
Physical and Chemical Properties Evaluation of Liquid Organic Fertilizers (LOFs) and Their Effects on Promoting Rice Growth

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Production of Liquid Organic Fertilizers (LOFs)
2.2. Analysis of Chemical Parameters
2.3. Determination of Microbial Parameters
2.4. Determination of Plant Hormones
2.5. Analysis of Polyamine (PA) Content
2.6. Evaluation of Phytotoxicity of LOF
2.7. Effects of LOF on Growth and Physiology of Rice Seedlings
2.8. Statistical Analysis
3. Results
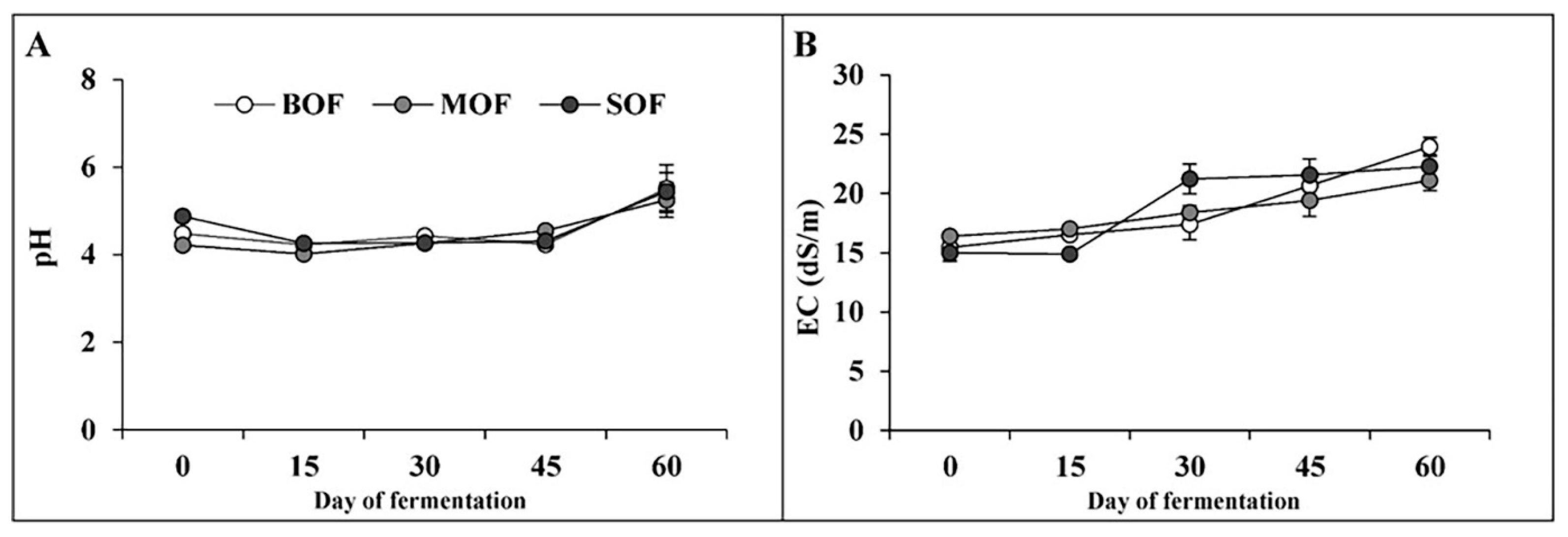
3.1. Physicochemical Parameters of Liquid Organic Fertilizers (LOFs)
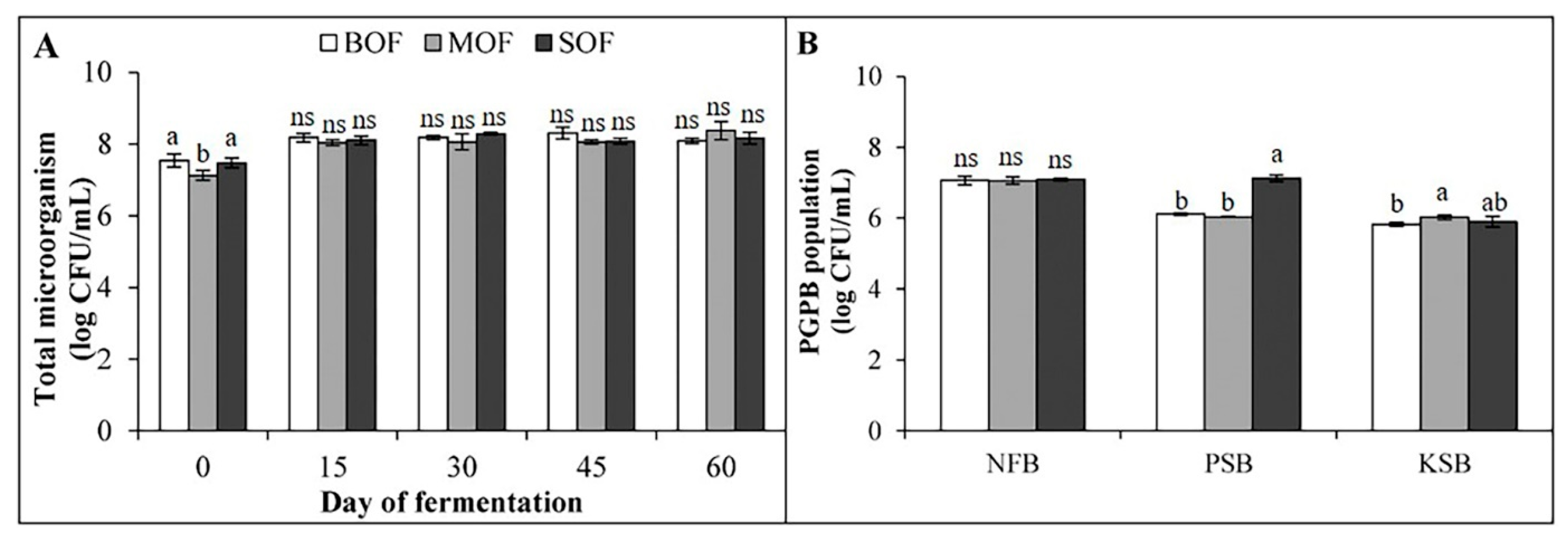
3.2. Change in Microbial Population Parameters in LOFs During Fermentation
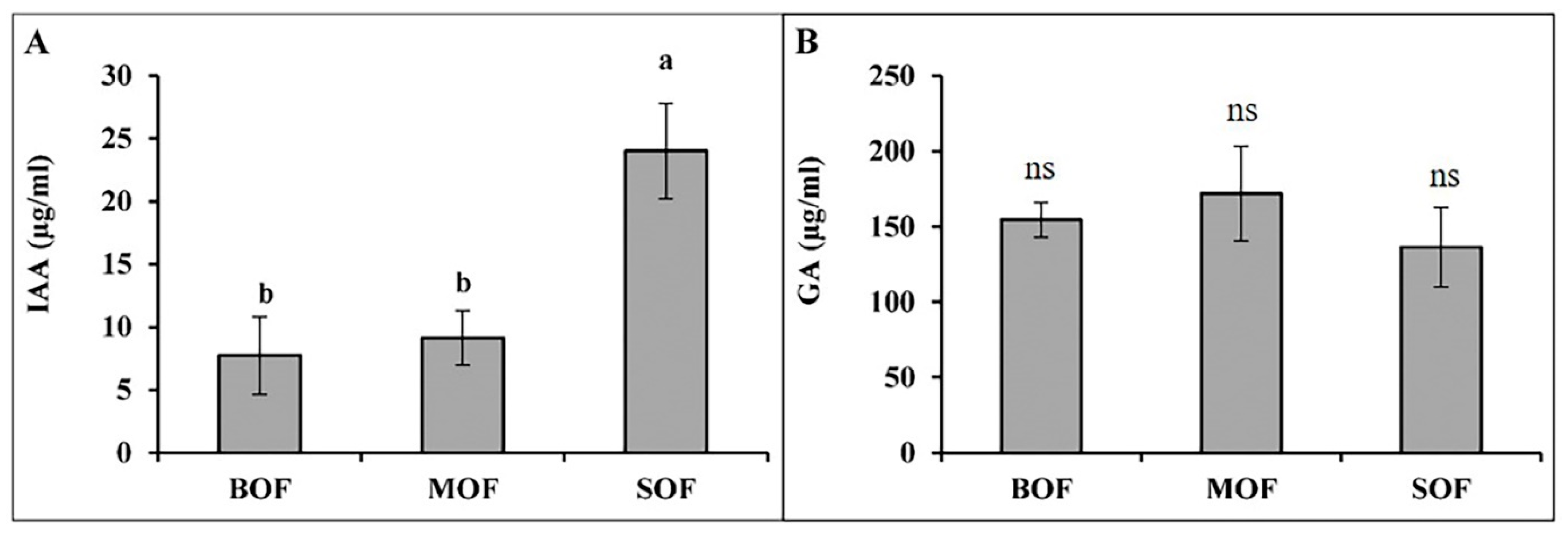
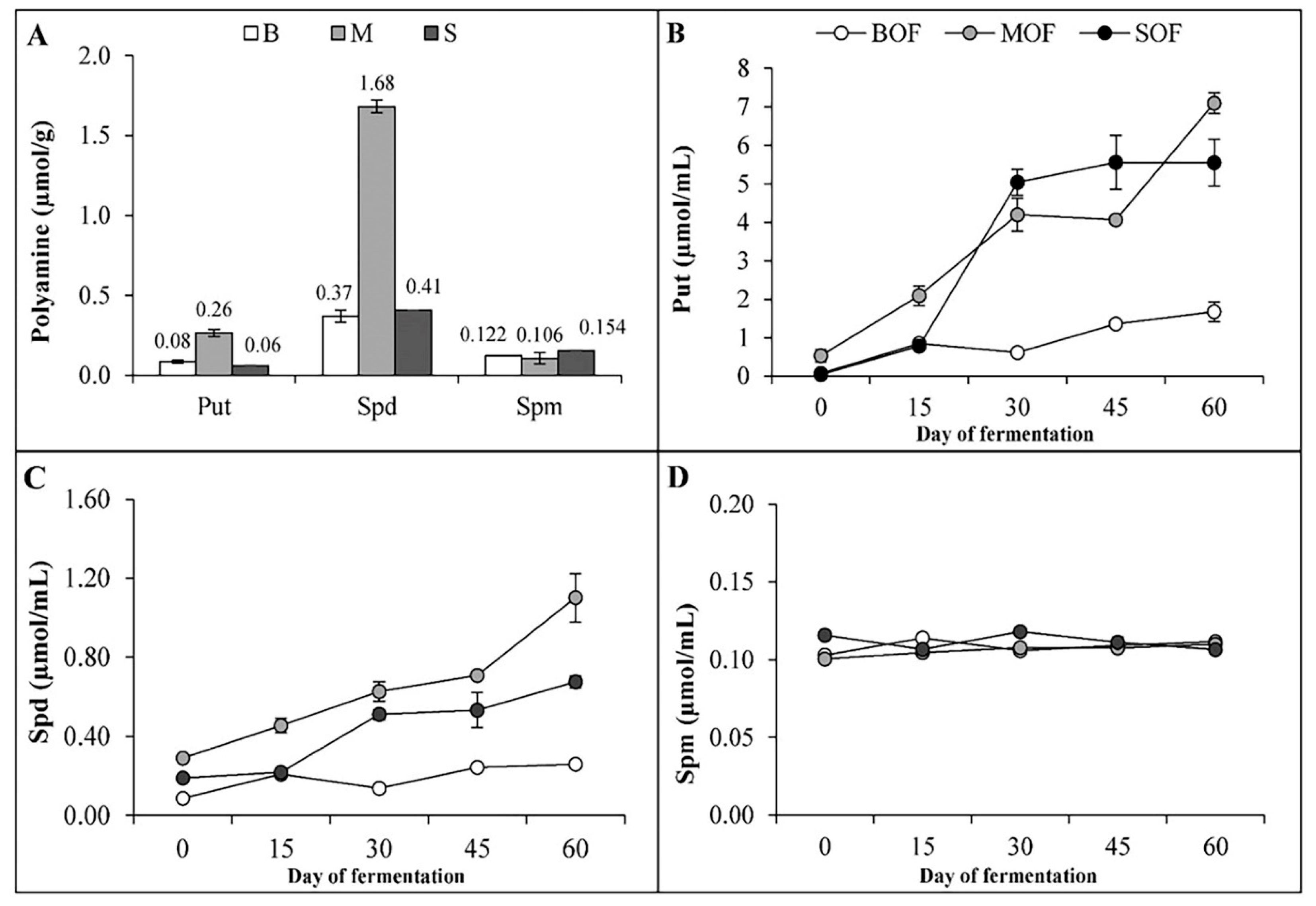
3.3. Phytohormones and Polyamines Concentration
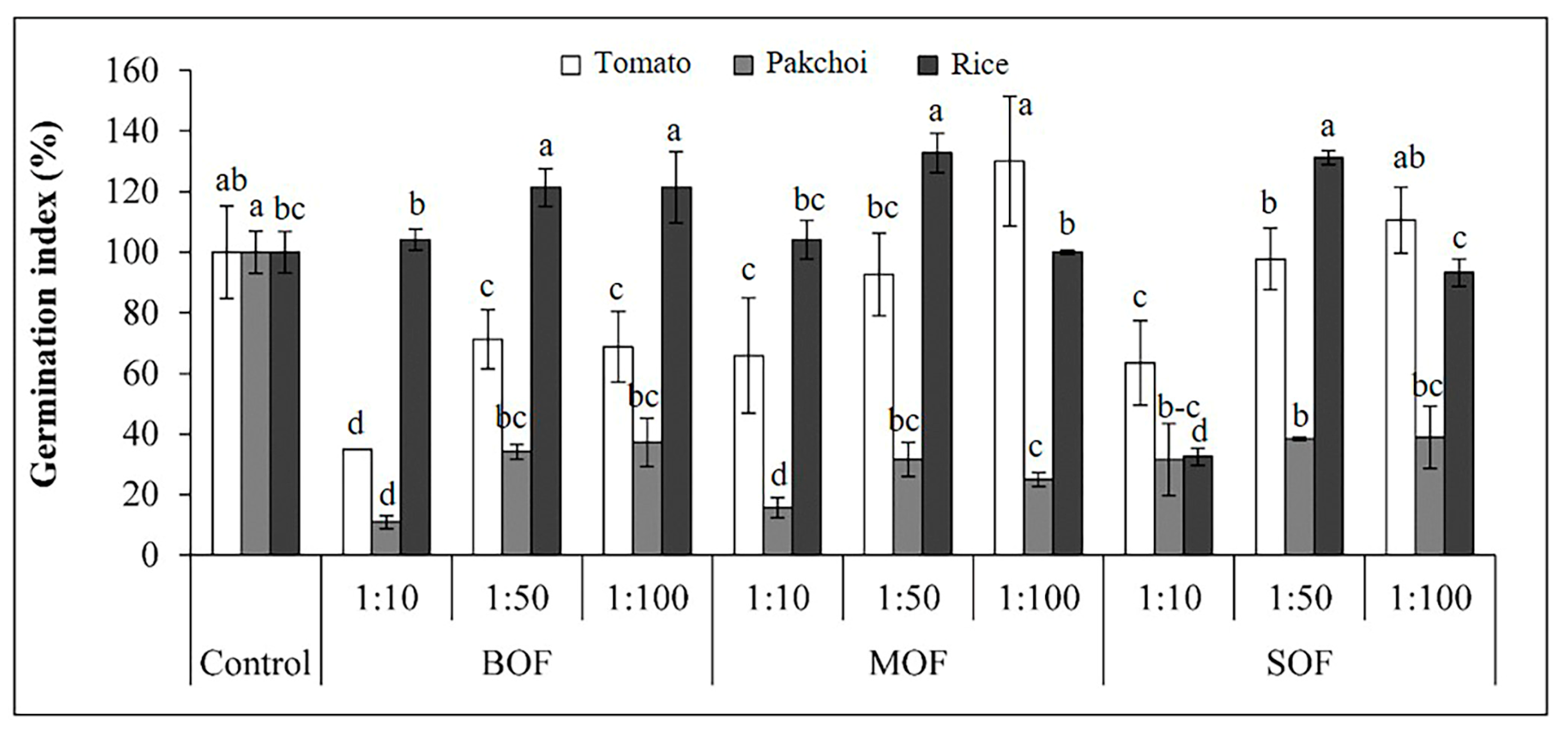
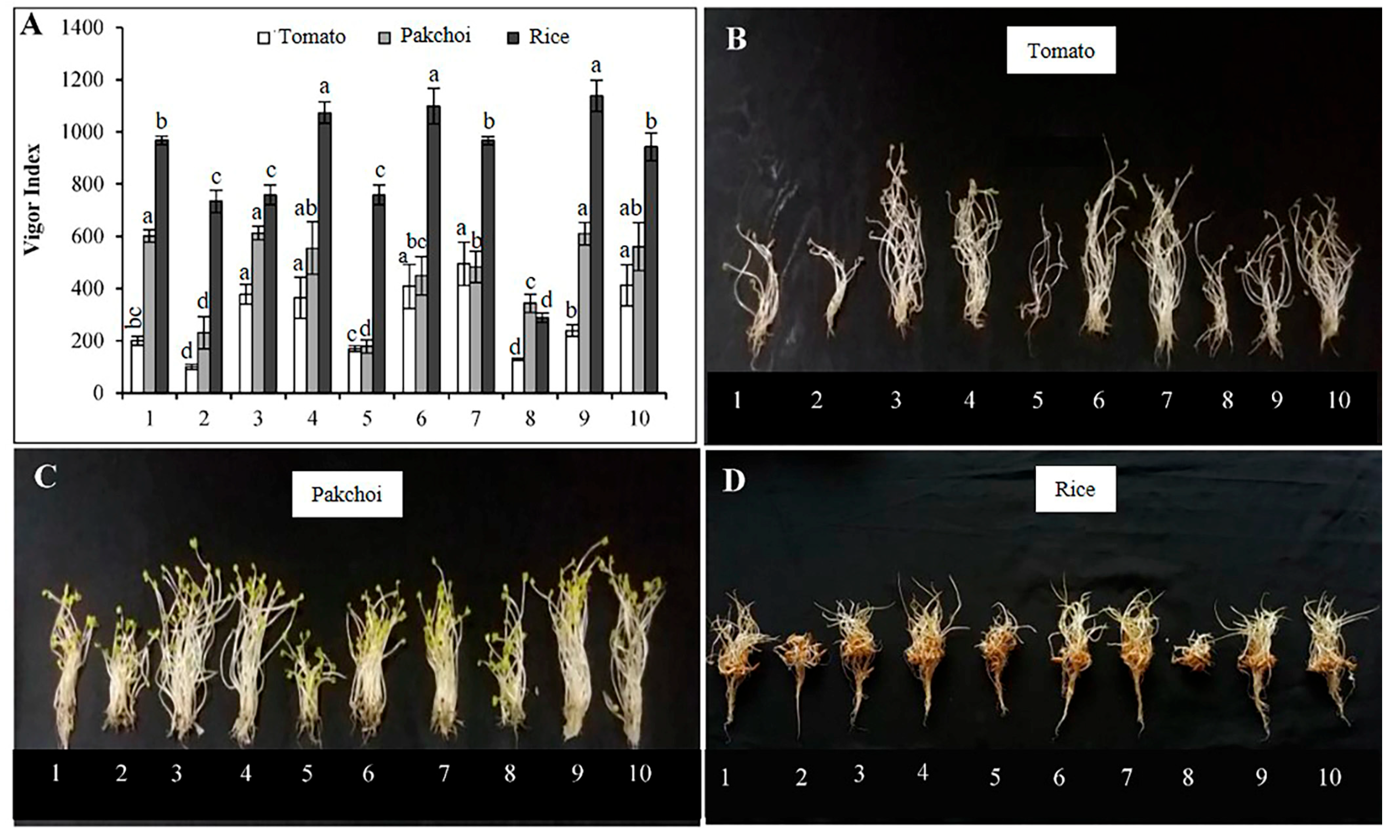
3.4. Effects of LOFs on Germination Index (GI), Vigor Index (VI), and Seedling Growth
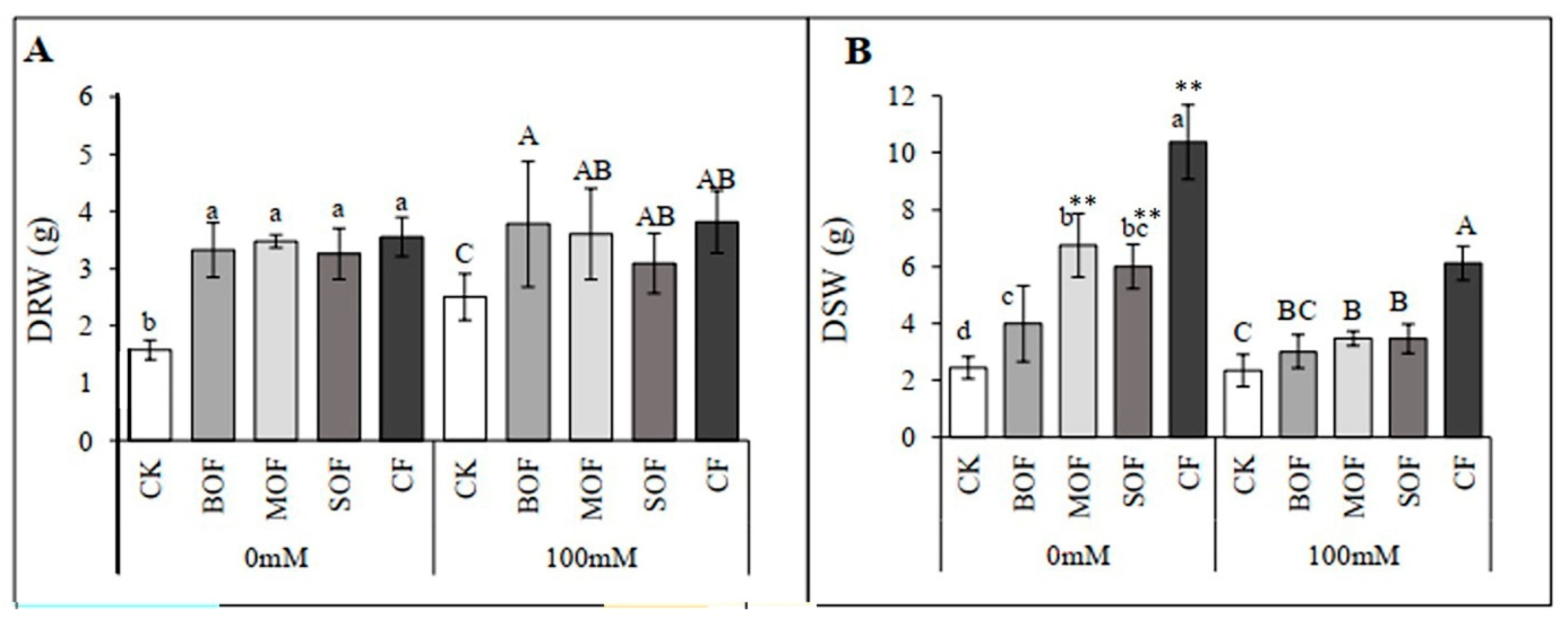
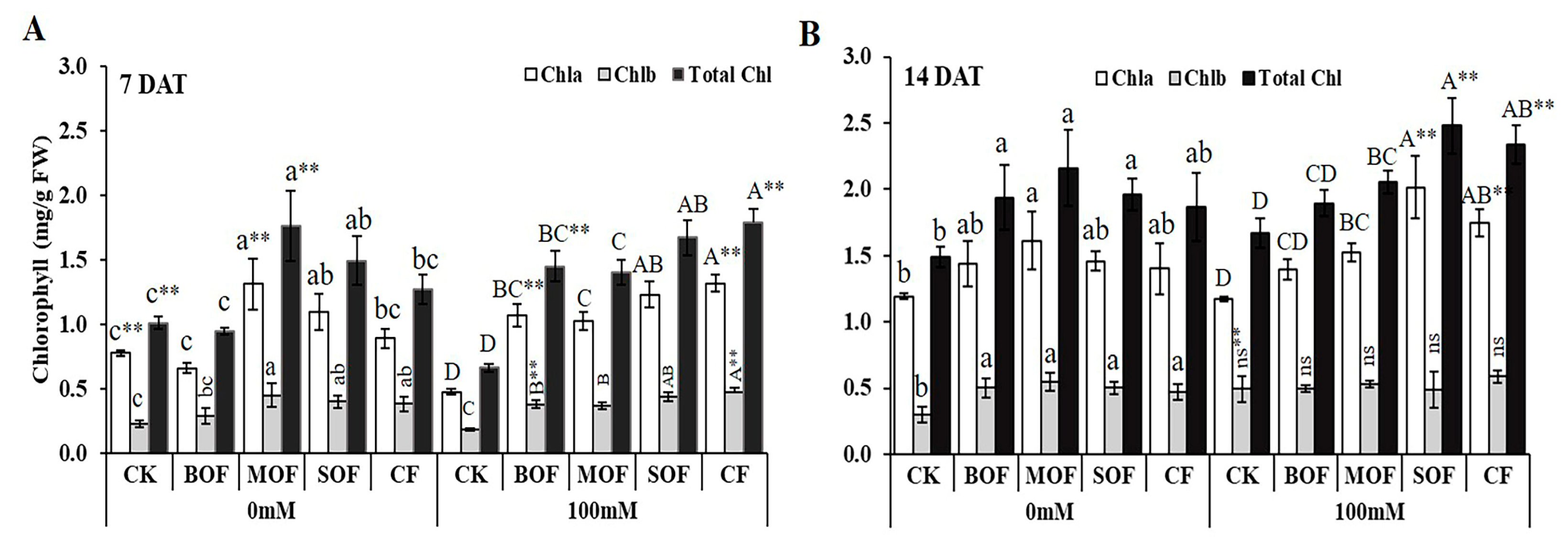
3.5. Effects of LOF on Growth and Physiology of Rice Under Salt Stress
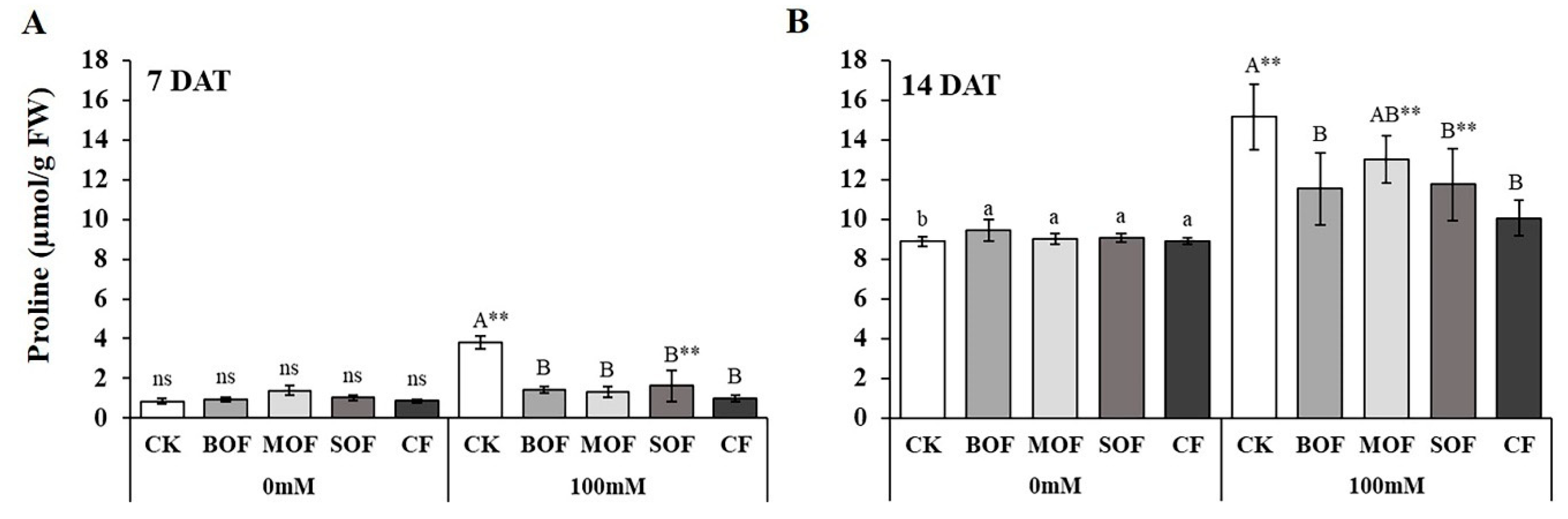
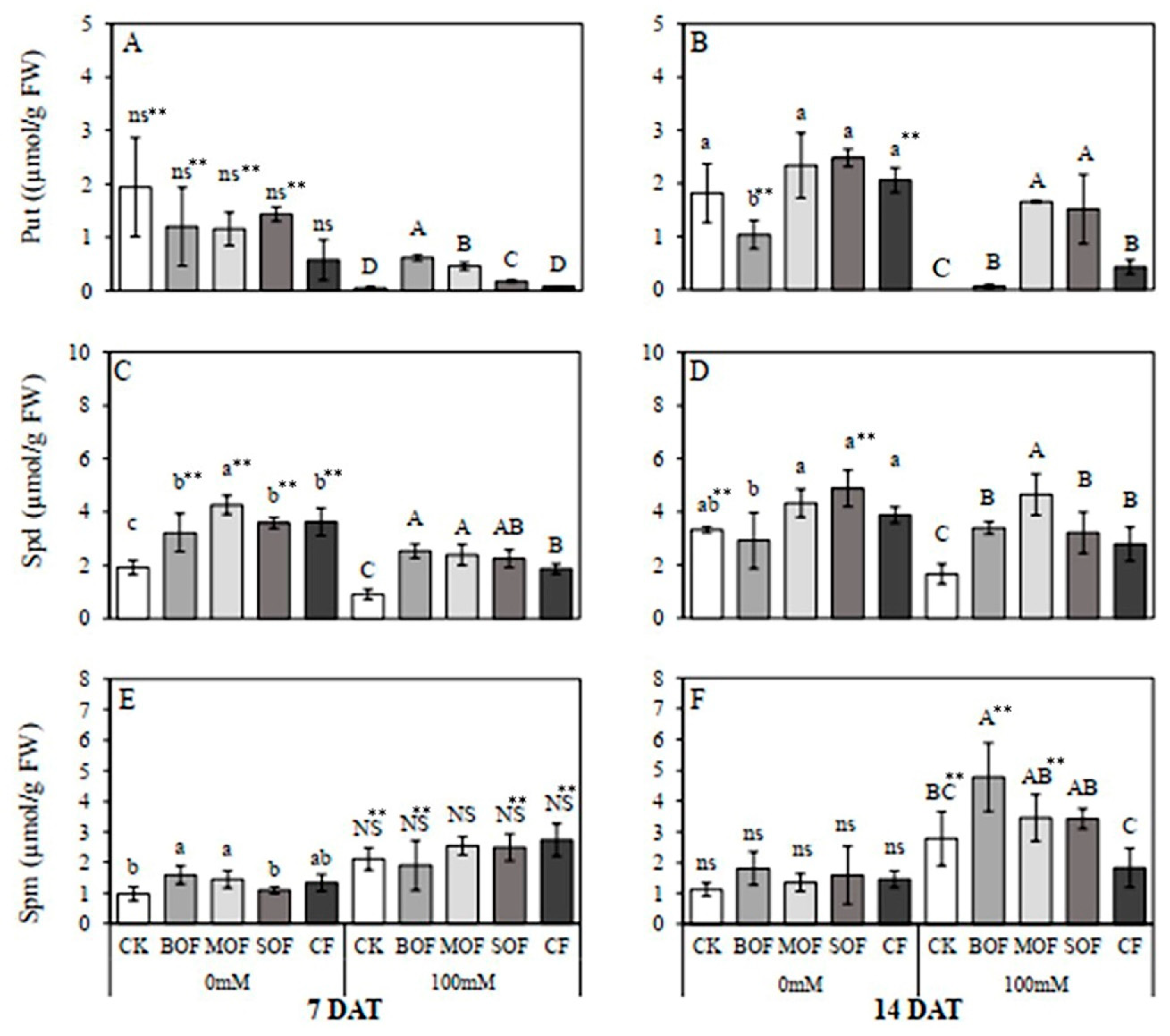
3.6. Effects of LOFs on Endogenous Polyamine (PA) Content in Rice Leaf
4. Discussion
4.1. Physico-Chemical Characteristics of Liquid Organic Fertilizers (LOFs)
4.2. Phytoxicity Evaluation of the LOFs Based on Seed Germination Test
4.3. LOFs Alleviated Salt-Stress Impact on Growth and Physiology of Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LOF | Liquid organic fertilizer |
| BOF | Brassica-based fertilizer |
| MOF | Mushroom-based fertilizer |
| SOF | Soybean-based fertilizer |
| CF | Chemical fertilizer |
| EC | Electrical conductivity |
| IAA | Indole acetic acid |
| GA | Gibberellic acid |
| PA | Polyamine |
| Put | Putrescine |
| Spd | Spermidine |
| Spm | Spermine |
| PGPB | Plant growth-promoting bacteria |
| NFB | Nitrogen-fixing bacteria |
| PSB | Phosphorus solubilizing bacteria |
| KSB | Potassium solubilizing bacteria |
| GI | Germination index |
| VI | Vigor index |
| CK | Control without fertilizer |
| DAT | Days after NaCl treatment |
| DRW | Dried root weight |
| DSW | Dried shoot weight |
| Pn | Net photosynthesis rate |
References
