
Resilience Assessment of Lowland Plantations Using an Ecosystem Modeling Approach




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Background
1.2. Restoring the Resilience of Reforested Areas
2. Experimental Section
2.1. Study Species
2.2. The FORECAST Model
2.3. Mixed Plantations Scenarios
2.4. Measuring Facilitation or Competition in Mixed Plantations
3. Results and Discussion
3.1. Model Performance
3.2. Interspecific Competition and Facilitation Processes


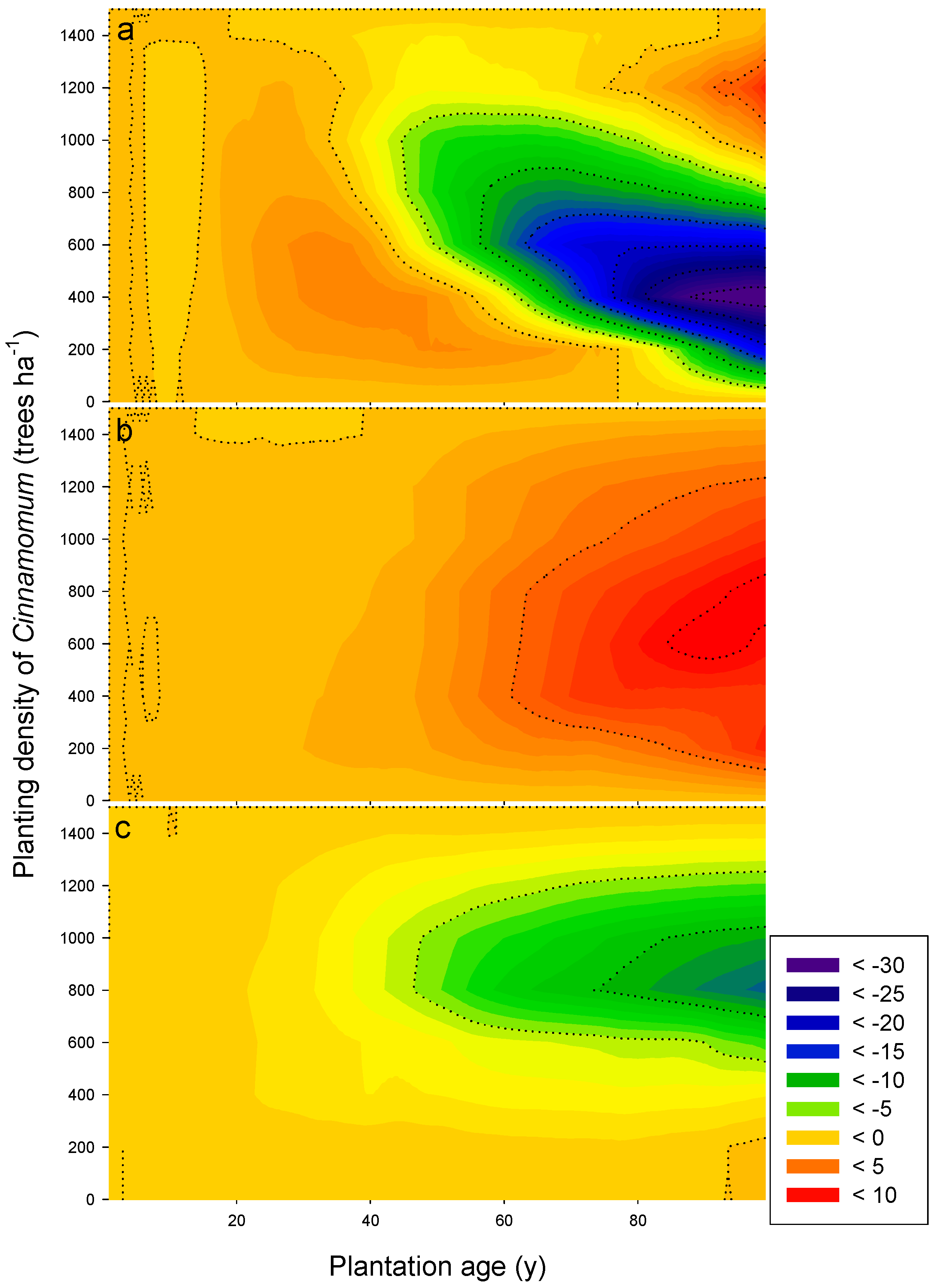
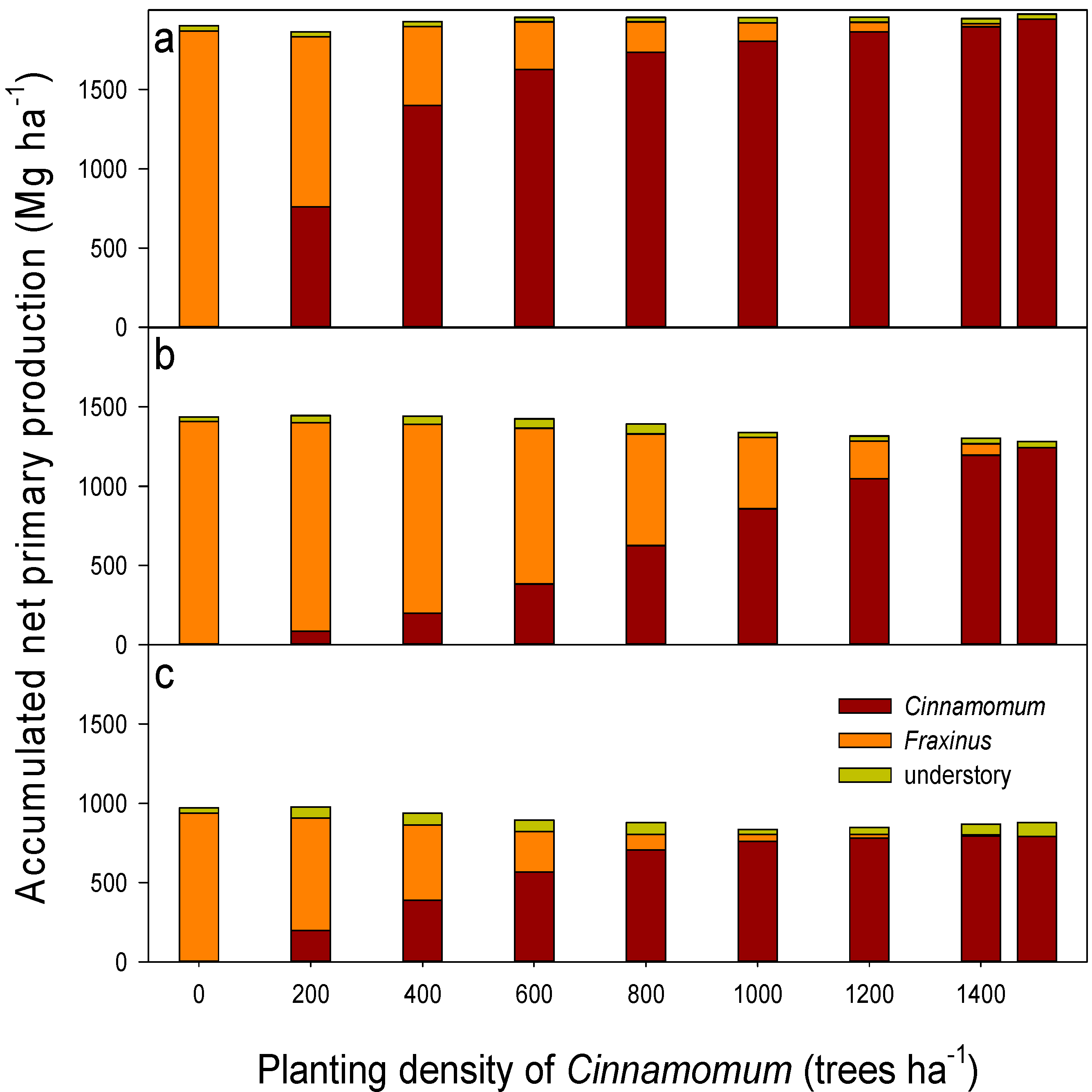
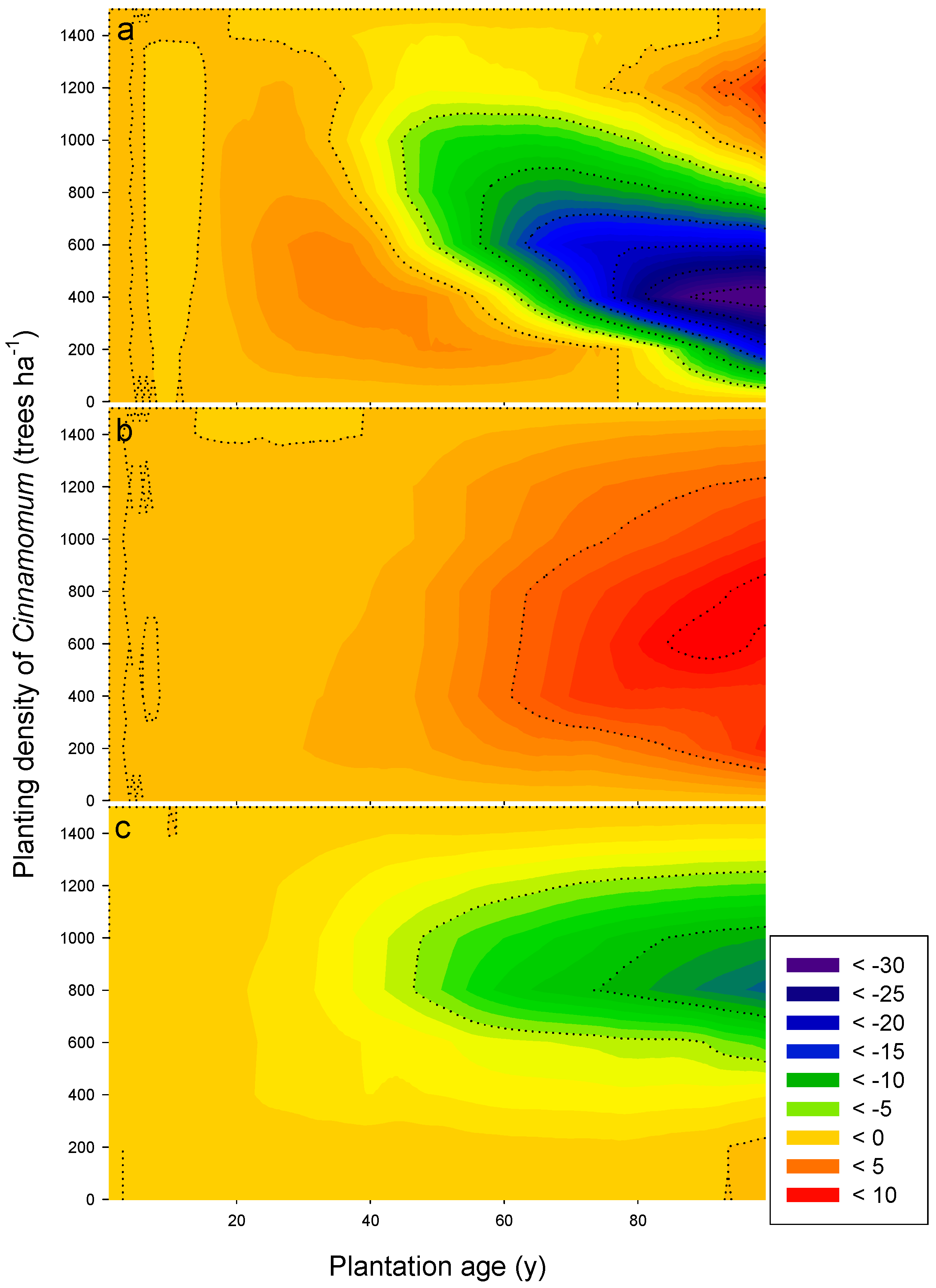
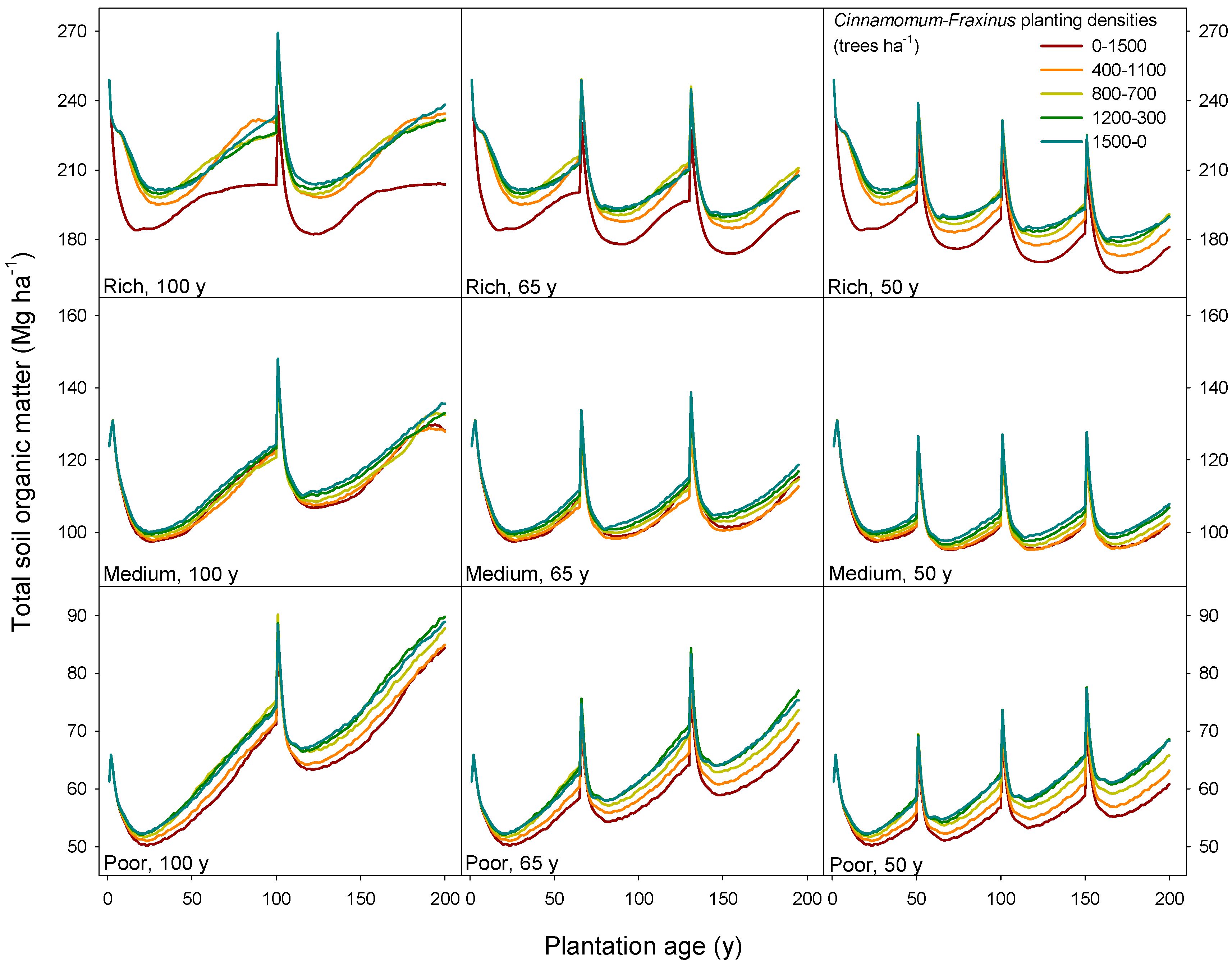
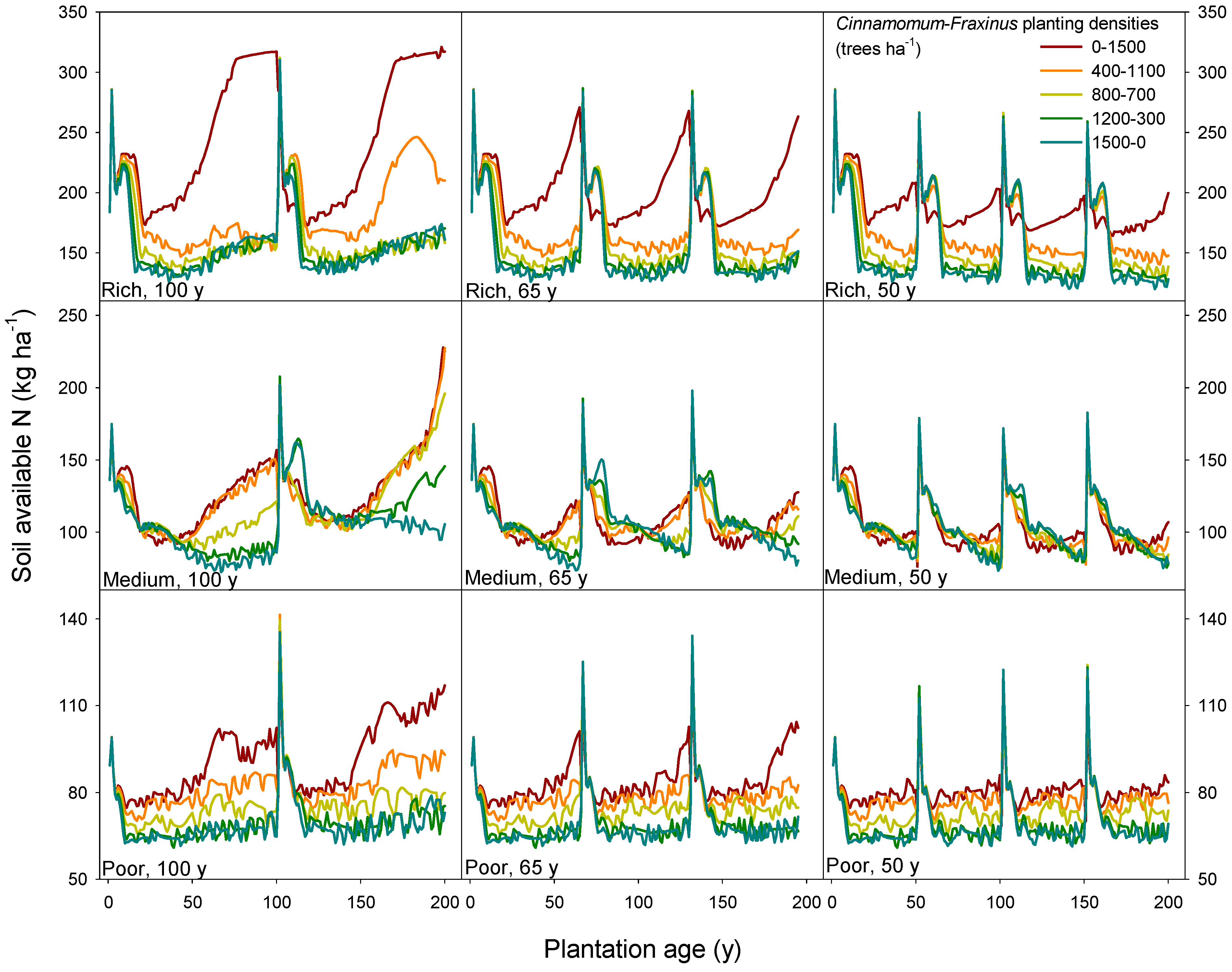
3.3. Long-Term Sustainability of the Productivity in Mixed Camphor Laurel-Himalayan Ash Plantations
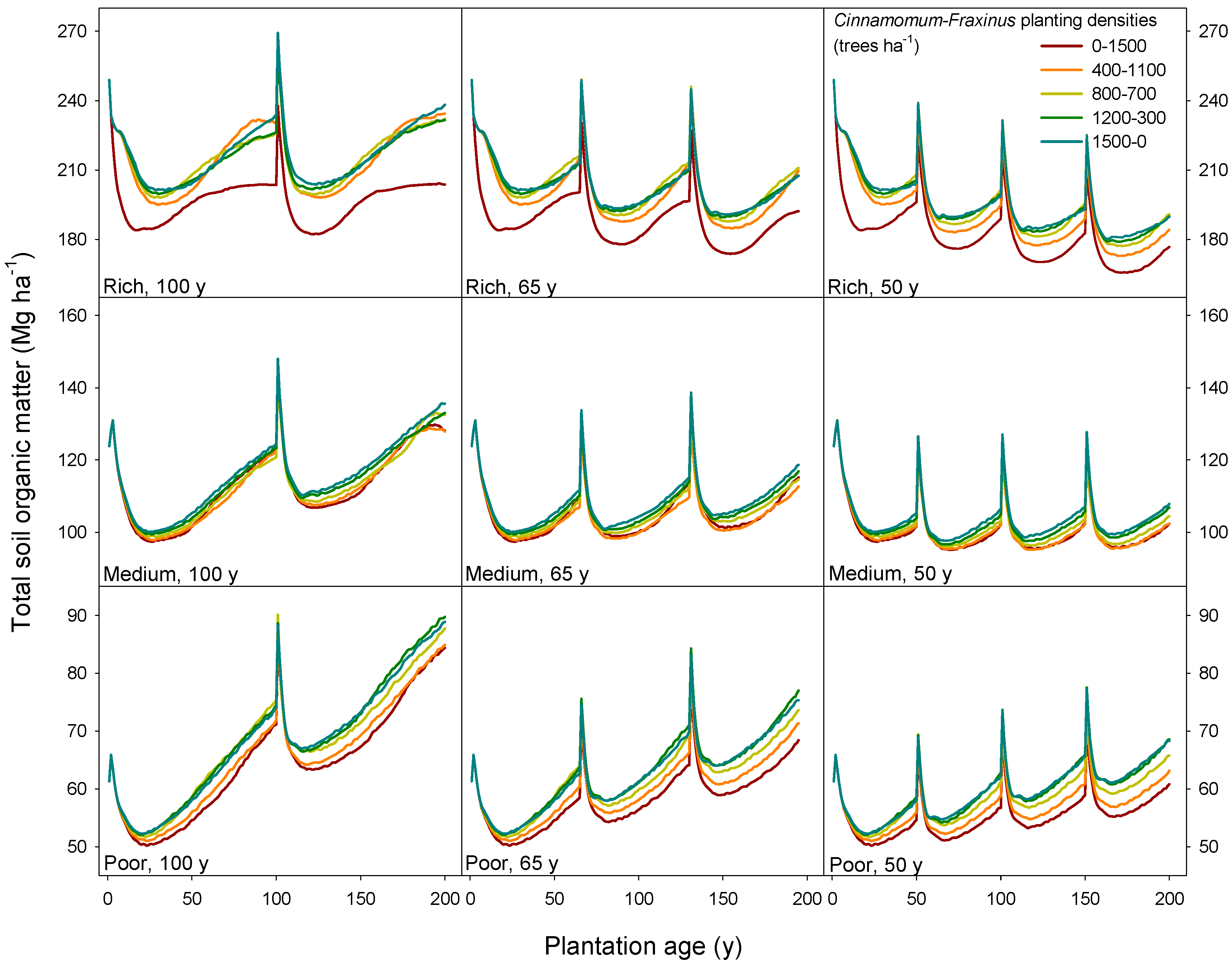
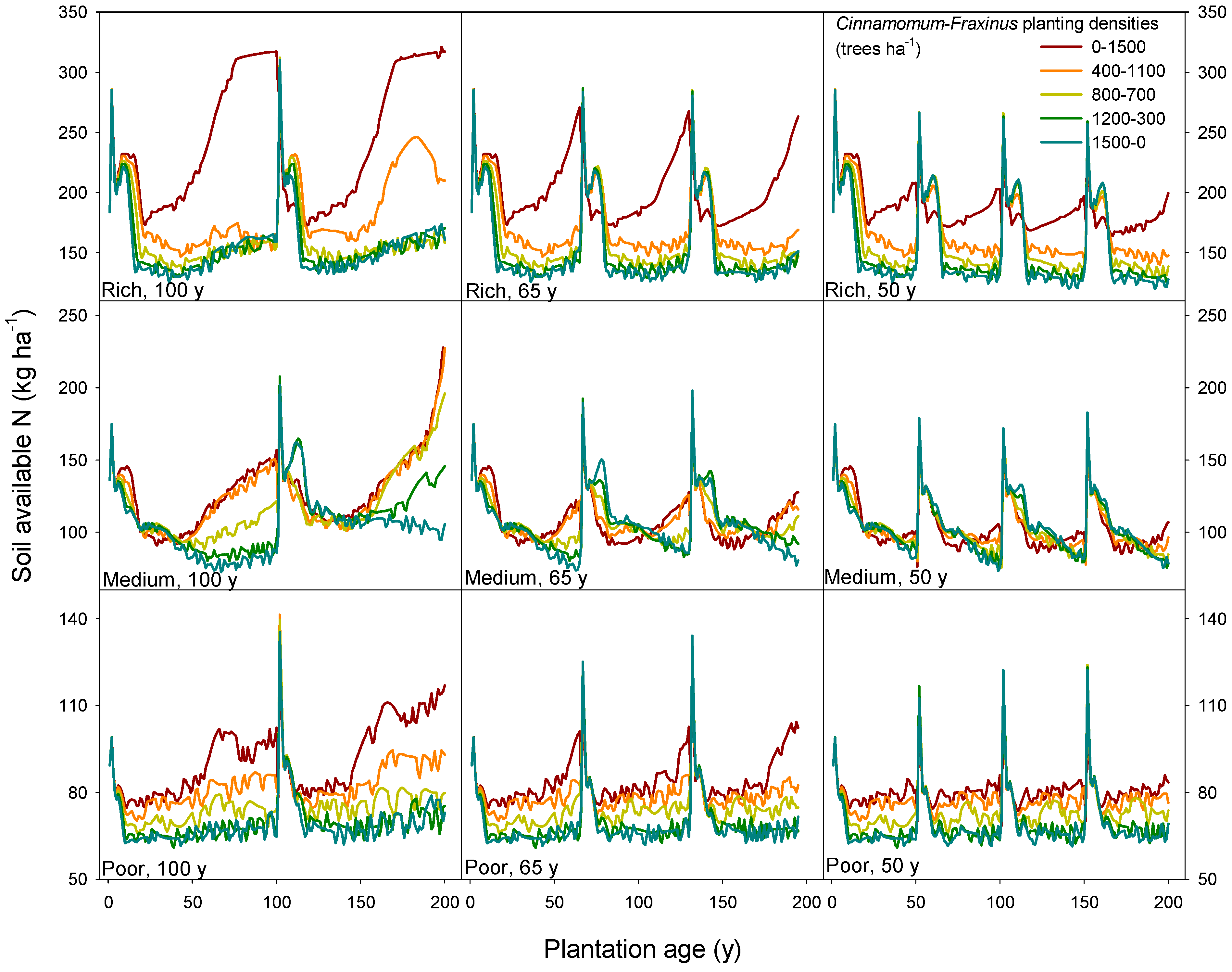
3.4. Model Limitations and Further Work
4. Conclusions and Management Implications
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rudel, T.K.; Perez-Lugo, M.; Zichal, H. When fields revert to forest: Development and spontaneous reforestation in post-war Puerto Rico. Prof. Geogr. 2000, 52, 386–397. [Google Scholar] [CrossRef]
- Lambin, E.F.; Meyfroidt, P. Global land use change, economic globalization, and the looming land scarcity. Proc. Natl. Acad. Sci. USA 2011, 108, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.B.; Turner, B.L. Human population growth and global land-use/cover change. Annu. Rev. Ecol. Syst. 1992, 23, 39–61. [Google Scholar] [CrossRef]
