
Tracking the Expansion of Sonneratia apetala and Its Impact on Local Mangroves Using Time-Series Remote Sensing Data

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
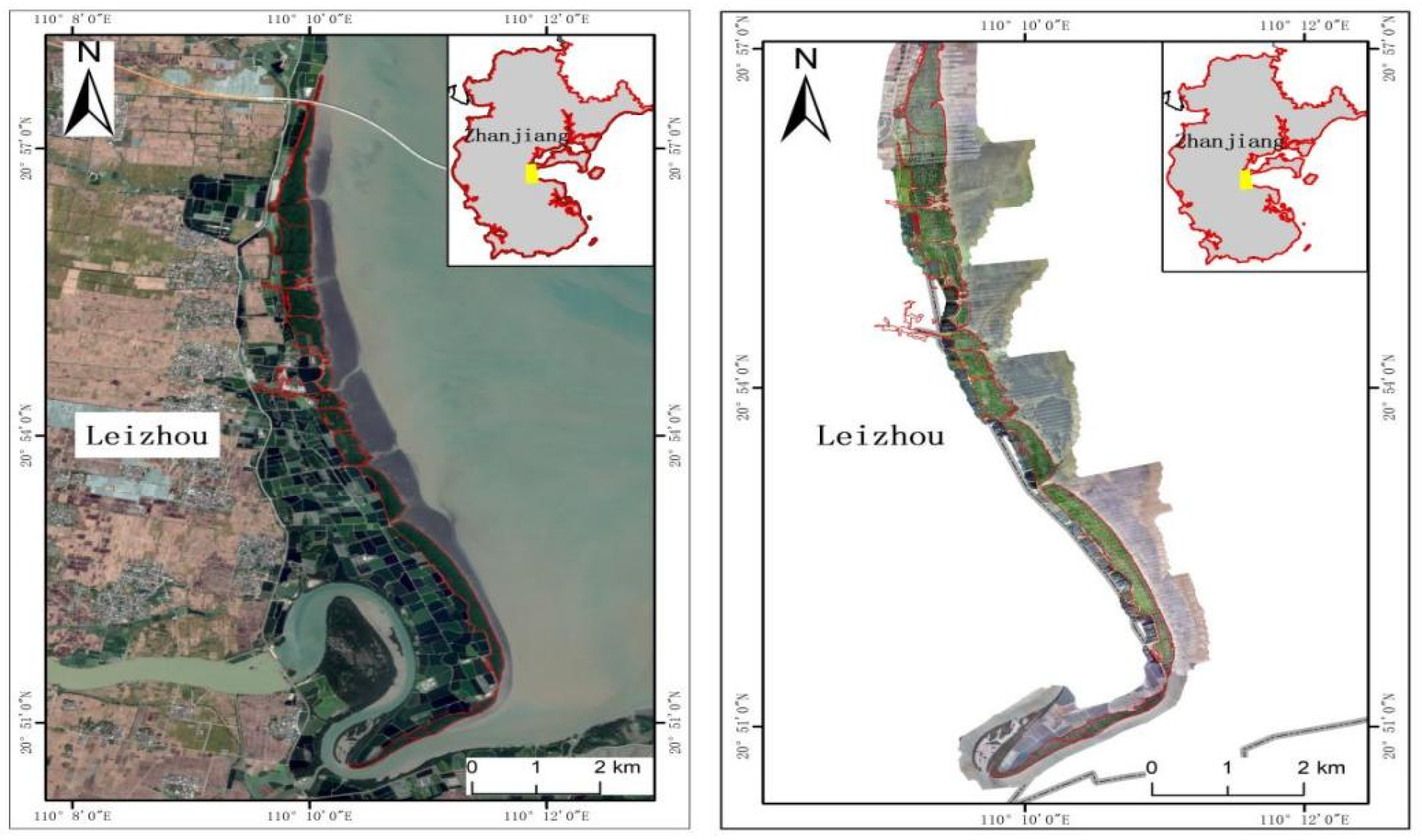
