
Recent NDVI-Based Variation in Growth of Boreal Intact Forest Landscapes and Its Correlation with Climatic Variables


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
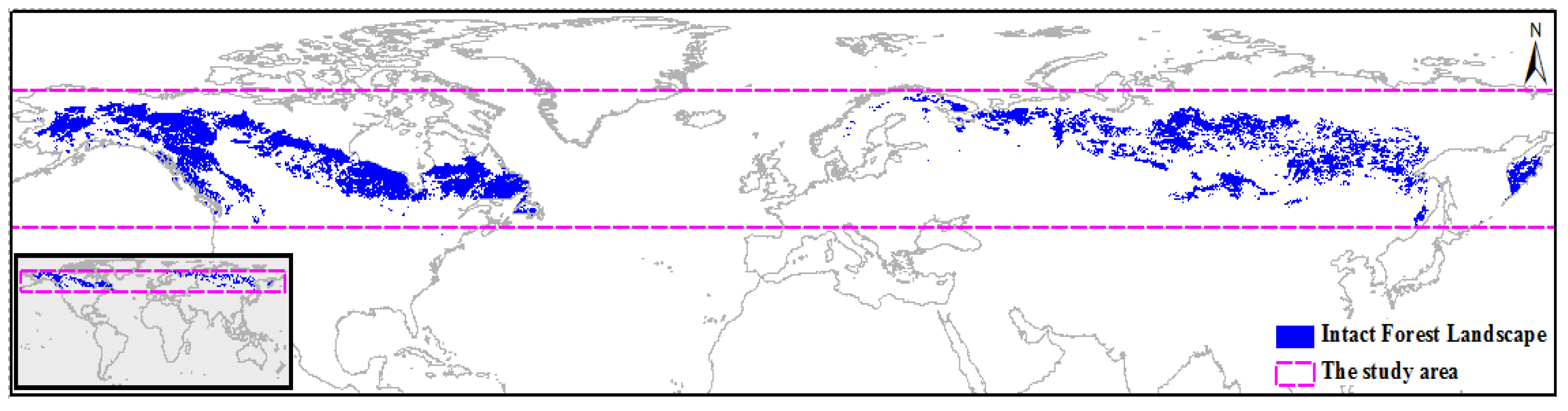
2.1. Study Area and Intact Forest Landscape
2.2. Climate Data
2.3. MODIS NDVI
2.4. Statistical Analyses Strategy
3. Results and Discussion
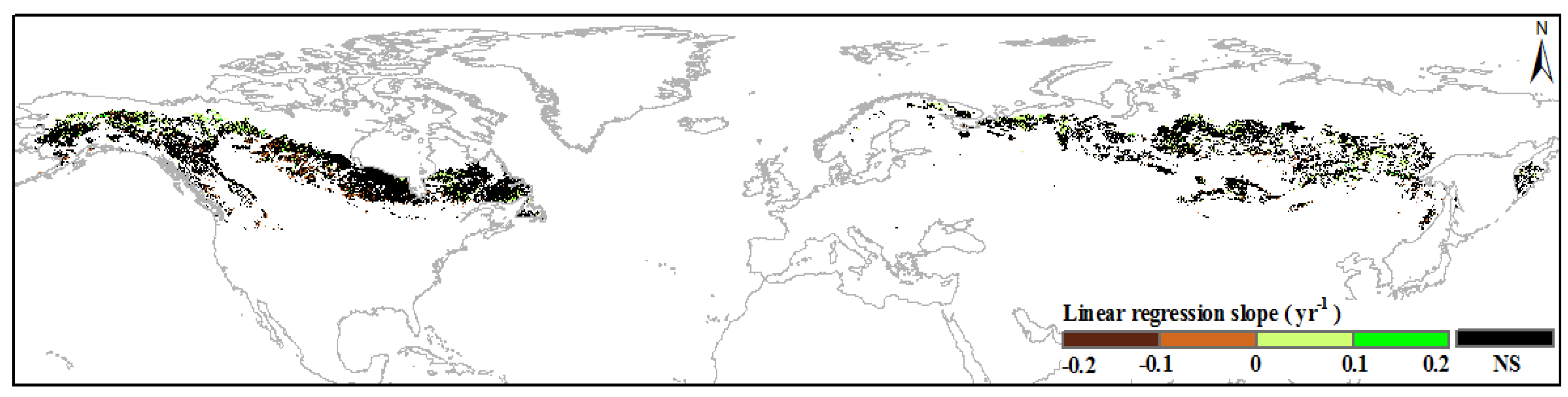
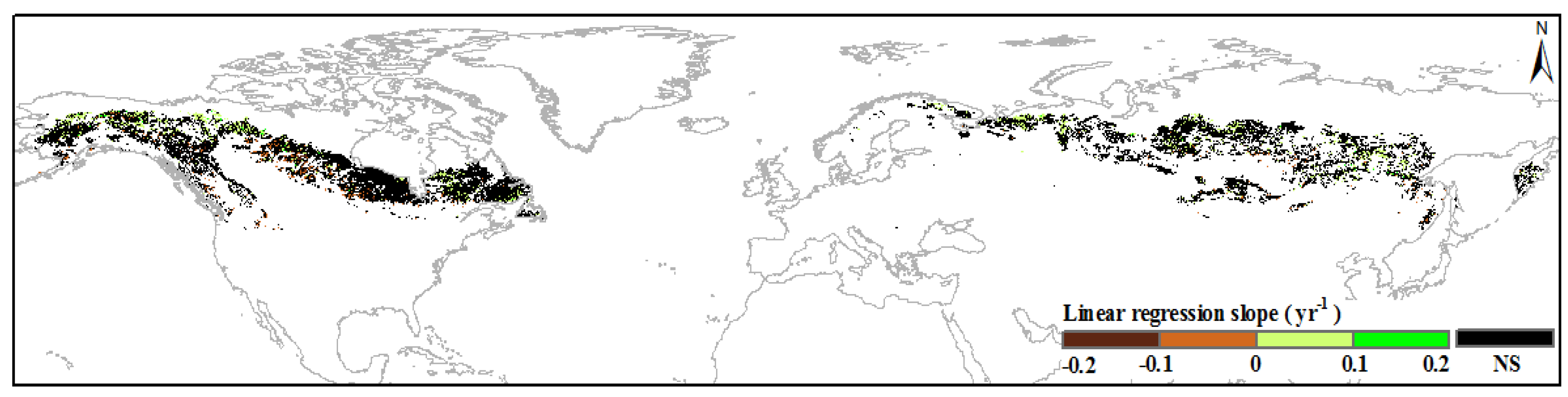
3.1. Annual Variations in Boreal IFLs
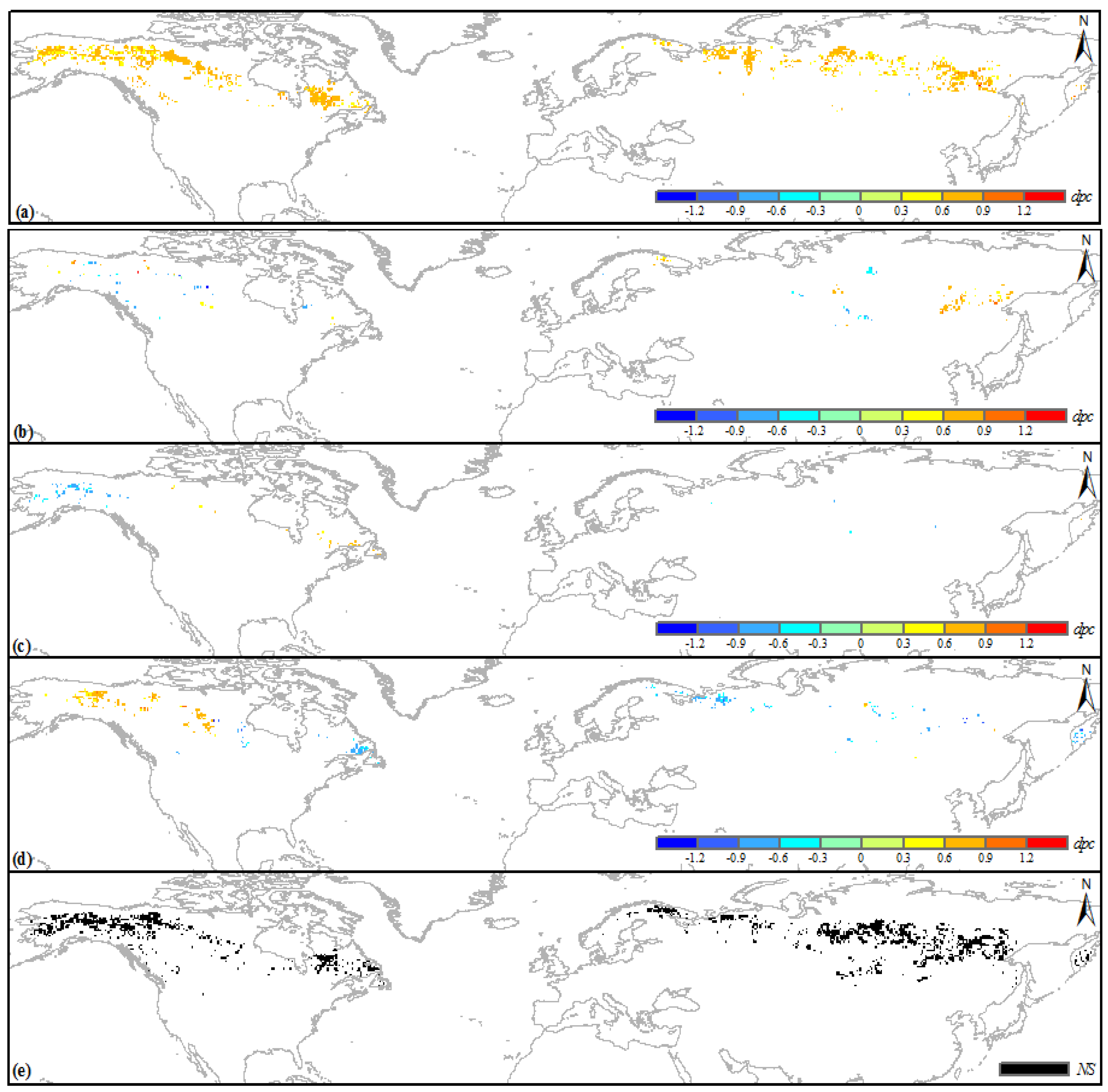
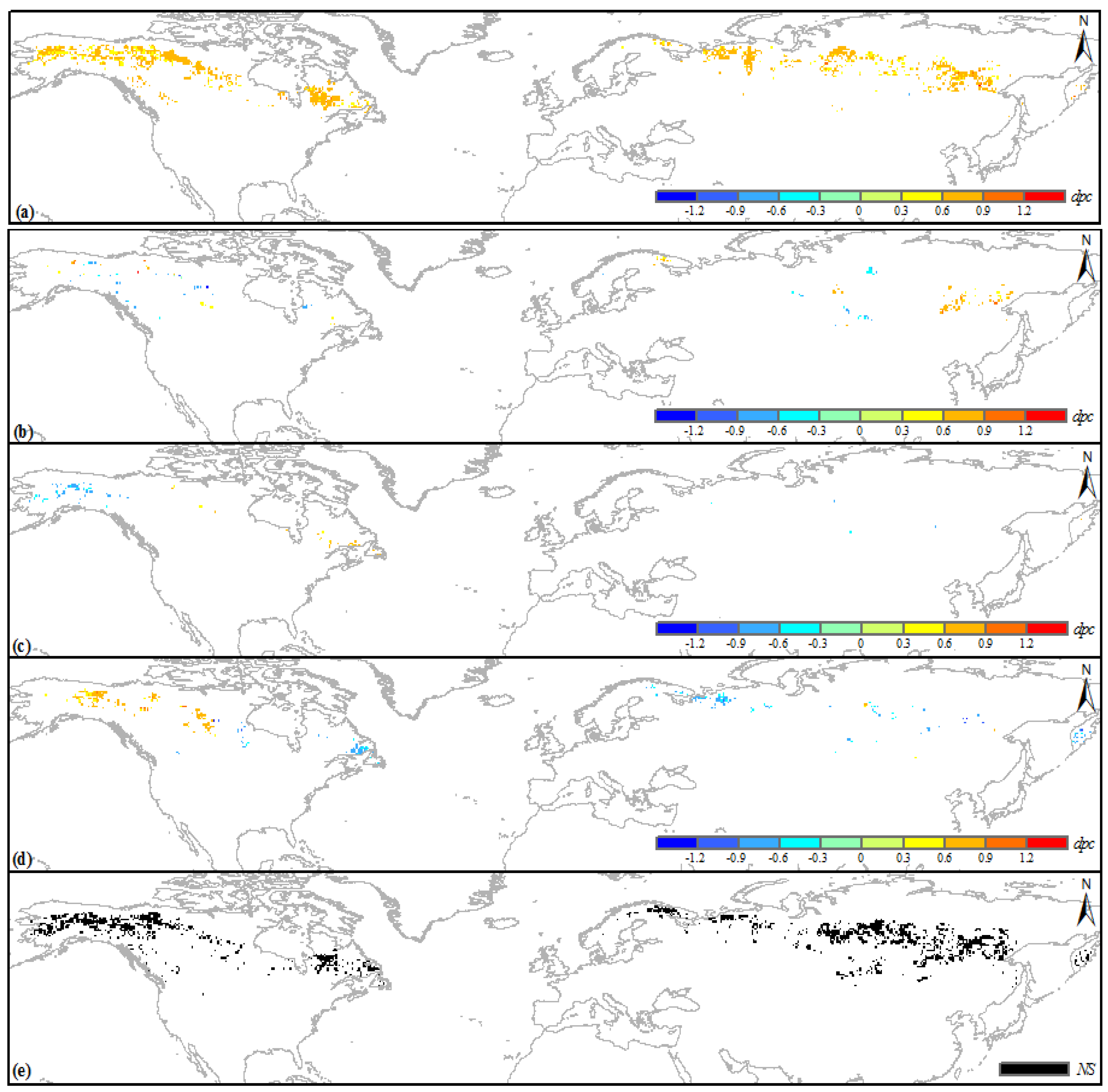
3.2. Climatic Sensitivity of the Greening Boreal IFLs
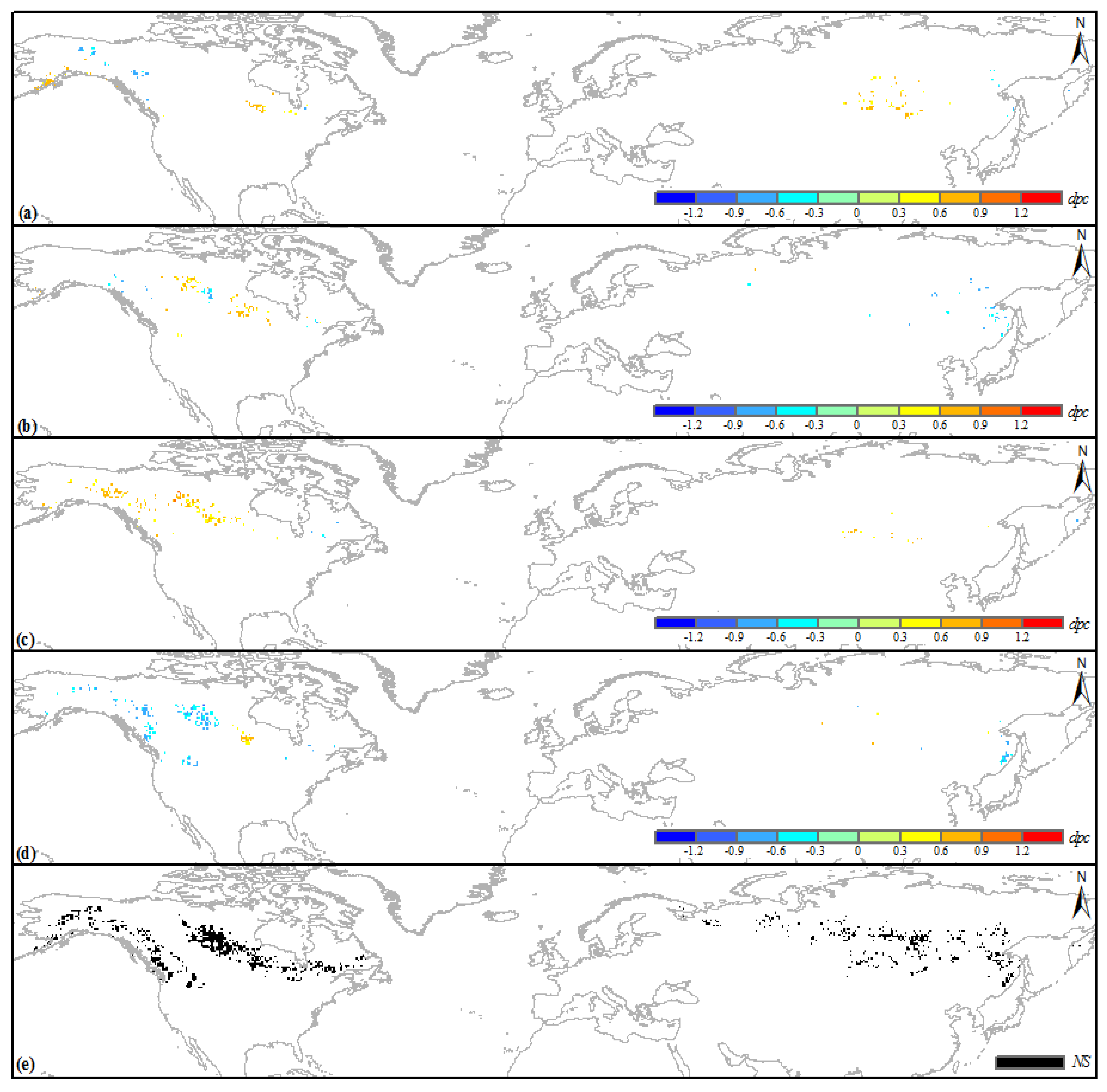
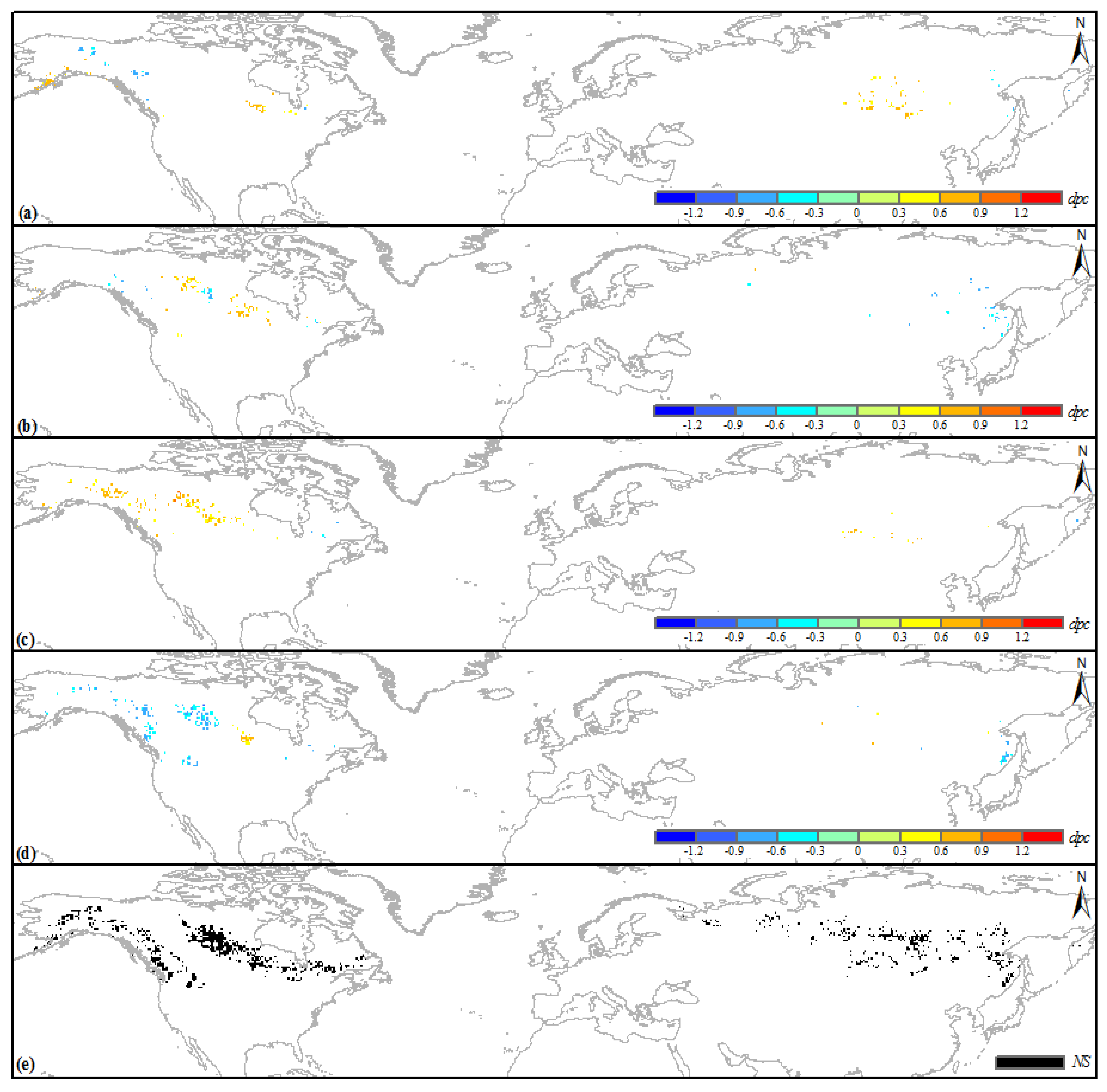
3.3. Climatic Sensitivity of the Browning Boreal IFLs
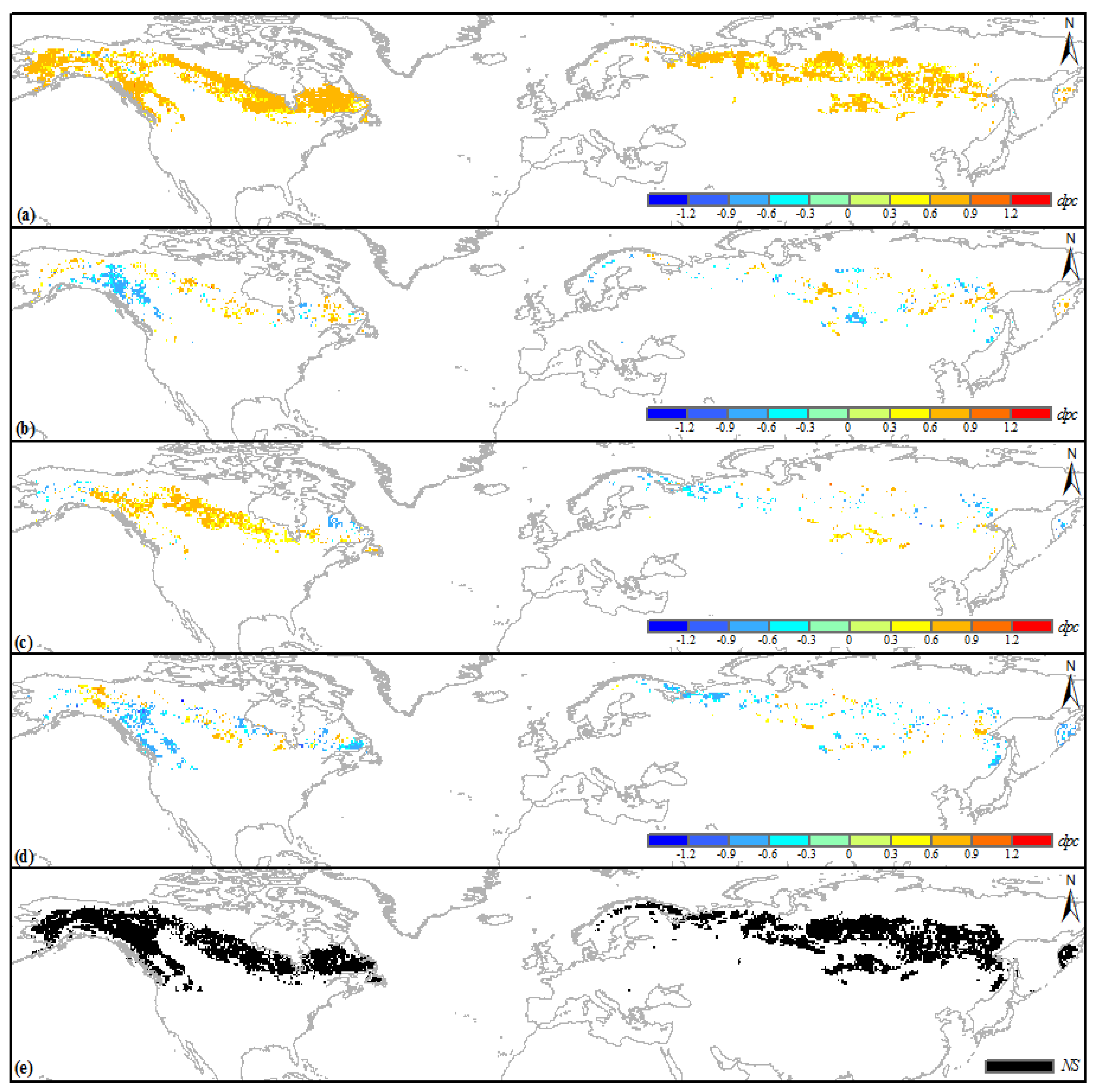