- FOREST EUROPE; UNECE; FAO. State of Europe’s Forests 2011. Status and Trends in Sustainable Forest Management in Europe; Ministerial Conference on the Protection of Forests in Europe: Aas, Norway, 2011; p. 337. [Google Scholar]
- FAO. State of the World’s Forests 2014; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; p. 119. [Google Scholar]
- Alston, M. Who is down on the farm? Social aspects of Australian agriculture in the 21st century. Agr. Hum. Values 2004, 21, 37–46. [Google Scholar] [CrossRef]
- Lin, K.-C.; Liu, W.-Y. An empirical study of afforestation policy in Taiwan. Socioecon. Law Inst. Rev. 2007, 40, 175–211. (In Chinese) [Google Scholar]
- Lo, K.-A. Agricultural landowners’ participation in the plain landscape afforestation program (PLAP): A case study in Taitung County, Taiwan. Q. J. For. Res. 2005, 27, 17–30. (In Chinese) [Google Scholar]
- Kimmins, J.P.; Blanco, J.A.; Seely, B.; Welham, C.; Scoullar, K. Forecasting Forest Futures: A Hybrid Modelling Approach to the Assessment of Sustainability of Forest Ecosystems and Their Values; Earthscan: London, UK, 2010; p. 281. [Google Scholar]
- Messier, C.; Puettmann, K.J.; Coates, K.D. Managing forests as complex adaptive systems: Building resilience to the challenge of global change. In The Earthscan Forest Library; Routledge: London, UK, 2013; p. 368. [Google Scholar]
- Herbert, D.A.; Fownes, J.H.; Vitousek, P.M. Hurricane damage to a Hawaiian forest: Nutrient supply rate affects resistance and resilience. Ecology 1999, 80, 908–920. [Google Scholar] [CrossRef]
