebelo, L.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, M. The Global Mangrove Watch—A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018. In submission. [Google Scholar]
- Pre-Processing of Landsat and RapidEye Data. Available online: https://arcsi.remotesensing.info (accessed on 13 September 2018).
- Clewley, D.; Bunting, P.; Shepherd, J.; Gillingham, S.; Flood, N.; Dymond, J.; Lucas, R.; Armston, J.; Moghaddam, M. A python-based open source system for geographic object-based image analysis (GEOBIA) utilizing raster attribute tables. Remote Sens. 2014, 6, 6111–6135. [Google Scholar] [CrossRef]
- Bunting, P.; Gillingham, S. The KEA image file format. Comput. Geosci. 2013, 57, 54–58. [Google Scholar] [CrossRef]
- Bunting, P.; Clewley, D.; Lucas, R.M.; Gillingham, S. The Remote Sensing and GIS Software Library (RSGISLib). Comput. Geosci. 2014, 62, 216–226. [Google Scholar] [CrossRef]
- Fatoyinbo, T.E.; Simard, M. Height and biomass of mangroves in Africa from ICESat/GLAS and SRTM. Int. J. Remote Sens. 2013, 34, 668–681. [Google Scholar] [CrossRef]
- Lee, J.S. Digital image enhancement and noise filtering by use of local statistics. Pattern Anal. Mach. Intell. IEEE Trans. 1980, PAMI-2, 165–168. [Google Scholar] [CrossRef]
- Dingle Robertson, L.; King, D.J. Comparison of pixel- and object-based classification in land cover change mapping. Int. J. Remote Sens. 2011, 32, 1505–1529. [Google Scholar] [CrossRef]
- Bruzzone, L.; Prieto, D.F. Automatic analysis of the difference image for unsupervised change detection. Geosci. Remote Sens. IEEE Trans. 2000, 38, 1171–1182. [Google Scholar] [CrossRef]
- Bruzzone, L.; Prieto, D.F. An adaptive semiparametric and context-based approach to unsupervised change detection in multitemporal remote-sensing images. Image Proc. IEEE Trans. 2002, 11, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New global forest/non-forest maps from ALOS PALSAR data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Wilson, E.B. Probable Inference, the Law of Succession, and Statistical Inference. J. Am. Stat. Assoc. 1927, 22, 209–212. [Google Scholar] [CrossRef]
- Rahman, A.F.; Dragoni, D.; Didan, K.; Barreto-Munoz, A.; Hutabarat, J.A. Detecting large scale conversion of mangroves to aquaculture with change point and mixed-pixel analyses of high-fidelity MODIS data. Remote Sens. Environ. 2013, 130, 96–107. [Google Scholar] [CrossRef]
- Thomas, N.; Lucas, R.; Itoh, T.; Simard, M.; Fatoyinbo, L.; Bunting, P.; Rosenqvist, A. An approach to monitoring mangrove extents through time-series comparison of JERS-1 SAR and ALOS PALSAR data. Wetl. Ecol. Manag. 2015, 23, 3–17. [Google Scholar] [CrossRef]
- Held, A.; Ticehurst, C.; Lymburner, L.; Williams, N. High resolution mapping of tropical mangrove ecosystems using hyperspectral and radar remote sensing. Int. J. Remote Sens. 2003, 24, 2739–2759. [Google Scholar] [CrossRef]
