1. Introduction
Despite its tropical latitude, the island of Hawaii has relatively low plant diversity due to its age and isolation. A large share of the species in the Hawaiian Islands have evolved in the region and are therefore unique to the archipelago. One of these endemic species,
Metrosideros polymorpha (‘ōhi‘a), is the most prevalent tree on Hawaii Island. Likely due to its high degree of evolved polymorphism [
1,
2], ‘ōhi‘a is present as a dominant or co-dominant tree in all the forested ecological zones on the island ranging from sea level to 2700 m above sea level, occupying an area larger than 250,000 ha [
3]. ‘Ōhi‘a accounts for more than 50% of the total basal area of woody plants throughout the Hawaiian archipelago [
4]. As such, this species supports a large number of associated endemic plant and animal communities found on the island [
3,
5], and it is deeply engrained into Hawaiian culture [
3]. Substantial losses of this keystone species would fundamentally and permanently change the ecological and cultural landscape of the island [
4,
6,
7].
In 2010, a large number of ‘ōhi‘a trees began exhibiting browning foliage, rapidly followed by tree mortality. In subsequent years, the incidence of ‘ōhi‘a canopy browning and mortality spread dramatically, affecting an area of more than 20,000 hectares by late 2016 [
8]. In response, a multi-agency team of scientists, forest managers, and concerned citizens assembled to determine the cause and review approaches to fight the spread of this disease, subsequently named Rapid ‘Ōhi‘a Death (ROD). Two distinct and previously unknown fungal species in the genus
Ceratocystis were eventually isolated as the pathogens causing ROD [
9]. In recent years, hundreds of confirmed cases of ROD resulting from at least one of two
Ceratocystis species have been documented in ‘ōhi‘a stands across the island. Infection by either of these two pathogens causes a similar progression of symptoms, although at a slightly different pace. The infection is first noticeable when leaves on one or more branches of the tree begin to yellow, wilt, and turn brown within days to weeks. This brown stage is quite noticeable, and it is relatively unique to ROD infection [
10] (this issue). Brown leaves then drop from the infected tree over several weeks to months until only bare branches remain.
While the cause and progression of the disease in individual ‘ōhi‘a are fairly well understood, many important questions remain with respect to how the disease spreads and how to slow down this spread. Knowledge of the full extent of ROD infection across the island, both current and past, is critical to the determination of how and where to apply limited resources to contain the disease. However, because of the span of spatial scale involved—‘ōhi‘a crowns are relatively small, and infections are scattered across an area of many thousands of hectares of remote forest—detection of ROD-infected trees across the island remains a challenge. Currently available tools fall short in accomplishing this task [
8]. In addition, the most extensive ‘ōhi‘a forests lie in very remote areas and on private property, making them difficult to reach on foot. Unmanned aerial vehicles are quick to deploy and offer very high-resolution, detailed imagery, but are only able to map small areas at a time and are not permitted over certain federal and most private land. The efficacy and coverage of helicopter surveys is limited by the ability of the human eye to spot single brown crowns at a distance, by a lack of spatial accuracy, and by the substantial cost of flight time. Conversely, data from Earth-orbiting satellites are readily available and could cover the full region, but are too spatially or spectrally coarse to pinpoint individual browning trees. These shortcomings suggest that appropriate mapping data collected from higher-altitude aircraft may be able to provide the needed coverage and resolution to identify potential ROD infections across the island.
The Carnegie Airborne Observatory (CAO) [
11] is uniquely able to fill this gap, as it is able to collect laser-guided imaging spectroscopy (LGIS) data from altitudes of 2000 or more meters above ground level. LGIS is the fusion of data from a high-fidelity visible-to-shortwave imaging spectrometer (VSWIR) to three-dimensional information provided by a light detection and ranging (LiDAR) scanner [
12,
13]. This combination of boresight-aligned instruments allows spectrometer readings from a forest canopy to be mapped to sub-meter precision. Asner et al. [
10] (in review) demonstrate that various stages of ROD infection exhibit distinct LGIS spectra at leaf to canopy scales, and they used this information to develop a spectral mapping signature for ‘ōhi‘a trees likely infected by ROD. Here, we show that those LGIS spectra can be used to produce maps of both current ROD infections (i.e., brown, desiccated ‘ōhi‘a tree crowns) and past ROD infections (i.e., fully-dead, leafless tree crowns). We apply the LGIS approach to map brown and leafless ‘ōhi‘a trees throughout Hawaii Island in 2017, and we assess its accuracy for future mapping efforts with the intent to track the spread of the disease in space and time.
2. Materials and Methods
2.1. Airborne Data Collection
We collected data with the CAO between 7 June and 5 July 2017 over the island of Hawaii with three co-aligned instruments: a visible-to-shortwave imaging spectrometer (VSWIR), a dual-channel airborne light-detection and ranging (LiDAR) scanner, and a high-resolution 3-band (red, green, and blue) 60-megapixel digital camera. For optimal flight coverage, the island was partitioned into 15 compartments of fairly uniform forest type ranging in size from 12,000 to 75,000 ha. During flight, aircraft speed was maintained within 10% of the 60 m s−1 nominal air speed and within 200 m of the nominal planned elevation of 2000 m above ground level. The LiDAR was operated at an effective pulse frequency of 200 kHz, a scan frequency of 34 Hz, 30 percent overlap, and total field-of-view of 38 degrees to match the 34-degree FOV of the VSWIR after clipping 2 degrees from the scan edges. These settings yielded a mean density of 3.3 pulses m−2 and a VSWIR resolution of 2 m. VSWIR data were collected in 427 channels between 350 and 2485 nm at 5 nm increments (full-width at half max). The high-resolution digital camera had an effective resolution of 24 cm.
2.2. Data Processing and Orthorectification
Estimated location error in LiDAR point vertical positioning from planar surface analysis was less than 10 cm (root mean squared error). LiDAR point cloud data were processed to 1-m resolution digital surface maps of ground elevation, canopy surface elevation, and canopy surface height above ground using the LAStools software suite (Rapidlasso GmbH; Gilching, Germany).
The VSWIR radiance data were first orthorectified using LiDAR surface maps, positon, and orientation data from the on-board GPS-IMU navigation system, and known boresight and position offsets between the two instruments. To improve the signal-to-noise ratio, the radiance data were reduced by averaging each pair neighboring bands. This yielded a dataset of 214 bands at increments of 10 nm. Using computed observation angles and elevation, atmospheric correction was performed using ACORN-6LX software (Imspec LLC; Palmdale, CA, USA) to transform the VSWIR radiance data into surface-level reflection spectra. For each flight line, we used an iterative procedure to find the optimal setting for the ACORN-6LX visibility parameter that is used to specify the amount of aerosols present at the time of collection. Beginning with a setting of 100 km, the visibility parameter was decreased until the resulting reflectance at 420 nm was <1%. The individual VSWIR reflectance data were mosaicked into one image for each of the 15 flight coverage compartments using the criteria of minimum phase angle (angle from sun to target to instrument) for overlap regions. These coverages were mosaicked into a full island map using the criteria of maximum normalized difference vegetation index (NDVI).