- Chapin, F.S.; McGuire, A.D.; Ruess, R.W.; Hollingsworth, T.N.; Mack, M.C.; Johnstone, J.F.; Kasischke, E.S.; Euskirchen, E.S.; Jones, J.B.; Jorgenson, M.T.; et al. Resilience of Alaska’s boreal forest to climatic change. Can. J. For. Res. 2010, 40, 1360–1370. [Google Scholar] [CrossRef]
- Blanco, J.A.; Dubois, D.; Littlejohn, D.; Flanders, D.N.; Robinson, P.; Moshofsky, M.; Welham, C. Soil organic matter: A sustainability indicator for wildfire control and bioenergy production in the urban/forest interface. Soil Sci. Soc. Am. J. 2014. [Google Scholar] [CrossRef]
- Dymond, C.C.; Tedder, S.; Spittlehouse, D.L.; Raymer, B.; Hopkins, K.; McCallion, K.; Sandland, J. Diversifying managed forests to increase resilience. Can. J. For. Res. 2014, 44, 1196–1205. [Google Scholar] [CrossRef]
- Kelty, M.J. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Springer: Berlin, Germany, 1992; pp. 125–141. [Google Scholar]
- Chomel, M.; DesRochers, A.; Baldy, V.; Larchevêque, M.; Gauquelin, T. Non-additive effects of mixing hybrid poplar and white spruce on aboveground and soil carbon storage in boreal plantations. For. Ecol. Manag. 2014, 328, 292–299. [Google Scholar] [CrossRef]
- Pretzsch, H. Forest Dynamics, Growth, and Yield: From Measurement to Model; Springer: Berlin, Germany, 2009; p. 664. [Google Scholar]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Lo, Y.-H.; Blanco, J.A.; Kimmins, J.P.; Seely, B.; Welham, C. Linking climate change and forest ecophysiology to project future trends in tree growth: A review of forest models. In Climate Change—Research and Technology for Adaptation and Mitigation; Blanco, J.A., Kheradmand, H., Eds.; InTech: Rijeka, Croatia, 2011; pp. 63–86. [Google Scholar]
