
Investigating the Impact of Salinity on Soil Organic Matter Dynamics Using Molecular Biomarkers and Principal Component Analysis

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
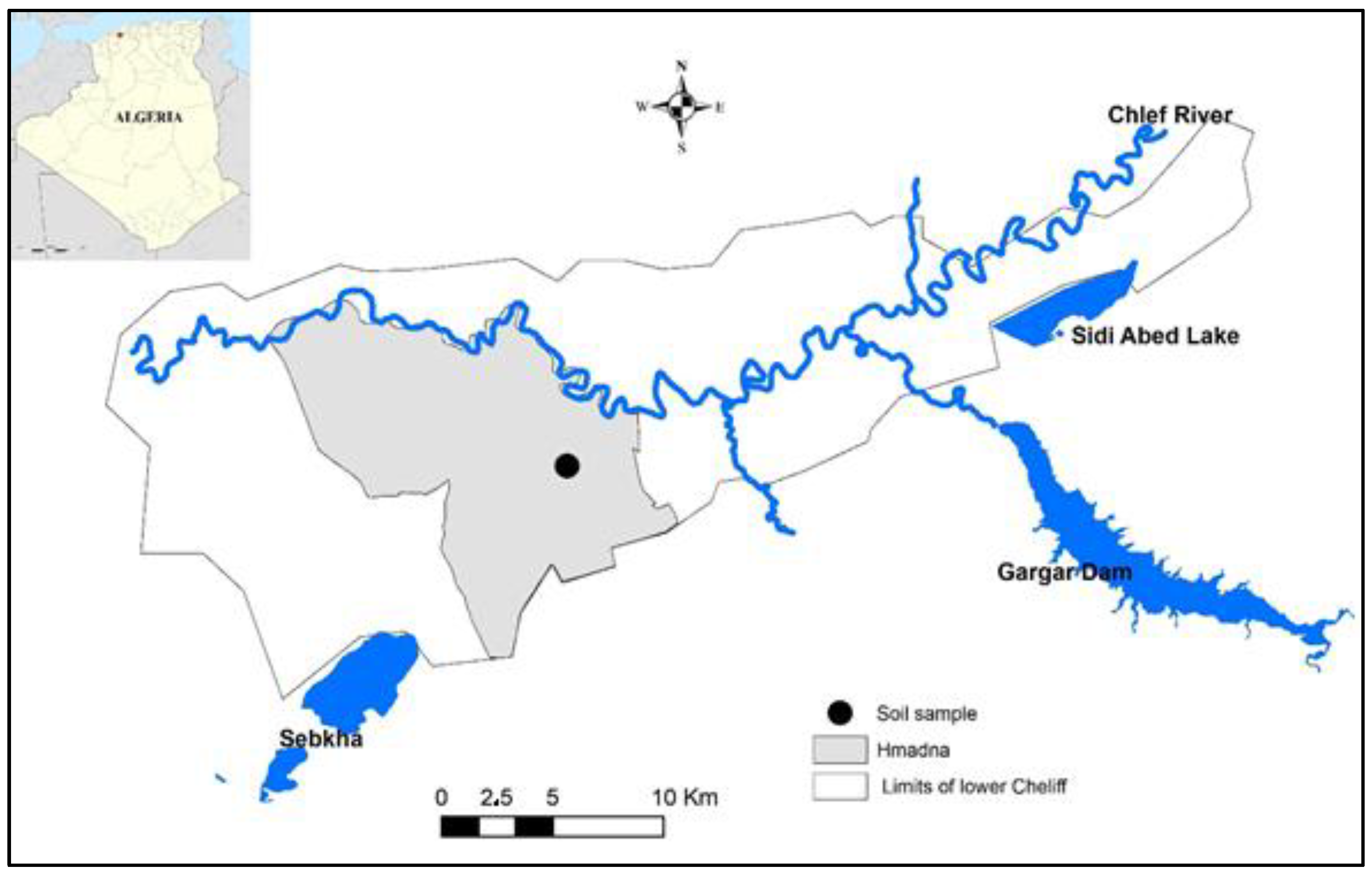
2.1. Site Description
2.2. Sampling Strategy
2.3. Salinity
2.4. Molecular Analysis
2.5. Gas Chromatography Coupled with Mass Spectrometry (GC-MS)
2.6. Molecular Biomarker Ratios
2.7. Data Analysis Method
3. Results and Discussion
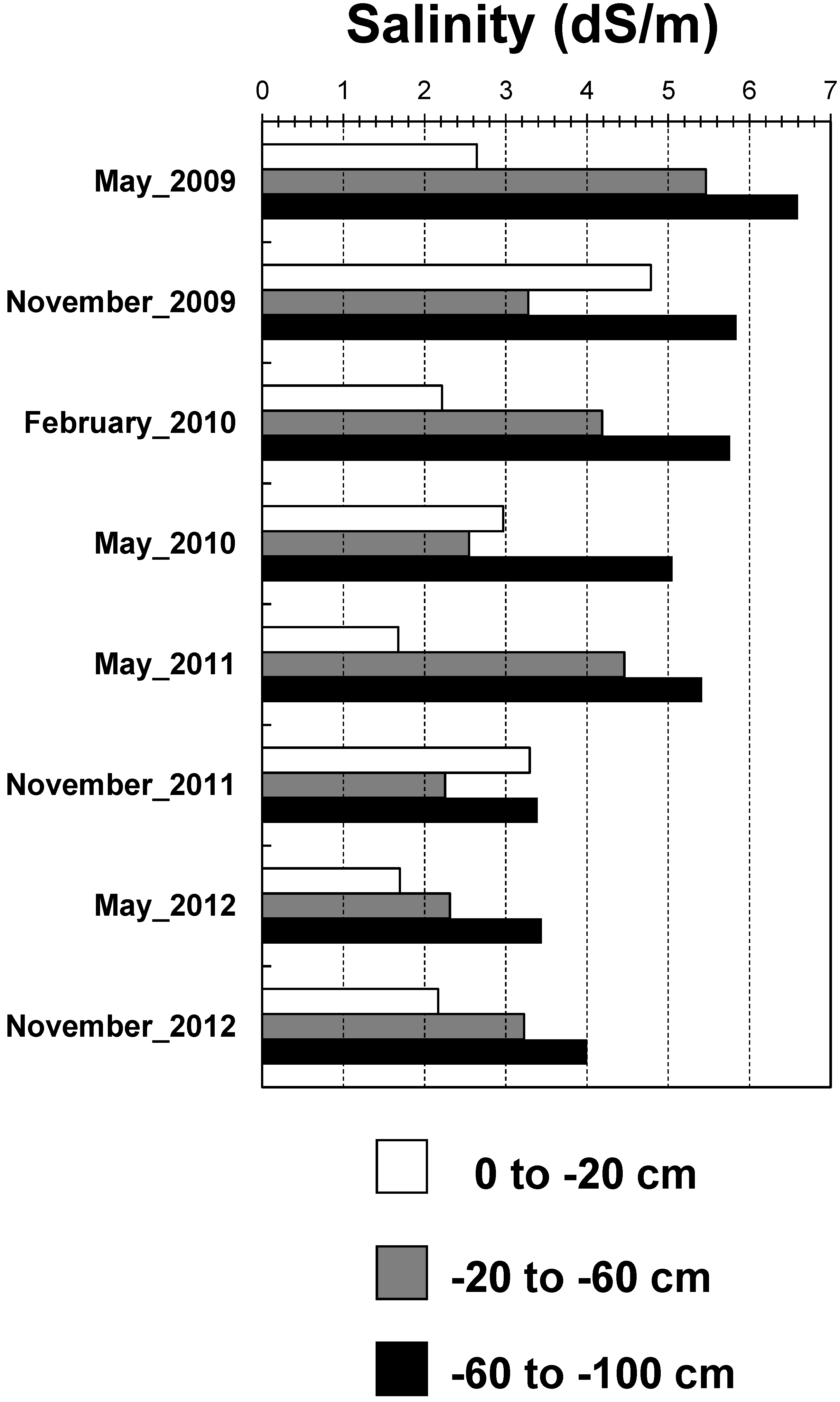
3.1. Temporal and Depth-Dependent Variations in Soil Salinity
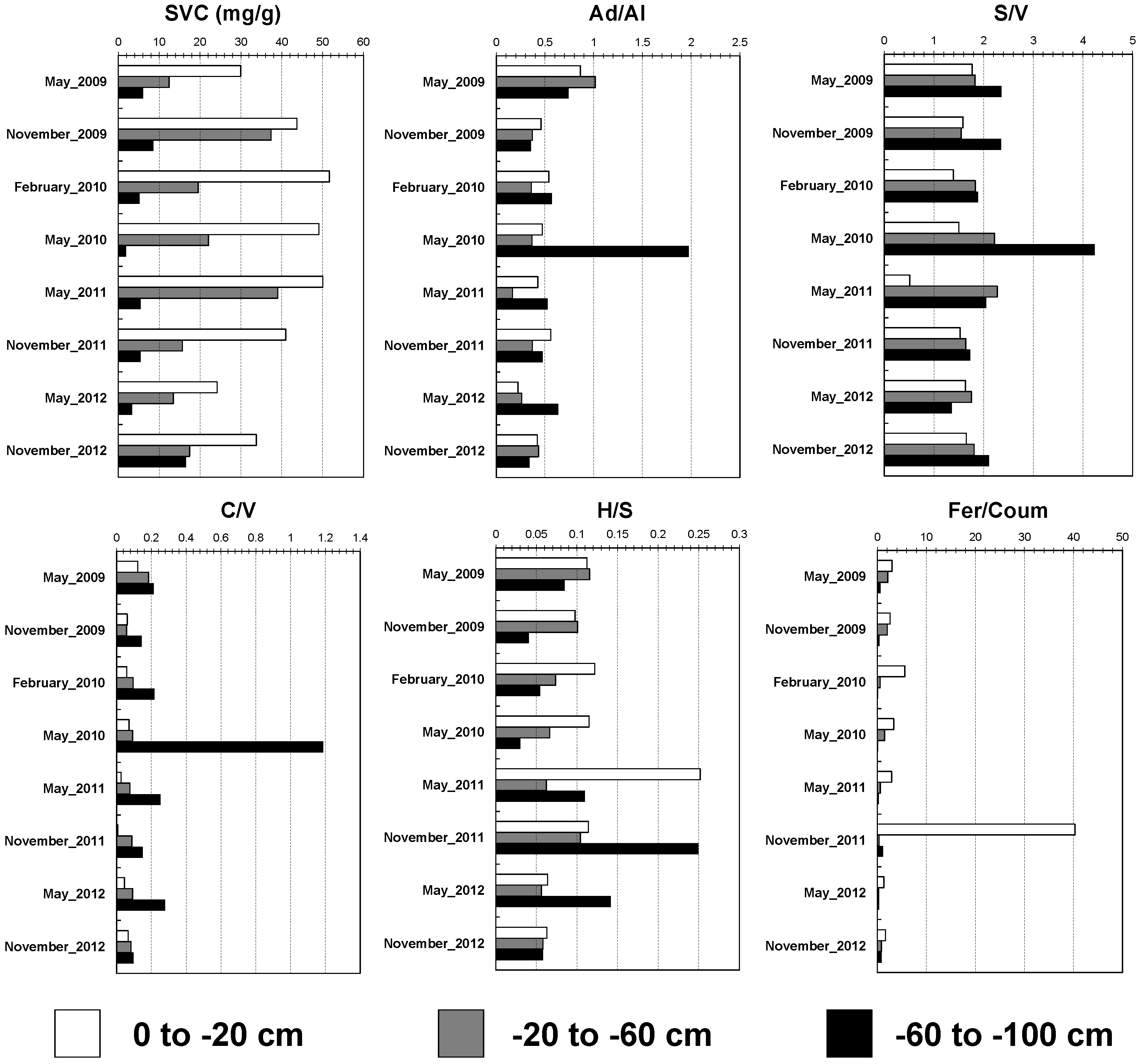
3.2. Depth-Dependent Dynamics of Lignin Biomarkers: Insights into Decomposition and Preservation in Soil Profiles
3.3. Depth and Temporal Variability of Lipid Biomarkers: Insights into Microbial Activity and Organic Matter Dynamics in Soil Profiles