3.4. Climatic Sensitivity of the No-Change Boreal IFLs
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Zhang, L.; Sun, X.; Fu, Y.; Wen, X.; Wang, Q.; Li, S.; Ren, C.; Song, X.; Liu, Y.; et al. Environmental controls over carbon exchange of three forest ecosystems in eastern china. Glob. Chang. Biol. 2008, 14, 2555–2571. [Google Scholar] [CrossRef]
- Potapov, P.; Yaroshenko, A.; Turubanova, S.; Dubinin, M.; Laestadius, L.; Thies, C.; Aksenov, D.; Egorov, A.; Yesipova, Y.; Glushkov, I.; et al. Mapping the world’s intact forest landscapes by remote sensing. Ecol. Soc. 2008, 13, 51. [Google Scholar]
- Zhuravleva, I.; Turubanova, S.; Potapov, P.; Hansen, M.; Tyukavina, A.; Minnemeyer, S.; Laporte, N.; Goetz, S.; Verbelen, F.; Thies, C. Satellite-based primary forest degradation assessment in the democratic Republic of the Congo, 2000–2010. Environ. Res. Lett. 2013, 8, 279–288. [Google Scholar] [CrossRef]
- IFL Method-Intact Forest Landscapes. Available online: http://www.intactforests.org/method.html (accessed on 1 February 2016).
- Turner, I.M. Species loss in fragments of tropical rain forest: A review of the evidence. J. Appl. Ecol. 1996, 33, 200–209. [Google Scholar] [CrossRef]
- Yaroshenko, A.Y.; Potapov, P.V.; Turubanova, S.A. The Last Intact Forest Landscapes of Northern European Russia; Greenpeace Russia and Global Forest Watch: Moscow, Russia, 2001. [Google Scholar]
- Mollicone, D.; Achard, F.; Federici, S.; Eva, H.D.; Grassi, G.; Belward, A.; Raes, F.; Seufert, G.; Stibig, H.-J.; Matteucci, G.; et al. An incentive mechanism for reducing emissions from conversion of intact and non-intact forests. Clim. Chang. 2007, 83, 477–493. [Google Scholar] [CrossRef]