- Kimmins, J.P.; Mailly, D.; Seely, B. Modelling forest ecosystem net primary production: The hybrid simulation approach used in FORECAST. Ecol. Model 1999, 122, 195–224. [Google Scholar] [CrossRef]
- Sachs, D.; Sollins, P. Potential effects of management practices on nitrogen nutrition and long-term productivity of western hemlock stands. For. Ecol. Manag. 1986, 17, 25–36. [Google Scholar] [CrossRef]
- Wang, J.R.; Comeau, P.; Kimmins, J.P. Simulation of mixedwood management of aspen and white spruce in northeastern British Columbia. Water Air Soil Pollut. 1995, 82, 171–178. [Google Scholar] [CrossRef]
- Morris, D.M.; Kimmins, J.P.; Duckert, D.R. The use of soil organic matter as a criterion of the relative sustainability of forest management alternatives: A modelling approach using FORECAST. For. Ecol. Manag. 1997, 94, 61–78. [Google Scholar] [CrossRef]
- Wei, X.; Liu, W.; Waterhouse, J.; Armleder, M. Simulations on impacts of different management strategies on long-term site productivity in lodgepole pine forests of the central interior of British Columbia. For. Ecol. Manag. 2000, 133, 217–229. [Google Scholar] [CrossRef]
- Wei, X.; Kimmins, J.P.; Zhou, G. Disturbances and the sustainability of long-term site productivity in lodgepole pine forests in the central interior of British Columbia—An ecosystem modeling approach. Ecol. Model. 2003, 164, 239–256. [Google Scholar] [CrossRef]
- Blanco, J.A.; Seely, B.; Welham, C.; Kimmins, J.P.; Seebacher, T.M. Testing the performance of a forest ecosystem model (FORECAST) against 29 years of field data in a Pseudotsuga menziesii plantation. Can. J. For. Res. 2007, 37, 1808–1820. [Google Scholar] [CrossRef]