2.1. Study Area
2.2. Data Sources and Preprocessing
2.3. Water Mask
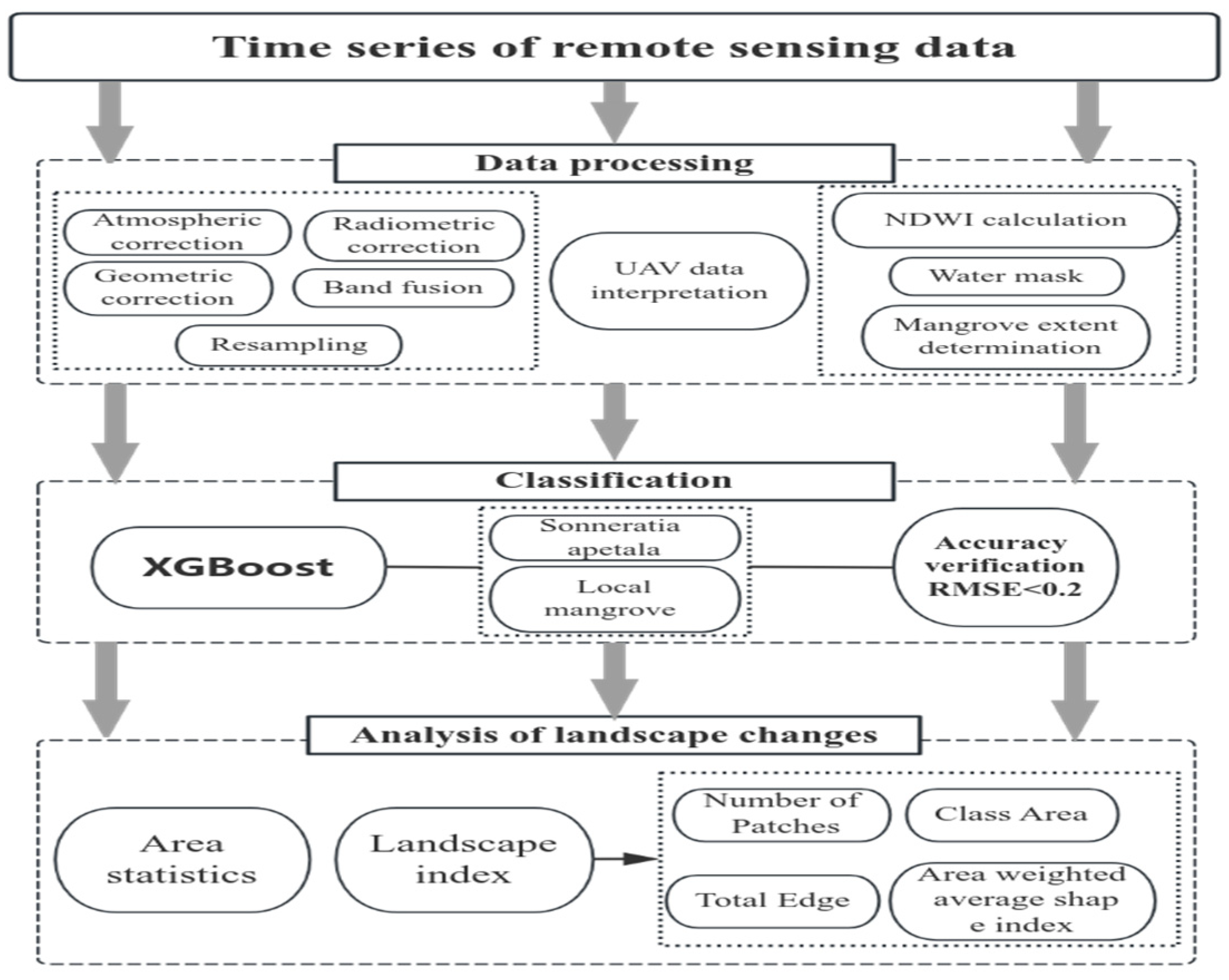
2.4. Methods
2.4.1. Classification of Mangrove Species
Mangrove Classification Rules