- Ramsey, E.W., III; Nelson, G.A.; Sapkota, S.K. Classifying coastal resources by integrating optical and radar imagery and color infrared photography. Mangroves Salt Marshes 1998, 2, 109–119. [Google Scholar] [CrossRef]
- Abdel-Hamid, A.; Dubovyk, O.; Abou El-Magd, I.; Menz, G. Mapping Mangroves Extents on the Red Sea Coastline in Egypt using Polarimetric SAR and High Resolution Optical Remote Sensing Data. Sustainability 2018, 10, 646. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, T.; Liu, M.; Jia, M.; Lin, H.; Chu, L.M.; Devlin, T.A. Potential of Combining Optical and Dual Polarimetric SAR Data for Improving Mangrove Species Discrimination Using Rotation Forest. Remote Sens. 2018, 10, 467. [Google Scholar] [CrossRef]
- Forkuor, G.; Conrad, C.; Thiel, M.; Ullmann, T.; Zoungrana, E. Integration of Optical and Synthetic Aperture Radar Imagery for Improving Crop Mapping in Northwestern Benin, West Africa. Remote Sens. 2014, 6, 6472–6499. [Google Scholar] [CrossRef]
- Van Beijma, S.; Comber, A.; Lamb, A. Random forest classification of salt marsh vegetation habitats using quad-polarimetric airborne SAR, elevation and optical RS data. Remote Sens. Environ. 2014, 149, 118–129. [Google Scholar] [CrossRef]
- Fatoyinbo, T.E.; Simard, M.; Washington-Allen, R.A.; Shugart, H.H. Landscape-scale extent, height, biomass, and carbon estimation of Mozambique’s mangrove forests with Landsat ETM+ and Shuttle Radar Topography Mission elevation data. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Hashim, M.; Hazli, W.; Kadir, W.; Yong, L.K. Global Rain Forest Mapping Activities in Malaysia: Radar Remote Sensing for Forest Survey and Biomass Indicators; Final Report JERS-1 Science Program; JAXA: Tokyo, Japan, 1999; Volume 6. [Google Scholar]
- Cornforth, W.A.; Fatoyinbo, T.E.; Freemantle, T.P.; Pettorelli, N. Advanced land observing satellite phased array type L-Band SAR (ALOS PALSAR) to inform the conservation of mangroves: Sundarbans as a case study. Remote Sens. 2013, 5, 224–237. [Google Scholar] [CrossRef]
- Kumar, T.; Patnaik, C. Discrimination of mangrove forests and characterization of adjoining land cover classes using temporal C-band Synthetic Aperture Radar data: A case study of Sundarbans. Int. J. Appl. Earth Obs. Geoinform. 2013, 23, 119–131. [Google Scholar] [CrossRef]
- Rao, B.R.M.; Dwivedi, R.S.; Kushwaha, S.P.S.; Bhattacharya, S.N.; Anand, J.B.; Dasgupta, S. Monitoring the spatial extent of coastal wetlands using ERS-1 SAR data. Int. J. Remote Sens. 1999, 20, 2509–2517. [Google Scholar] [CrossRef]
- Lucas, R.M.; Mitchell, A.L.; Rosenqvist, A.; Proisy, C.; Melius, A.; Ticehurst, C. The potential of L-band SAR for quantifying mangrove characteristics and change: case studies from the tropics. Aquatic Conserv. Mar. Freshw. Ecosyst. 2007, 17, 245–264. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Simard, M.; Grandi, G.D.; Saatchi, S.; Mayaux, P. Mapping tropical coastal vegetation using JERS-1 and ERS-1 radar data with a decision tree classifier. Int. J. Remote Sens. 2002, 23, 1461–1474. [Google Scholar] [CrossRef]