- Seely, B.; Welham, C.; Blanco, J.A. Towards the application of soil organic matter as an indicator of forest ecosystem productivity: Deriving thresholds, developing monitoring systems, and evaluating practices. Ecol. Indic. 2010, 10, 999–1008. [Google Scholar] [CrossRef]
- Bi, J.; Blanco, J.A.; Seely, B.; Kimmins, J.P.; Ding, Y.; Welham, C. Yield decline in Chinese-fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 37, 1615–1630. [Google Scholar] [CrossRef]
- Blanco, J.A.; González, E. Exploring the sustainability of current management prescriptions for Pinus caribaea plantations in Cuba: A modelling approach. J. Trop. For. Sci. 2010, 22, 139–154. [Google Scholar]
- Blanco, J.A.; González, E. The legacy of forest management in tropical forests: Analysis of its long-term influence with ecosystem models. For. Syst. 2010, 19, 249–262. [Google Scholar]
- Xin, Z.H.; Jiang, H.; Jie, C.Y.; Wei, X.H.; Blanco, J.; Zhou, G.M. Simulated nitrogen dynamics for a Cunninghamia lanceolata plantation with selected rotation ages. J. Zhejiang Agric. For. Univ. 2011, 28, 855–862. [Google Scholar]
- Blanco, J.A.; Wei, X.; Jiang, H.; Jie, C.-Y.; Xin, Z.-H. Impacts of enhanced nitrogen deposition and soil acidification on biomass production and nitrogen leaching in Chinese fir plantations. Can. J. For. Res. 2012, 42, 437–450. [Google Scholar] [CrossRef]