- Pptapov, P.; Laestadius, L.; Yaroshenko, A.; Turubanova, S. Global mapping and monitoring the extent of forest alteration: The intact forest landscapes method. Forest Resources Assessment Working Paper. 2009. Available online: http://www.fao.org/docrep/012/k7611e/k7611e00.pdf (accessed on 30 March 2016).
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Angertz, A.; Biraud, S.; Bonfils, C.; Henning, C.C.; Buermann, W.; Pinzon, J.; Tucker, C.J.; Fung, I. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proc. Natl. Acad. Sci. USA 2005, 102, 10823–10827. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Wang, X.; Ciais, P.; ZHU, B.; Wang, T.; Liu, J. Changes in satellite-derived vegetation growth trend in temperate and boreal eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Jin, J.; Jiang, H.; Peng, C.; Zhang, X.; Wang, Y.; Wang, J. Climate change contribution to forest growth in eastern china over past two decades. Terr. Atmos. Ocean. Sci. 2014, 25, 49–60. [Google Scholar] [CrossRef]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Cadule, P.; Viovy, N.; Wang, T. Spatiotemporal patterns of terrestrial carbon cycle during the 20th century. Glob. Biogeochem. Cycles 2009, 23, 2191–2196. [Google Scholar] [CrossRef]
- Tan, K.; Piao, S.; Peng, C.; Fang, J. Satellite-based estimation of biomass carbon stocks for northeast china’s forests between 1982 and 1999. For. Ecol. Manag. 2007, 240, 114–121. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S.; et al. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Chang. Boil. 2015, 21, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.; Pinzon, J.; Brown, M.; Slayback, D.; Pak, E.; Mahoney, R.; Vermote, E.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- De Jong, R.; Verbesselt, J.; Zeileis, A.; Schaepman, M.E. Shifts in global vegetation activity trends. Remote Sens. 2013, 5, 1117–1133. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5. Article 5018. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Piao, S.; Ciais, P.; Li, J.; Friedlingstein, P.; Koven, C.; Chen, A. Spring temperature change and its implication in the change of vegetation growth in north america from 1982 to 2006. Proc. Natl. Acad. Sci. USA 2011, 108, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Sohn, B.J. Recent trends in changes of vegetation over East Asia coupled with temperature and rainfall variations. J. Geophys. Res. 2010, 115, 1307–1314. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Chen, A.; Xu, L.; Cao, C.; Fang, J.; Myneni, R.B.; Pinzon, J.E.; Tucker, C.J.; Piao, S. Recent change of vegetation growth trend in China. Environ. Res. Lett. 2011, 6, 44027–44039. [Google Scholar] [CrossRef]
- Bryant, D.; Nielsen, D.; Tangley, L.; Sizer, N.; Miranda, M.; Brown, P.; Johnson, N.; Malk, A.; Miller, K. The last frontier forests: Ecosystems and economies on the edge. What is the status of the worlds remaining large natural forest ecosystems? 1997. Available online: http://www.popline.org/node/526187 (accessed on 20 April 2015).