- Liu, Y.; Lan, X.; Hou, H.; Ji, J.; Liu, X.; Lv, Z. Multifaceted ability of organic fertilizers to improve crop productivity and abiotic stress tolerance: Review and perspectives. Agronomy 2024, 14, 1141. [Google Scholar] [CrossRef]
- Culas, R.J.; Anwar, M.R.; Maraseni, T.N. A framework for evaluating benefits of organic fertilizer use in agriculture. J. Agric. Food Res. 2025, 19, 101576. [Google Scholar] [CrossRef]
- Wang, L.; Sun, X.; Li, S.; Zhang, T.; Zhang, W.; Zhai, P. Application of organic amendments to a coastal saline soil in North China: Effects on soil physical and chemical properties and tree growth. PLoS ONE 2014, 98, e89185. [Google Scholar] [CrossRef] [PubMed]
- Wichern, F.R.; Islam, M.; Hemkemeyer, M.; Watson, C.; Joergensen, R. Organic amendments alleviate salinity effects on soil microorganisms and mineralisation processes in aerobic and anaerobic paddy rice soils. Front. Sustain. Food Syst. 2020, 4, 30. [Google Scholar] [CrossRef]
- Zayed, B.A.; Elkhoby, W.M.; Salem, A.K.; Ceesay, M.; Uphoff, N.T. Effect of integrated nitrogen fertilizer on rice productivity and soil fertility under saline soil conditions. J. Plant Biol. Res. 2013, 2, 14–24. [Google Scholar]
- Litardo, R.C.M.; Bendezú, S.J.G.; Zenteno, M.D.C.; Pérez-Almeida, I.B.; Parismoreno, L.L.; García, E.D.L. Effect of mineral and organic amendments on rice growth and yield in saline soils. J. Saudi Soc. Agric. Sci. 2022, 21, 29–37. [Google Scholar] [CrossRef]
- Phibunwatthanawong, T.; Riddech, N. Liquid organic fertilizer production for growing vegetables under hydroponic condition. Int. J. Recycl. Org. Waste Agric. 2019, 8, 369–380. [Google Scholar] [CrossRef]
- Rengasamy, K.R.R.; Kulkarni, M.G.; Papenfus, H.B.; Van Staden, J. Quantification of plant growth biostimulants, phloroglucinol and eckol, in four commercial seaweed liquid fertilizers and some by-products. Algal Res. 2016, 20, 57–60. [Google Scholar] [CrossRef]
- Martinez-Alcantara, B.; Marinez-Cuenca, M.-R.; Bermejo, A.; Legaz, F.; Quiñones, A. Liquid organic fertilizers for sustainable agriculture: Nutrient uptake of organic versus mineral fertilizers in citrus trees. PLoS ONE 2016, 11, e0161619. [Google Scholar] [CrossRef]
- Vinoth, S.; Sundari; Gurusaravanan, P.; Sivakumar, S.; Siva, G.; Kumar, G.P.; Manju; Velmurugan, K.; Lakshminarayana, V.; Jayabalan, N. Evaluation of seagrass liquid extract on salt stress alleviation in tomato plants. Asian J. Plant Sci. 2017, 16, 172–183. [Google Scholar] [CrossRef]
