Modeling the Directional Clumping Index of Crop and Forest

 , ,
, ,
Abstract
:
1. Introduction
2. Modeling and Evaluation
2.1. Analytical Method for Crop and Forest
2.1.1. Crop Model
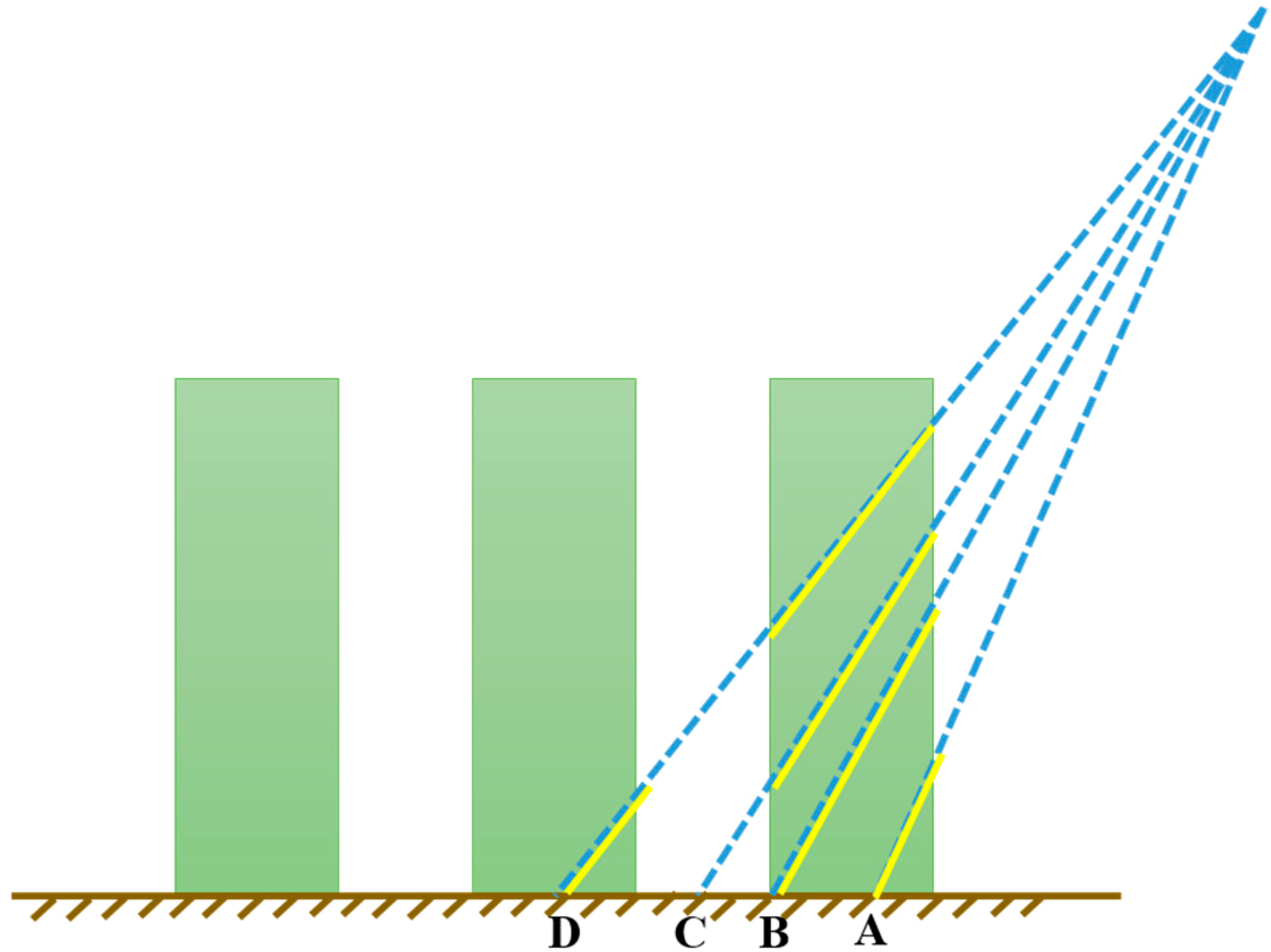
2.1.2. Forest Model
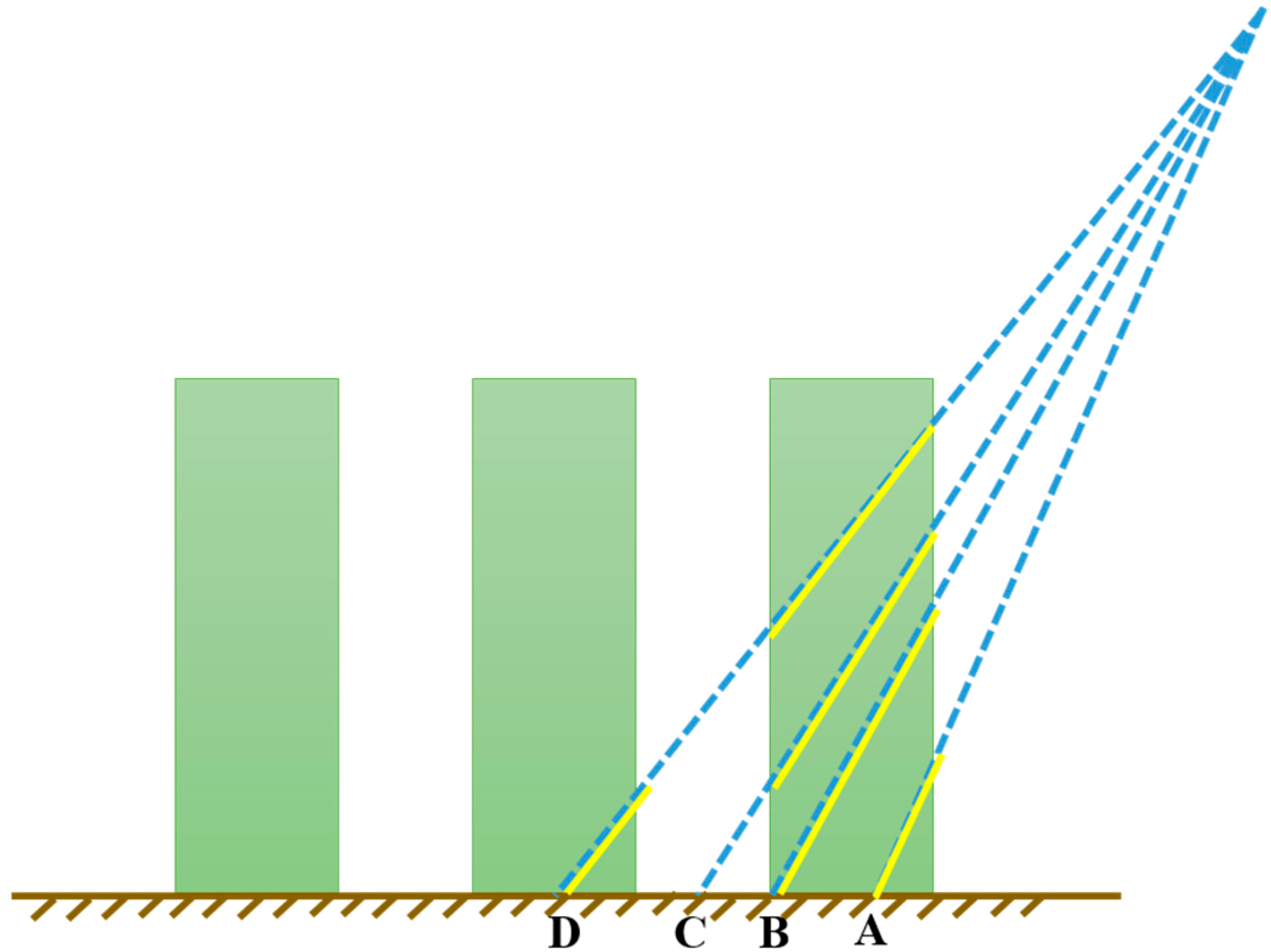
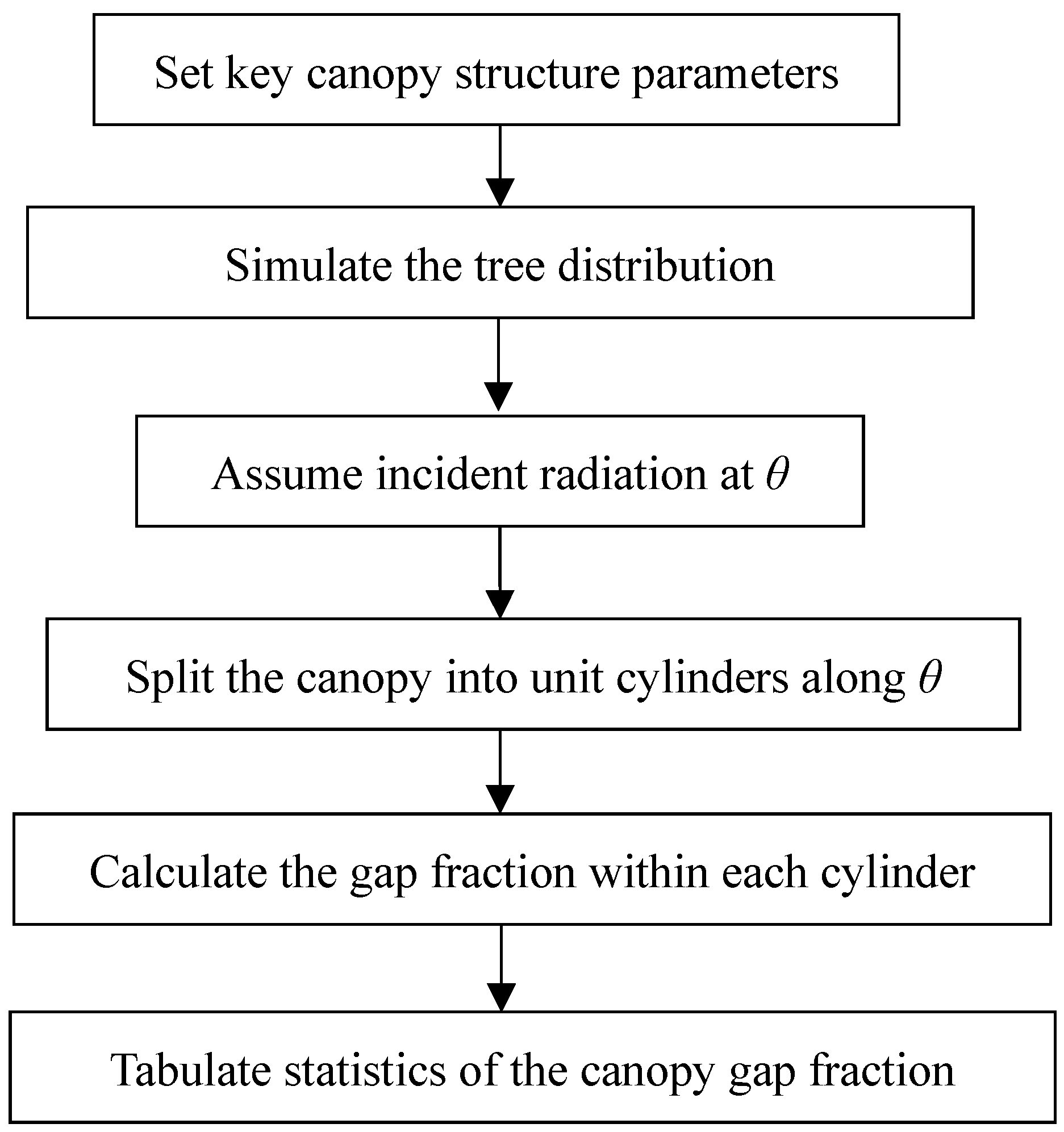