3.4. Influence of Salinity on Lipid and Lignin Biomarkers: Implications for Microbial Activity and Organic Matter Stabilization
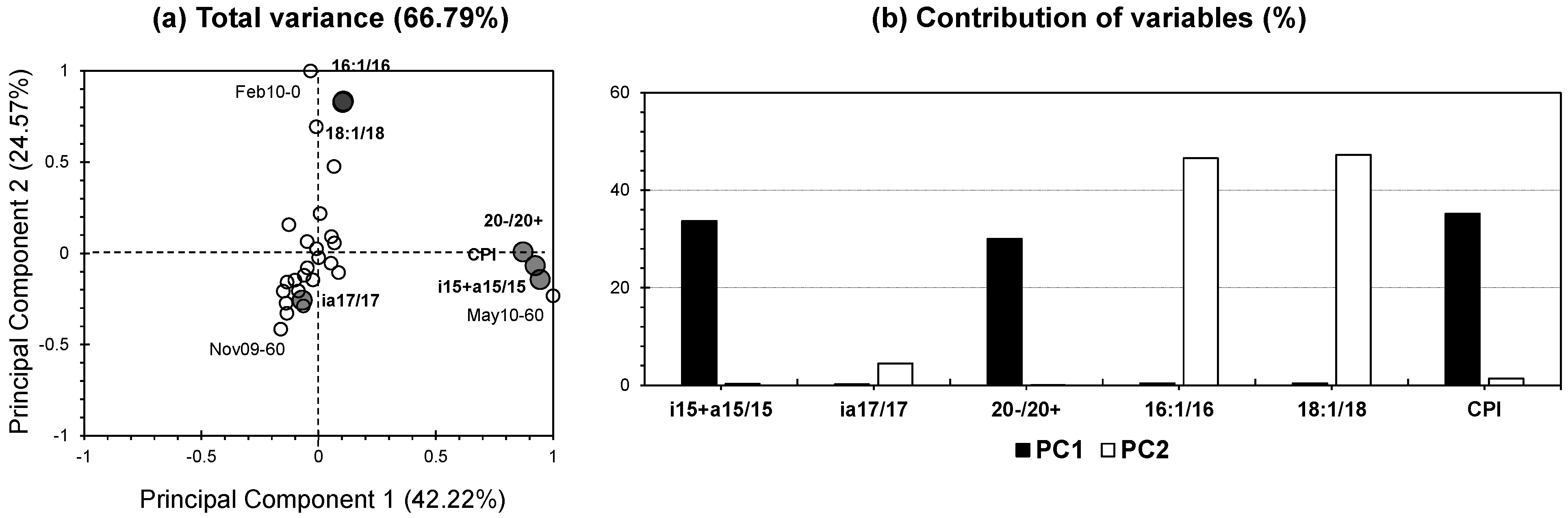
3.5. PCA for Molecular Biomarkers
3.5.1. Linking Lignin and Lipid Dynamics with Soil Biochemical Processes
3.5.2. PCA Analysis of Lignin Biomarkers: Enhanced Insights into Decomposition and Preservation Dynamics
3.5.3. PCA Analysis of Lipid Biomarkers: Insights into Microbial Activity and Lipid Dynamics in Soil Profiles
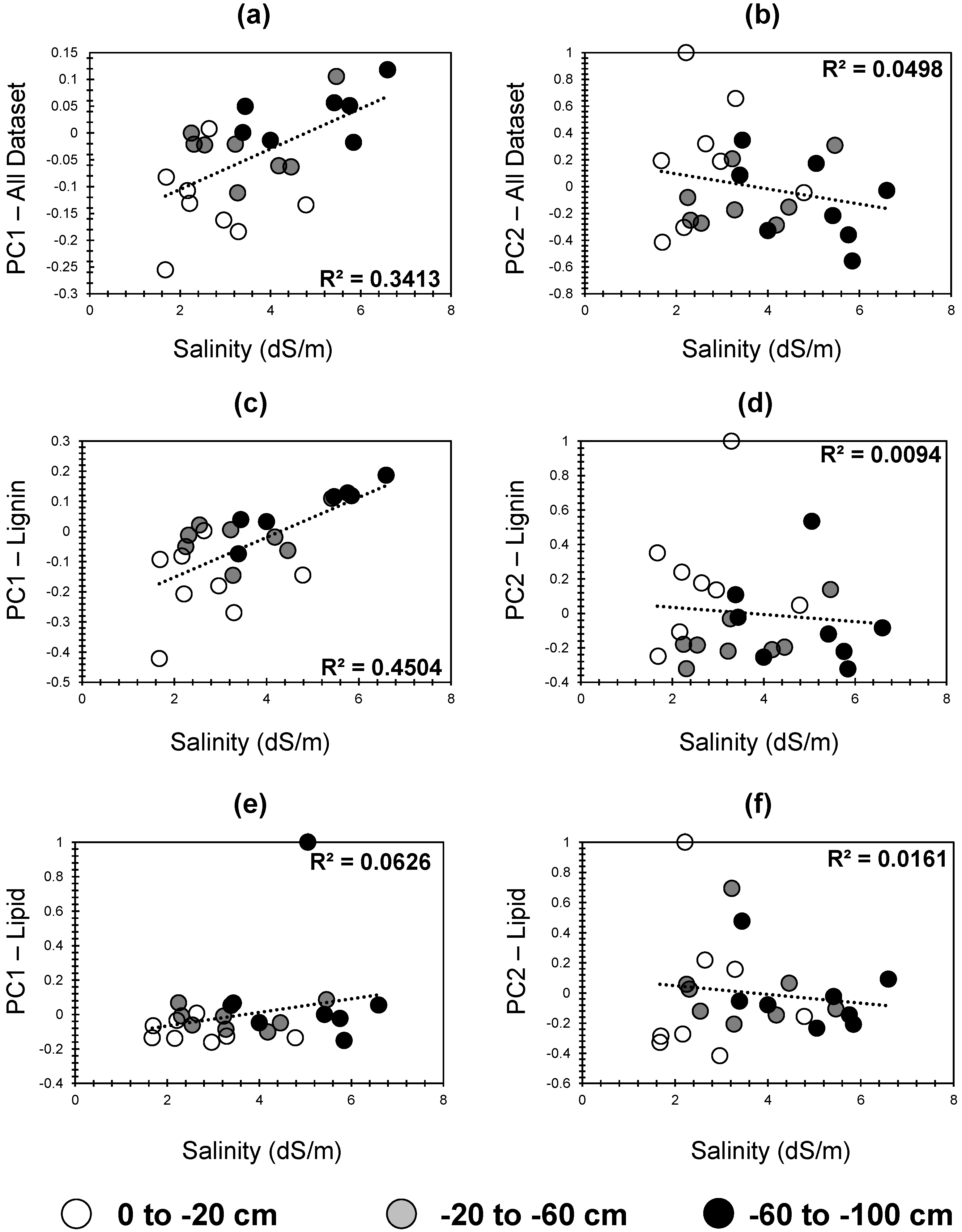
3.5.4. Correlation Between Principal Components and Salinity: Insights into Temporal and Depth-Based Soil Organic Matter Dynamics
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahri, H.; Dignac, M.-F.; Rumpel, C.; Rasse, D.P.; Chenu, C.; Mariotti, A. Lignin Turnover Kinetics in an Agricultural Soil Is Monomer Specific. Soil Biol. Biochem. 2006, 38, 1977–1988. [Google Scholar] [CrossRef]