- Sunaryo, Y.; Purnomo, D.; Darini, M.T.; Cahyani, V.R. Nutrients content and quality of liquid fertilizer made from goat manure. J. Phys. Conf. Ser. 2018, 1022, 012053. [Google Scholar] [CrossRef]
- Widjajanto, D.W.; Purbajanti, E.D.; Sumarsono; Utama, C.S. The role of local microorganisms generated from rotten fruits and vegetables in producing liquid organic fertilizer. J. Appl. Chem. Sci. 2017, 4, 325–332. [Google Scholar] [CrossRef]
- Kebrom, T.H.; Woldesenbet, S.; Bayabil, H.K.; Garcia, M.; Gao, M.; Ampim, P.; Awal, R.; Fares, A. Evaluation of phytotoxicity of three organic amendments to collard greens using the seed germination bioassay. Environ. Sci. Pollut. Res. 2019, 26, 5454–5462. [Google Scholar] [CrossRef]
- Wong, W.S.; Tan, S.N.; Ge, L.; Chen, X.; Yong, J.W.H. The importance of phytohormones and microbes in biofertilizers. In Bacterial Metabolites in Sustainable Agroecosystem; Maheshwari, D.K., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 105–158. [Google Scholar] [CrossRef]
- Biondi, S.; Antognoni, F.; Marincich, L.; Lianza, M.; Tejos, R.; Ruiz, K.B. The polyamine “multiverse” and stress mitigation in crops: A case study with seed priming in quinoa. Sci. Hortic. 2022, 304, 111292. [Google Scholar] [CrossRef]
- Amiri, H.; Banakar, M.H.; Hemmati Hassan Gavyar, P. Polyamines: New plant growth regulators promoting salt stress tolerance in plants. J. Plant Growth Regul. 2024, 43, 4923–4940. [Google Scholar] [CrossRef]
- Pottosin, I.; Olivas-Aguirre, M.; Dobrovinskaya, O.; Zepeda-Jazo, I.; Shabala, S. Modulation of ion transport across plant membranes by polyamines: Understanding specific modes of action under stress. Front. Plant Sci. 2021, 11, 616077. [Google Scholar] [CrossRef]
- Wu, J.; Shu, S.; Li, C.; Sun, J.; Guo, S. Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol. Biochem. 2018, 128, 152–162. [Google Scholar] [CrossRef]
- Shabala, S.; Munns, R. Salinity stress: Physiological constraints and adaptive mechanisms. In Plant Stress Physiology; Shabala, S., Ed.; CABI: Allen, TX, USA, 2012; pp. 59–93. [Google Scholar] [CrossRef]
- Hassan, N.; Ebeed, H.; Aljaarany, A. Exogenous application of spermine and putrescine mitigate adversities of drought stress in wheat by protecting membranes and chloroplast ultra-structure. Physiol. Mol. Biol. Plants 2020, 26, 233–245. [Google Scholar] [CrossRef]