2.2. Simulation Method of Directional Clumping Index
2.2.1. Crop Directional Clumping Index Simulation
2.2.2. Forest Scenario Settings
2.2.3. Forest Directional Clumping Index Simulation
2.3 Forest Ground Measurements
3. Results and Evaluation
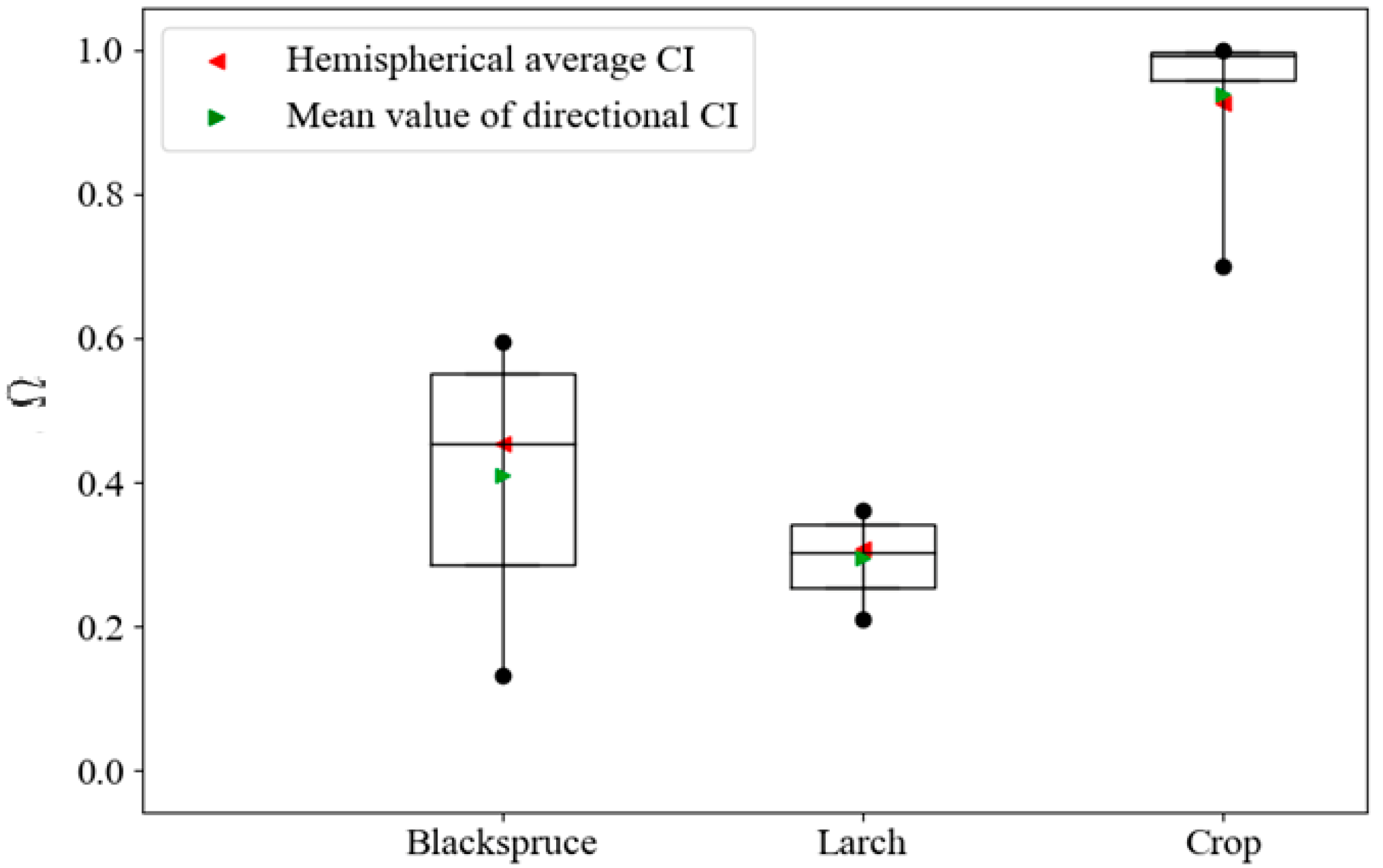
3.1. Crop Directional Clumping Index Validation
