Exploring the Inclusion of Small Regenerating Trees to Improve Above-Ground Forest Biomass Estimation Using Geospatial Data


Abstract
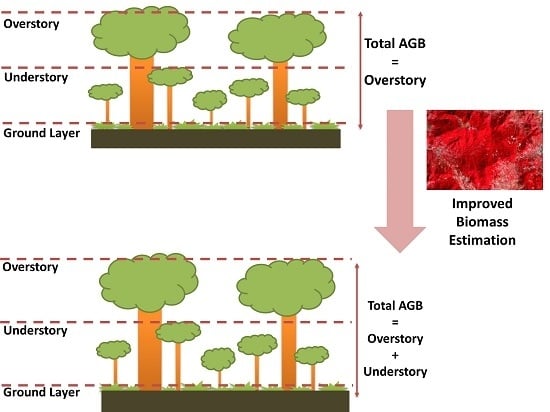
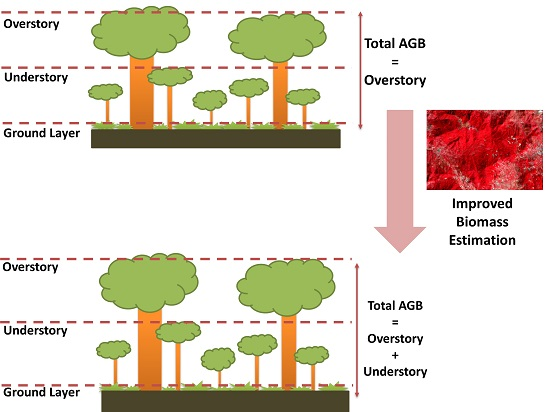
1. Introduction
2. Study Area and Data Used
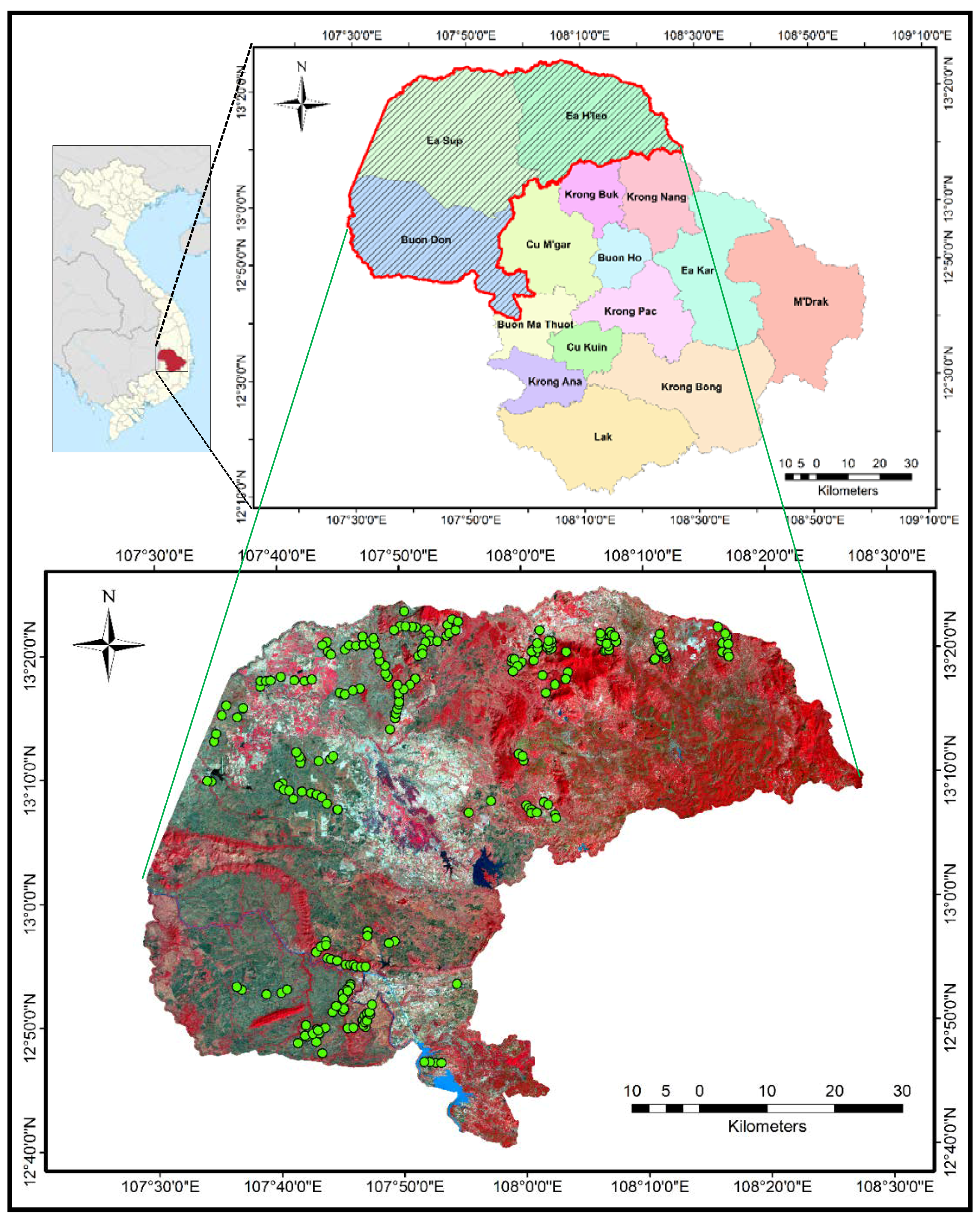
2.1. Site Description
2.2. Field Data Collection
2.3. Remote Sensing and Ancillary Data
3. Methodology
3.1. In Situ AGB Estimation
3.2. Remotely Sensed Data Pre-Processing and Creation of Derived Variables
3.3. Regression Algorithms
3.4. Modeling Approaches
- Test 1: Test 11 (Raw spectral bands) versus Test 12 (Raw spectral bands + topographic variables)
- Test 2: Test 21 (Vegetation Indices) versus Test 22 (Vegetation Indices + topographic variables)
- Test 3: Test 31 (Raw spectral band ratios) versus Test 32 (Raw spectral band ratios + topographic variables)
- Test 4: Test 41 (PC bands) versus Test 42 (PC bands + topographic variables)
- Test 5: Test 51 (Texture) versus Test 52 (Texture + topographic variables)
- Test 6: Test 61 (All Landsat-derived variables combined) versus Test 62 (All Landsat-derived variables combined + topographic variables)
4. Results
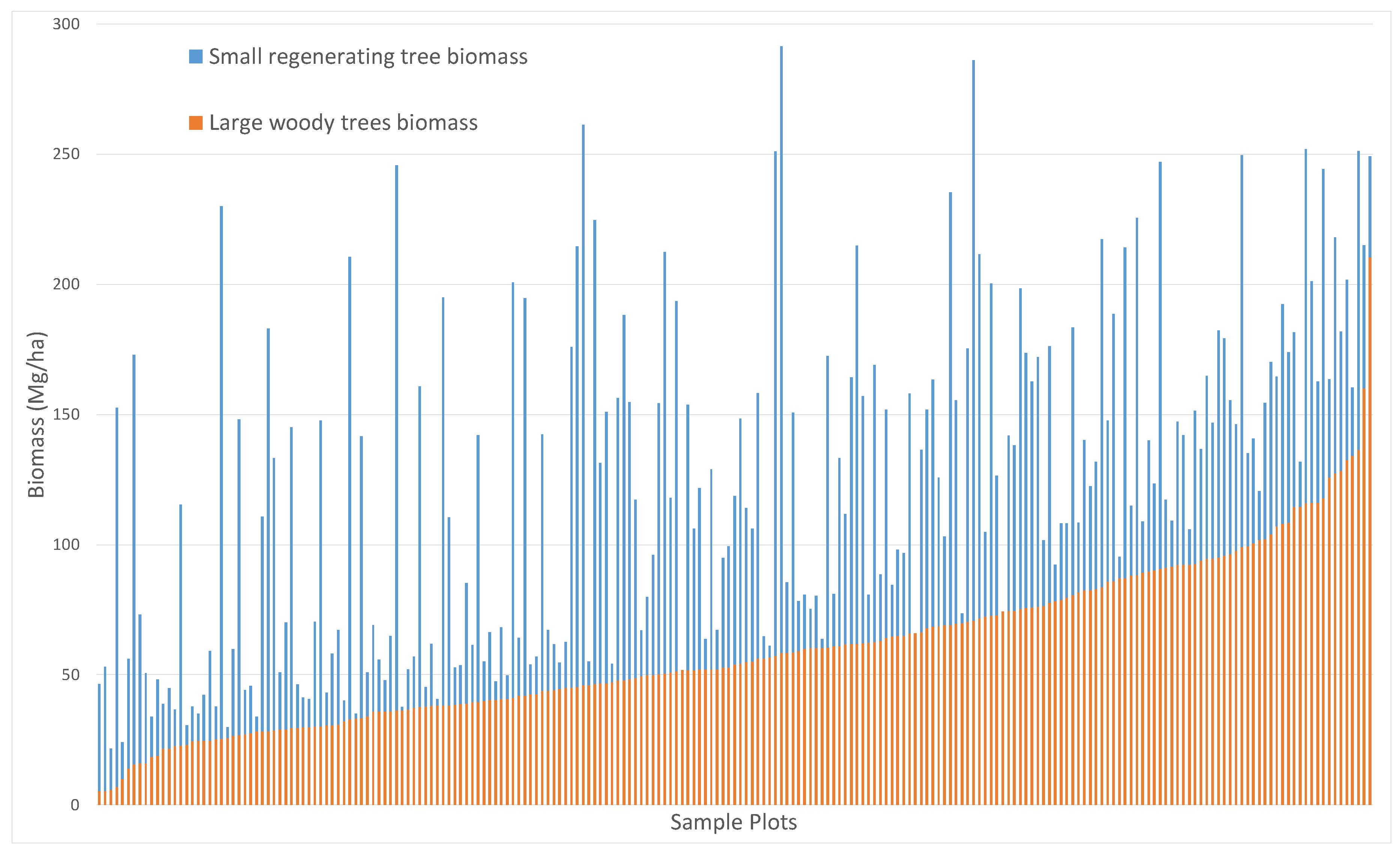
4.1. In Situ AGB Estimation from the Forest Inventory
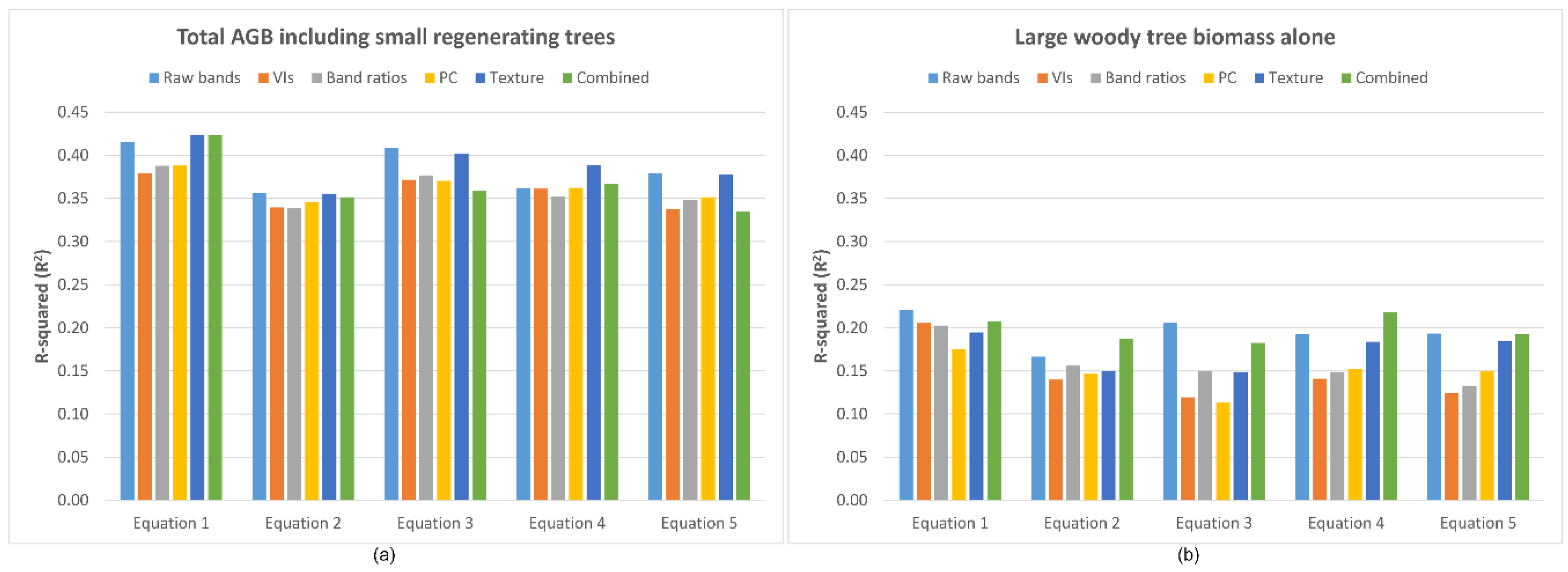
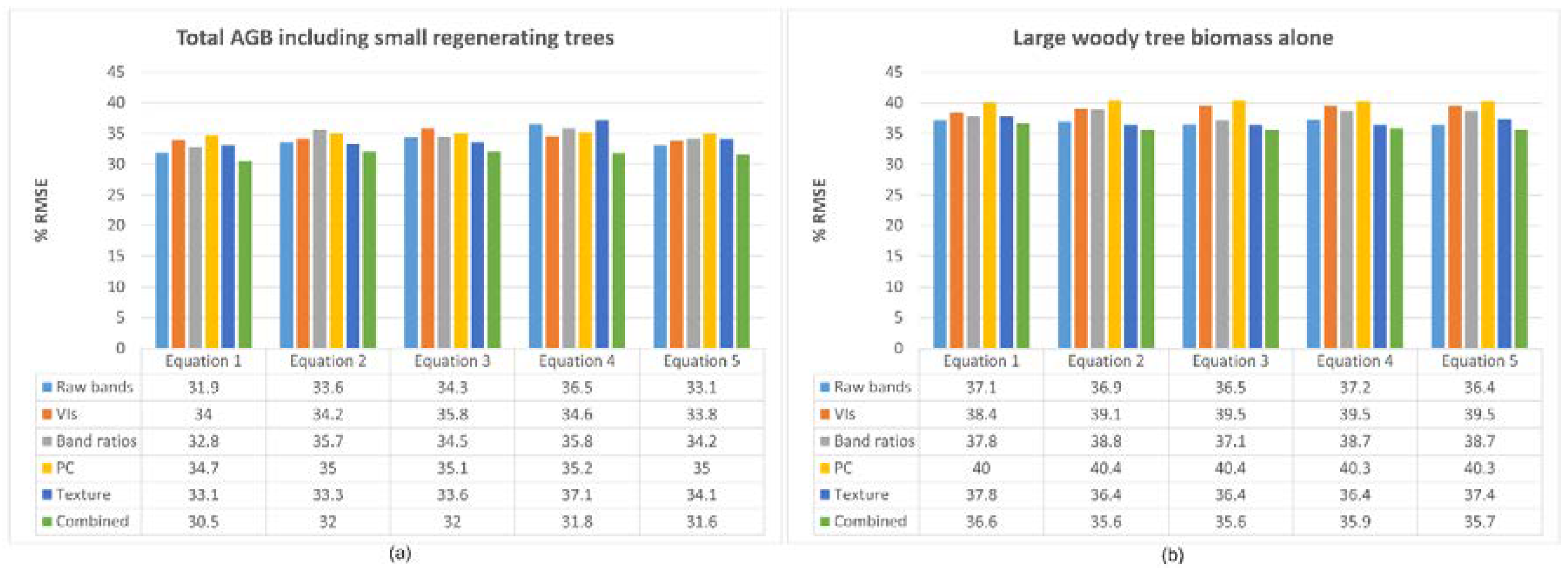
4.2. AGB Estimation from Satellite Data
4.3. Assessment of Optical Imagery-Derived Variables