- Atiya Ali, M.; Poortvliet, E.; Strömberg, R.; Yngve, A. Polyamines in foods: Development of a food database. Food Nutr. Res. 2011, 55, 5572. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Sarwar, M.; Arshad, M.; Martens, D.A.; Frankenberger, W.T. Tryptophan-dependent biosynthesis of auxins in soil. Plant Soil 1992, 147, 207–215. [Google Scholar] [CrossRef]
- Holbrook, A.A.; Edge, W.J.W.; Bailey, F. Spectrophotometric method for determination of gibberellic acid. Adv. Chem. Ser. 2009, 28, 159–167. [Google Scholar] [CrossRef]
- Pedrol, N.; Tiburcio, A.F. Polyamines determination by TLC and HPLC. In Handbook of Plant Ecophysiology Techniques; Roger, M.J.R., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 335–363. [Google Scholar] [CrossRef]
- Zucconi, F.; Forte, M.; Monaco, A.; De Bertoldi, M. Biological evaluation of compost maturity. Biocycle 1981, 22, 27–29. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Wynne, A.T.; Meyer, J.H. An economic assessment of using molasses and condensed molasses solids as a fertiliser in the South African sugar industry. In Proceedings of the South African Sugar Technology Association, Mount Edgecombe, South Africa, 30 July–2 August 2002; pp. 71–78. [Google Scholar]
- Olstorpe, M.; Lyberg, K.; Lindberg, J.E.; Schnürer, J.; Passoth, V. Population diversity of yeasts and lactic acid bacteria in pig feed fermented with whey, wet wheat distillers’ grains, or water at different temperatures. Appl. Environ. Microbiol. 2008, 74, 1696–1703. [Google Scholar] [CrossRef]
- Sundberg, C.; Smårs, S.; Jönsson, H. Low pH as an inhibiting factor in the transition from mesophilic to thermophilic phase in composting. Bioresour. Technol. 2004, 95, 145–150. [Google Scholar] [CrossRef]
- Kadir, A.A.; Rahman, N.A.; Azhari, N.W. The utilization of Banana Peel in the fermentation liquid in food waste composting. IOP Conf. Ser. Mater. Sci. Eng. 2016, 136, 012055. [Google Scholar] [CrossRef]
- Okamoto, A.; Sugi, E.; Koizumi, Y.; Yanagida, F.; Udaka, S. Polyamine content of ordinary foodstuffs and various fermented foods. Biosci. Biotechnol. Biochem. 2009, 61, 1582–1584. [Google Scholar] [CrossRef]
- Prachyakij, P.; Charernjiratrakul, W.; Kantachote, D. Improvement in the quality of a fermented seaweed beverage using an antiyeast starter of Lactobacillus plantarum DW3 and partial sterilization. World J. Microbiol. Biotechnol. 2008, 24, 1713–1720. [Google Scholar] [CrossRef]
- Pan, I.; Dam, B.; Sen, S.K. Composting of common organic wastes using microbial inoculants. 3 Biotech 2012, 2, 127–134. [Google Scholar] [CrossRef]