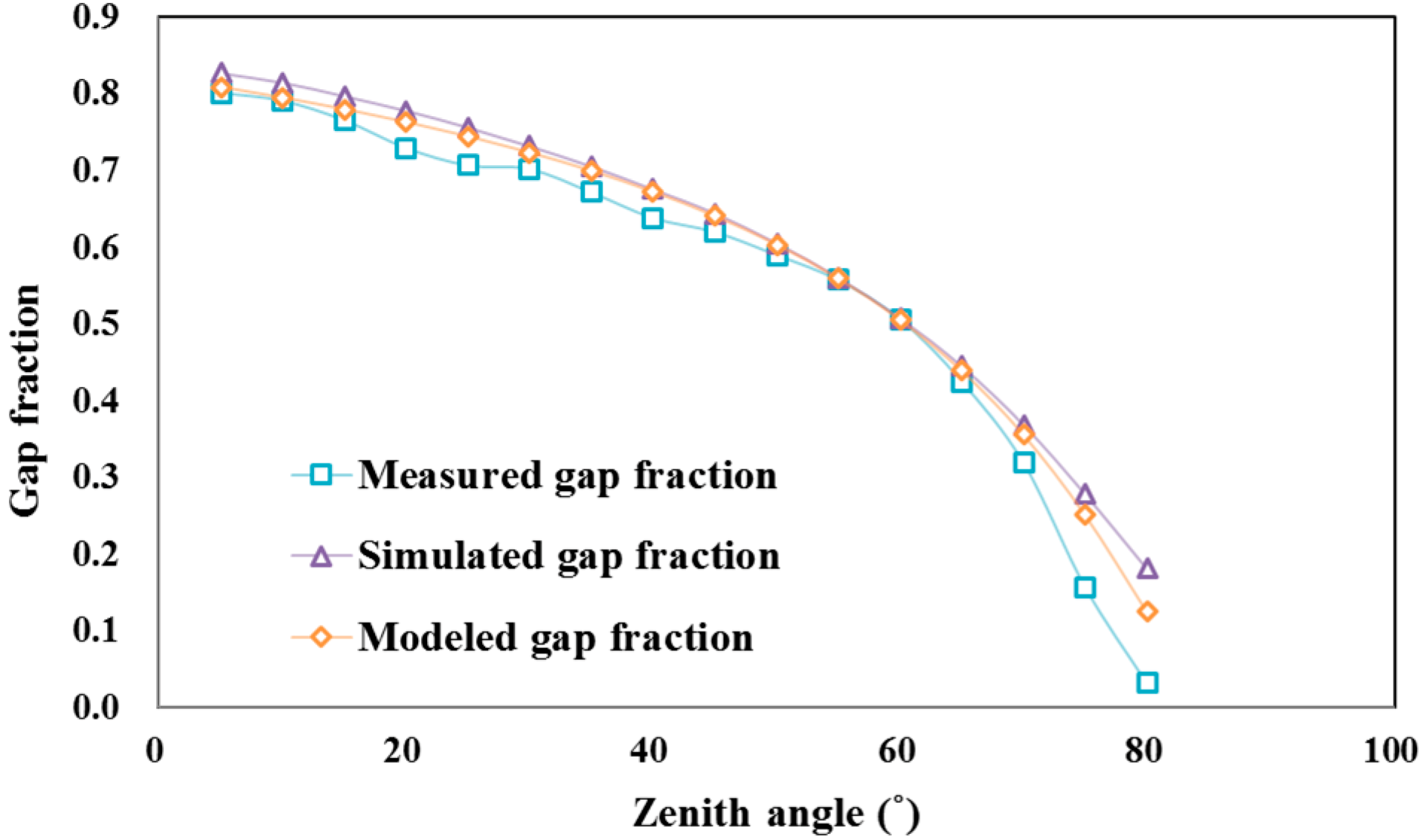
3.2. Forest Directional Clumping Index Validation
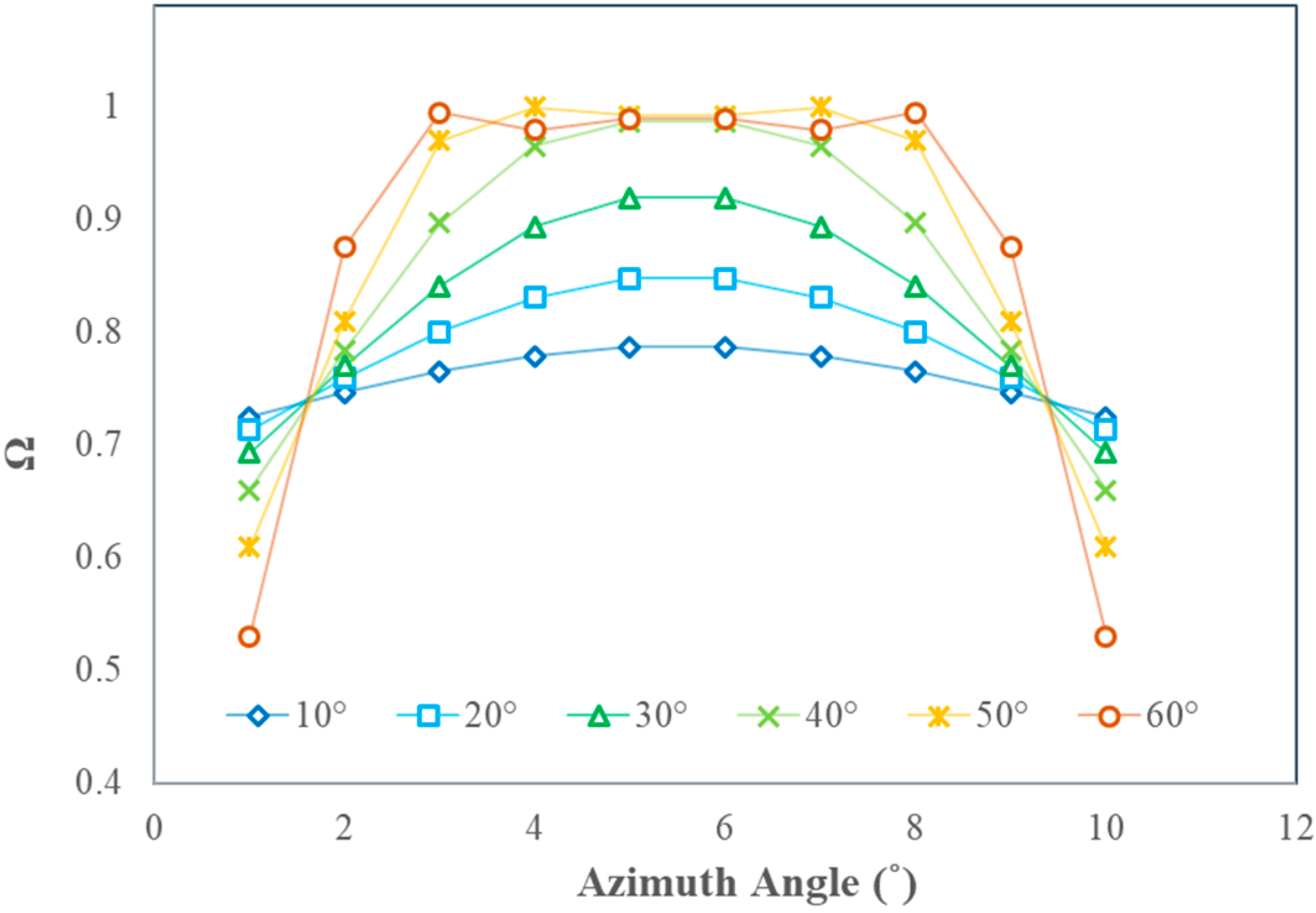
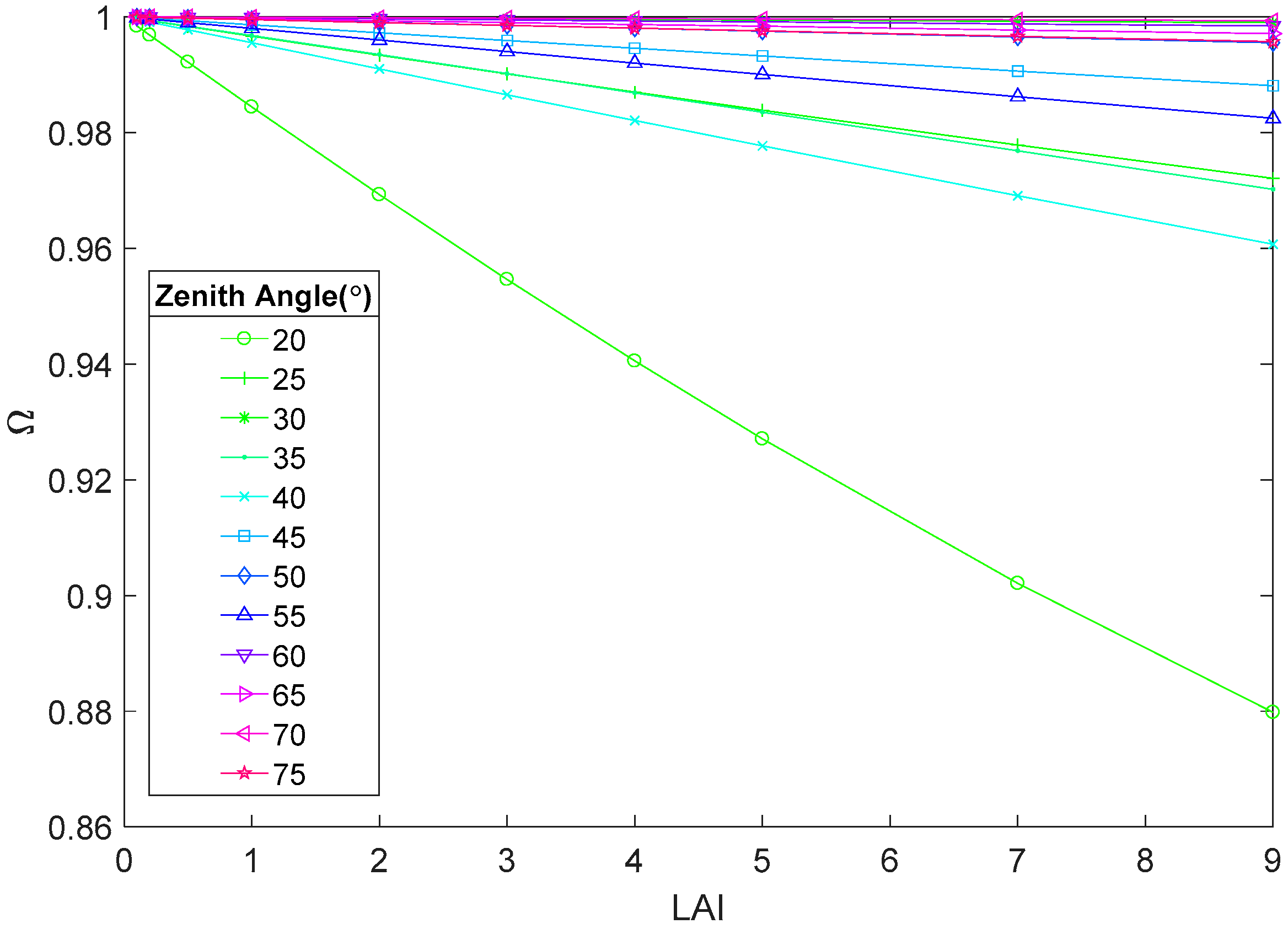
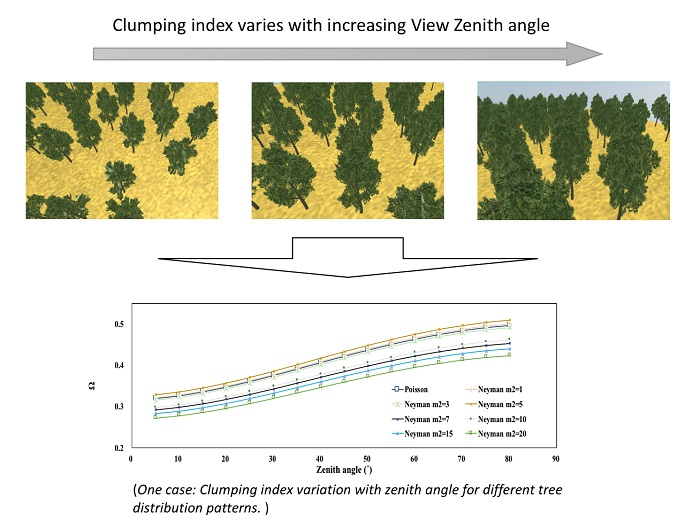
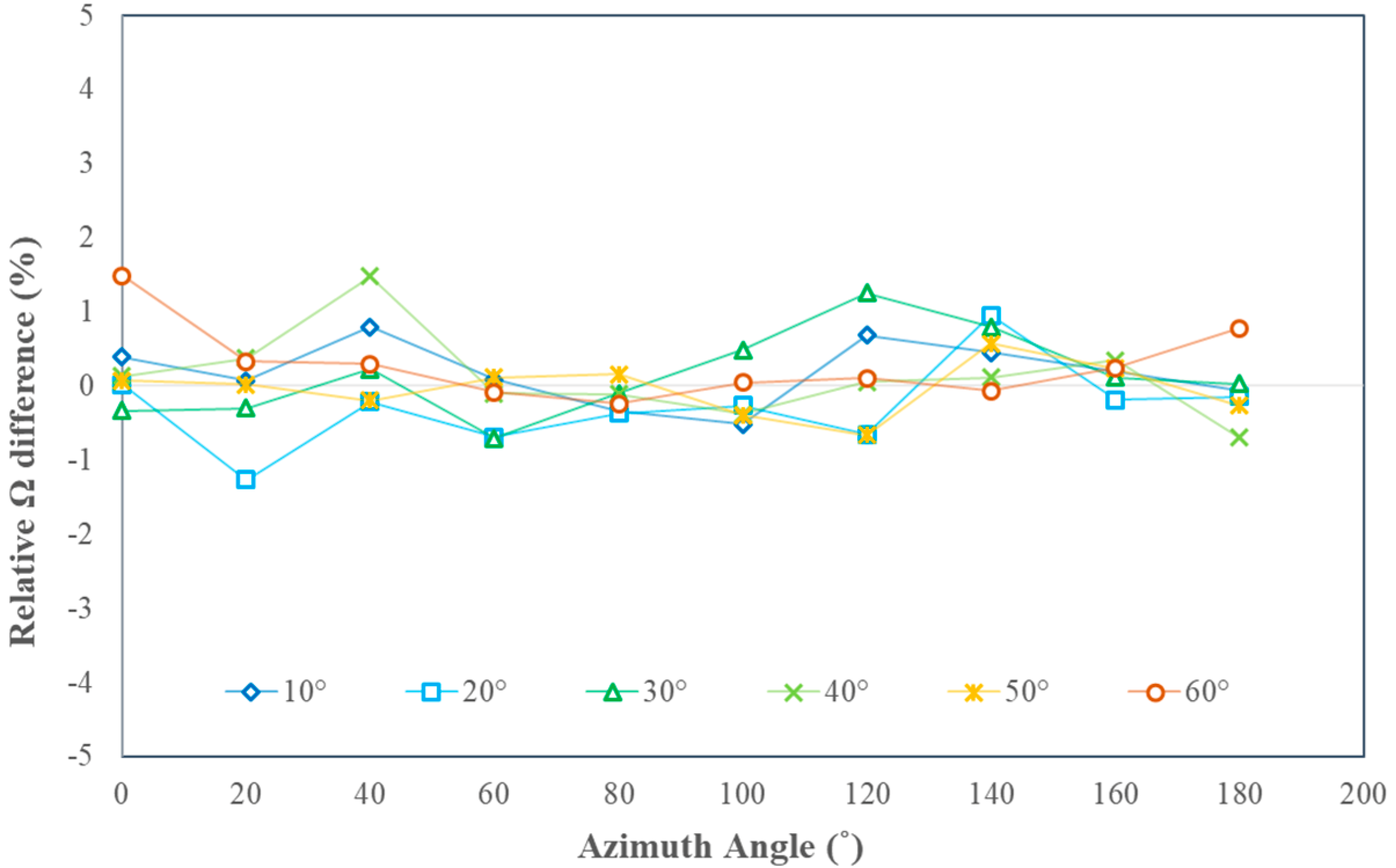
3.3. Assessing the Variation Magnitude of Ω(θ)
4. Sensitivity Analysis