- Wei, X.H.; Blanco, J.A.; Jiang, H.; Kimmins, J.P. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci. Total Environ. 2012, 416, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Seely, B.; Hawkins, C.; Blanco, J.; Welham, C.; Kimmins, J. Evaluation of a mechanistic approach to mixedwood modelling. For. Chron. 2008, 84, 181–193. [Google Scholar] [CrossRef]
- Wei, X.H.; Blanco, J.A. Significant increase in ecosystem C can be achieved with sustainable forest management in subtropical plantation forests. PLoS One 2014, 9, e89688. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.C. Lauraceae. In Flora of Taiwan, 2nd ed.; Editorial Committee of the Flora of Taiwan, Ed.; Department of Botany, Nation Taiwan University: Taipei, Taiwan, 1996; p. 481. [Google Scholar]
- Chen, Z.H.; Wu, B.; Li, J.Y.; Zhao, J.G.; Zhou, X.Y.; Zhang, Y.K. Germination of the seeds and growth of seedlings of Cinnamomum camphora (L.) Presl. Plant Species Biol. 2004, 19, 55–58. [Google Scholar] [CrossRef]
- Miyawaki, A.; Fujiwara, K.; Hada, Y.; Itoh, S.; Miyata, I.; Miyoshi, N.; Murakami, Y.; Nakagoshi, N.; Nakanishi, H.; Nakamura, Y. Vegetation of Japan. Vol. 4. Chugoku; Shibundo: Tokyo, Japan, 1983; p. 540. [Google Scholar]
- Pakenham, T. Remarkable Trees of the World; Weidenfeld & Nicolson: London, UK, 2002. [Google Scholar]
- Penman, J.; Gytarsky, M.; Hiraishi, T.; Krug, T.; Kruger, D.; Pipatti, R.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K.; et al. Good Practice Guidance for Land Use, Land-Use Change and Forestry; IPCC National Greenhouse Gas Inventories Programme and Institute for Global Environmental Strategies: Kanagawa, Japan, 2003. [Google Scholar]