- Liu, K.; Li, X.; Shi, X.; Wang, S. Monitoring mangrove forest changes using remote sensing and GIS data with decision-tree learning. Wetlands 2008, 28, 336–346. [Google Scholar] [CrossRef]
- Clewley, D.; Whitcomb, J.; Moghaddam, M.; McDonald, K.; Chapman, B.; Bunting, P. Evaluation of ALOS PALSAR data for high-resolution mapping of vegetated wetlands in Alaska. Remote Sens. 2015, 7, 7272–7297. [Google Scholar] [CrossRef]
- Conchedda, G.; Durieux, L.; Mayaux, P. An object-based method for mapping and change analysis in mangrove ecosystems. ISPRS J. Photogramm. Remote Sens. 2008, 63, 578–589. [Google Scholar] [CrossRef]
- Nascimento, W.R.; Souza-Filho, P.W.M.; Proisy, C.; Lucas, R.M.; Rosenqvist, A. Mapping changes in the largest continuous Amazonian mangrove belt using object-based classification of multisensor satellite imagery. Estuar. Coast. Shelf Sci. 2013, 117, 83–93. [Google Scholar] [CrossRef]
- Kamal, M.; Phinn, S. Hyperspectral data for mangrove species mapping: A comparison of pixel-based and object-based approach. Remote Sens. 2011, 3, 2222–2242. [Google Scholar] [CrossRef]
- Myint, S.W.; Giri, C.P.; Wang, L.; Zhu, Z.; Gillette, S.C. Identifying mangrove species and their surrounding land use and land cover classes using an object-oriented approach with a lacunarity spatial measure. GISci. Remote Sens. 2008, 45, 188–208. [Google Scholar] [CrossRef]
- Alesheikh, A.A.; Ghorbanali, A.; Nouri, N. Coastline change detection using remote sensing. Int. J. Environ. Sci. Technol. 2007, 4, 61–66. [Google Scholar] [CrossRef]
- Walter, V. Object-based classification of remote sensing data for change detection. ISPRS J. Photogramm. Remote Sens. 2004, 58, 225–238. [Google Scholar] [CrossRef]
- Duke, N.C.; Meynecke, J.O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; et al. A World Without Mangroves? Science 2007, 317, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.A. The regularized iteratively reweighted MAD method for change detection in multi-and hyperspectral data. IEEE Trans. Image Process. 2007, 16, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, J.; Manica, A.; Swetnam, R.; Balmford, A.; Spalding, M. Predicting global patterns in mangrove forest biomass. Conserv. Lett. 2014, 7, 233–240. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Surrounding Environment | Dominant Species | Forest Extent | Forest Setting | Fragmentation | Condition | |
|---|---|---|---|---|---|---|---|
| Gulf of Fonseca | Open mudflats | Rhizophora mangle, Avicennia germinans | coastal fringes | Estuarine | Continuous stands | Anthropogenic | |
| Honduras | and agri/aquaculture | Laguncularia racemosa | fragmented in places | disturbance | |||
| Bragança (Pará), | Tropical Savannah | Rhizophora mangle, Avicennia germinans | Limited on peninsulas within | Coastal | Continuous stands | Small natural | |