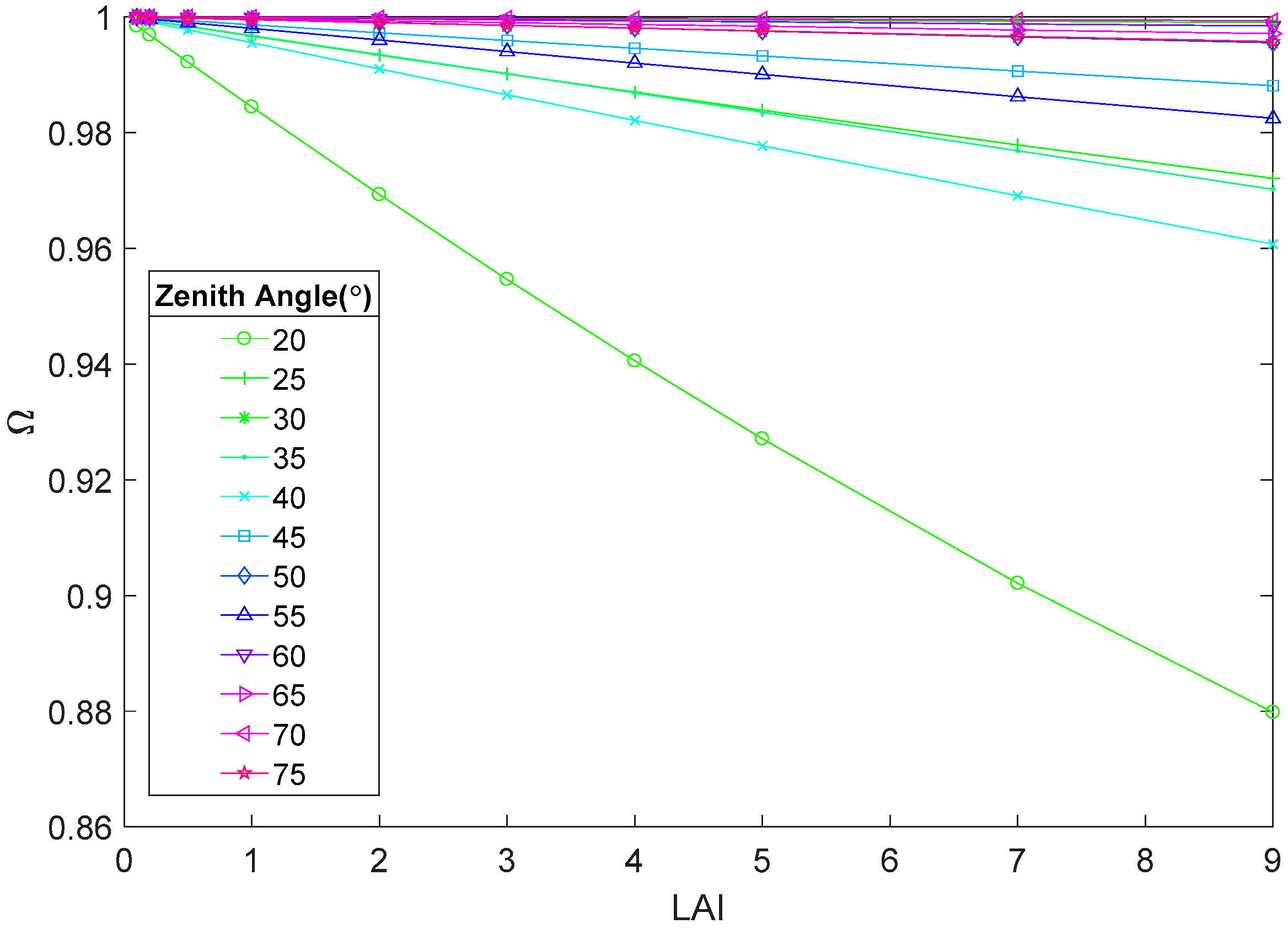
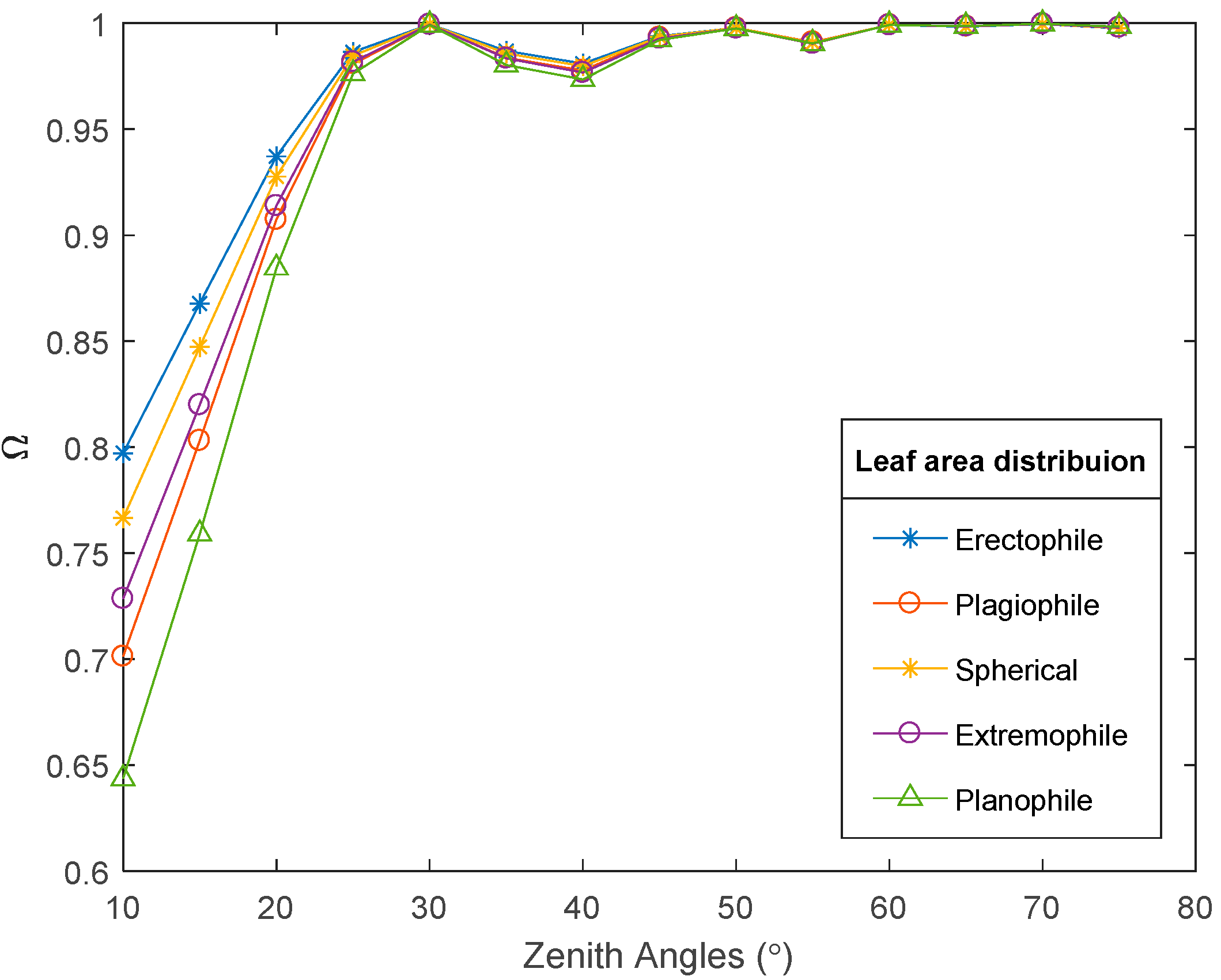
4.1. Factors Influencing Crop Clumping Index Variation
4.1.1. Leaf Area Index
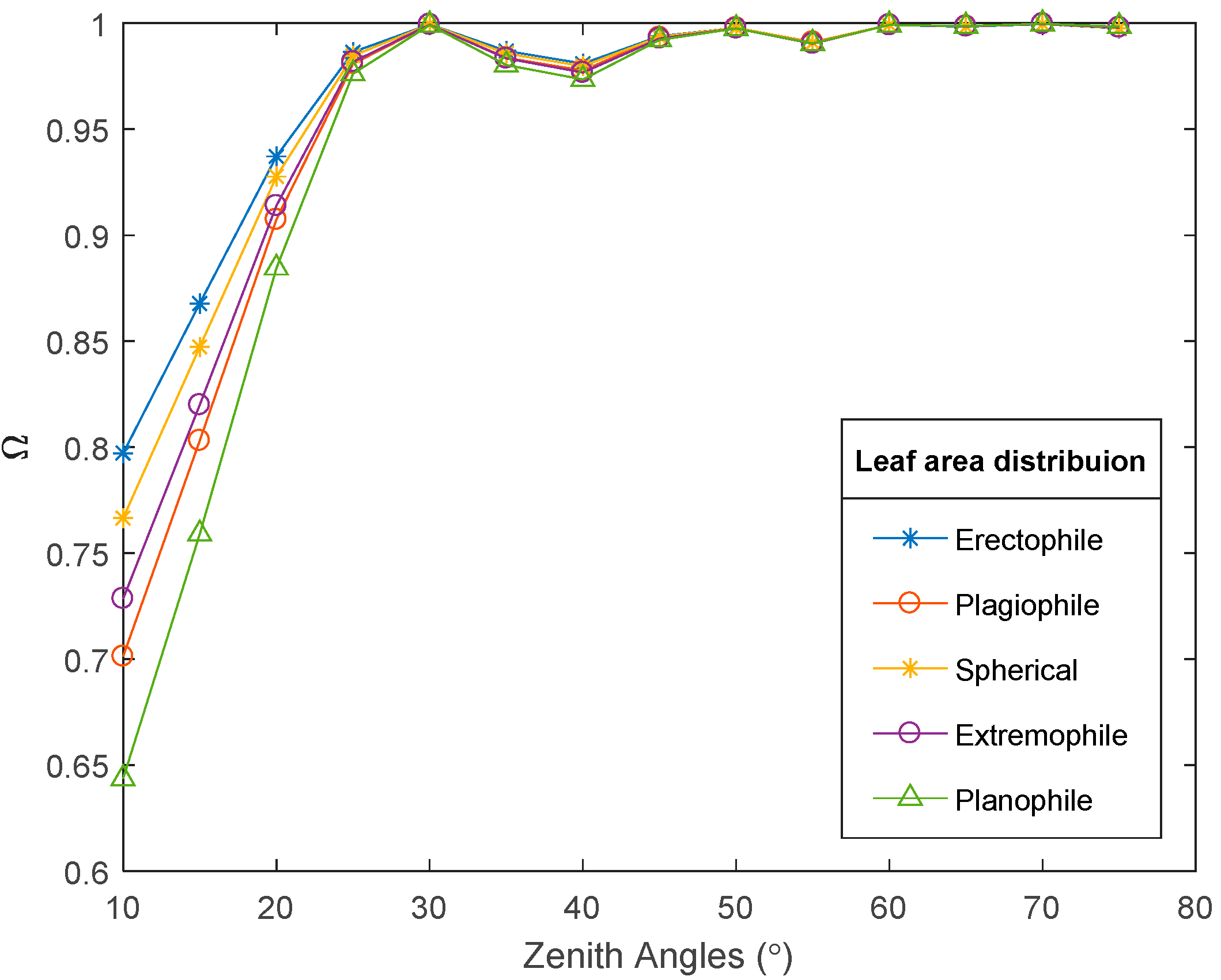
4.1.2. Leaf Angle Distribution
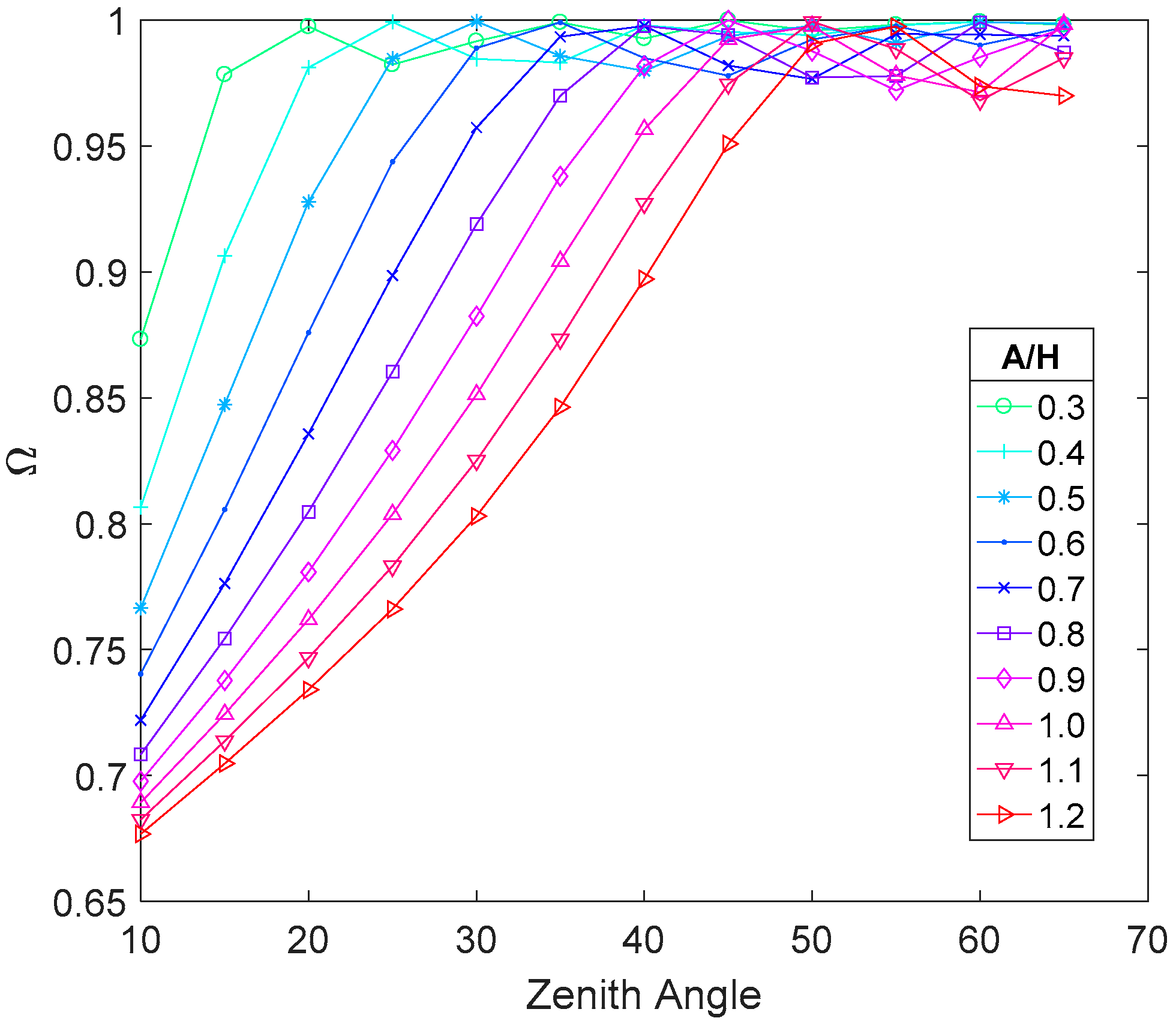
4.1.3. Row Structure
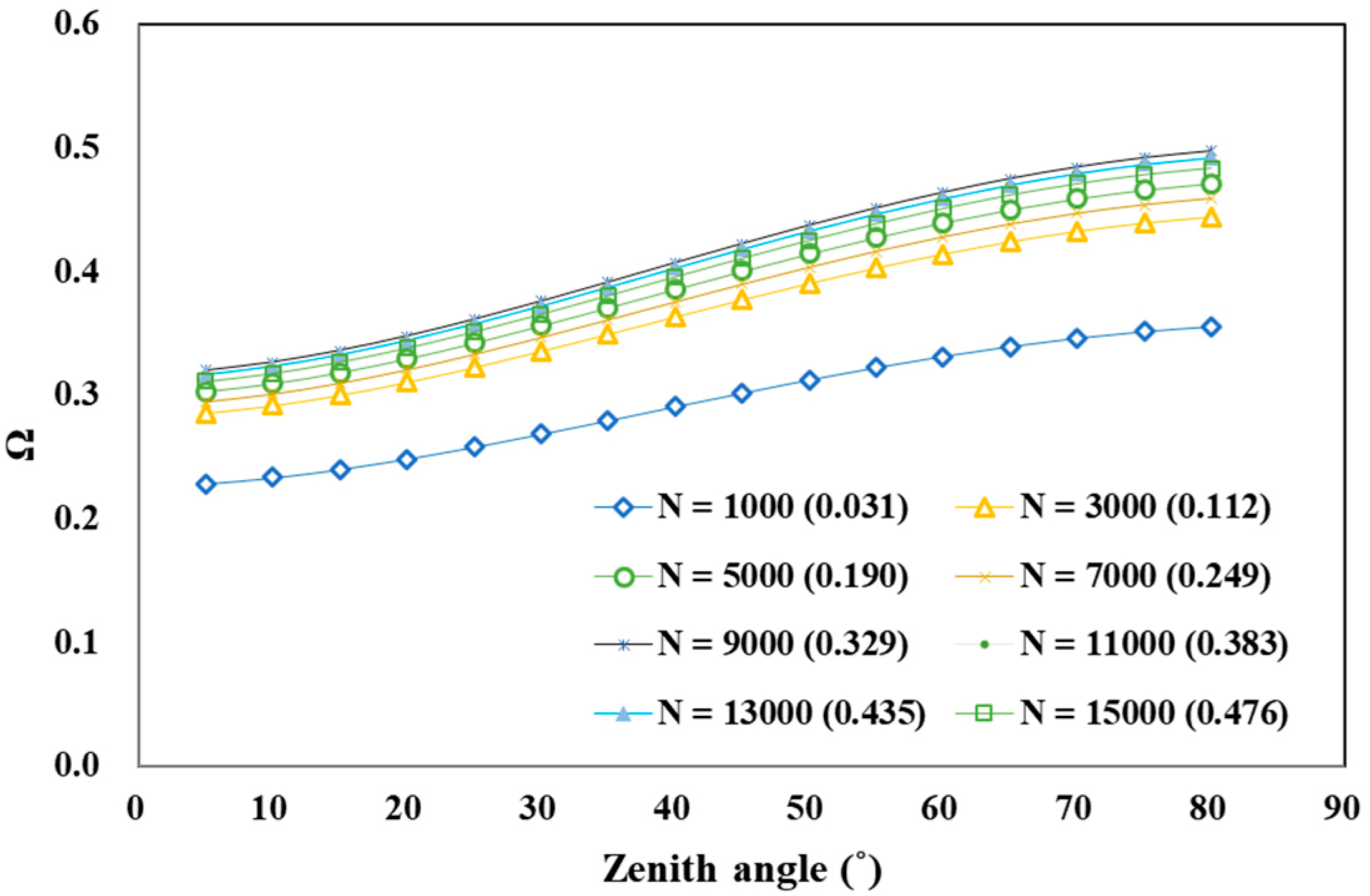