2.3. Training Data
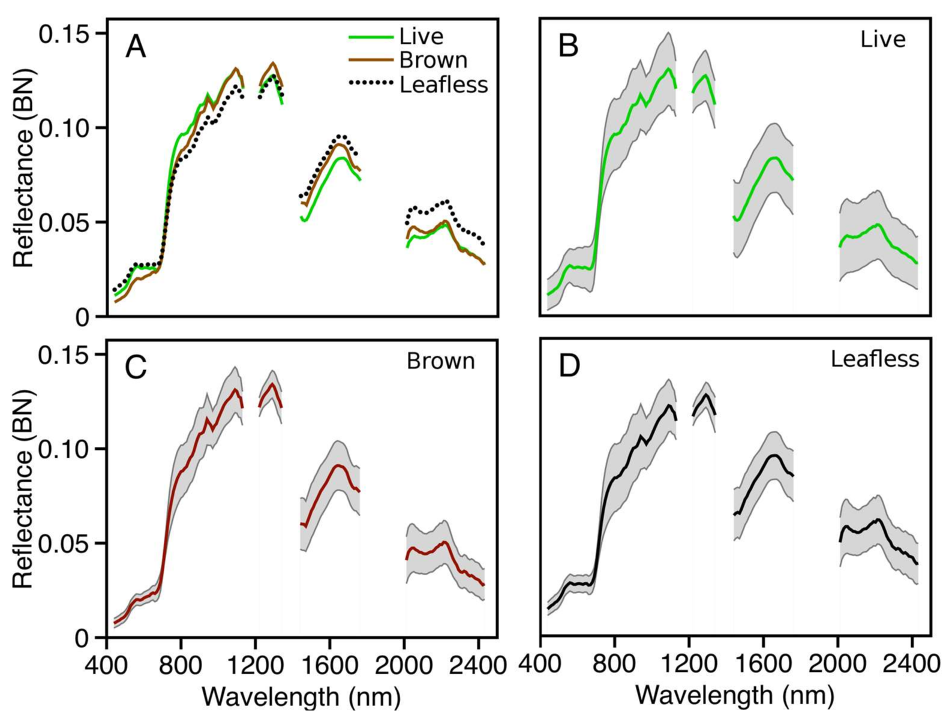
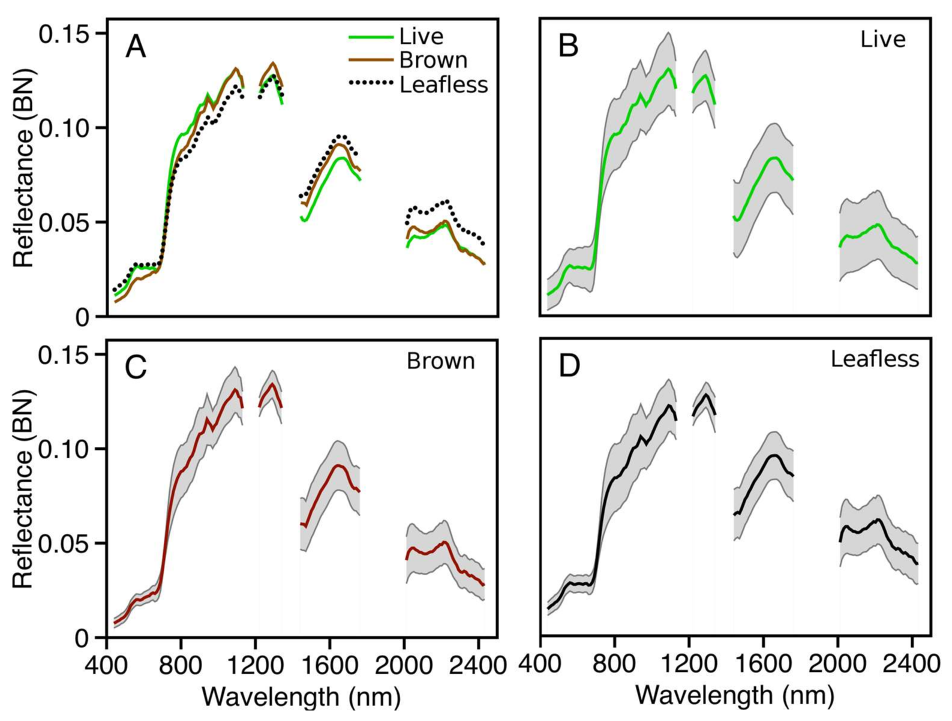
Using orthorectified color images from the on-board digital camera as a reference, we identified 688 VSWIR pixels within crowns exhibiting the brown state of ROD infection, labeled “
Brown”, and 903 VSWIR pixels within leafless crowns, labeled “
Leafless”, across the entirety of ‘ōhi‘a stand distribution on Hawaii Island (
Figure 1). Because signal from background vegetation can have a large impact on observed spectra, especially in leafless crowns, care was taken to select training pixels from central regions of crowns and that tree branching was dense enough to ensure that the resulting spectra would be dominated by the class of interest. We also identified 13,946 pixels representing other entities we were not interested in mapping; these were primarily living canopies of other plant species, along with a small number of non-vegetation targets such as bare ground, manmade structures, and other surfaces, all of which provided the model with a third target class, labeled “
Live”. The non-vegetated pixels make up <1% of the target set; therefore, despite their spectral characteristics being different from each other and living vegetation, we chose to include them in the living category, because we were not interested in separating samples classified into this category. A few shaded pixels were included for each of the three classes to reduce any artifact caused by decreased signal-to-noise ratios in darker pixels.
The full VSWIR reflectance spectrum was extracted for each of these training pixels and labeled by class (
Figure 2). Reflectance values of wavelengths known to be absorbed or scattered by atmospheric constituents were removed, leaving 159 bands representing four regions of the measured spectrum (
Table 1). Cleaned individual spectra were subsequently brightness-normalized (BN), such that the magnitude (Euclidean norm) of each BN spectrum was 1.0. This was done for each spectrum (pixel) using the formula
in which
is the original reflectance spectrum as a vector,
is the brightness-normalized reflectance spectrum.
2.4. Modelling
Both false detections and false omissions have the potential to waste resources when final maps are used in the field; we sought to maximize the model results under dual criteria of high sensitivity and high precision. To meet this goal, we combined two model forms into an ensemble model for differentiating pixels within the
Brown and
Leafless classes from those in the
Live class. The first model form was a Support Vector Machine (SVM), which finds optimal divisions between the data points in n-dimensional vector space [
14]. This machine learning tool has been demonstrated to be an effective classification tool with hyperspectral data [
15,
16] and has performed well with CAO data in particular [
17,
18,
19]. SVMs naturally incorporate information from all wavelengths of each input spectrum and offer the benefit of being computationally simple to train and apply. The second model form used was the Gradient Boosting Machine (GBM) [
20,
21]. GBM models are created by generating an ensemble of fairly weak (shallow) decision trees. Because each successive tree is fit to the remaining residuals from the previous combination of trees, a process known as ‘boosting’, GBMs incorporate both bias and variance reduction, distinguishing them from other decision-tree based approaches such as a Random Forest (RF), which are focused on variance reduction. GBM models are able to incorporate nonlinear interactions between spectral features while also being robust to spectral collinearity. We found GBM models outperformed RF models during exploratory testing in this study.
We found optimal model parameters for both the SVM and GBM fits using a multi-stage grid search technique. The first stage grid covered a broadest region of the available parameter space with the intent of surrounding the most-optimal parameters. The search grid for each successive stage used finer granularity, covering a small region of the grid that proved most optimal in the last stage. A 10-fold cross-validation was performed for each set of parameters in the grid to get a reasonable estimate of how well the model would perform on new data. All model training was done using the package scikit-learn [
22] for Python. As the same tree canopy can simultaneously contain both brown foliage and leafless branches, leaving the canopy state rather ambiguous, we focused less on misclassifications between these two states and more on misclassified living trees. In addition, errors of commission regarding living trees would be much more problematic to users of the final maps. Thus, the criteria used during model parameter selection for both models was the number of correctly classified
Brown and
Leafless pixels (combined) while holding the false positive rate of each of the
Brown and
Leafless classes to less than 1% (independently). Error rates were assessed on a per-pixel, rather than a per-crown, basis because of the relatively large number of small, single-pixel instances in the training data. Once optimal parameter values for each model were obtained, we trained final models with the full set of training pixels using these parameters to build models for subsequent full-island map application.