4.4. Performance of Topographic Variables
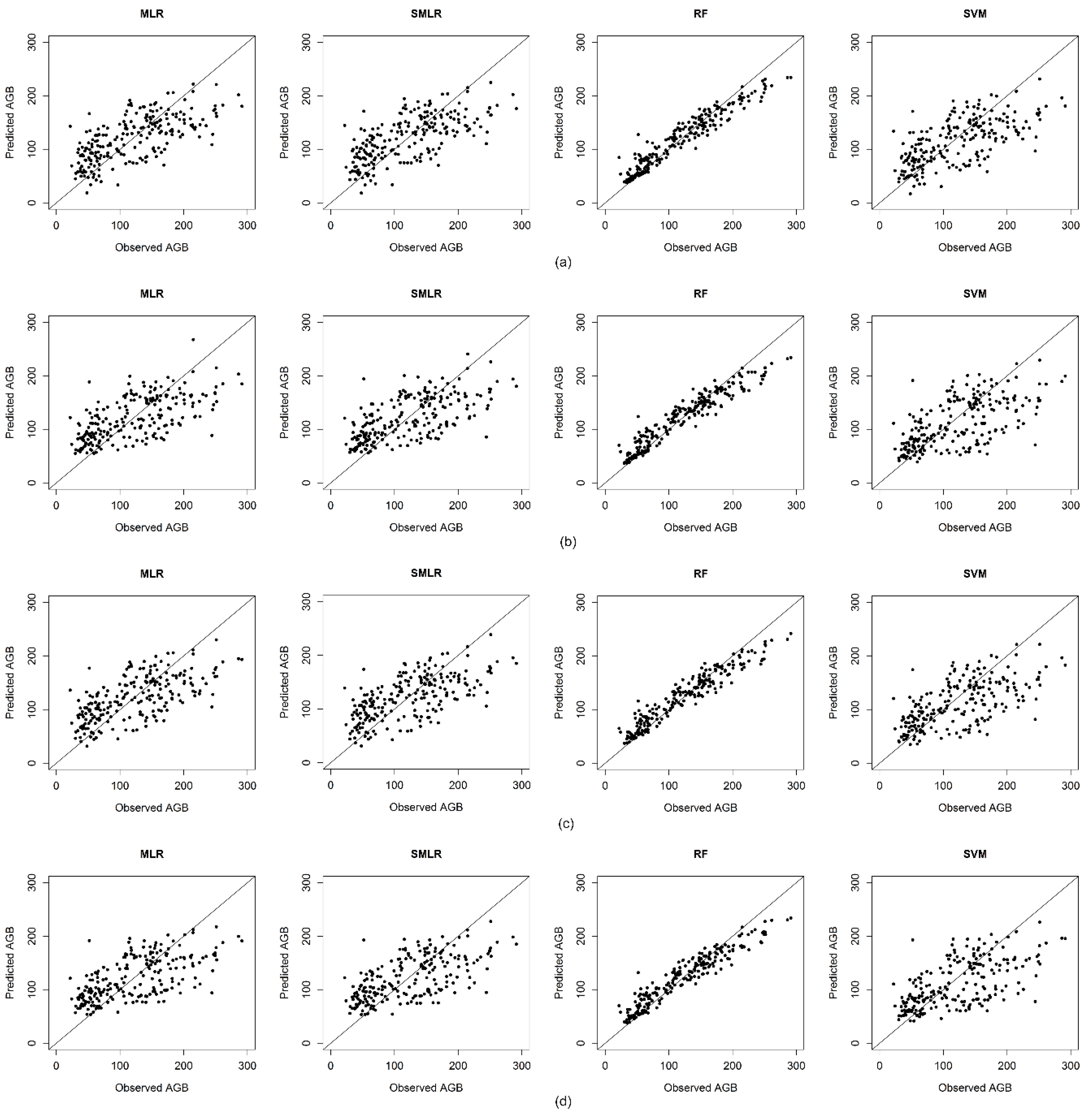
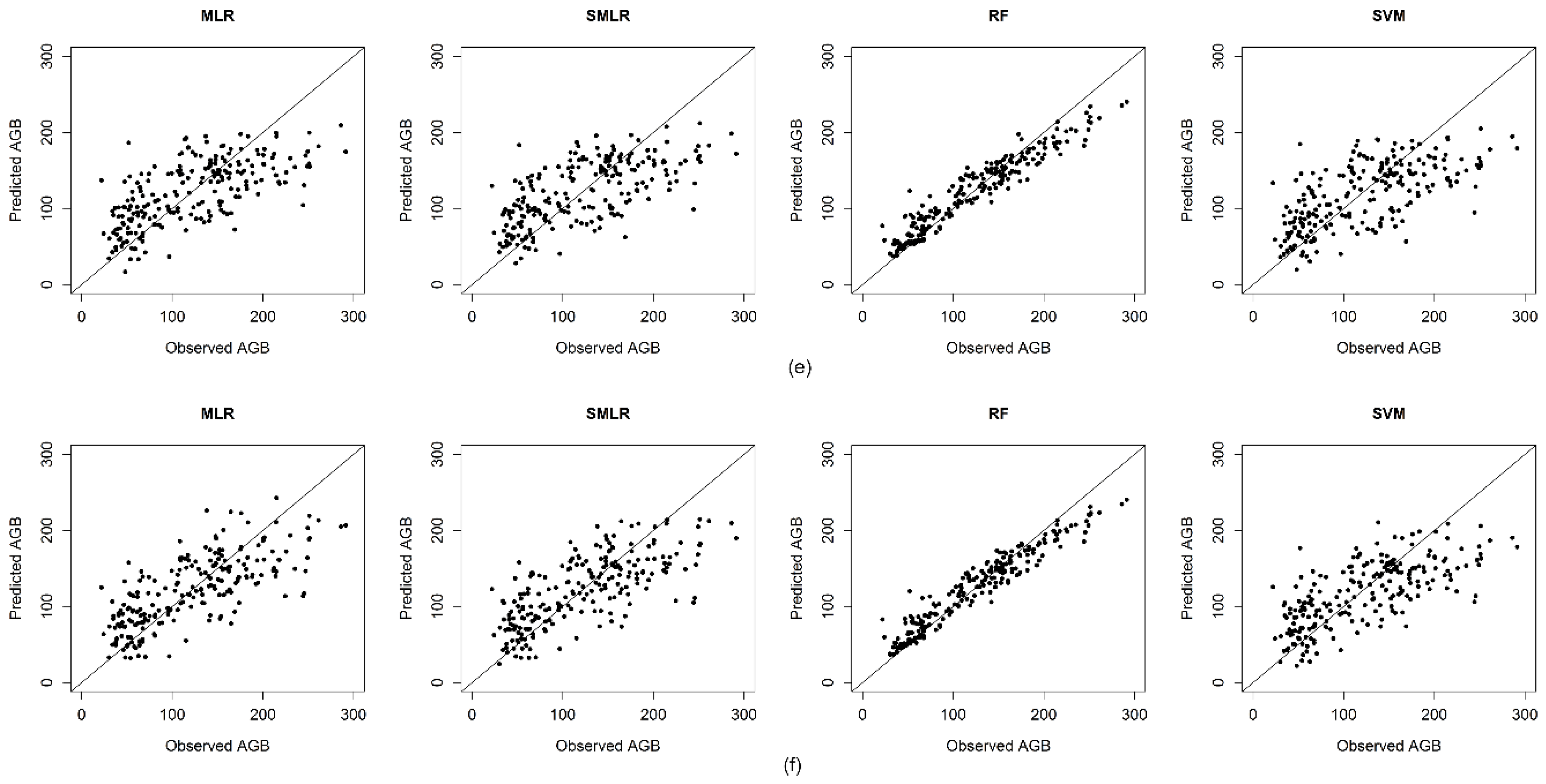
4.5. Performance of Regression Methods
5. Discussion
5.1. In Situ AGB Estimation from the Forest Inventory
5.2. AGB Estimation from the Satellite Data
5.3. Assessment of Optical Imagery-Derived Variables
5.4. Performance of Topographic Variables
5.5. Performance of Regression Methods
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gibbs, H.; Brown, S.; Niles, J.; Foley, J. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 045023. [Google Scholar] [CrossRef]
- Brown, S.; Gillespie, A.J.; Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. For. Sci. 1989, 35, 881–902. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Foody, G.M.; Cutler, M.E.; McMorrow, J.; Pelz, D.; Tangki, H.; Boyd, D.S.; Douglas, I. Mapping the biomass of Bornean tropical rain forest from remotely sensed data. Glob. Ecol. Biogeogr. 2001, 10, 379–387. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Christensen, N.L.; Bourgeauchavez, L.L. Correlating radar backscatter with components of biomass in loblolly-pine forests. IEEE Trans. Geosci. Remote Sens. 1995, 33, 643–659. [Google Scholar] [CrossRef]