4.2. Factors Influencing Forest Clumping Index Variation
4.2.1. Tree Density
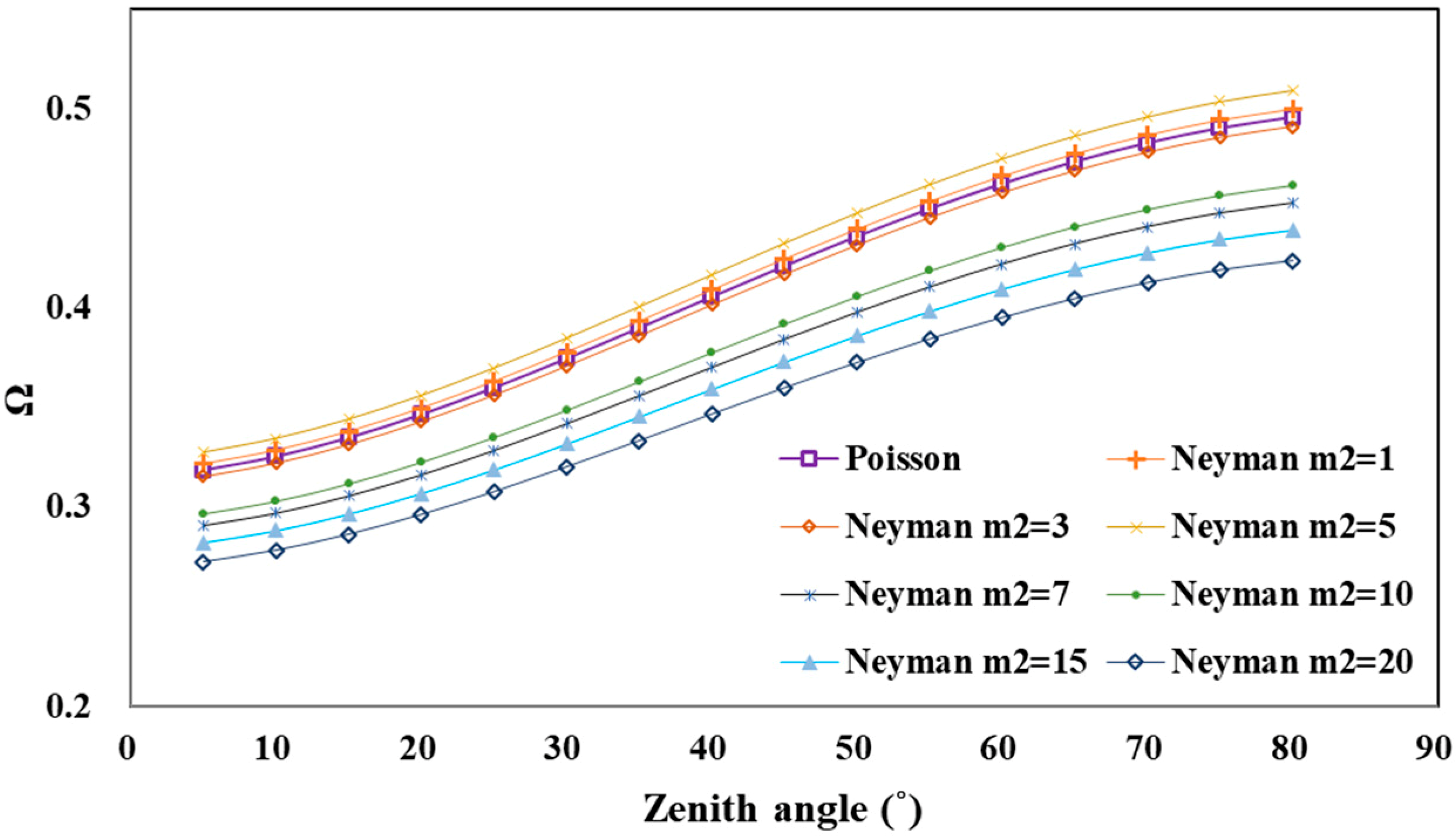
4.2.2. Tree Distribution Pattern
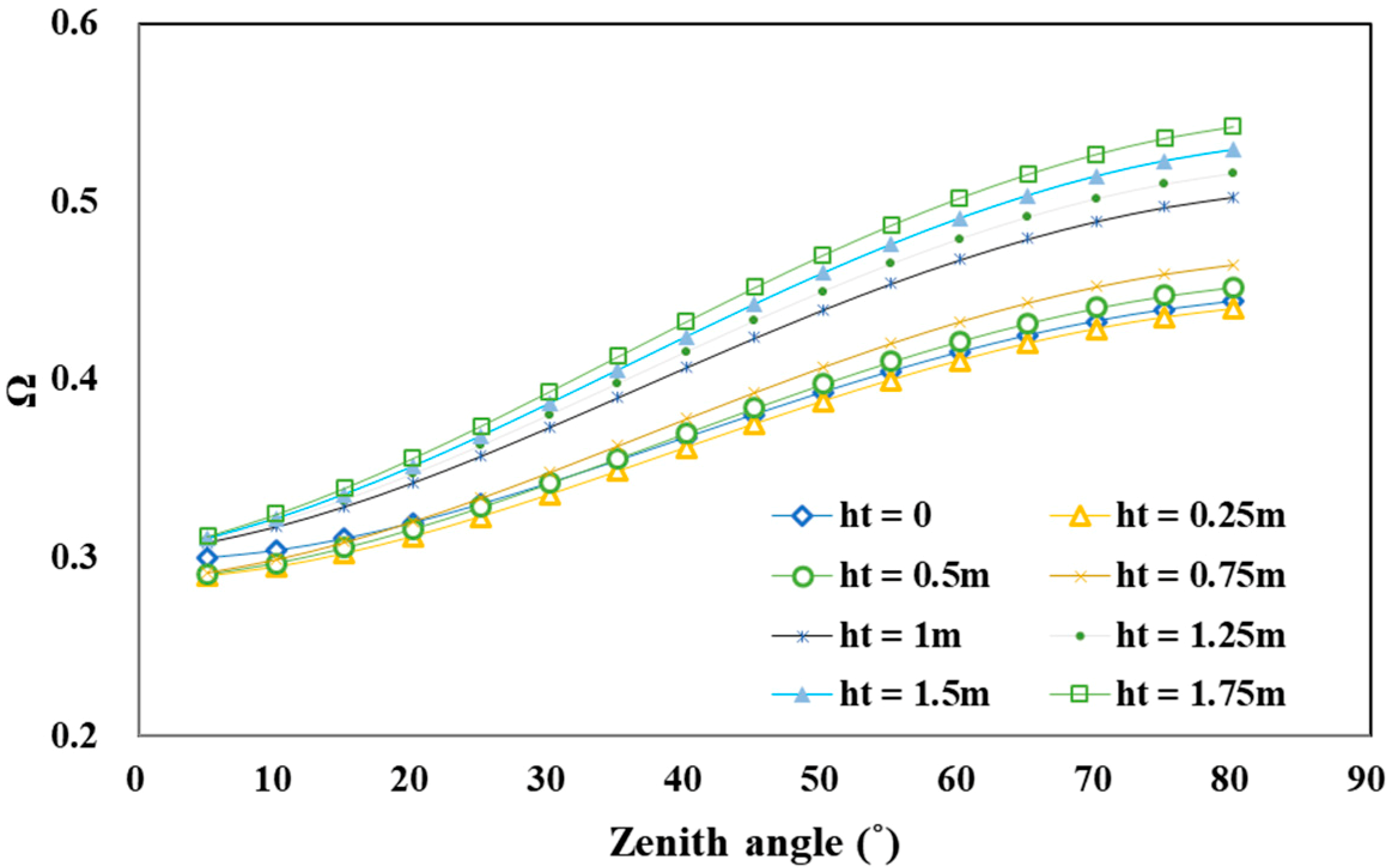
4.2.3. Trunk Height
4.2.4. Crown Shape
5. Conclusions
- (1)
- The crop model was developed for row structure with homogenous plant elements within the hedgerow. It is not suited for other structures or intercropping of multiple crops. The forest model has not considered the complementary tree species. The tree distribution pattern, LAI, and gap fraction will be quite different in a scenario with the mixture of complementary trees [53].