| Brazil | close proximity to the coast | change | |||||
| Sáo Luis, (Maranhäo) Brazil | Arid/Tropical Savannah | Rhizophora mangle, Avicennia germinans | Large and small fringes | Riverine/ | Continuous stands | Large natural | |
| and wetlands | along a river estuary | estuarine | change | ||||
| and islands | |||||||
| Amapá, | Savannah and riparian | Rhizophora mangle, Rhizophora harisonni | large stands and | Coastal | Continuous stands and fringes | Pristine | |
| Brazil | tropical forest | Avicennia germinans, Laguncularia racemosa | small fringes | ||||
| Baia de Todos os Santos | Tropical vegetation on | Laguncularia racemosa | Fine fringes that | Estuarine | Isolated fragmented | Small natural | |
| (Bahia) Brazil | elevated slopes | line river banks | extents | change | |||
| Guayaquil, | Arid Savannah and | Avicennia germinans, Rhizophora mangle | Large stands with | Estuarine | Fragmented by | Anthropogenic | |
| Ecuador | agriculture | Limited fringes | aquaculture | disturbance | |||
| French Guiana | Tropical rainforest | Laguncularia racemosa, Avicennia germinans, | coastal fringes | Coastal | Continuous stands | Large natural | |
| Rhizophora mangle, Rhizophora racemosa | change | ||||||
| Gulf of Paria, Venezuela | Tropical | Not available | Coastal fringes | Coastal | Continuous stands | Large natural | |
| and Trinidad and Tobago | rainforest | change | |||||
| Guinea Bissau | Arid Savannah | Rhizophora mangle, Laguncalaria racemosa, | Large and small | Coastal/ | Continuous and | Large natural | |
| Avicennia germinans | riverside fringes | riverine | fragmented stands | change | |||
| and islands | |||||||
| Zambezia, Mozambique | Tropical savannah | Avicennia racemosa, Rhizophora mangle | Riverine fringes | Riverine | Naturally | Pristine | |
| vegetation | Bruguiera gymnorrhiza, Heritiera littoralis | fragmented | |||||
| Niger delta, | Tropical savannah | Avicennia africana, Rhizophora racemosa | Large forest stand | Deltaic/ | Continuous | Pristine | |
| Nigeria | vegetation | Rhizophora mangle, Rhizophora harrisonii | coastal | forest | |||
| Riau, Indonesia | Plantation and | Not Available | Coastal island and | Coastal/ | Continuous | Large natural | |
| peatland | riverine fringes | riverine | stands | change | |||
| Mahakam delta and Balikpapan | Agriculture and | Avicennia sonneratia, Rhizophora Bruguiera, | Large forest stands | Deltaic | Heavily | Anthropogenic | |
| East Kalimantan | tropical savannah | Xylocarpus sp., Nypa sp. | fragmented | disturbance | |||
| Indonesia | |||||||
| Perak, Malaysia | Urban and | Rhizophora apiculata, R. apiculata Blume., | Large forest stands | Coastal | Fragmented by | Anthropogenic | |
| agriculture | Bruguiera gymnorrhiza, B. parviflora | logging | disturbance | ||||
| Kakadu National Park | Arid Savannah/ | Sonneratia alba, Rhizophra stylosa, | Coastal and | Coastal/ | Naturally | Pristine | |