- Brown, S.; Gaston, G. Use of forest inventories and geographic information systems to estimate biomass density of tropical forests: Application to tropical Africa. Environ. Monit. Assess. 1995, 38, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Iverson, L.R.; Lugo, A.E. Land-use and biomass changes of forests in Peninsular Malaysia from 1972 to 1982: A GIS approach. In Effects of Land-Use Change on Atmospheric CO2 Concentrations: South and Southeast Asia as A Case Study; Dale, V.H., Ed.; Springer: New York, NY, USA, 1994; pp. 117–143. [Google Scholar]
- Wijaya, A.; Kusnadi, S.; Gloaguen, R.; Heilmeier, H. Improved strategy for estimating stem volume and forest biomass using moderate resolution remote sensing data and GIS. J. For. Res. 2010, 21, 1–12. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed]
- Houghton, R.A. How well do we know the flux of CO2 from land-use change? Tellus B Chem. Phys. Meteorol. 2010, 62, 337–351. [Google Scholar] [CrossRef]
- Lu, D. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar] [CrossRef]
- MacLean, D.A.; Wein, R.W. Changes in understory vegetation with increasing stand age in New Brunswick forests: Species composition, cover, biomass, and nutrients. Can. J. Bot. 1977, 55, 2818–2831. [Google Scholar] [CrossRef]
- Ferraz, A.; Gonçalves, G.; Soares, P.; Tomé, M.; Mallet, C.; Jacquemoud, S.; Bretar, F.; Pereira, L. Comparing small-footprint lidar and forest inventory data for single strata biomass estimation—a case study over a multi-layered mediterranean forest. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium, Munich, Germany, 22–27 July 2012; pp. 6384–6387. [Google Scholar]