- (2)
- The influence of the crown shape model needs further investigation. Currently, we have assumed the tree crown as vertical ellipsoid. For most species, the crowns can quite well be represented by ellipsoids; however, some exceptions, such as the boreal needle forest, can be fit better by cones McPherson [54] stated that the mean difference between the crown volume measures from the assumptions of crown shape as paraboloids, vertical ellipsoids, and horizontal ellipsoids are 10%. In tropical forest, the multi-stem trees are very common. The current crown shape model might be not valid for them. The further effect of crown shape model on the clumping index needs to be discussed for different species.
- (3)
- Further comprehensive sensitivity analysis is expected. The analysis in this paper is restricted to be a mono parameter analysis and the variation range of all the parameters is independent. This is only the first step to observe the influence of each single parameter under specific conditions. In the natural environment, there is a size-density allometry of plants under self-thinning as the resources for growing are limited. The crown projection area scales to stem diameter [53]. Besides, LAI and LAD many vary simultaneously. Therefore, a multiple simultaneous analysis is needed to avoid the risk to misconstrue simple size effects as changes in the crown morphology.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, J.M.; Liu, J.; Leblanc, S.G.; Lacaze, R.; Roujean, J. Multi-angular optical remote sensing for assessing vegetation structure and carbon absorption. Remote Sens. Environ. 2003, 84, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Duthoit, S.; Demarez, V.; Gastellu-Etchegorry, J.; Martin, E.; Roujean, J. Assessing the effects of the clumping phenomenon on BRDF of a maize crop based on 3D numerical scenes using DART model. Agric. For. Meteorol. 2008, 148, 1341–1352. [Google Scholar] [CrossRef]
- Qin, Z.; Li, Z.; Cheng, F.; Chen, J.; Liang, B. Influence of canopy structural characteristics on cooling and humidifying effects of Populus tomentosa community on calm sunny summer days. Landsc. Urban Plan. 2014, 127, 75–82. [Google Scholar] [CrossRef]
- Chen, J.M.; Menges, C.H.; Leblanc, S.G. Global mapping of foliage clumping index using multi-angular satellite data. Remote Sens. Environ. 2005, 97, 447–457. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Fassnacht, K.S.; Gower, S.T.; Norman, J.M.; McMurtric, R.E. A comparison of optical and direct methods for estimating foliage surface area index in forests. Agric. For. Meteorol. 1994, 71, 183–207. [Google Scholar] [CrossRef]
- Nilson, T. Inversion of gap frequency data in forest stands. Agric. For. Meteorol. 1999, 98–99, 437–448. [Google Scholar] [CrossRef]
- Chen, J.M.; Leblanc, S.G. A four-scale bidirectional reflectance model based on canopy architecture. IEEE Trans. Geosci. Remote Sens. 1997, 35, 1316–1337. [Google Scholar] [CrossRef] [Green Version]
- Pisek, J.; Chen, J.M.; Lacaze, R.; Sonnentag, O.; Alikas, K. Expanding global mapping of the foliage clumping index with multi-angular POLDER three measurements: Evaluation and topographic compensation. ISPRS J. Photogramm. Remote Sens. 2010, 65, 341–346. [Google Scholar] [CrossRef]
- Miller, J.B. A formula for average foliage density. Aust. J. Bot. 1967, 15, 141–144. [Google Scholar] [CrossRef]
- Watson, D.J. Comparative physiological studies on the growth of field crops: I. Variation in net assimilation rate and leaf area between species and varieties, and within and between years. Ann. Bot. 1947, 11, 41–76. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A. Defining leaf area index for non-flat leaves. Plant Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Zheng, G.; Moskal, L.M. Retrieving leaf area index (LAI) using remote sensing: Theories, methods and sensors. Sensors 2009, 9, 2719–2745. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Baret, F.; Smith, G.J.; Jonckheere, I.; Coppin, P. Review of methods for in situ leaf area index (LAI) determination: Part II. Estimation of LAI, errors and sampling. Agric. For. Meteorol. 2004, 121, 37–53. [Google Scholar] [CrossRef]
- Bréda, N.J. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. J. Exp. Bot. 2003, 54, 2403–2417. [Google Scholar] [CrossRef] [PubMed]
- Kucharik, C.J.; Norman, J.M.; Murdock, L.M.; Gower, S.T. Characterizing canopy nonrandomness with a multiband vegetation imager (MVI). J. Geophys. Res. Atmos. (1984–2012) 1997, 102, 29455–29473. [Google Scholar] [CrossRef] [Green Version]
- Norman, J.M.; Jarvis, P.G. Photosynthesis in Sitka spruce (Picea sitchensis (Bong.) Carr.). III. Measurements of canopy structure and interception of radiation. J. Appl. Ecol. 1974, 11, 375–398. [Google Scholar] [CrossRef]
- Chen, J.M. Optically-based methods for measuring seasonal variation of leaf area index in boreal conifer stands. Agric. For. Meteorol. 1996, 80, 135–163. [Google Scholar] [CrossRef]
- Ryu, Y.; Nilson, T.; Kobayashi, H.; Sonnentag, O.; Law, B.E.; Baldocchi, D.D. On the correct estimation of effective leaf area index: Does it reveal information on clumping effects? Agric. For. Meteorol. 2010, 150, 463–472. [Google Scholar] [CrossRef]
- Sinoquet, H.; Sonohat, G.; Phattaralerphong, J.; Godin, C. Foliage randomness and light interception in 3-D digitized trees: An analysis from multiscale discretization of the canopy. Plant Cell Environ. 2005, 28, 1158–1170. [Google Scholar] [CrossRef]
- Kucharik, C.J.; Norman, J.M.; Gower, S.T. Characterization of radiation regimes in nonrandom forest canopies: Theory, measurements, and a simplified modeling approach. Tree Physiol. 1999, 19, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Kuusk, A. A fast, invertible canopy reflectance model. Remote Sens. Environ. 1995, 51, 342–350. [Google Scholar] [CrossRef]
- Leblanc, S.G.; Chen, J.M. A practical scheme for correcting multiple scattering effects on optical LAI measurements. Agric. For. Meteorol. 2001, 110, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Gonsamo, A.; Pellikka, P. The computation of foliage clumping index using hemispherical photography. Agric. For. Meteorol. 2009, 149, 1781–1787. [Google Scholar] [CrossRef]
- Wang, Y.; Woodcock, C.E.; Buermann, W.; Stenberg, P.; Voipio, P.; Smolander, H.; Häme, T.; Tian, Y.; Hu, J.; Knyazikhin, Y.; et al. Evaluation of the MODIS LAI algorithm at a coniferous forest site in Finland. Remote Sens. Environ. 2004, 91, 114–127. [Google Scholar] [CrossRef]
- He, L.; Chen, J.M.; Pisek, J.; Schaaf, C.B.; Strahler, A.H. Global clumping index map derived from the MODIS BRDF product. Remote Sens. Environ. 2012, 119, 118–130. [Google Scholar] [CrossRef]
- Pisek, J.; Ryu, Y.; Sprintsin, M.; He, L.; Oliphant, A.J.; Korhonen, L.; Kuusk, J.; Kuusk, A.; Bergstrom, R.; Verrelst, J.; et al. Retrieving vegetation clumping index from Multi-angle Imaging SpectroRadiometer (MISR) data at 275m resolution. Remote Sens. Environ. 2013, 138, 126–133. [Google Scholar] [CrossRef]
- Graefe, J.; Sandmann, M. Shortwave radiation transfer through a plant canopy covered by single and double layers of plastic. Agric. For. Meteorol. 2015, 201, 196–208. [Google Scholar] [CrossRef]
- Rautiainen, M.; Stenberg, P. On the angular dependency of canopy gap fractions in pine, spruce and birch stands. Agric. For. Meteorol. 2015, 206, 1–3. [Google Scholar] [CrossRef]
- Stenberg, P.; Manninen, T. The effect of clumping on canopy scattering and its directional properties: A model simulation using spectral invariants. Int. J. Remote Sens. 2015, 36, 5178–5191. [Google Scholar] [CrossRef]