- Landsberg, J. Modelling forest ecosystems: State of the art, challenges, and future directions. Can. J. For. Res. 2003, 33, 385–397. [Google Scholar] [CrossRef]
- Kimmins, J.P. Scientific Foundations for the Simulation of Ecosystem Function and Management in FORCYTE-11; Information Report NOR-X-328; Forestry Canada: Edmonton, AB, Canada, 1993; p. 88. [Google Scholar]
- Yao, Y.J.; Kang, W.X.; Tian, D.L. Study of the biomass and productivity of Cinnamomum camphora plantation. J. Cent. South For. Univ. 2003, 23, 1–5. (In Chinese) [Google Scholar]
- Lee, H.-T.; Feng, F.-L. A forest carbon sequestration inventory system: An example of camphor trees in Taiwan. Taiwan J. For. Sci. 2008, 23 (Supplement), S11–S22. (In Chinese) [Google Scholar]
- Lei, P.F.; Xiang, W.H.; Tian, D.L.; Fang, X. Carbon storage and distribution in Cinnamomum camphor plantation. Chin. J. Ecol. 2004, 23, 25–30. (In Chinese) [Google Scholar]
- Lin, K.C.; Duh, C.T.; Huang, C.M. Estimate of carbon storage and sequestration of Fraxinus griffithii plantations in Taiwan. Q. J. Chin. For. 2010, 43, 261–276. (In Chinese) [Google Scholar]
- Welham, C.; van Rees, K.; Seely, B.; Kimmins, H. Projected long-term productivity in Saskatchewan hybrid poplar plantations: Weed competition and fertilizer effects. Can. J. For. Res. 2007, 37, 356–370. [Google Scholar] [CrossRef]
- Castrillón, Y.; Blanco, J.A.; Primicia, I.; Ansó, M.; Imbert, J.B.; Castillo, F.J. Evaluation of the ecological model FORECAST to simulate litter production in pine stands in the Pyrenees. In Proceedings of the 11th Congress of the AEET—Invitation to Ecology: Strengthening links with Society, Pamplona, Spain, 6–10 May 2013.