- Gonzalez, M.; Augusto, L.; Gallet-Budynek, A.; Xue, J.; Yauschew-Raguenes, N.; Guyon, D.; Trichet, P.; Delerue, F.; Niollet, S.; Andreasson, F.; et al. Contribution of understory species to total ecosystem aboveground and belowground biomass in temperate Pinus pinaster Ait. Forests. For. Ecol. Manag. 2013, 289, 38–47. [Google Scholar] [CrossRef]
- Meyfroidt, P.; Lambin, E.F. The causes of the reforestation in Vietnam. Land Use Policy 2008, 25, 182–197. [Google Scholar] [CrossRef]
- Khuc, Q.V.; Tran, B.Q.; Meyfroidt, P.; Paschke, M.W. Drivers of deforestation and forest degradation in Vietnam: An exploratory analysis at the national level. For. Policy Econ. 2018, 90, 128–141. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Baker, P.J. Structure and composition of deciduous dipterocarp forest in central Vietnam: Patterns of species dominance and regeneration failure. Plant Ecol. Divers. 2016, 9, 589–601. [Google Scholar] [CrossRef]
- Russell, M.B.; D’Amato, A.W.; Schulz, B.K.; Woodall, C.W.; Domke, G.M.; Bradford, J.B. Quantifying understorey vegetation in the US Lake States: A proposed framework to inform regional forest carbon stocks. Forestry 2014, 87, 629–638. [Google Scholar] [CrossRef]
- Helms, J.A. The Dictionary of Forestry. 1998. Available online: http://www.citeulike.org/group/7954/article/4089380 (accessed on 21 June 2018).