| (NT) Australia | saltpan | Avicennia Marina | riverine fringes | Riverine | fragmented | ||
| and islands |
| Number | Study Site | Dist to Water (m) | Max Elevation (m) |
|---|---|---|---|
| 1. | Gulf of Fonesca, Honduras | 10,468 | 50 |
| 2. | Bragança (Pará), Brazil | 4928 | 29 |
| 3. | Sáo Luis (marahháo), Brazil | 6252 | 35 |
| 4. | Amapá State, Brazil | 42,388 | 30 |
| 5. | Todos os Santos, Brazil | 5100 | 25 |
| 6. | Guayaquil, Ecuador | 4996 | 50 |
| 7. | French Guiana | 16,160 | 39 |
| 8. | Gulf of Paria Venezuela/Trinidad and Tobago | 8918 | 50 |
| 9. | Guinea Bissau | 9670 | 19 |
| 10. | Zambezia, Mozambique | 31,260 | 18 |
| 11. | Niger delta, Nigeria | 9736 | 23 |
| 12. | Riau, Indonesia | 4702 | 19 |
| 13. | East Kalimantan, Indonesia | 3752 | 17 |
| 14. | Balikpapan, Indonesia | 3752 | 17 |
| 15. | Perak, Malaysia | 4334 | 23 |
| 16. | Kakadu NP (NT), Australia | 13,874 | 15 |
| Study Site | Area (ha) | Giri Area (ha) | Giri Difference (%) | Stratified Random Accuracy (%) | Border Accuracy (%) | Overall Accuracy (%) | Kappa | Wilson Range (%) | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Amapá state | 130,870 | 92,900 | 29.0 | 94.3 | 97.4 | 95.5 | 0.90 | 93.9–97.0 | 0.019 |
| Bragança | 251,420 | 234,730 | 6.6 | 92.1 | 95.4 | 93.2 | 0.86 | 91.3–95.2 | 0.048 |
| Sáo Luis | 172,820 | 169,670 | 1.8 | 93.5 | 90.5 | 90.1 | 0.79 | 87.5–92.6 | 0.099 |
| Biai de Todos os Santos | 55,270 | 35,920 | 35.0 | 89.8 | 71.8 | 86.5 | 0.71 | 83.6–89.5 | 0.147 |
| Guayaquil | 134,130 | 152,100 | 11.3 | 98.0 | 99.6 | 98.5 | 0.97 | 97.4–99.6 | <0.001 |
| French Guiana | 142,100 | 150,000 | 5.3 | 90.7 | 89.9 | 90.4 | 0.79 | 88.0–92.7 | 0.100 |
| Guinea Bissau | 592,990 | 740,430 | 19.9 | 93.9 | 93.3 | 93.7 | 0.86 | 92.1–95.2 | 0.046 |
| Honduras | 104,030 | 89,360 | 14.1 | 94.2 | 90.0 | 92.7 | 0.84 | 90.5–94.9 | 0.062 |
| Mozambique | 24,230 | 41,710 | 41.7 | 89.7 | 76.0 | 85.4 | 0.68 | 82.3–88.5 | 0.169 |
| Riau | 150,570 | 111,590 | 25.9 | 91.8 | 87.7 | 90.5 | 0.79 | 87.9–93.0 | 0.095 |
| Mahakam Delta | 69,980 | 76,980 | 9.1 | 93.7 | 98.1 | 94.6 | 0.88 | 92.5–96.7 | 0.032 |
| Balikpapan | 58,770 | 51,250 | 12.8 | 91.3 | 89.3 | 90.6 | 0.79 | 88.2–93.1 | 0.013 |
| Perak | 44,050 | 43,930 | 0.3 | 95.5 | 98.0 | 96.0 | 0.92 | 94.2–97.9 | 0.012 |
| Niger delta | 372,440 | 317,670 | 14.7 | 95.7 | 96.2 | 95.8 | 0.92 | 94.1–97.6 | 0.093 |
| Kakadu | 9800 | 21,350 | 54.1 | 97.3 | 95.8 | 97.0 | 0.93 | 95.4–98.6 | 0.006 |
| Gulf of Paria | 216,290 | 288,260 | 25.0 | 94.0 | 91.5 | 93.4 | 0.85 | 91.6–95.3 | 0.055 |
| Combined | 2,529,760 | 2,517,850 | 0.5 | 93.5 | 91.2 | 92.8 | 0.84 | 92.2–93.3 | 0.059 |
| Study Site | 1996–2010 | 1996–2010 | 2007–2010 | 2007–2010 | 2007–2008 | 2007–2008 | 2008–2009 | 2008–2009 | 2009–2010 | 2009–2010 |
|---|---|---|---|---|---|---|---|---|---|---|
| Gain (ha) | Loss (ha) | Gain (ha) | Loss (ha) | Gain (ha) | Loss (ha) | Gain (ha) | Loss (ha) | Gain (ha) | Loss (ha) | |