- Podile, A.; Kishore, G. Plant growth-promoting rhizobacteria. In Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2006; pp. 195–230. [Google Scholar] [CrossRef]
- Maheshwari, D.M.; Dheeman, S.; Agarwal, M. Phytohormone-producing PGPR for sustainable agriculture. In Bacterial Metabolites in Sustainable Agroecosystem; Maheshwari, D.K., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 159–182. [Google Scholar] [CrossRef]
- Kantachote, D.; Kowpong, K.; Charernjiratrakul, W.; Pengnoo, A. Microbial succession in a fermenting of wild forest noni (Morinda coreia Ham) fruit plus molasses and its role in producing a liquid fertilizer. Electron. J. Biotechnol. 2009, 12, 1–11. [Google Scholar] [CrossRef]
- Shirasawa, H.; Nishiyama, C.; Hirano, R.; Koyanagi, T.; Okuda, S.; Takagi, H.; Kurihara, S. Isolation of the high polyamine-producing bacterium Staphylococcus epidermidis FB146 from fermented foods and identification of polyamine-related genes. Biosci. Microbiota Food Health 2023, 42, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-molecules with diverse functions in plant and human health and disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef]
- Zaman, M.Z.; Abu Bakar, F.; Jinap, S.; Bakar, J. Novel starter cultures to inhibit biogenic amines accumulation during fish sauce fermentation. Int. J. Food Microbiol. 2011, 145, 84–91. [Google Scholar] [CrossRef]
- Latorre-Moratalla, M.L.; Bover-Cid, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Control of biogenic amines in fermented sausages: Role of starter cultures. Front. Microbiol. 2012, 3, 169. [Google Scholar] [CrossRef]
- Jin, Y.H.; Lee, J.H.; Park, Y.K.; Lee, J.-H.; Mah, J.-H. The occurrence of biogenic amines and determination of biogenic amine-producing lactic acid bacteria in Kkakdugi and Chonggak Kimchi. Foods 2019, 8, 73. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y.; Hodgkiss, I.J. Effects of composting on phytotoxicity of spent pig-manure sawdut litter. Environ. Pollut. 1996, 93, 249–256. [Google Scholar] [CrossRef]
- Warman, P.R. Evaluation of seed germination and growth tests for assessing compost maturity. Compost Sci. Util. 1999, 7, 33–37. [Google Scholar] [CrossRef]
- Selim, S.M.; Zayed, M.S.; Houssam, M.A. Evaluation of phytotoxicity of compost during composting process. Nat. Sci. 2012, 10, 69–77. [Google Scholar]