- Thevenot, M.; Dignac, M.-F.; Rumpel, C. Fate of Lignins in Soils: A Review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar]
- Assembly, G. Sustainable Development Goals. SDGs Transform. Our World 2015, 2030, 6–28. [Google Scholar]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer International Publishing: Cham, Switzerland, 2018; pp. 43–53. ISBN 978-3-319-96189-7. [Google Scholar]
- Collard, F.-X.; Blin, J. A Review on Pyrolysis of Biomass Constituents: Mechanisms and Composition of the Products Obtained from the Conversion of Cellulose, Hemicelluloses and Lignin. Renew. Sustain. Energy Rev. 2014, 38, 594–608. [Google Scholar]
- Corwin, D.L. Climate Change Impacts on Soil Salinity in Agricultural Areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Boulaine, J. Carte Des Sols Des Plaines Du Cheliff Au 1/50.000 e, Feuilles 1 à 5. Inspection générale de l’Agriculture du Gouvernement Général de l’Algérie 1956. [Google Scholar]
- Douaoui, A. Variabilité Spatiale de la Salinité et Sa Relation Avec Certaines Caractéristiques Des Sols de la Plaine du Bas-Chéliff. Ph.D. Thesis, University Center of Tipaza Morsli Abdallah, Tipaza, Algeria, 2005. [Google Scholar]
- Abd El Kader Douaoui, H.N.; Walter, C. Detecting Salinity Hazards within a Semiarid Context by Means of Combining Soil and Remote-Sensing Data. Geoderma 2006, 134, 217–230. [Google Scholar] [CrossRef]
- Younes, K.; Moghrabi, A.; Moghnie, S.; Mouhtady, O.; Murshid, N.; Grasset, L. Assessment of the Efficiency of Chemical and Thermochemical Depolymerization Methods for Lignin Valorization: Principal Component Analysis (PCA) Approach. Polymers 2022, 14, 194. [Google Scholar] [CrossRef]
- Schellekens, J.; Bindler, R.; Martínez-Cortizas, A.; McClymont, E.L.; Abbott, G.D.; Biester, H.; Pontevedra-Pombal, X.; Buurman, P. Preferential Degradation of Polyphenols from Sphagnum–4-Isopropenylphenol as a Proxy for Past Hydrological Conditions in Sphagnum-Dominated Peat. Geochim. Cosmochim. Acta 2015, 150, 74–89. [Google Scholar]
- Bahreininejad, B.; Allahdadi, M. Effects of Saline Irrigated Water on Forage Quality of Globe Artichoke (Cynara cardunculus var. scolymus L.). Iran Agric. Res. 2020, 39, 59–66. [Google Scholar]
- Hardie, M.; Doyle, R. Measuring Soil Salinity. In Plant Salt Tolerance; Shabala, S., Cuin, T.A., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 913, pp. 415–425. ISBN 978-1-61779-985-3. [Google Scholar]
- Wang, J.J.; Provin, T.; Zhang, H. Measurement of Soil Salinity and Sodicity. In Soil Test Methods from the Southeastern United States; University of Kentucky: Lexington, KY, USA; Clemson University: Clemson, SC, USA, 2014; p. 185. Available online: https://aesl.ces.uga.edu/Sera6/PUB/Methodsmanualfinalsera6.Pdf#page=193 (accessed on 26 February 2025).