| Amapá State | 4360 | 7560 | 180 | 1300 | 10 | 0 | 30 | 1 | 1 | 0 |
| Bragança | 5370 | 3920 | 330 | 5 | 1 | 0 | 5 | 0 | 5 | 0 |
| Sáo Luis | 3260 | 5770 | 160 | 1280 | 3 | 4 | 2 | 0 | 8 | 2 |
| Baia de Todos os Santos | 4580 | 3340 | 3 | 2 | 2 | 0 | 1 | 0 | 4 | 0 |
| Guayaquil | 10,170 | 5280 | 600 | 870 | 1 | 0 | 4 | 0 | 7 | 0 |
| French Guiana | 15,570 | 8570 | 3250 | 3120 | 8 | 0 | 70 | 0 | 60 | 20 |
| Guinea Bissau | 47,930 | 28,420 | 2090 | 1510 | 9 | 20 | 50 | 2 | 120 | 0 |
| Honduras | 8560 | 2620 | 540 | 490 | 10 | 20 | 10 | 4 | 20 | 0 |
| Mozambique | 740 | 820 | 20 | 240 | 0 | 0 | 1 | 0 | 7 | 0 |
| Riau | 5060 | 950 | 390 | 240 | 0 | 0 | 30 | 0 | 20 | 1 |
| Mahakam delta | 1490 | 23,003 | 990 | 920 | 2 | 0 | 70 | 0 | 4 | 0 |
| Balikpapan | 2820 | 3950 | 5 | 320 | 3 | 0 | 6 | 0 | 7 | 0 |
| Perak | 1050 | 240 | 50 | 70 | 0 | 0 | 3 | 0 | 2 | 0 |
| Niger delta | 9660 | 1170 | 980 | 70 | 3 | 0 | 2 | 0 | 10 | 0 |
| Kakadu | 2210 | 570 | 30 | 40 | 1 | 0 | 10 | 0 | 7 | 0 |
| Gulf of Paria | 5160 | 3360 | 580 | 620 | 0 | 0 | 7 | 0 | 7 | 0 |
| Total | 127,990 | 76,860 | 10,198 | 11,097 | 53 | 44 | 301 | 7 | 289 | 41 |
| Study Site | 1996–2010 | 1996–2010 | 2007–2010 | 2007–2010 | 2007–2008 | 2007–2008 | 2008–2009 | 2008–2009 | 2009–2010 | 2009–2010 |
|---|---|---|---|---|---|---|---|---|---|---|
| Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | |
| Amapá State | 78.0 | 71.8 | 88.5 | 73.8 | 100 | 96.8 | 98.9 | 96.8 | 100 | 96.8 |
| Bragança | 66.4 | 70.2 | 79.6 | 74.3 | 100 | 98.0 | 100 | 98.0 | 99.6 | 98.0 |
| Sáo Luis | 61.0 | 69.6 | 96.8 | 68.2 | 100 | 96.0 | 98.8 | 96.8 | 99.2 | 96.5 |
| Baia de Todos os Santos | 64.0 | 47.4 | 93.8 | 91.8 | 100 | 92.4 | 99.6 | 92.4 | 99.6 | 92.4 |
| Guayaquil | 61.9 | 84.0 | 69.1 | 76.4 | 99.7 | 96.8 | 98.1 | 96.8 | 99.0 | 96.8 |
| French Guiana | 69.6 | 79.2 | 85.4 | 96.2 | 100 | 100 | 99.3 | 100 | 98.5 | 98.8 |
| Guinea Bissau | 67.8 | 50.0 | 84.5 | 74.5 | 99.3 | 91.8 | 98.3 | 93.6 | 95.8 | 93.6 |
| Honduras | 66.5 | 93.2 | 70.5 | 88.0 | 100 | 96.1 | 100 | 96.8 | 97.8 | 96.8 |
| Mozambique | 58.9 | 58.3 | 95.6 | 89.7 | 100 | 94.4 | 100 | 94.4 | 98.4 | 94.4 |
| Riau | 57.6 | 52.0 | 84.5 | 69.6 | 100 | 95.2 | 98.5 | 95.2 | 99.6 | 94.8 |
| Mahakam delta | 68.0 | 83.2 | 94.2 | 88.2 | 100 | 98.4 | 94.4 | 98.4 | 99.7 | 98.4 |
| Balikpapan | 57.4 | 85.8 | 80.1 | 93.9 | 100 | 98.4 | 99.6 | 98.4 | 100 | 98.4 |
| Perak | 52.8 | 79.1 | 97.7 | 88.2 | 100 | 94.0 | 100 | 94.0 | 100 | 94.0 |
| Niger delta | 50.4 | 54.3 | 55.8 | 52.9 | 99.2 | 99.7 | 100 | 99.7 | 100 | 99.7 |
| Kakadu | 59.6 | 66.9 | 87.5 | 94.7 | 99.6 | 96.0 | 100 | 96.0 | 99.2 | 96.0 |
| Gulf of Paria | 62.0 | 53.0 | 67.4 | 67.2 | 100 | 98.8 | 100 | 98.8 | 100 | 98.8 |
| Total | 62.7 | 68.3 | 81.5 | 78.3 | 99.9 | 96.5 | 99 | 96.7 | 99.1 | 96.5 |
| Kappa | 0.24 | 0.35 | 0.53 | 0.45 | 0.89 | NA | 0.86 | NA | 0.84 | NA |