- Irik, H.A.; Bikmaz, G. Effect of different salinity on seed germination, growth parameters and biochemical contents of pumpkin (Cucurbita pepo L.) seeds cultivars. Sci. Rep. 2024, 14, 6929. [Google Scholar] [CrossRef] [PubMed]
- Hepsibha, B.T.; Tha, A.G. Revealing the non-phytotoxic effect of gunapaselam (fermented fish waste) by a dose dependent in vitro study. Int. J. Pharma Bio Sci. 2017, 8, 643–648. [Google Scholar] [CrossRef]
- Jagadabhi, P.S.; Wani, S.P.; Kaushal, M.; Patil, M.; Vemula, A.K.; Rathore, A. Physico-chemical, microbial and phytotoxicity evaluation of composts from sorghum, finger millet and soybean straws. Int. J. Recycl. Org. Waste Agric. 2019, 8, 279–293. [Google Scholar] [CrossRef]
- Tanimoto, E. Regulation of root growth by plant hormones—Roles for auxin and gibberellin. Crit. Rev. Plant Sci. 2005, 24, 249–265. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Mirza, J.I.; Bagni, N. Effects of exogenous polyamines and difluoromethylornithine on seed germination and root growth of Arabidopsis thaliana. Plant Growth Regul. 1991, 10, 163–168. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Rehman, H.; Hussain, M. Seed priming with polyamines improves the germination and early seedling growth in fine rice. J. New Seeds 2008, 9, 145–155. [Google Scholar] [CrossRef]
- Afzal, I.; Munir, F.; Ayub, C.M.; Basra, S.M.A.; Hameed, A.; Nawaz, A. Changes in antioxidant enzymes, germination capacity and vigour of tomato seeds in response of priming with polyamines. Seed Sci. Technol. 2009, 37, 765–770. [Google Scholar] [CrossRef]
- Jang, S.J.; Choi, Y.J.; Park, K.Y. Effects of polyamines on shoot and root development in arabidopsis seedlings and carnation cultures. J. Plant Biol. 2002, 45, 230–236. [Google Scholar] [CrossRef]
- Saleethong, P.; Sanitchon, J.; Kong-ngern, K.; Theerakulpisut, P. Pretreatment with spermidine reverses inhibitory effects of salt stress in two rice (Oryza sativa L.) cultivars differing in salinity tolerance. Asian J. Plant Sci. 2011, 10, 245–254. [Google Scholar] [CrossRef]
- Islam, A.; Pang, J.; Meng, F.; Li, Y.; Xu, N.; Yang, C.; Liu, J. Putrescine, spermidine, and spermine play distinct roles in rice salt tolerance. J. Integr. Agric. 2020, 19, 643–655. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Peng, Y.; Viña, A.; Arkebauer, T.; Schepers, J.S. Efficiency of chlorophyll in gross primary productivity: A proof of concept and application in crops. J. Plant Physiol. 2016, 201, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Roychoudhury, A. Seed priming with spermine and spermidine regulates the expression of diverse groups of abiotic stress-responsive genes during salinity stress in the seedlings of indica rice varieties. Plant Gene 2017, 11, 124–132. [Google Scholar] [CrossRef]