- Ertel, J.R.; Hedges, J.I. The Lignin Component of Humic Substances: Distribution among Soil and Sedimentary Humic, Fulvic, and Base-Insoluble Fractions. Geochim. Cosmochim. Acta 1984, 48, 2065–2074. [Google Scholar]
- Jørgensen, P.-E.; Eriksen, T.; Jensen, B.K. Estimation of Viable Biomass in Wastewater and Activated Sludge by Determination of ATP, Oxygen Utilization Rate and FDA Hydrolysis. Water Res. 1992, 26, 1495–1501. [Google Scholar]
- Hedges, J.I.; Mann, D.C. The Characterization of Plant Tissues by Their Lignin Oxidation Products. Geochim. Cosmochim. Acta 1979, 43, 1803–1807. [Google Scholar] [CrossRef]
- Otto, A.; Simpson, M.J. Evaluation of CuO Oxidation Parameters for Determining the Source and Stage of Lignin Degradation in Soil. Biogeochemistry 2006, 80, 121–142. [Google Scholar] [CrossRef]
- Collard, M.; Teychené, B.; Lemée, L. Comparison of Three Different Wastewater Sludge and Their Respective Drying Processes: Solar, Thermal and Reed Beds–Impact on Organic Matter Characteristics. J. Environ. Manag. 2017, 203, 760–767. [Google Scholar]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of Salt-induced Land Degradation and Restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Massoud, F.I. Salt Affected Soils at a Global Scale and Concepts for Control; FAO: Rome, Italy, 1981. [Google Scholar]
- Bourg, I.C.; Ajo-Franklin, J.B. Clay, Water, and Salt: Controls on the Permeability of Fine-Grained Sedimentary Rocks. Acc. Chem. Res. 2017, 50, 2067–2074. [Google Scholar] [CrossRef]
- Corwin, D.L.; Lesch, S.M.; Oster, J.D.; Kaffka, S.R. Short-Term Sustainability of Drainage Water Reuse: Spatio-Temporal Impacts on Soil Chemical Properties. J. Environ. Qual. 2008, 37, S-8-S-24. [Google Scholar] [CrossRef]
- Wichelns, D.; Qadir, M. Achieving Sustainable Irrigation Requires Effective Management of Salts, Soil Salinity, and Shallow Groundwater. Agric. Water Manag. 2015, 157, 31–38. [Google Scholar]
- Younes, K.; Laduranty, J.; Descostes, M.; Grasset, L. Molecular Biomarkers Study of an Ombrotrophic Peatland Impacted by an Anthropogenic Clay Deposit. Org. Geochem. 2017, 105, 20–32. [Google Scholar] [CrossRef]
- Kaal, J.; Pérez-Rodríguez, M.; Biester, H. Molecular Probing of DOM Indicates a Key Role of Spruce-Derived Lignin in the DOM and Metal Cycles of a Headwater Catchment: Can Spruce Forest Dieback Exacerbate Future Trends in the Browning of Central European Surface Waters? Environ. Sci. Technol. 2022, 56, 2747–2759. [Google Scholar] [CrossRef]
- Grünewald, G.; Kaiser, K.; Jahn, R.; Guggenberger, G. Organic Matter Stabilization in Young Calcareous Soils as Revealed by Density Fractionation and Analysis of Lignin-Derived Constituents. Org. Geochem. 2006, 37, 1573–1589. [Google Scholar]
- Gordon, E.S.; Goñi, M.A. Sources and Distribution of Terrigenous Organic Matter Delivered by the Atchafalaya River to Sediments in the Northern Gulf of Mexico. Geochim. Cosmochim. Acta 2003, 67, 2359–2375. [Google Scholar]
- Hedges, J.I.; Clark, W.A.; Come, G.L. Organic Matter Sources to the Water Column and Surficial Sediments of a Marine Bay: Organic Matter Sources. Limnol. Oceanogr. 1988, 33, 1116–1136. [Google Scholar] [CrossRef]