- Archer, J.K. Understory Vegetation and Soil Response to Silvicultural Activity in A Southeastern Mixed Pine Forest: A Chronosequence Study. Master’s Thesis, University of Florida, Gainesville, FL, USA, May 2003. [Google Scholar]
- Svoboda, M.; Matějka, K.; Kopáček, J. Biomass and element pools of understory vegetation in the catchments of čertovo lake and plešné lake in the bohemian forest. Biologia 2006, 61, S509–S521. [Google Scholar] [CrossRef]
- Tremblay, N.O.; Larocque, G.R. Seasonal dynamics of understory vegetation in four eastern Canadian forest types. Int. J. Plant Sci. 2001, 162, 271–286. [Google Scholar] [CrossRef]
- Estornell, J.; Ruiz, L.A.; Velázquez-Martí, B.; Fernández-Sarría, A. Estimation of shrub biomass by airborne lidar data in small forest stands. For. Ecol. Manag. 2011, 262, 1697–1703. [Google Scholar] [CrossRef]
- Halpern, C.B.; Spies, T.A. Plant species diversity in natural and managed forests of the pacific northwest. Ecol. Appl. 1995, 5, 913–934. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A.R. Aboveground biomass of understorey vegetation has a negligible or negative association with overstorey tree species diversity in natural forests. Glob. Ecol. Biogeogr. 2016, 25, 141–150. [Google Scholar] [CrossRef]
- Zavitkovski, J. Ground vegetation biomass, production, and efficiency of energy utilization in some northern Wisconsin forest ecosystems. Ecology 1976, 57, 694–706. [Google Scholar] [CrossRef]
- Zeng, H.Q.; Liu, Q.J.; Feng, Z.W.; Ma, Z.Q. Biomass equations for four shrub species in subtropical China. J. For. Res. 2010, 15, 83–90. [Google Scholar] [CrossRef]
- Joyce, L.A.; Mitchell, J.E. Understory cover/biomass relationships in Alabama forest types. Agrofor. Syst. 1989, 9, 205–210. [Google Scholar] [CrossRef]
- Zheng, D.; Rademacher, J.; Chen, J.; Crow, T.; Bresee, M.; Le Moine, J.; Ryu, S.R. Estimating aboveground biomass using Landsat 7 ETM+ data across a managed landscape in northern Wisconsin, USA. Remote Sens. Environ. 2004, 93, 402–411. [Google Scholar] [CrossRef]
- Porté, A.; Samalens, J.C.; Dulhoste, R.; Teissier Du Cros, R.; Bosc, A.; Meredieu, C. Using cover measurements to estimate aboveground understorey biomass in Maritime pine stands. Ann. For. Sci. 2009, 66, 307p1–307p11. [Google Scholar] [CrossRef]
- Ducey, M.J.; Astrup, R. Rapid, nondestructive estimation of forest understory biomass using a handheld laser rangefinder. Can. J. For.t Res. 2018, 48, 803–808. [Google Scholar] [CrossRef]
- Muukkonen, P.; Mäkipää, R.; Laiho, R.; Minkkinen, K.; Vasander, H.; Finér, L. Relationship between biomass and percentage cover in understorey vegetation of boreal coniferous forests. Silva Fenn. 2006, 40, 231–245. [Google Scholar] [CrossRef]
- Hermy, M. Accuracy of visual cover assessments in predieting standing crop and environmental correlation in deciduous forests. Vegetatio 1988, 75, 57–64. [Google Scholar] [CrossRef]
- Chiarucci, A.; Wilson, J.B.; Anderson, B.J.; Dominicis, V. Cover versus biomass as an estimate of species abundance: Does it make a difference to the conclusions? J. Veg. Sci. 1999, 10, 35–42. [Google Scholar] [CrossRef]
- Chave, J.; RiÉRa, B.; Dubois, M.A. Estimation of biomass in a neotropical forest of French Guiana: Spatial and temporal variability. J. Trop. Ecol. 2001, 17, 79–96. [Google Scholar] [CrossRef]
- Avtar, R.; Suzuki, R.; Sawada, H. Natural forest biomass estimation based on plantation information using PALSAR data. PLoS ONE 2014, 9, e86121. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Lu, D. Aboveground biomass estimation using Landsat TM data in the Brazilian amazon. Int. J. Remote Sens. 2005, 26, 2509–2525. [Google Scholar] [CrossRef]
- Main-Knorn, M.; Cohen, W.B.; Kennedy, R.E.; Grodzki, W.; Pflugmacher, D.; Griffiths, P.; Hostert, P. Monitoring coniferous forest biomass change using a Landsat trajectory-based approach. Remote Sens. Environ. 2013, 139, 277–290. [Google Scholar] [CrossRef]
- Powell, S.L.; Cohen, W.B.; Healey, S.P.; Kennedy, R.E.; Moisen, G.G.; Pierce, K.B.; Ohmann, J.L. Quantification of live aboveground forest biomass dynamics with Landsat time-series and field inventory data: A comparison of empirical modeling approaches. Remote Sens. Environ. 2010, 114, 1053–1068. [Google Scholar] [CrossRef]
- Vicharnakorn, P.; Shrestha, R.; Nagai, M.; Salam, A.; Kiratiprayoon, S. Carbon stock assessment using remote sensing and forest inventory data in Savannakhet, Lao PDR. Remote Sens. 2014, 6, 5452. [Google Scholar] [CrossRef]
- Wohlfart, C.; Wegmann, M.; Leimgruber, P. Mapping threatened dry deciduous dipterocarp forest in south-east Asia for conservation management. Trop. Conserv. Sci. 2014, 7, 597–613. [Google Scholar] [CrossRef]
- Bunyavejchewin, S. Canopy structure of the dry dipterocarp forest of Thailand. Thai For. Bull. 1983, 14, 1–93. [Google Scholar]
- Sodhi, N.S.; Brook, B.W. Fragile southeast Asian biotas. Biol. Conserv. 2008, 141, 883–884. [Google Scholar] [CrossRef]
- Bunyavejchewin, S.; Baker, P.; Davies, S.J. Seasonally dry tropical forest in Continental Southeast Asia: Structure, composition, and dynamics. In The Ecology and Conservation of Seasonally Dry Forests in Asia; Mc-Shea, W.J., Davis, S.J., Bhumpakphan, N., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011; pp. 9–35. [Google Scholar]
- Huy, B.; Poudel, K.; Kralicek, K.; Hung, N.; Khoa, P.; Phương, V.; Temesgen, H. Allometric equations for estimating tree aboveground biomass in tropical dipterocarp forests of Vietnam. Forests 2016, 7, 180. [Google Scholar] [CrossRef]
- Huy, B.; Tri, P.C.; Triet, T. Assessment of enrichment planting of teak (Tectona grandis) in degraded dry deciduous dipterocarp forest in the Central Highlands, Vietnam. South. For. J. For. Sci. 2018, 80, 75–84. [Google Scholar] [CrossRef]
- Tổng Cục Lâm Nghiệp; BỘ NÔNG NGHIỆP VÀ PHÁT TRIỂN NÔNG THÔN. Tài liệu tập huấn hướng dẫn kỹ thuật điều tra rừng. 2013. Available online: http://hocday.com/ti-liu-tp-hun-hng-dn-k-thut-iu-tra-rng.html (accessed on 21 June 2018).