- Intact Forest Landscapes. Available online: http://www.intactforests.org/data.ifl.html (accessed on 10 October 2015).
- Centre for Environmental Data Analysis. Available online: http://browse.ceda.ac.uk/browse/badc/cru/data/cru_ts/cru_ts_3.23/data (accessed on 30 June 2015).
- New, M.; Hulme, M.; Jones, P. Representing twentieth-century space-time climate variability. Part ii: Development of 1901–96 monthly grids of terrestrial surface climate. J. Clim. 2000, 13, 2217–2238. [Google Scholar] [CrossRef]
- Land Processes Distributed Active Archive Center. Available online: https://lpdaac.usgs.gov/dataset_discovery/modis/modis_products_table/mod13c2 (accessed on 28 April 2015).
- Farrer, E.C.; Ashton, I.W.; Spasojevic, M.J.; Fu, S.Y.; Gonzalez, D.J.X.; Suding, K.N. Indirect effects of global change accumulate to alter plant diversity but not ecosystem function in alpine tundra. J. Ecol. 2015, 103, 351–360. [Google Scholar] [CrossRef]
- Chen, C.; Chen, D.L.; Pan, J.J.; Lam, S.K. Analysis of factors controlling soil N2O emission by principal component and path analysis method. Environ. Earth Sci. 2014, 72, 1511–1517. [Google Scholar] [CrossRef]
- Brown, R.D.; Robinson, D.A. Northern hemisphere spring snow cover variability and change over 1922–2010 including an assessment of uncertainty. Cryosphere Discuss. 2010, 5, 219–229. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Friedlingstein, P.; Zhou, L.; Wang, T. Change in snow phenology and its potential feedback to temperature in the northern hemisphere over the last three decades. Environ. Res. Lett. 2013, 8, 1880–1885. [Google Scholar] [CrossRef]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.H.; Ciais, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z.; Jeong, S.-J.; et al. Leaf onset in the northern hemisphere triggered by daytime temperature. Nat. Commun. 2015. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Wang, Y.; Jiang, H.; Cheng, M. Recent NDVI-Based Variation in Growth of Boreal Intact Forest Landscapes and Its Correlation with Climatic Variables. Sustainability 2016, 8, 326. https://doi.org/10.3390/su8040326
Jin J, Wang Y, Jiang H, Cheng M. Recent NDVI-Based Variation in Growth of Boreal Intact Forest Landscapes and Its Correlation with Climatic Variables. Sustainability. 2016; 8(4):326. https://doi.org/10.3390/su8040326
Chicago/Turabian StyleJin, Jiaxin, Ying Wang, Hong Jiang, and Min Cheng. 2016. "Recent NDVI-Based Variation in Growth of Boreal Intact Forest Landscapes and Its Correlation with Climatic Variables" Sustainability 8, no. 4: 326. https://doi.org/10.3390/su8040326
APA StyleJin, J., Wang, Y., Jiang, H., & Cheng, M. (2016). Recent NDVI-Based Variation in Growth of Boreal Intact Forest Landscapes and Its Correlation with Climatic Variables. Sustainability, 8(4), 326. https://doi.org/10.3390/su8040326