- Shao, J.; Huang, K.; Batool, M.; Idrees, F.; Afzal, R.; Haroon, M.; Noushahi, H.A.; Wu, W.; Hu, Q.; Lu, X.; et al. Versatile roles of polyamines in improving abiotic stress tolerance of plants. Front. Plant Sci. 2022, 13, 1003155. [Google Scholar] [CrossRef]
- Do, P.T.; Drechsel, O.; Heyer, A.G.; Hincha, D.K.; Zuther, E. Changes in free polyamine levels, expression of polyaminebiosynthesis genes, and performance of rice cultivars under salt stress: A comparison with responses to drought. Front. Plant Sci. 2014, 5, 182. [Google Scholar] [CrossRef]
- Chuntaburee, S.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Alleviation of salt stress in seedlings of black glutinous rice by seed priming with spermidine and gibberellic acid. Not. Bot. Horti Agrobot. Cluj-Napoca 2014, 42, 405–413. [Google Scholar] [CrossRef]
- Theerakulpisut, P.; Madee, P.; Pamuta, D.; Nounjan, N. Exogenous application of spermidine (SPD) and wood vinegar improves salt tolerance in salt-sensitive rice (Oryza sativa L.). Pak. J. Bot. 2021, 53, 1–9. [Google Scholar] [CrossRef]
- Ghosh, N.; Das, S.P.; Mandal, C.; Gupta, S.; Das, K.; Dey, N.; Adak, M.K. Variations of antioxidative responses in two rice cultivars with polyamine treatment under salinity stress. Physiol. Mol. Biol. Plants 2012, 18, 301–313. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Amelioration of salinity stress by exogenously applied spermidine or spermine in three varieties of indica rice differing in their level of salt tolerance. J. Plant Physiol. 2011, 168, 317–328. [Google Scholar] [CrossRef]
- Jiang, D.X.; Chu, X.; Li, M.; Hou, J.J.; Tong, X.; Gao, Z.P.; Chen, G.X. Exogenous spermidine enhances salt-stressed rice photosynthetic performance by stabilizing structure and function of chloroplast and thylakoid membranes. Photosynthetica 2020, 58, 61–71. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Maodong, Q.; Dong, X.X.; Ahmad, P.; Abd_Allah, E.F.; Zhang, L. Spermine application alleviates salinity induced growth and photosynthetic inhibition in Solanum lycopersicum by modulating osmolyte and secondary metabolite accumulation and differentially regulating antioxidant metabolism. Plant Physiol. Biochem. 