We next devised a way to combine the two models in order to improve the applied model predictions. Both models, when applied at the whole-island scale, left patches of false-positives for either the
Brown or
Leafless classes due to heavily site-specific vegetation or observation conditions. We sought to eliminate these artifacts, while preserving model accuracy. Initial testing suggested that the SVM model was better at detecting
Brown and
Leafless pixels yet more prone to false positives. We found that the best performance with the least amount of artifact occurred when the GBM model was able to ‘veto’ perceived spurious predictions of the SVM. For each pixel and for each class, each of the two models produced a likelihood value between 0 and 1, which effectively portrays the model’s confidence that a given pixel belongs to a given class. We found that the vast majority of false positives in maps produced by the SVM model had only moderate likelihood values, yet corresponding likelihood values from the GBM model were extremely low. Using this feature to our advantage, any SVM likelihood value for
Brown or
Leafless classes of less than 0.975 was replaced by the associated GBM likelihood, if the GBM likelihood value was less than 0.01. For a given class
, this model combination rule can be written as
in which
is the combined model likelihood value for class
{
Brown,
Leafless},
is the SVM model likelihood value for class
, and
is the GBM model likelihood value for class
. For the likelihood values produced by the above combination approach, still in the range 0 to 1, we used the following decision rule to assign a class to each pixel:
in which
,
, and
are likelihood values for classes
Live,
Brown, and
Leafless, respectively,
is a specified likelihood threshold value, and
C is the predicted classification of a given pixel.
2.5. Model Application
We used a combination of two metrics to evaluate potential values of the likelihood threshold value, , for application of the combined model to full maps of Hawaii Island. For a given value of , we computed classification sensitivity (producer accuracy) and classification precision (user accuracy) for the Brown and Leafless classes using the SVM, GBM, and combined model predictions from the 10-fold cross-validation training under the optimal parameters for each model. We repeated this calculation for each of 19 values, , to produce response curves of both metrics for both Brown and Leafless classes.
Ultimately, we selected threshold of 0.65, with which we applied the combined model decision rule to the model prediction likelihood values of the full CAO VSWIR coverage of the island. Because VSWIR pixels are 4 m2 in area, light reflecting from brown vegetation on the ground can blend with light reflecting from the edges of tree vegetation, causing the spectra for a number of edge pixels to be identified as Brown or Leafless. We filtered out false detections due to such edges by using the canopy height data built from the LiDAR data. If the LiDAR-derived height of the top of the vegetation surface was <1.5 m in any of the four 1 m × 1 m quadrant subregions within a VSWIR pixel, then the pixel was re-classified as Live when building output maps.
2.6. Additional Spectral Filtering
Field verification of early versions of the final map indicated that excessive false-positives remained for the Brown class. There were three scenarios in which a crown in the map was not observed to be a leafless ‘ōhi‘a in the field later in the same year. Each of these scenarios below involved vegetation not represented in the training data sets.
A small number of instances of crowns in the far north region, Kohala, were classified as Brown but observed to be green three months after the CAO flight coverage. The most likely cause of this was that these were ‘ōhi‘a exhibiting unusually heavy flowering (‘ōhi‘a flowers are typically bright red) along with relatively low foliage. This situation may have been exacerbated by an extremely low sun angle at the time of overflight.
In the region of Hawaii Volcanoes National Parks, some instances of tall or climbing brown Dicranopteris linearis fern (uluhe) were found to be misclassified as Brown by the model.
Also in the region of Hawaii Volcanoes National Park, a large number Morella faya (firetree) trees are known to be dying in the area. These trees also appear dry and brown—similar, but not identical, to ‘ōhi‘a in the brown-state of ROD infection—and the model classified map pixels, covering them as Brown with high likelihood values.
We created a filter to reduce detected crowns under these false-positive scenarios using an analysis of the spectra of the affected pixels. The pixels from brown
D. linearis fern and flowering ‘ōhi‘a in scenarios 1 and 2 all had highly-negative slopes in the red-green region of the visible spectrum (600–680 nm), whereas brown ‘ōhi‘a and
M. faya both consistently had positive slopes in the same spectral region. We therefore used the following criteria to identify and remove such pixels from the map.
in which
is the reflectance at the indicated wavelength
in nm.
M. faya pixels classified as
Brown (scenario 3 above) were separable from verified
Brown ‘ōhi‘a pixels using a combination of two features: (1) the peak height difference between the infrared and first shortwave regions of the spectrum was greater for ‘ōhi‘a pixels and (2) the slope of the peak in the first shortwave region (1640–1680 nm). We employed the following criteria to identify and remove brown
M. faya pixels from the map.
While we do not believe any examples of brown D. linearis fern, brown M. faya, or flowering ‘ōhi‘a were represented in the training data, some of the Brown training pixels were caught by one or more of the two spectral filter criteria. For the purposes of evaluating map accuracy, it was unclear how to represent these ambiguous pixels in a confusion matrix. Applying the filter criteria to just the predicted classes would show degraded mapping performance over the pre-filter results, while applying the filter criteria to both observed and predicted classes would show improvement. We therefore simply kept track of the number of pixels affected, and performed an analysis of the potential effects of this filtering on model sensitivity of Brown pixels.
We made final maps of the locations of suspected ROD-infected brown ‘ōhi‘a tree crowns (as clusters of predicted Brown pixels) and leafless tree crowns (as clusters of predicted Leafless pixels) from the remaining pixel-level detections. Centroid points of all contiguous, adjacent (no diagonal connections) clusters of Brown or Leafless pixels greater than or equal to two pixels (8 m2) in size were used as spatial coordinates for suspected brown-state ‘ōhi‘a or leafless tree crowns. Although only a small number of points were found in areas known to not contain ‘ōhi‘a, these points were considered false detections and were therefore masked out in the final maps.
3. Results
We found both the SVM and GBM models to be capable of detecting a high number of Brown and Leafless training pixels with minimal false detection. Use of a radial basis function (RBF) for the SVM model yielded the best results. Under this kernel, the optimal parameter values were a gamma of 0.0075 and cost of 200. For the GBM approach, optimal parameter values were a learning rate of 0.225, a maximum tree depth of 6, and the assembly of 1750 trees.
Cross validation results indicated that the SVM was more sensitive to spectral features that define both
Brown and
Leafless classes (
Table 2). Sensitivity values were 90.1% and 88.0% for
Brown and
Leafless classes, respectively. The sensitivity rate for both these classes combined, compared to the
Live class, was 90.5%. GBM was considerably more conservative during training, with many more false-negatives for the
Brown class, though it seemed to be slightly better at distinguishing
Brown from
Leafless pixels. The GBM
Brown sensitivity was 70.8%,
Leafless sensitivity was 65.1%, and combined
Brown and
Leafless sensitivity was 69.5%. Upon application, the SVM was much more prone to erroneous detections of both
Brown and
Leafless pixels due to lighting or forest canopy conditions not represented in the training dataset.