| Wilson Range (%) | 61.3–64.0 | 66.9–69.7 | 80.2–82.7 | 77.0–79.7 | 99.7–100 | 96.7–97.2 | 98.6–99.4 | 95.9–97.4 | 98.8–99.5 | 95.8–97.3 |
| Study Site | 1996–2010 | 1996–2010 | 2007–2010 | 2007–2010 | 2007–2008 | 2007–2008 | 2008–2009 | 2008–2009 | 2009–2010 | 2009–2010 |
|---|---|---|---|---|---|---|---|---|---|---|
| Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | Gain (%) | Loss (%) | |
| Amapá State | 94.6 | 93.0 | 93.9 | 96.3 | 100 | 96.8 | 97.4 | 96.8 | 100 | 96.8 |
| Bragança | 95.4 | 72.8 | 91.0 | 96.6 | 100 | 98.0 | 100 | 98.0 | 100 | 98.0 |
| Sáo Luis | 95.0 | 96.0 | 99.6 | 96.3 | 100 | 96.8 | 98.1 | 96.8 | 99.2 | 96.5 |
| Baia de Todos os Santos | 90.0 | 94.4 | 96.7 | 92.6 | 100 | 92.4 | 100 | 92.4 | 99.6 | 92.4 |
| Guayaquil | 95.0 | 85.2 | 95.5 | 96.6 | 100 | 96.8 | 99.7 | 96.8 | 99.3 | 96.8 |
| French Guiana | 88.8 | 96.6 | 97.9 | 99.5 | 100 | 100 | 99.3 | 100 | 98.2 | 98.2 |
| Guinea Bissau | 97.5 | 95.4 | 91.5 | 93.4 | 99.3 | 92.2 | 97.7 | 93.6 | 95.2 | 93.6 |
| Honduras | 95.5 | 94.4 | 92.9 | 97.6 | 100 | 96.1 | 100 | 96.8 | 97.8 | 96.8 |
| Mozambique | 93.9 | 95.3 | 98.6 | 93.8 | 100 | 94.4 | 100 | 94.4 | 98.8 | 94.4 |
| Riau | 91.4 | 95.6 | 92.0 | 95.1 | 100 | 95.2 | 98.5 | 95.2 | 99.6 | 95.2 |
| Mahakam delta | 98.3 | 97.2 | 95.9 | 97.8 | 100 | 98.4 | 94.5 | 98.4 | 99.7 | 98.4 |
| Balikpapan | 95.4 | 95.7 | 94.4 | 98.2 | 100 | 98.4 | 99.2 | 98.4 | 100 | 98.4 |
| Perak | 81.6 | 94.7 | 100 | 92.9 | 100 | 94.0 | 99.6 | 94.0 | 100 | 94.0 |
| Niger delta | 99.8 | 99.8 | 90.0 | 98.8 | 100 | 99.7 | 100 | 99.7 | 98.1 | 99.7 |
| Kakadu | 97.6 | 96.0 | 97.4 | 96.2 | 99.6 | 96.0 | 100 | 96.0 | 100 | 96.0 |
| Gulf of Paria | 93.8 | 97.0 | 96.5 | 98.3 | 100 | 98.8 | 99.6 | 98.8 | 99.6 | 98.8 |
| Total | 94.0 | 93.7 | 94.9 | 96.5 | 99.9 | 96.5 | 98.9 | 96.7 | 99 | 96.6 |
| Kappa | 0.88 | 0.87 | 0.88 | 0.92 | 0.95 | NA | 0.84 | NA | 0.81 | NA |
| Wilson Range (%) | 93.3–94.7 | 92.9–94.4 | 94.2–95.6 | 95.9–97.1 | 99.8–100 | 95.8–97.3 | 98.5–99.3 | 95.9–97.4 | 98.6–99.4 | 95.8–97.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, N.; Bunting, P.; Lucas, R.; Hardy, A.; Rosenqvist, A.; Fatoyinbo, T. Mapping Mangrove Extent and Change: A Globally Applicable Approach. Remote Sens. 2018, 10, 1466. https://doi.org/10.3390/rs10091466
Thomas N, Bunting P, Lucas R, Hardy A, Rosenqvist A, Fatoyinbo T. Mapping Mangrove Extent and Change: A Globally Applicable Approach. Remote Sensing. 2018; 10(9):1466. https://doi.org/10.3390/rs10091466
Chicago/Turabian StyleThomas, Nathan, Peter Bunting, Richard Lucas, Andy Hardy, Ake Rosenqvist, and Temilola Fatoyinbo. 2018. "Mapping Mangrove Extent and Change: A Globally Applicable Approach" Remote Sensing 10, no. 9: 1466. https://doi.org/10.3390/rs10091466
APA StyleThomas, N., Bunting, P., Lucas, R., Hardy, A., Rosenqvist, A., & Fatoyinbo, T. (2018). Mapping Mangrove Extent and Change: A Globally Applicable Approach. Remote Sensing, 10(9), 1466. https://doi.org/10.3390/rs10091466