2019, 144, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Das Bhowmik, S.; Long, H.; Cheng, Y.; Mundree, S.; Hoang, L.T.M. Rapid accumulation of proline enhances salinity tolerance in Australian wild rice Oryza australiensis domin. Plants 2021, 10, 2044. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Original Fresh Materials (Dry Weight) | After 60 Days of Fermentation (in Liquid Extraction) | |||||
|---|---|---|---|---|---|---|
| Parameters | Brassica | Mushrooms | Soybean | BOF | MOF | SOF |
| Total N (%) | 0.41 ± 0.013 c | 0.56 ± 0.03 b | 0.65 ± 0.006 a | 0.32 ± 0.02 b | 0.44 ± 0.10 b | 0.99 ± 0.11 a |
| Total P (%) | 0.07 ± 0.003 b | 0.10 ± 0.01 a | 0.05 ± 0.001 c | 0.06 ± 0.004 b | 0.1 ± 0.06 a | 0.11 ± 0.02 a |
| Total K (%) | 0.51 ± 0.03 a | 0.46 ± 0.004 b | 0.23 ± 0.001 c | 1.23 ± 0.02 a | 1.34 ± 0.01 a | 1.2 ± 0.01 a |
| Total Na (mg/L) | - | - | - | 218 ± 9.8 ab | 236 ± 28 a | 182 ± 14 b |
| OM (%) | 78.13 ± 3.08 b | 76.47 ± 1.50 b | 88.02 ± 1.35 a | 6.94 ± 0.24 b | 9.39 ± 1.47 a | 11.62 ± 1.27 a |
| OC (%) | 45.24 ± 1.92 b | 44.39 ± 0.82 b | 50.89 ± 2.78 a | 4.06 ± 0.12 c | 5.48 ± 0.76 b | 6.75 ± 0.77 a |
| C/N | 107.3 ± 0.46 a | 80.4 ± 0.35 b | 79.2 ± 0.32 c | 12.96 ± 0.56 a | 12.77 ± 2.16 a | 6.87 ± 0.95 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riddech, N.; Ngo, M.N.; Boonlue, S.; Dongsansuk, A.; Santanoo, S.; Theerakulpisut, P. Physical and Chemical Properties Evaluation of Liquid Organic Fertilizers (LOFs) and Their Effects on Promoting Rice Growth. Sustainability 2025, 17, 3087. https://doi.org/10.3390/su17073087
Riddech N, Ngo MN, Boonlue S, Dongsansuk A, Santanoo S, Theerakulpisut P. Physical and Chemical Properties Evaluation of Liquid Organic Fertilizers (LOFs) and Their Effects on Promoting Rice Growth. Sustainability. 2025; 17(7):3087. https://doi.org/10.3390/su17073087
Chicago/Turabian StyleRiddech, Nuntavun, My Ngan Ngo, Sophon Boonlue, Anoma Dongsansuk, Supranee Santanoo, and Piyada Theerakulpisut. 2025. "Physical and Chemical Properties Evaluation of Liquid Organic Fertilizers (LOFs) and Their Effects on Promoting Rice Growth" Sustainability 17, no. 7: 3087. https://doi.org/10.3390/su17073087
APA StyleRiddech, N., Ngo, M. N., Boonlue, S., Dongsansuk, A., Santanoo, S., & Theerakulpisut, P. (2025). Physical and Chemical Properties Evaluation of Liquid Organic Fertilizers (LOFs) and Their Effects on Promoting Rice Growth. Sustainability, 17(7), 3087. https://doi.org/10.3390/su17073087