- Hedges, J.I.; Blanchette, R.A.; Weliky, K.; Devol, A.H. Effects of Fungal Degradation on the CuO Oxidation Products of Lignin: A Controlled Laboratory Study. Geochim. Cosmochim. Acta 1988, 52, 2717–2726. [Google Scholar]
- Jeffries, T.W. Biodegradation of Lignin-Carbohydrate Complexes. In Physiology of Biodegradative Microorganisms; Ratledge, C., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 1991; pp. 163–176. ISBN 978-94-010-5527-7. [Google Scholar]
- Skyba, O.; Douglas, C.J.; Mansfield, S.D. Syringyl-Rich Lignin Renders Poplars More Resistant to Degradation by Wood Decay Fungi. Appl. Environ. Microbiol. 2013, 79, 2560–2571. [Google Scholar] [CrossRef]
- Chang, B.V.; Chang, S.W.; Yuan, S.Y. Anaerobic Degradation of Polycyclic Aromatic Hydrocarbons in Sludge. Adv. Environ. Res. 2003, 7, 623–628. [Google Scholar]
- Orem, W.H.; Colman, S.M.; Lerch, H.E. Lignin Phenols in Sediments of Lake Baikal, Siberia: Application to Paleoenvironmental Studies. Org. Geochem. 1997, 27, 153–172. [Google Scholar]
- Tortosa-Díaz, L.; Saura-Martínez, J.; Taboada-Rodríguez, A.; Martínez-Hernández, G.B.; López-Gómez, A.; Marín-Iniesta, F. Influence of Industrial Processing of Artichoke and By-Products on The Bioactive and Nutritional Compounds. Food Eng. Rev. 2025, 1–24. [Google Scholar] [CrossRef]
- Elouaqoudi, F.Z.; El Fels, L.; Amir, S.; Merlina, G.; Meddich, A.; Lemee, L.; Ambles, A.; Hafidi, M. Lipid Signature of the Microbial Community Structure during Composting of Date Palm Waste Alone or Mixed with Couch Grass Clippings. Int. Biodeterior. Biodegrad. 2015, 97, 75–84. [Google Scholar]
- Kögel, I.; Hempfling, R.; Zech, W.; Hatcher, P.G.; Schulten, H.-R. Chemical Composition of the Organic Matter in Forest Soils: 1. Forest Litter. Soil Sci. 1988, 146, 124–136. [Google Scholar]
- Kögel-Knabner, I. The Macromolecular Organic Composition of Plant and Microbial Residues as Inputs to Soil Organic Matter: Fourteen Years On. Soil Biol. Biochem. 2017, 105, A3–A8. [Google Scholar] [CrossRef]
- Eckmeier, E.; Wiesenberg, G.L. Short-Chain n-Alkanes (C16–20) in Ancient Soil Are Useful Molecular Markers for Prehistoric Biomass Burning. J. Archaeol. Sci. 2009, 36, 1590–1596. [Google Scholar] [CrossRef]
- Wiesenberg, G.L.; Lehndorff, E.; Schwark, L. Thermal Degradation of Rye and Maize Straw: Lipid Pattern Changes as a Function of Temperature. Org. Geochem. 2009, 40, 167–174. [Google Scholar] [CrossRef]
- Bhattarai, S.P.; Su, N.; Midmore, D.J. Oxygation Unlocks Yield Potentials of Crops in Oxygen-Limited Soil Environments. Adv. Agron. 2005, 88, 313–377. [Google Scholar]
- Balesdent, J.; Chenu, C.; Balabane, M. Relationship of Soil Organic Matter Dynamics to Physical Protection and Tillage. Soil Tillage Res. 2000, 53, 215–230. [Google Scholar] [CrossRef]
- Estournel-Pelardy, C.; El-Mufleh Al Husseini, A.; Doskočil, L.; Grasset, L. A Two-Step Thermochemolysis for Soil Organic Matter Analysis. Application to Lipid-Free Organic Fraction and Humic Substances from an Ombrotrophic Peatland. J. Anal. Appl. Pyrolysis 2013, 104, 103–110. [Google Scholar] [CrossRef]