- Castro, K.L.; Sanchez-Azofeifa, G.A.; Rivard, B. Monitoring secondary tropical forests using space-borne data: Implications for Central America. Int. J. Remote Sens. 2003, 24, 1853–1894. [Google Scholar] [CrossRef]
- Lu, D.; Chen, Q.; Wang, G.; Liu, L.; Li, G.; Moran, E. A survey of remote sensing-based aboveground biomass estimation methods in forest ecosystems. Int. J. Digit. Earth 2016, 9, 63–105. [Google Scholar] [CrossRef]
- Morel, A.C.; Saatchi, S.S.; Malhi, Y.; Berry, N.J.; Banin, L.; Burslem, D.; Nilus, R.; Ong, R.C. Estimating aboveground biomass in forest and oil palm plantation in Sabah, Malaysian Borneo using Alos Palsar data. For. Ecol. Manag. 2011, 262, 1786–1798. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Aguilar, S.; Hernandez, A.; Lao, S.; Perez, R. Error propagation and scaling for tropical forest biomass estimates. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Tanase, M.A.; Panciera, R.; Lowell, K.; Tian, S.; Hacker, J.M.; Walker, J.P. Airborne multi-temporal l-band polarimetric SAR data for biomass estimation in semi-arid forests. Remote Sens. Environ. 2014, 145, 93–104. [Google Scholar] [CrossRef]
- Avitabile, V.; Baccini, A.; Friedl, M.A.; Schmullius, C. Capabilities and limitations of Landsat and land cover data for aboveground woody biomass estimation of Uganda. Remote Sens. Environ. 2012, 117, 366–380. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Houghton, R.A.; Dos Santos Alvalá, R.C.; Soares, J.V.; Yu, Y. Distribution of aboveground live biomass in the Amazon basin. Glob. Chang. Biol. 2007, 13, 816–837. [Google Scholar] [CrossRef]
- Shen, W.; Li, M.; Huang, C.; Wei, A. Quantifying live aboveground biomass and forest disturbance of mountainous natural and plantation forests in Northern Guangdong, China, based on multi-temporal Landsat, PALSAR and field plot data. Remote Sens. 2016, 8, 595. [Google Scholar] [CrossRef]
- Steininger, M.K. Satellite estimation of tropical secondary forest above-ground biomass: Data from Brazil and Bolivia. Int. J. Remote Sens. 2000, 21, 1139–1157. [Google Scholar] [CrossRef]
- Dolph, K.L. Height-Diameter Equations for Young-Growth Red Fir in California and Southern Oregon; Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1989. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef]
- Anh, L.V.; Paull, D.J.; Griffin, A.L. Investigating the capabilities of new microwave alos-2/palsar-2 data for biomass estimation. In Proceedings of the SPIE Remote Sensing, Edinburgh, UK, 18 October 2016. [Google Scholar] [CrossRef]
- Dube, T.; Mutanga, O. Investigating the robustness of the new Landsat-8 operational land imager derived texture metrics in estimating plantation forest aboveground biomass in resource constrained areas. ISPRS J. Photogramm. Remote Sens. 2015, 108, 12–32. [Google Scholar] [CrossRef]
- Inoguchi, A.; Henry, M.; Birigazzi, L.; Sola, G. Tree Allometric Equation Development for Estimation of Forest Above-Ground Biomass in Vietnam; UN-REDD Programm: Hanoi, Vietnam, 2012. [Google Scholar]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a worldwide wood economics spectrum. Dryad Data Repos. 2009. [Google Scholar] [CrossRef]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer. 1997. Available online: http://www.fao.org/docrep/w4095e/w4095e00.htm (accessed on 21 June 2018).
- Zhang, X. On the estimation of biomass of submerged vegetation using Landsat thematic mapper (TM) imagery: A case study of the Honghu Lake, PR China. Int. J. Remote Sens. 1998, 19, 11–20. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. ISPRS J. Photogramm. Remote Sens. 2015, 102, 222–231. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the NASA. Goddard Space Flight Center 3d ERTS-1 Symposium, Washington, DC, USA; 1973; pp. 309–317. [Google Scholar]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Tucker, C.J. A spectral method for determining the percentage of green herbage material in clipped samples. Remote Sens. Environ. 1980, 9, 175–181. [Google Scholar] [CrossRef]
- Pearson, R.L.; Miller, L.D.; Program, U.S.I.B. Remote mapping of standing crop biomass for estimation of the productivity of the shortgrass prairie, Pawnee National Grasslands, Colorado. In Proceedings of the Eighth International Symposium on Remote Sensing of Environment, Ann Arbor, MI, USA, 2–6 October 1972. [Google Scholar]
- Kaufman, Y.J.; Tanré, D. Strategy for direct and indirect methods for correcting the aerosol effect on remote sensing: From AVHRR to EOS-MODIS. Remote Sens. Environ. 1996, 55, 65–79. [Google Scholar] [CrossRef]
- Pinty, B.; Verstraete, M.M. Gemi: A non-linear index to monitor global vegetation from satellites. Vegetatio 1992, 101, 15–20. [Google Scholar] [CrossRef]
- Ji, L.; Wylie, B.K.; Nossov, D.R.; Peterson, B.; Waldrop, M.P.; McFarland, J.W.; Rover, J.; Hollingsworth, T.N. Estimating aboveground biomass in interior Alaska with Landsat data and field measurements. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 451–461. [Google Scholar] [CrossRef]
- Labrecque, S.; Fournier, R.A.; Luther, J.E.; Piercey, D. A comparison of four methods to map biomass from Landsat-TM and inventory data in western newfoundland. For. Ecol. Manag. 2006, 226, 129–144. [Google Scholar] [CrossRef]
- Dube, T.; Mutanga, O. Evaluating the utility of the medium-spatial resolution Landsat 8 multispectral sensor in quantifying aboveground biomass in Umgeni catchment, south Africa. ISPRS J. Photogramm. Remote Sens. 2015, 101, 36–46. [Google Scholar] [CrossRef]
- Kelsey, K.; Neff, J. Estimates of aboveground biomass from texture analysis of Landsat imagery. Remote Sens. 2014, 6, 6407. [Google Scholar] [CrossRef]
- Sarker, L.R.; Nichol, J.E. Improved forest biomass estimates using ALOS AVNIR-2 texture indices. Remote Sens. Environ. 2011, 115, 968–977. [Google Scholar] [CrossRef]
- Neumann, M.; Saatchi, S.S.; Ulander, L.M.H.; Fransson, J.E.S. Assessing performance of L- and P-band polarimetric interferometric SAR data in estimating boreal forest above-ground biomass. IEEE Trans. Geosci. Remote Sens. 2012, 50, 714–726. [Google Scholar] [CrossRef]
- Nichol, J.E.; Sarker, M.L.R. Improved biomass estimation using the texture parameters of two high-resolution optical sensors. IEEE Trans. Geosci. Remote Sens. 2011, 49, 930–948. [Google Scholar] [CrossRef]
- Akinwande, O.; Dikko, H.G.; Gulumbe, U. Identifying the limitation of stepwise selection for variable selection in regression analysis. Am. J. Theor. Appl. Stat. 2015, 4, 414–419. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Axelsson, C.; Skidmore, A.K.; Schlerf, M.; Fauzi, A.; Verhoef, W. Hyperspectral analysis of mangrove foliar chemistry using PLSR and support vector regression. Int. J. Remote Sens. 2013, 34, 1724–1743. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by random forest. R-News 2002, 2/3, 18–22. [Google Scholar]
- Dimitriadou, E.; Hornik, K.; Leisch, F.; Meyer, D.; Weingessel, A. E1071: Misc Functions of the Department of Statistics (e1071), TU Wien. R Package Version 1.5-25. 2011. Available online: http://www.citeulike.org/user/nrv2/article/12239162 (accessed on 21 June 2018).