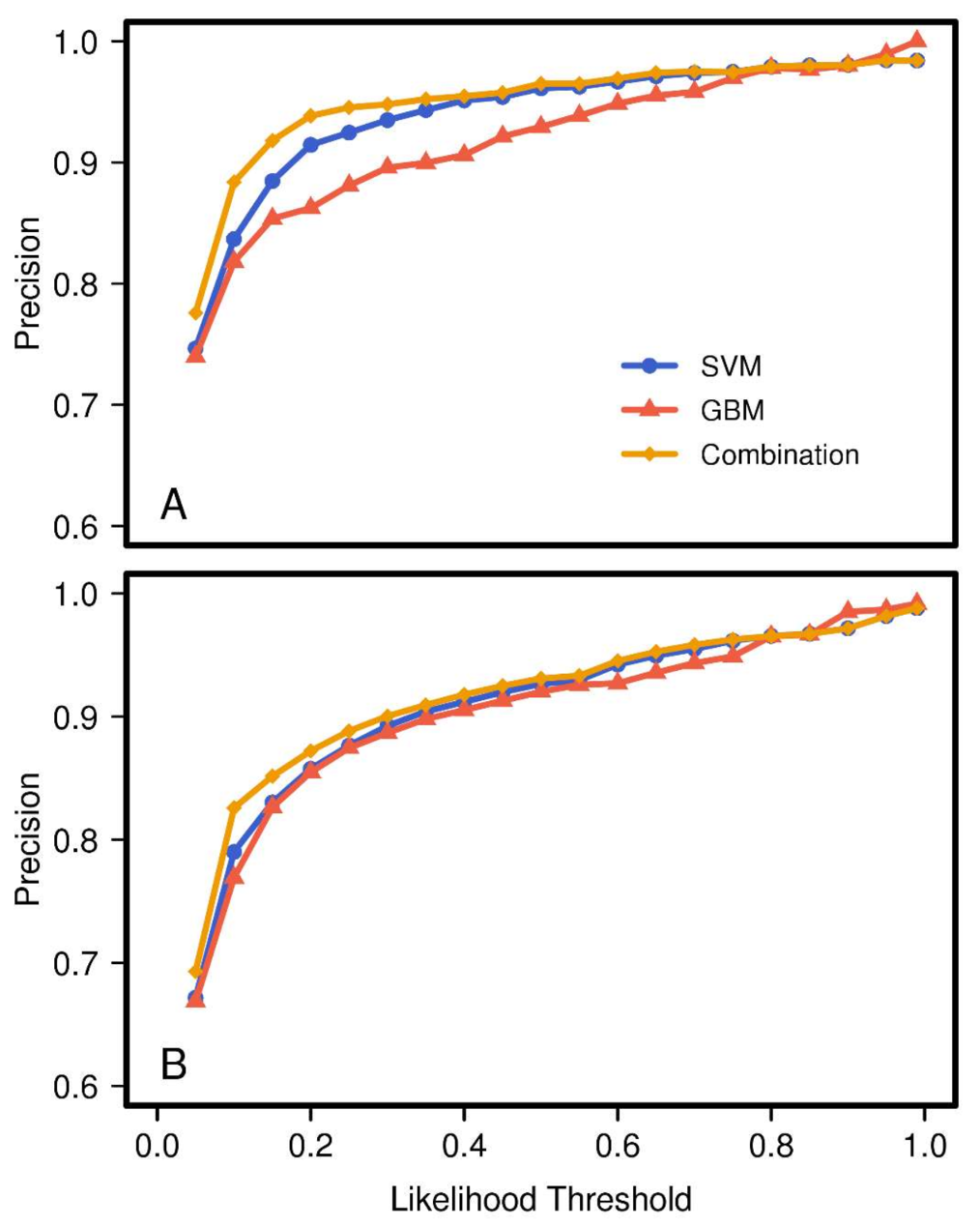
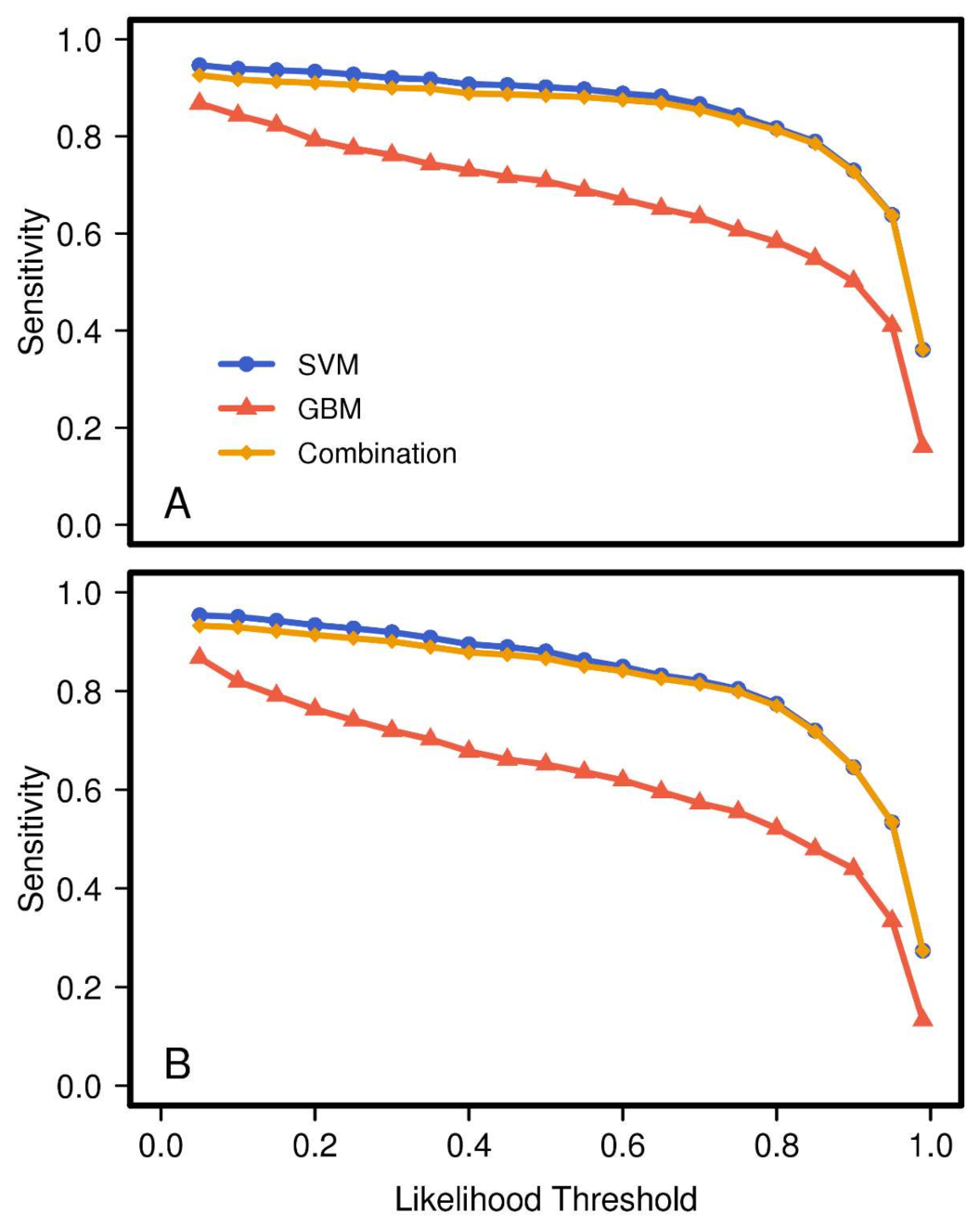
When analyzing the effect of threshold value on the different models, we found a generally slow decline in true detection rates for all models as the likelihood threshold
increased from 0 to approximately 0.8 (
Figure 3). Thus, selection of any value within this range will still result in a high number of true positives. Additionally, the rate of decline was significantly lower for the GBM model compared to the SVM model. This analysis showed that the combined model retained nearly all of the superior sensitivity of the SVM model, indicating that for most of the truly
Brown or truly
Leafless training pixels the GBM was in at least partial agreement.