- Garrigues, S.; Shabanov, N.V.; Swanson, K.; Morisette, J.T.; Baret, F.; Myneni, R.B. Intercomparison and sensitivity analysis of Leaf Area Index retrievals from LAI-2000, AccuPAR, and digital hemispherical photography over croplands. Agric. For. Meteorol. 2008, 148, 1193–1209. [Google Scholar] [CrossRef] [Green Version]
- Piayda, A.; Dubbert, M.; Werner, C.; Correia, A.V.; Pereira, J.S.; Cuntz, M. Influence of woody tissue and leaf clumping on vertically resolved leaf area index and angular gap probability estimates. For. Ecol. Manag. 2015, 340, 103–113. [Google Scholar] [CrossRef]
- Chen, J.M.; Mo, G.; Pisek, J.; Liu, J.; Deng, F.; Ishizawa, M.; Chan, D. Effects of foliage clumping on the estimation of global terrestrial gross primary productivity. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, S.G.; Fernandes, R.; Chen, J.M. Recent advancements in optical field leaf area index, foliage heterogeneity, and foliage angular distribution measurements. In Proceedings of the 2002 IEEE International Geoscience and Remote Sensing Symposium (IGARSS’02), Toronto, ON, Canada, 24–28 June 2002; pp. 2902–2904. [Google Scholar]
- Saitoh, T.M.; Nagai, S.; Noda, H.M.; Muraoka, H.; Nasahara, K.N. Examination of the extinction coefficient in the Beer-Lambert law for an accurate estimation of the forest canopy leaf area index. For. Sci. Technol. 2012, 8, 67–76. [Google Scholar] [CrossRef]
- Andrieu, B.; Sinoquet, H. Evaluation of structure description requirements for predicting gap fraction of vegetation canopies. Agric. For. Meteorol. 1993, 65, 207–227. [Google Scholar] [CrossRef]
- Fan, W.; Yan, B.; Xu, X. Crop area and leaf area index simultaneous retrieval based on spatial scaling transformation. Sci. China Earth Sci. 2010, 53, 1709–1716. [Google Scholar] [CrossRef]
- Peng, J.; Fan, W.; Xu, X.; Wang, L.; Liu, Q.; Li, J.; Zhao, P. Estimating Crop Albedo in the Application of a Physical Model Based on the Law of Energy Conservation and Spectral Invariants. Remote Sens. 2015, 7, 15536–15560. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Strahler, A.H. Geometric-optical bidirectional reflectance modeling of a conifer forest canopy. IEEE Trans. Geosci. Remote 1986, 6, 906–919. [Google Scholar] [CrossRef]
- Xu, X.; Fan, W.; Li, J.; Zhao, P.; Chen, G. A unified model of bidirectional reflectance distribution function for the vegetation canopy. Sci. China Earth Sci. 2017, 60, 463–477. [Google Scholar] [CrossRef]
- Yan, B.; Xu, X.; Fan, W. A unified canopy bidirectional reflectance (BRDF) model for row ceops. Sci. China Earth Sci. 2012, 55, 824–836. [Google Scholar] [CrossRef]
- Nilson, T. A theoretical analysis of the frequency of gaps in plant stands. Agric. Meteorol. 1971, 8, 25–38. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using Landsat TM images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Uria Diez, J.; Wiegand, K. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam. J. Veg. Sci. 2015, 27, 318–327. [Google Scholar] [CrossRef]
- Nilson, T.; Peterson, U. A forest canopy reflectance model and a test case. Remote Sens. Environ. 1991, 37, 131–142. [Google Scholar] [CrossRef]
- Franklin, J.; Michaelsen, J.; Strahler, A.H. Spatial analysis of density dependent pattern in coniferous forest stands. Vegetatio 1985, 64, 29–36. [Google Scholar] [CrossRef]
- Neyman, J. On a new class of “contagious” distributions, applicable in entomology and bacteriology. Ann. Math. Stat. 1939, 10, 35–57. [Google Scholar] [CrossRef]
- Li, J.; Fan, W.; Liu, Y.; Zhu, G.; Peng, J.; Xu, X. Estimating Savanna Clumping Index Using Hemispherical Photographs Integrated with High Resolution Remote Sensing Images. Remote Sens. 2016, 9, 52. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Plant canopy gap-size analysis theory for improving optical measurements of leaf-area index. Appl. Opt. 1995, 34, 6211–6222. [Google Scholar] [CrossRef] [PubMed]
- Kotchenova, S.Y.; Song, X.; Shabanov, N.V.; Potter, C.S.; Knyazikhin, Y.; Myneni, R.B. Lidar remote sensing for modeling gross primary production of deciduous forests. Remote Sens. Environ. 2004, 92, 158–172. [Google Scholar] [CrossRef]
- Ross, J.K. The Radiation Regime and the Architecture of Plant Stands; Junk W. In Pubs: The Hague, The Netherlands, 1981. [Google Scholar]
- Popescu, S.C.; Zhao, K. A voxel-based lidar method for estimating crown base height for deciduous and pine trees. Remote Sens. Environ. 2008, 112, 767–781. [Google Scholar] [CrossRef]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- McPherson, E.G.; Rowntree, R.A. Geometric solids for simulation of tree crowns. Landsc. Urban Plan. 1988, 15, 79–83. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Unit |
|---|---|---|
| Zenith angle, | degree | |
| Azimuth angle | degree | |
| Gap fraction | dimensionless | |
| Clumping index (Ω) | dimensionless | |
| LAI | Leaf area index | dimensionless |
| u | Leaf volume density | m−1 |
| Mean projection of unit foliage area along the direction of zenith angle | dimensionless | |
| Row period width, row width, gap width, and crop height, respectively. for any | m | |
| Row width and gap width, respectively, in the direction perpendicular to the row | m | |
| R | Radius of the tree crown | m |
| H | Height (or depth) of the tree crown | m |
| Trunk diameter at breast height, DBH | m | |
| Trunk height | m | |
| Crown closure, counting overlapped crown projection multiple times | dimensionless | |
| Crown closure, counts overlapped crown projection once | dimensionless | |
| Probability | dimensionless | |
| N | Average number of trees in unit area | dimensionless |
| M | Average number of trees in a quadrat defined by the Poisson distribution | dimensionless |
| m1 | Average group number of trees in a sample area defined by the Neyman distribution | dimensionless |
| m2 | Average number of trees in each group | dimensionless |
| Leaf reflectance | dimensionless | |
| Leaf transmittance | dimensionless | |
| Bi-directional reflectance of soil | dimensionless | |
| Directional-hemispherical reflectance of soil | dimensionless |
| Column ID | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Height of trunk (m) | 0.5 | 5 | 0.5 | 0.5 | 0, 0.25, …, 1.75 | 0.5 |
| Crown depth (m) | 6.5 | 1.5 | 2.28 | 2.28 | 2.28 | 2.07, 2.76, 3.62, 4.38, 5.09, 5.75, 6.37, 6.96 |
| Crown radius (m) | 0.45 | 0.75 | 0.76 | 0.76 | 0.76 | 0.80, 0.69, 0.60, 0.55, 0.51, 0.48, 0.45, 0.43 |
| H/2R ([-]) | 7.22 | 1 | 1.5 | 1.5 | 1.5 | 1.3, 2, 3, 4, 5, 6, 7, 8 |
| Radius at breast height (m) | 0.16 | 0.15 | 0.16 | 0.16 | 0.16 | 0.16 |
| ([-]) | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Average LAI ([-]) | 4.5 | 2 | 0.28, 0.84, 1.40, 1.97, 2.53, 3.09, 3.65, 4.22 | 2.25 | 2.25 | 2.25 |
| Area of research field (ha) | 1 | 5 | 4 | 4 | 4 | 4 |
| Distribution | Neyman m2 = 4 | Poisson | Neyman m2 = 4 | Poisson, Neyman m2 = 1, 3, 5, 7, 10, 15, 20 | Neyman m2 = 4 | Neyman m2 = 4 |
| Tree Number (per ha) | 4000 | 1011 | 250, 750, …, 3750 | 2000 | 2000 | 2000 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, J.; Fan, W.; Wang, L.; Xu, X.; Li, J.; Zhang, B.; Tian, D. Modeling the Directional Clumping Index of Crop and Forest. Remote Sens. 2018, 10, 1576. https://doi.org/10.3390/rs10101576
Peng J, Fan W, Wang L, Xu X, Li J, Zhang B, Tian D. Modeling the Directional Clumping Index of Crop and Forest. Remote Sensing. 2018; 10(10):1576. https://doi.org/10.3390/rs10101576
Chicago/Turabian StylePeng, Jingjing, Wenjie Fan, Lizhao Wang, Xiru Xu, Jvcai Li, Beitong Zhang, and Dingfang Tian. 2018. "Modeling the Directional Clumping Index of Crop and Forest" Remote Sensing 10, no. 10: 1576. https://doi.org/10.3390/rs10101576
APA StylePeng, J., Fan, W., Wang, L., Xu, X., Li, J., Zhang, B., & Tian, D. (2018). Modeling the Directional Clumping Index of Crop and Forest. Remote Sensing, 10(10), 1576. https://doi.org/10.3390/rs10101576