- Delucia, E.; Sipe, T.; Herrick, J.; Maherali, H. Sapling biomass allocation and growth in the understory of a deciduous hardwood forest. Am. J. Bot. 1998, 85, 955. [Google Scholar] [CrossRef] [PubMed]
- Trædal, L.T.; Vedeld, P.O.; Pétursson, J.G. Analyzing the transformations of forest PES in Vietnam: Implications for REDD+. For. Policy Econ. 2016, 62, 109–117. [Google Scholar] [CrossRef]
- Kajisa, T.; Murakami, T.; Mizoue, N.; Top, N.; Yoshida, S. Object-based forest biomass estimation using Landsat ETM+ in Kampong Thom Province, Cambodia. J. For. Res. 2009, 14, 203–211. [Google Scholar] [CrossRef]
- Lu, D.; Batistella, M. Exploring TM image texture and its relationships with biomass estimation in Rondônia, Brazilian Amazon. Acta Amazon. 2005, 35, 249–257. [Google Scholar] [CrossRef]
- Chirici, G.; Barbati, A.; Corona, P.; Marchetti, M.; Travaglini, D.; Maselli, F.; Bertini, R. Non-parametric and parametric methods using satellite images for estimating growing stock volume in alpine and Mediterranean forest ecosystems. Remote Sens. Environ. 2008, 112, 2686–2700. [Google Scholar] [CrossRef]
- Luther, J.E.; Fournier, R.A.; Piercey, D.E.; Guindon, L.; Hall, R.J. Biomass mapping using forest type and structure derived from Landsat TM imagery. Int. J. Appl. Earth Obs. Geoinf. 2006, 8, 173–187. [Google Scholar] [CrossRef]
- Woodcock, C.E.; Allen, R.; Anderson, M.; Belward, A.; Bindschadler, R.; Cohen, W.; Gao, F.; Goward, S.N.; Helder, D.; Helmer, E.; et al. Free access to Landsat imagery. Science 2008, 320, 1011. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, D.; Cohen, W.B.; Kennedy, R.E.; Yang, Z. Using Landsat-derived disturbance and recovery history and lidar to map forest biomass dynamics. Remote Sens. Environ. 2014, 151, 124–137. [Google Scholar] [CrossRef]
- Pham, L.T.H.; Brabyn, L. Monitoring mangrove biomass change in vietnam using SPOT images and an object-based approach combined with machine learning algorithms. ISPRS J. Photogramm. Remote Sens. 2017, 128, 86–97. [Google Scholar] [CrossRef]
- Luong, N.V.; Tateishi, R.; Hoan, N.T.; To, T.T.; Son, L.M. An analysis of forest biomass changes using geospatial tools and ground survey data: A case study in Yok Don national park, Central Highlands of Vietnam. Vietnam J. Earth Sci. 2014, 36, 439–450. [Google Scholar] [CrossRef]
- Foody, G.M.; Boyd, D.S.; Cutler, M.E.J. Predictive relations of tropical forest biomass from Landsat TM data and their transferability between regions. Remote Sens. Environ. 2003, 85, 463–474. [Google Scholar] [CrossRef]
- Tian, X.; Li, Z.; Su, Z.; Chen, E.; van der Tol, C.; Li, X.; Guo, Y.; Li, L.; Ling, F. Estimating montane forest above-ground biomass in the upper reaches of the Heihe River Basin using Landsat-TM data. Int. J. Remote Sens. 2014, 35, 7339–7362. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Gale, J. Availability of carbon dioxide for photosynthesis at high altitudes: Theoretical considerations. Ecology 1972, 53, 494–497. [Google Scholar] [CrossRef]
- Mohd Zaki, N.A.; Abd Latif, Z. Carbon sinks and tropical forest biomass estimation: A review on role of remote sensing in aboveground-biomass modelling. Geocarto Int. 2017, 32, 701–716. [Google Scholar] [CrossRef]
- Van der Laan, C.; Verweij, P.A.; Quiñones, M.J.; Faaij, A.P. Analysis of biophysical and anthropogenic variables and their relation to the regional spatial variation of aboveground biomass illustrated for north and east Kalimantan, Borneo. Carbon Balance Manag. 2014, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Lefsky, M.A.; Turner, D.P.; Guzy, M.; Cohen, W.B. Combining lidar estimates of aboveground biomass and Landsat estimates of stand age for spatially extensive validation of modeled forest productivity. Remote Sens. Environ. 2005, 95, 549–558. [Google Scholar] [CrossRef]
- Propastin, P. Modifying geographically weighted regression for estimating aboveground biomass in tropical rainforests by multispectral remote sensing data. Int. J. Appl. Earth Obs. Geoinfor. 2012, 18, 82–90. [Google Scholar] [CrossRef]
- Wang, X.Y.; Guo, Y.G.; He, J. Estimation of forest biomass by integrating ALOS PALSAR and HJ1B data. In Proceedings of the SPIE 9260, Land Surface Remote Sensing II, Beijing, China, 8 November 2014. [Google Scholar]
- Laurin, G.V.; Balling, J.; Corona, P.; Mattioli, W.; Papale, D.; Puletti, N.; Rizzo, M.; Truckenbrodt, J.; Urban, M. Above-ground biomass prediction by Sentinel-1 multitemporal data in central Italy with integration of ALOS2 and Sentinel-2 data, J. Appl. Remote Sens. 2018, 12, 016008. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
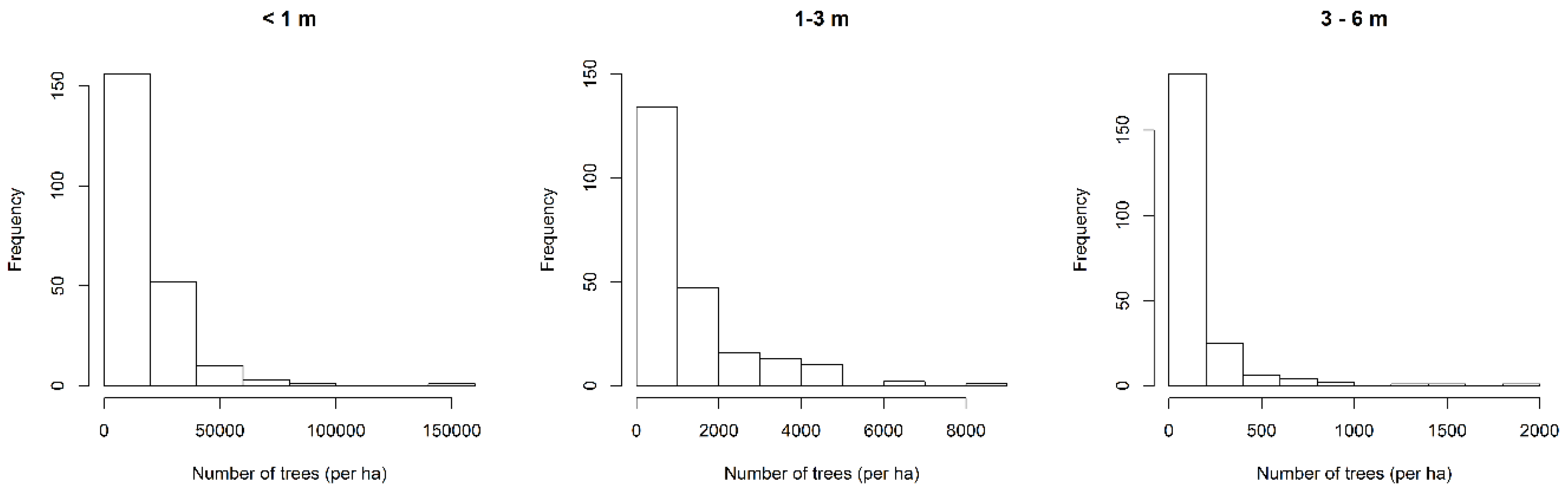
| Height | Min | Max | Mean | Standard Deviation |
|---|---|---|---|---|
| <1 m | 300 | 141,100 | 15,385 | 17,292 |
| 1–3 m | 0 | 8700 | 1219 | 1376 |
| 3–6 m | 0 | 2000 | 136 | 252 |
| No | Equations | References | R2/R2 adj | Remarks |
|---|---|---|---|---|
| Allometric equations used for calculating large woody tree biomass | ||||
| 1 | Equation (1) | [62] | 0.93 | UN-REDD—developed specifically for deciduous forests in the Central Highlands in Vietnam |
| 2 | Equation (2) | [64] | 0.97 | For tropical and humid regions |
| 3 | Equation (3) | [2] | 0.97 | For humid regions |
| 4 | Equation (4) | [3] | N/A | |