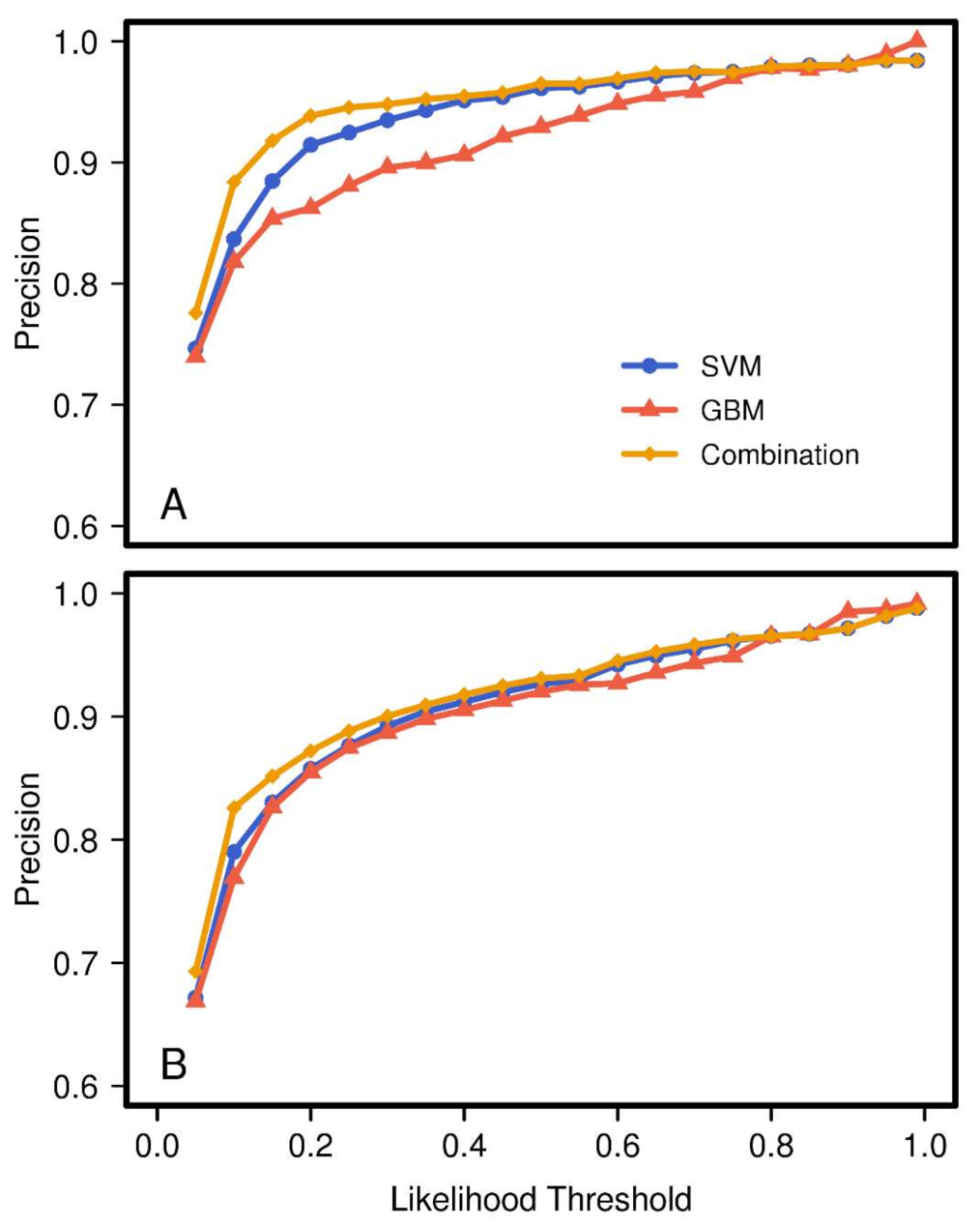
Similarly, precision rapidly increased with
to >90% for all models in the range 0 to ca. 0.4 (
Figure 4). However, compared to the values for sensitivity, the difference between models was not as large. The rate of precision for
Brown pixels was lower for the GBM than for the SVM. However, with respect to
Leafless pixels, the two models had nearly the same values for precision. We found the combined model not only kept the properties of the best of its component models, it improved upon them. Here, the combined model
Brown pixel precision values most rapidly improve with increasing threshold, reaching a semi-plateau by a threshold value of 0.2. To a lesser degree the same was true for
Leafless pixels.
Given the above results, and upon inspection of several sample map areas, we selected a likelihood threshold of 0.65. When applied to the combined model, this threshold maintained desirable detection rates (i.e., sensitivity values of 86.9% and 82.5% for
Brown and
Leafless pixels, respectively), while greatly reducing the false positives (i.e., precision values >95% for both classes) to acceptable levels (
Table 3). The kappa statistic for this classification was 0.895. Only 42 of the 13,946
Live training samples were misclassified as class
Brown or
Leafless, indicating the model is more likely to classify ambiguous trees as
Live.
The spectral filtering to remove D. linearis fern, M. faya tree, and flowering ‘ōhi‘a affected a noticeable number of Brown training pixels. We found that 48 of the 688 Brown training pixels met the criteria for one or both of the two spectral filters (Equations (1) and (2)). Thus, assuming our training pixels truly contain no examples of D. linearis fern, M. faya tree, or flowering ‘ōhi‘a, the model may lose up to 7.0% of Brown pixel sensitivity by applying these filters. However, 19 of these 48 pixels were not even identified as Brown by the combined model before filtering, indicating the actual loss in sensitivity might be closer to 4.2%. Of the 8316 Live pixels that met one or more of the filter criteria, none were mistakenly classified as Brown. This indicates that, outside the region of Hawaii Volcanoes National Park, this particular type of misclassification is probably rare.