- Akkermans, A.D.L.; Van Elsas, J.D.; De Bruijn, F.J. (Eds.) Molecular Microbial Ecology Manual; Springer Netherlands: Dordrecht, The Netherlands, 1996; ISBN 978-94-011-7660-6. [Google Scholar]
- Tremblay, L.; Benner, R. Microbial Contributions to N-Immobilization and Organic Matter Preservation in Decaying Plant Detritus. Geochim. Cosmochim. Acta 2006, 70, 133–146. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Robert, M.; Chenu, C. Interactions between Soil Minerals and Microorganisms. In Soil Biochemistry; CRC Press: Boca Raton, FL, USA, 2021; pp. 307–404. [Google Scholar]
- Quinn, P.J.; Joo, F.; Vigh, L. The Role of Unsaturated Lipids in Membrane Structure and Stability. Prog. Biophys. Mol. Biol. 1989, 53, 71–103. [Google Scholar] [CrossRef]
- Younes, K.; Grasset, L. Carbohydrates as Proxies in Ombrotrophic Peatland: DFRC Molecular Method Coupled with PCA. Chem. Geol. 2022, 606, 120994. [Google Scholar] [CrossRef]
- Harji, R.R.; Bhosle, N.B.; Garg, A.; Sawant, S.S.; Venkat, K. Sources of Organic Matter and Microbial Community Structure in the Sediments of the Visakhapatnam Harbour, East Coast of India. Chem. Geol. 2010, 276, 309–317. [Google Scholar]
- El Fels, L.; Lemee, L.; Ambles, A.; Hafidi, M. Identification and Biotransformation of Aliphatic Hydrocarbons during Co-Composting of Sewage Sludge-Date Palm Waste Using Pyrolysis-GC/MS Technique. Environ. Sci. Pollut. Res. 2016, 23, 16857–16864. [Google Scholar]
- Srivastava, M.; Mishra, A.K. Comparative Analysis of Paddy Soil Denitrifying Bacteria with Soil Phospholipid Fatty Acid Profile. Geomicrobiol. J. 2021, 38, 404–414. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Geographical Coordinates | Surface | Agricultural Acitvity | Overall Salinity EC (dS/m) | Soil Texture | ||
|---|---|---|---|---|---|---|---|
| Type | Periods | ||||||
| X | Y | ||||||
| El Hmadna | 296,645 | 3,979,661 | 2 ha | Artichoke and Cereals | 2009–2012 | 4.17 | Clay |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akkacha, A.; Douaoui, A.; Younes, K.; El Sawda, C.; Alsyouri, H.; El-Zahab, S.; Grasset, L. Investigating the Impact of Salinity on Soil Organic Matter Dynamics Using Molecular Biomarkers and Principal Component Analysis. Sustainability 2025, 17, 2940. https://doi.org/10.3390/su17072940
Akkacha A, Douaoui A, Younes K, El Sawda C, Alsyouri H, El-Zahab S, Grasset L. Investigating the Impact of Salinity on Soil Organic Matter Dynamics Using Molecular Biomarkers and Principal Component Analysis. Sustainability. 2025; 17(7):2940. https://doi.org/10.3390/su17072940
Chicago/Turabian StyleAkkacha, Abderrhamen, Abdelkader Douaoui, Khaled Younes, Christina El Sawda, Hatem Alsyouri, Samer El-Zahab, and Laurent Grasset. 2025. "Investigating the Impact of Salinity on Soil Organic Matter Dynamics Using Molecular Biomarkers and Principal Component Analysis" Sustainability 17, no. 7: 2940. https://doi.org/10.3390/su17072940
APA StyleAkkacha, A., Douaoui, A., Younes, K., El Sawda, C., Alsyouri, H., El-Zahab, S., & Grasset, L. (2025). Investigating the Impact of Salinity on Soil Organic Matter Dynamics Using Molecular Biomarkers and Principal Component Analysis. Sustainability, 17(7), 2940. https://doi.org/10.3390/su17072940