| 5 | Equation (5) | [2] | 0.99 | |
| Allometric equations used for calculating small regenerating tree biomass | ||||
| 6 | Equation (6) | [23] | ||
| Variables | Formula | References |
|---|---|---|
| Six raw spectral band reflectances | Band 2, Blue; Band 3, Green; Band 4, Red; Band 5, Near Infrared (NIR); Band 6, Shortwave Infrared (SWIR) 1; Band 7, SWIR 2 | |
| Seven Vegetation Indices | ||
| Normalized Difference Vegetation Index (NDVI) | [67] | |
| Soil Adjusted Vegetation Index (SAVI) | where L = 0.5 | [68] |
| Enhanced Vegetation Index (EVI) | [69] | |
| Difference Vegetation Index (DVI) | NIR − RED | [70] |
| Ratio Vegetation Index (RVI) | NIR/RED | [71] |
| Atmospherically Resistant Vegetation Index (ARVI) | (NIR − RB)/(NIR + RB) where RB = RED − 1 × (BLUE − RED) | [72] |
| Global Environment Monitoring Index (GEMI) | N × (1 − 0.25 × n) − (RED − 0.125)/(1 − RED) where n = [2(NIR2 − RED2) + 1.5NIR + 0.5RED]/(NIR + RED + 0.5) | [73] |
| Six Simple Band Ratios | BLUE/GREEN BLUE/RED BLUE/NIR GREEN/RED GREEN/NIR RED/NIR | |
| Three Principal Component Bands (PC1–PC3) | ||
| Eight Grey Level Co-occurrence Matrix (GLCM) texture parameters (window sizes from 3 × 3 to 11 × 11) from six raw spectral bands | ||
| Mean (ME) | ||
| Variance (VA) | ||
| Homogeneity (HO) | ||
| Contrast (CO) | ||
| Dissimilarity (DI) | ||
| Entropy (EN) | ||
| Second Moment (SE) | ||
| Correlation (CC) |
| No | Equations | References | Min | Max | Mean | Std Dev. |
|---|---|---|---|---|---|---|
| Large woody tree biomass | ||||||
| 1 | Equation (1) | [62] | 5.37 | 210.31 | 59.70 | 31.06 |
| 2 | Equation (2) | [64] | 7.37 | 424.56 | 102.26 | 59.18 |
| 3 | Equation (3) | [2] | 4.34 | 264.41 | 62.57 | 38.36 |
| 4 | Equation (4) | [3] | 4.58 | 323.48 | 74.78 | 45.63 |
| 5 | Equation (5) | [2] | 5.27 | 335.40 | 81.31 | 48.20 |
| Small regenerating tree biomass | ||||||
| 6 | Equation (6) | [23] | 1.08 | 233.01 | 62.76 | 51.94 |
| Biomass Range (Mg/ha) | Percentage of Plots (%) Equation (1) | Percentage of Plots (%) Equation (2) | Percentage of Plots (%) Equation (3) | Percentage of Plots (%) Equation (5) | Percentage of Plots (%) Equation (4) |
|---|---|---|---|---|---|
| Large woody tree biomass | |||||
| 0–50 | 42.04 | 16.81 | 41.59 | 34.51 | 29.65 |
| 50–100 | 45.58 | 37.17 | 40.27 | 39.82 | 39.82 |
| 100–150 | 8.41 | 24.78 | 12.39 | 16.37 | 18.14 |
| 150–200 | 0.44 | 12.39 | 1.77 | 4.42 | 7.08 |
| >200 | 0.44 | 5.75 | 0.88 | 1.77 | 2.21 |
| Total AGB (including biomass of both large woody trees and small regenerating trees) | |||||
| 0–50 | 12.83 | 3.54 | 14.60 | 11.50 | 9.73 |
| 50–100 | 26.99 | 25.22 | 23.89 | 24.34 | 23.89 |
| 100–150 | 23.45 | 14.16 | 23.45 | 18.58 | 18.58 |
| 150–200 | 21.24 | 23.89 | 22.57 | 23.01 | 23.01 |
| >200 | 12.39 | 30.09 | 12.39 | 19.47 | 21.68 |
| Test Variables | MLR | SMLR | RF | SVM | |||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | ||
| 1 | Raw spectral bands | 0.40 | 48.82 | 0.41 | 48.72 | 0.39 | 49.87 | 0.40 | 49.76 |
| 2 | VIs | 0.36 | 50.81 | 0.36 | 50.52 | 0.30 | 54.14 | 0.36 | 51.78 |
| 3 | Band ratios | 0.37 | 50.17 | 0.37 | 50.18 | 0.39 | 49.89 | 0.37 | 51.43 |
| 4 | PC | 0.37 | 49.89 | 0.37 | 49.89 | 0.27 | 55.01 | 0.37 | 51.01 |
| 5 | Texture | 0.40 | 49.01 | 0.41 | 48.74 | 0.33 | 52.23 | 0.40 | 49.28 |
| 6 | Combined | 0.40 | 51.23 | 0.40 | 51.28 | 0.38 | 49.61 | 0.41 | 49.52 |
| Test Variables | MLR | SMLR | RF | SVM | |||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | R2 | RMSE (Mg/ha) | ||
| 1 | Raw spectral bands | 0.42 | 48.06 | 0.44 | 47.42 | 0.48 | 46.22 | 0.42 | 49.21 |
| 2 | VIs | 0.37 | 50.22 | 0.36 | 50.54 | 0.41 | 49.12 | 0.38 | 51.28 |
| 3 | Band ratios | 0.40 | 49.09 | 0.39 | 49.28 | 0.47 | 46.42 | 0.39 | 50.90 |
| 4 | PC | 0.39 | 49.26 | 0.38 | 49.58 | 0.40 | 48.89 | 0.39 | 50.84 |
| 5 | Texture | 0.42 | 48.14 | 0.40 | 48.74 | 0.42 | 48.18 | 0.42 | 48.66 |
| 6 | Combined | 0.40 | 51.53 | 0.43 | 49.67 | 0.46 | 46.49 | 0.42 | 48.79 |
| Topographic Variables | Equation (1) | Equation (2) | Equation (3) | Equation (4) | Equation (5) |
|---|---|---|---|---|---|
| Elevation | 0.49 | 0.45 | 0.46 | 0.45 | 0.45 |
| Slope | 0.47 | 0.40 | 0.43 | 0.40 | 0.39 |
| Aspect | 0.03 | 0.01 | 0.00 | −0.03 | −0.03 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, A.V.; Paull, D.J.; Griffin, A.L. Exploring the Inclusion of Small Regenerating Trees to Improve Above-Ground Forest Biomass Estimation Using Geospatial Data. Remote Sens. 2018, 10, 1446. https://doi.org/10.3390/rs10091446
Le AV, Paull DJ, Griffin AL. Exploring the Inclusion of Small Regenerating Trees to Improve Above-Ground Forest Biomass Estimation Using Geospatial Data. Remote Sensing. 2018; 10(9):1446. https://doi.org/10.3390/rs10091446
Chicago/Turabian StyleLe, Anh V., David J. Paull, and Amy L. Griffin. 2018. "Exploring the Inclusion of Small Regenerating Trees to Improve Above-Ground Forest Biomass Estimation Using Geospatial Data" Remote Sensing 10, no. 9: 1446. https://doi.org/10.3390/rs10091446
APA StyleLe, A. V., Paull, D. J., & Griffin, A. L. (2018). Exploring the Inclusion of Small Regenerating Trees to Improve Above-Ground Forest Biomass Estimation Using Geospatial Data. Remote Sensing, 10(9), 1446. https://doi.org/10.3390/rs10091446