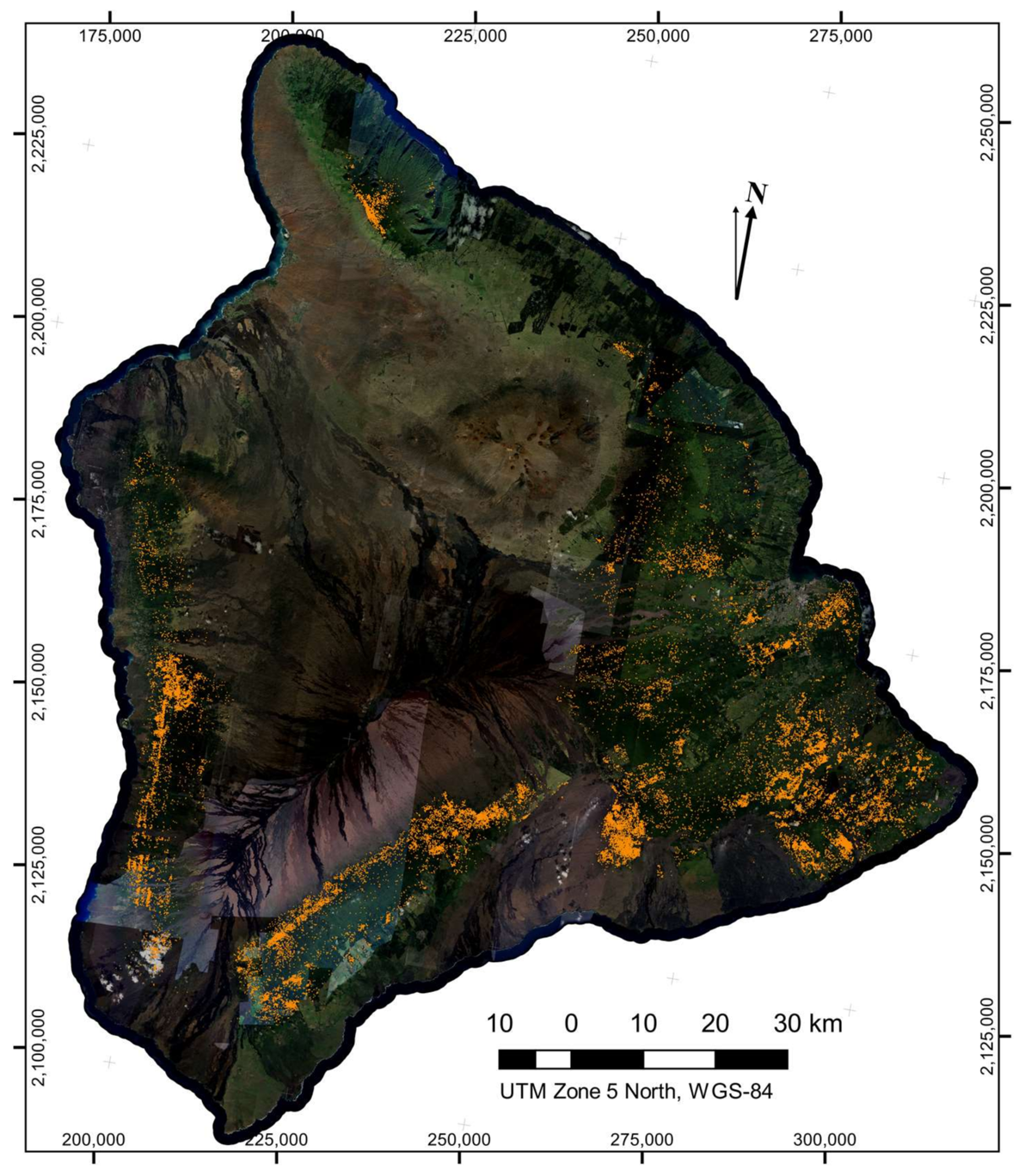
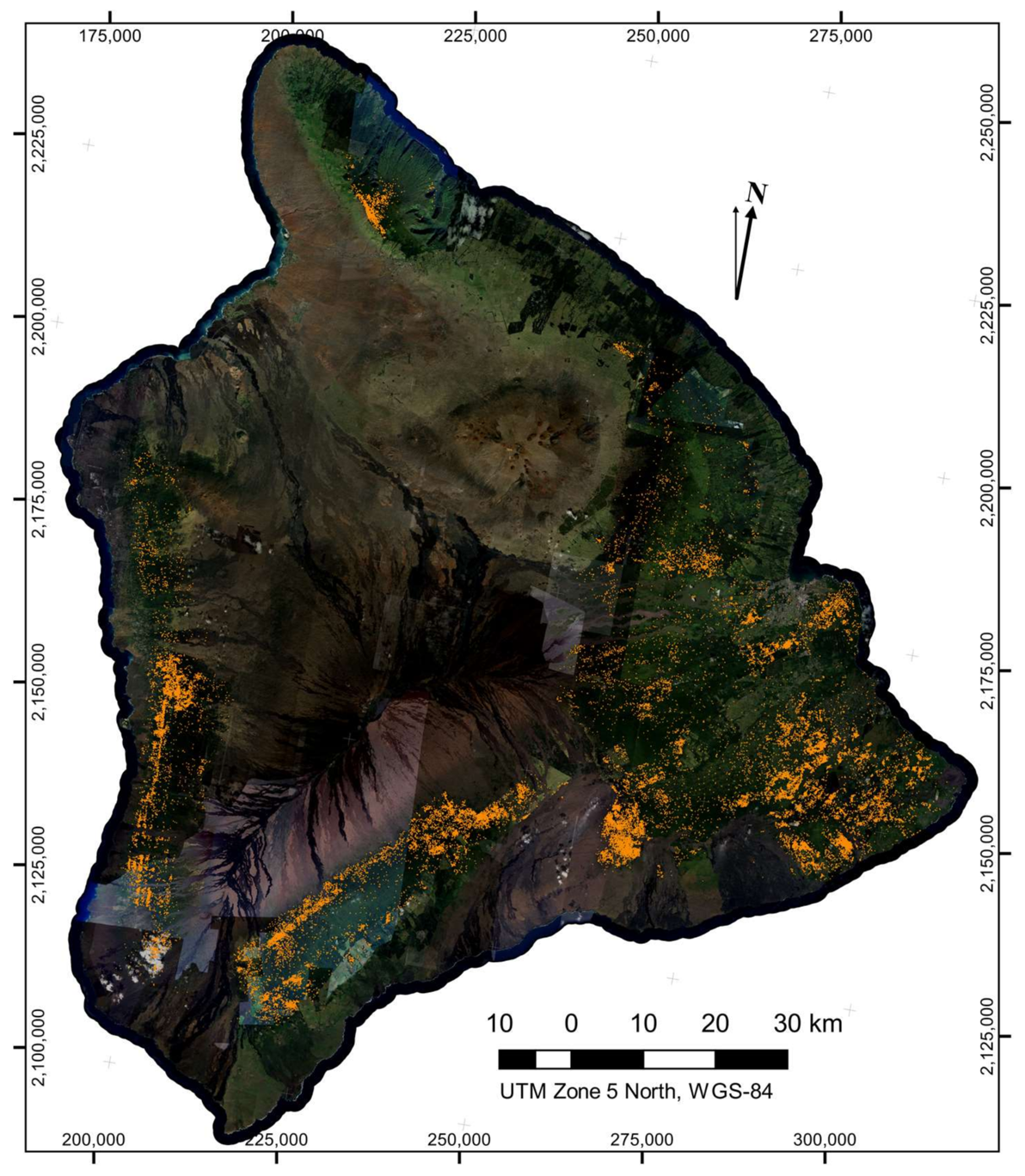
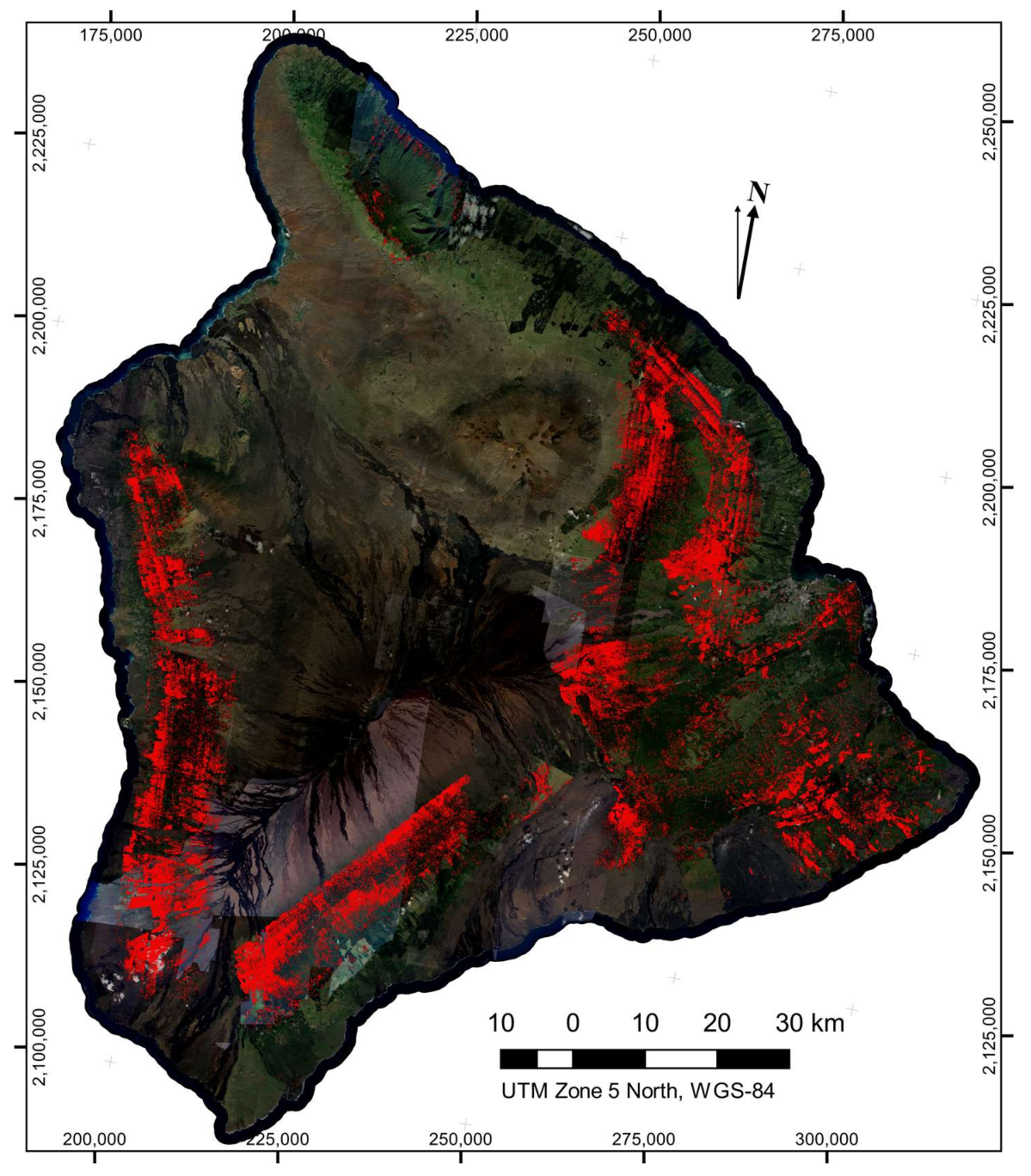
The LiDAR-derived canopy height and the spectral filtering both reduced the number of isolated
Brown pixel detections, resulting in a statistically robust map with very little visual artifact present (
Figure 5). Across the island of Hawaii, we found 43,134 individual crowns suspected of exhibiting the brown state of ROD infection. Hotspots of potential ROD infection are apparent in the maps. The peninsula on the eastern side of Hawaii known as the Puna district, where the ROD outbreak likely originated, contained a very high density of brown-state crown detections.
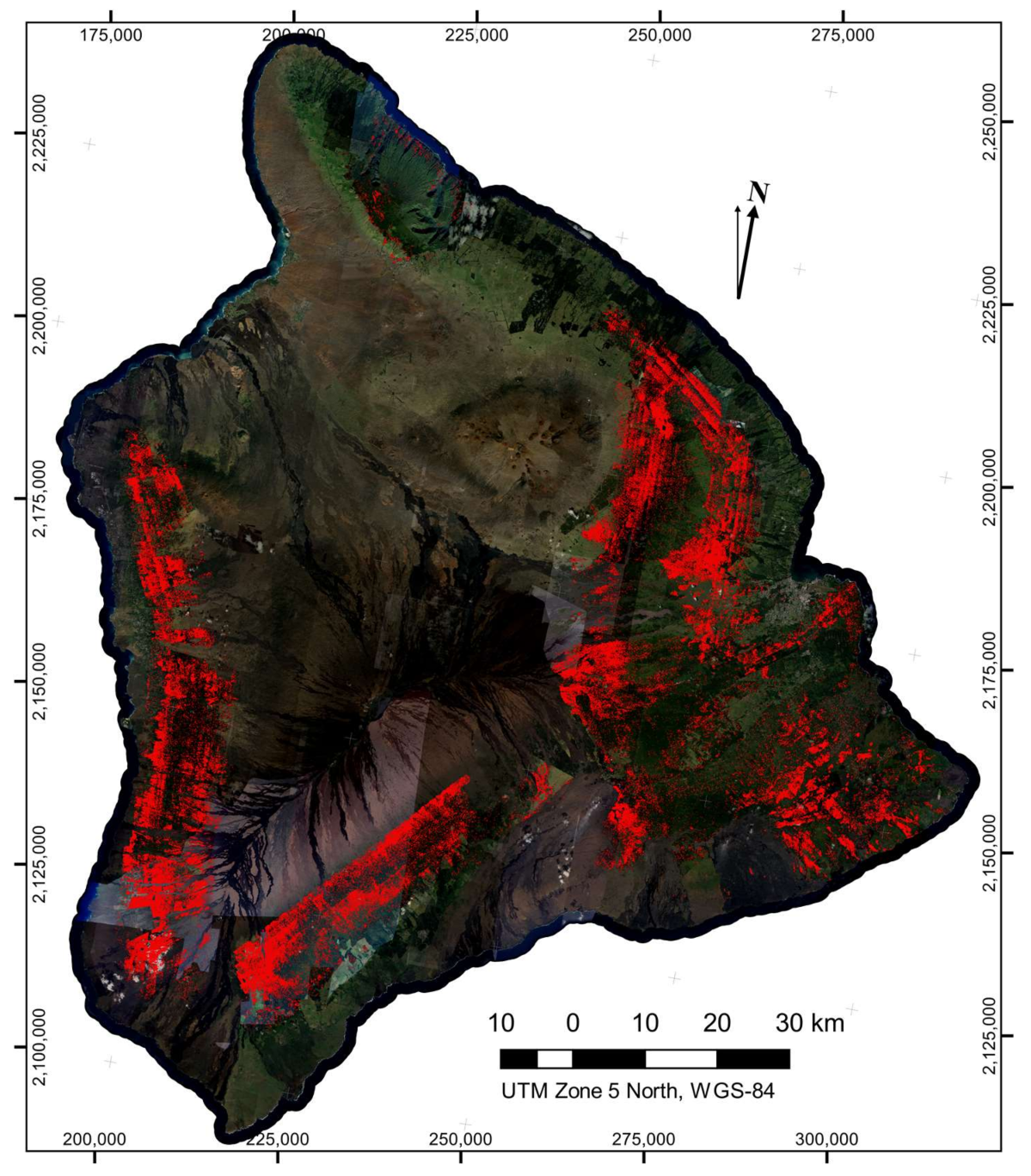
In comparison, leafless crown detections were much more numerous (547,666 detected leafless crowns in total) and more dispersed across the island (
Figure 6). It was apparent in the map (
Figure 6) that detection of leafless crowns was more sensitive to cross-flightline observation angle effects, leading to some linear artifact in the maps. However, we saw no evidence of an increase in false-positive rates of leafless crowns due to these viewing angle effect, but rather we observed that a decrease in detection on one side of each flightline was the cause of the artifacts. As viewing angle and solar angles approach nadir, green undergrowth beneath a thin, leafless crown has a more dominant effect on reflectance, leading to this decreased detection. Since we were unable to distinguish the cause of death for so many trees, we cannot ascertain what percentage of leafless crowns were the result of ROD. However, dense areas of detected leafless crowns can be found in regions containing higher counts of brown-state crown detections, and ROD is expected to be the dominant cause of death in these locations.