- Jie, C.; Jiang, H.; Zhou, G.; Wei, X.; Blanco, J.; Jiang, Z.; Xin, Z. Simulating the carbon storage of spruce forests based on the FORECAST model and remotely sensed data. In Proceedings of the Geoinformatics,2011 19th International Conference on Geoinformatics, Shanghai, China, 24–26 June 2011; IEEE: Piscateway, NJ, USA, 2011; pp. 1–6. [Google Scholar]
- Amoroso, M.M.; Turnblom, E.C. Comparing productivity of pure and mixed Douglas-fir and western hemlock plantations in the Pacific Northwest. Can. J. For. Res. 2006, 36, 1484–1496. [Google Scholar] [CrossRef]
- Lugo, A.E. Comparison of tropical tree plantations with secondary forests of similar age. Ecol. Monogr. 1992, 62, 1–41. [Google Scholar] [CrossRef]
- DeBell, D.S.; Whitesell, C.D.; Schubert, T.H. Using N2-fixing Albizia to increase growth of Eucalyptus plantations in Hawaii. For. Sci. 1989, 35, 64–75. [Google Scholar]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Mitchell, P.A.; Brockwell, J. Productivity of three young mixed-species plantations containing N2-fixing Acacia and non-N2-fixing Eucalyptus and Pinus trees in Southeastern Australia. For. Sci. 2007, 53, 426–434. [Google Scholar]
- Frivold, L.H.; Frank, J. Growth of mixed birch-coniferous stands in relation to pure coniferous stands at similar sites in south-eastern Norway. Scand. J. For. Res. 2002, 17, 139–149. [Google Scholar] [CrossRef]
- Kuo, Y.L.; Fan, K.S.; Hwang, C.W.; Lee, Y.P.; Wu, H.L.; Tsay, R.F. Gas exchange potential in sun-exposed leaves of 30 broadleaf tree species in Taiwan. Taiwan J. For. Sci. 2004, 19, 375–386. (In Chinese) [Google Scholar]
- Kanowski, J.; Catterall, C.P. Converting Stands of Camphor Laurel to raInforest: What are the Costs and Outcomes of Different Control Methods? Griffith University: Nathan, Australia, 2007; p. 16. [Google Scholar]
- Kanowski, J.; Catterall, C.P.; Neilan, W. Potential value of weedy regrowth for rainforest restoration. Ecol. Manag. Restor. 2008, 9, 88–99. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Hein, S.; Oosterbaan, A.; Wagner, S.; Clark, J.; Skovsgaard, J.P. A review of European ash (Fraxinus excelsior L.): Implications for silviculture. Forestry 2011, 84, 133–148. [Google Scholar] [CrossRef]
- Perry, E.; Hickman, G.W. A survey to determine the leaf nitrogen concentrations of 25 landscape tree species. J. Arboric. 2001, 27, 152–159. [Google Scholar]
- Wiedemann, E. Ertragskundliche und Waldbauliche Grundlagen der Forstwirtschaft; J. D. Sauerländer’s Verlag: Frankfurt, Germany, 1950. [Google Scholar]
- Mitscherlich, G. Wald, Wachstum und Umwelt. v. 1,Form und Wachstum von Baum und Bestand; J. D. Sauerländer’s Verlag: Frankfurt, Germany, 1978. [Google Scholar]
- Welham, C.; Seely, B.; Kimmins, H. The utility of the two-pass harvesting system: An analysis using the ecosystem simulation model FORECAST. Can. J. For. Res. 2002, 32, 1071–1079. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; McGraw-Hill: New York, NY, USA, 1990. [Google Scholar]
- Kimmins, J.P. Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Blanco, J.A. Forests may need centuries to recover their original productivity after continuous intensive management: An example from Douglas-fir stands. Sci. Total Environ. 2012, 437, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Hooker, T.D.; Compton, J.E. Forest ecosystem carbon and nitrogen accumulation during the first century after agricultural abandonment. Ecol. Appl. 2003, 13, 299–313. [Google Scholar] [CrossRef]
- Blanco, J.A.; Zavala, M.A.; Imbert, J.B.; Castillo, F.J. Sustainability of forest management practices: Evaluation through a simulation model of nutrient cycling. For. Ecol. Manag. 2005, 213, 209–228. [Google Scholar] [CrossRef]
- Josefsson, T.; Olsson, J.; Östlund, L. Linking forest history and conservation efforts: Long-term impact of low-intensity timber harvest on forest structure and wood-inhabiting fungi in northern Sweden. Biol. Conserv. 2010, 143, 1803–1811. [Google Scholar] [CrossRef]
- Gárate, M.; Blanco, J.A. Importancia de la caracterización de la masa de raíces en la simulación de ecosistemas forestales. Revista Ecosistemas 2013, 22, 66–73. (In Spanish) [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-H.; Lo, Y.-H.; Blanco, J.A.; Chang, S.-C. Resilience Assessment of Lowland Plantations Using an Ecosystem Modeling Approach. Sustainability 2015, 7, 3801-3822. https://doi.org/10.3390/su7043801
Wu C-H, Lo Y-H, Blanco JA, Chang S-C. Resilience Assessment of Lowland Plantations Using an Ecosystem Modeling Approach. Sustainability. 2015; 7(4):3801-3822. https://doi.org/10.3390/su7043801
Chicago/Turabian StyleWu, Chia-Hsin, Yueh-Hsin Lo, Juan A. Blanco, and Shih-Chieh Chang. 2015. "Resilience Assessment of Lowland Plantations Using an Ecosystem Modeling Approach" Sustainability 7, no. 4: 3801-3822. https://doi.org/10.3390/su7043801
APA StyleWu, C.-H., Lo, Y.-H., Blanco, J. A., & Chang, S.-C. (2015). Resilience Assessment of Lowland Plantations Using an Ecosystem Modeling Approach. Sustainability, 7(4), 3801-3822. https://doi.org/10.3390/su7043801