4. Discussion
Using CAO laser-guided imaging spectroscopy, we obtained very high classification accuracy for detecting brown-state, potential ROD-infected, and leafless trees with both the GBM and SVM models and their combination. This suggests that the spectra of these two classes were highly distinguishable from surrounding vegetation. While it was possible to visually identify particular brown and leafless crowns with certainty in the three-band (red, green, and blue) 24 cm aerial imagery due to the location and pattern of occurrence, this was not the case for all images captured across the island. Due to differences in lighting conditions and aircraft movement, our RGB imagery was insufficient to determine whether some trees were truly exhibiting the brown state of ROD infection. This is why hyperspectral data are often sought for difficult classification problems [
7,
23,
24]. As their canopies turn brown, ROD-infected ‘ōhi‘a trees also experience a characteristic change in leaf chemical composition and shape, both of which highly affect the reflection of light across the full spectrum measurable by imaging spectroscopy [
25] (
Figure 2).
The ensemble approach of mixing two model forms greatly improved whole-island brown-state and leafless detection maps. SVM models have been used extensively in remote sensing classification applications [
16], and GBM models have more recently shown promise in the field [
18,
26,
27]. Yet, individually, each of the two model forms was prone to misclassification when applied to regions of the island not represented in the training data. An increase in training data would likely help address this issue, but generating a training set that completely spans the extremely diverse climate and biogeographical conditions represented on the island of Hawaii [
28,
29] presents a major challenge. Fortunately, each model form was overactive in different parts of the island and under different conditions. We were able to take advantage of this effect by combining the models into an ensemble. Ensemble models in general have been shown to improve performance over individual component models [
30,
31,
32]. In fact, a GBM itself is a form of ensemble modeling, as classification trees are sequentially added during boosting to incrementally improve fit [
33]. Additionally, more diversity in the component models of an ensemble has been shown to improve ensemble model performance and further reduce overfitting [
34]. Here, we mixed two distinct model forms and were able to reduce overfitting (manifested most-critically as artifacts in the final maps) while retaining high target detection capabilities.
The range of spatial scale of this project was extreme in the sense of applied geospatial classification models. VSWIR data collected by the CAO in 2017 covered an area of approximately 525,000 ha, about 320,000 ha of which is considered forested. This suggests that our maps cover nearly 800 million pixels potentially able to contain infected ‘ōhi‘a. There are few examples of classification maps covering such a large area at high spatial resolution. This is not surprising given the challenges encountered in such a project. In addition to the challenging problem of minimizing flight coverage costs subject to unpredictable tropical weather conditions, merging data from numerous flights occurring under diverse lighting conditions is quite difficult. The boundary between two different collections can easily result in visible artifacts in maps [
35,
36]. Neighboring areas may be covered by flight lines collected under extremely different conditions. Additionally, effects of the bi-directional reflectance distribution function (BRDF) may significantly alter the shape of recorded spectra [
37] and affect model predictions, although the “pushbroom” style of collection of the VSWIR instrument essentially keeps this a cross-track problem. In this project we chose to handle these challenges with a custom flight line mosaic scheme, manual cloud masking, and most importantly, training data identification across regions of diverse lighting and view angle conditions. While not entirely free of viewing angle artifact (see
Figure 5 and
Figure 6), the resulting maps of the crown classes of interest, with surprisingly little artifact, across nearly a full month of data collection.
We chose a pixel-based approach for mapping brown and leafless trees, primarily because our target of interest was partial crowns. ROD symptoms typically spread from whole branch to whole branch [
8]; as such, we believed a focus on pixel-sized was essential to keep model sensitivity high and consistent across the landscape. Working at the crown level was an alternative approach to this problem, and there are several tools available for the automatic delineation of individual tree crowns from LiDAR data [
38,
39,
40], but had we incorporated this methodology, there would have been several setbacks. First, these tools are most-often used for small regions of fairly simple, consistent vegetation growthform (e.g., boreal forest). In contrast, the variation in growthform among living ‘ōhi‘a across Hawaii Island is incredibly high. Adding more variation from dead or dying trees would only increase this complexity. In addition, there are many instances of multiple tree crowns blending together into a single uniform canopy, which severely limits the efficacy of individual crown segmentation [
38,
39].
Our resulting maps detailing the extent and distribution of both brown-state crowns and leafless crowns across broad regions of Hawaii Island reveal a startling extent of mortality. The extent and distribution of brown-state canopies presented here agree well with Hawaii Island-wide helicopter surveys conducted in January 2017 that estimated that ca. 30,000 ha of ‘ōhi‘a forest stands had recently experienced levels of
Ceratocystis-induced mortality >10% (unpublished data, Hawaii Department of Land and Natural Resources). Although approximately half a million leafless crowns documented across Hawaii Island cannot and should not be uniformly attributed to
Ceratocystis-induced Rapid ‘Ōhi‘a Death (ROD), it is likely that the majority of the 43,134 trees exhibiting brown canopies in our 2017 imagery were experiencing the lethal effects of such
Ceratocystis infection. Furthermore, these brown-state canopies constitute relatively recent mortality, certainly within the last year, and likely within 6 months prior to image collection. We employed numerous measures to reduce false-positive detection rates, such as a high likelihood threshold of 0.65 for class inclusion and minimum cluster size of 8 m
2. These measures likely affect small trees more than large trees. However, this may not be a large problem, because prior results suggest that medium-sized trees are the most affected by ROD [
8]. In addition, instances of isolated small brown-state trees should be minimal, because the disease spreads so quickly [
9]. If small individual crowns were not detected, we feel it is very likely that many infected crowns in the surrounding area were detected, and final maps should accurately reflect the relative density of ROD infection.
A prior, well-acknowledged, and well-studied process of ‘ōhi‘a mortality characterized as “‘ōhi‘a dieback” was responsible for high levels of mortality across an estimated 50,000 ha of windward portions of Hawaii Island during the thirty years between the mid-1950’s to the mid-1980’s [
41]. By comparison,
Ceratocystis-induced ROD has affected more than half that amount of area (i.e., 30,000 ha) in approximately one-tenth the time. Our brown-state tree crown maps clearly indicate that mortality from the disease has spread well beyond the area of infection that in 2014 was restricted to the easternmost Puna District of Hawaii Island; currently, ‘ōhi‘a stands experiencing
Ceratocystis-induced mortality are found on each of the five main volcanoes of the Island. Preliminary results from forest inventory plot data indicate that, on average, ‘ōhi‘a stands in which infected trees are present exhibit annual mortality rates of 10%, with some stands experiencing rates as high as 42% [
42].
The final brown-state and leafless crown maps will support efforts to combat ROD throughout Hawaii Island, and on other islands if the disease is transported westward. Final maps show several hotspots where either the extent or the existence of infection was previously unknown. Recent developments have allowed the cause of death of these crowns to be rapidly confirmed as ROD using field kits that can genetically screen for
Ceratocystis fungus [
43]. The location of confirmed hotspots of ROD infections can help pathologists, ecologists, and land managers to better understand mechanisms of how ROD spreads, what regions are most vulnerable, and where and how to best allocate resources to reduce damage [
8]. Additionally, research is underway to determine the feasibility of restoring ‘ōhi‘a to infected areas using genetic resistance techniques [
44]. These maps will help to identify examples of ‘ōhi‘a trees that are that may be resistant to the
Ceratocystis species causing the disease. As a single snapshot in time, these maps integrate the past progression of ROD across the ‘ōhi‘a forests of Hawaii Island. However, remapping the island using laser-guided imaging spectroscopy will enable a more-refined and up-to-date understanding of the rate of ROD proliferation. While the general methodology presented here can be readily transferred to such a remapping or any new applications, several specific facets of this methodology should be transferred with care. Just as highly-optimized models should not be blindly applied to new data—whether they come from different instruments, different time periods, or collected over different locations—the transferability of specific values in the classification rule or in the different post-processing filters will need to be reevaluated during each application as well.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}