Classification
2.4.2. Accuracy Verification
2.4.3. Analysis of Landscape Pattern Change
3. Results and Discussion
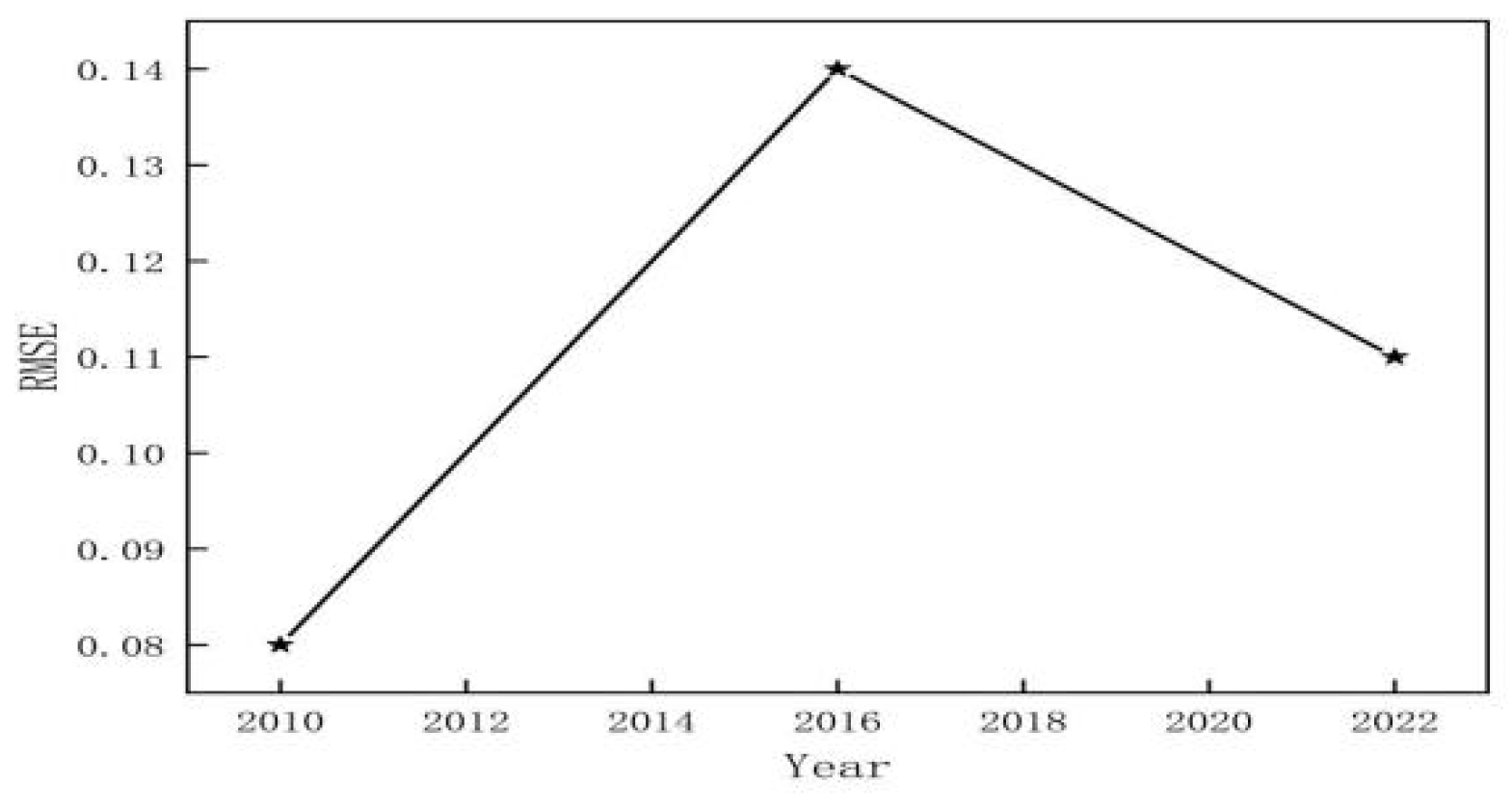
3.1. Classification Accuracy
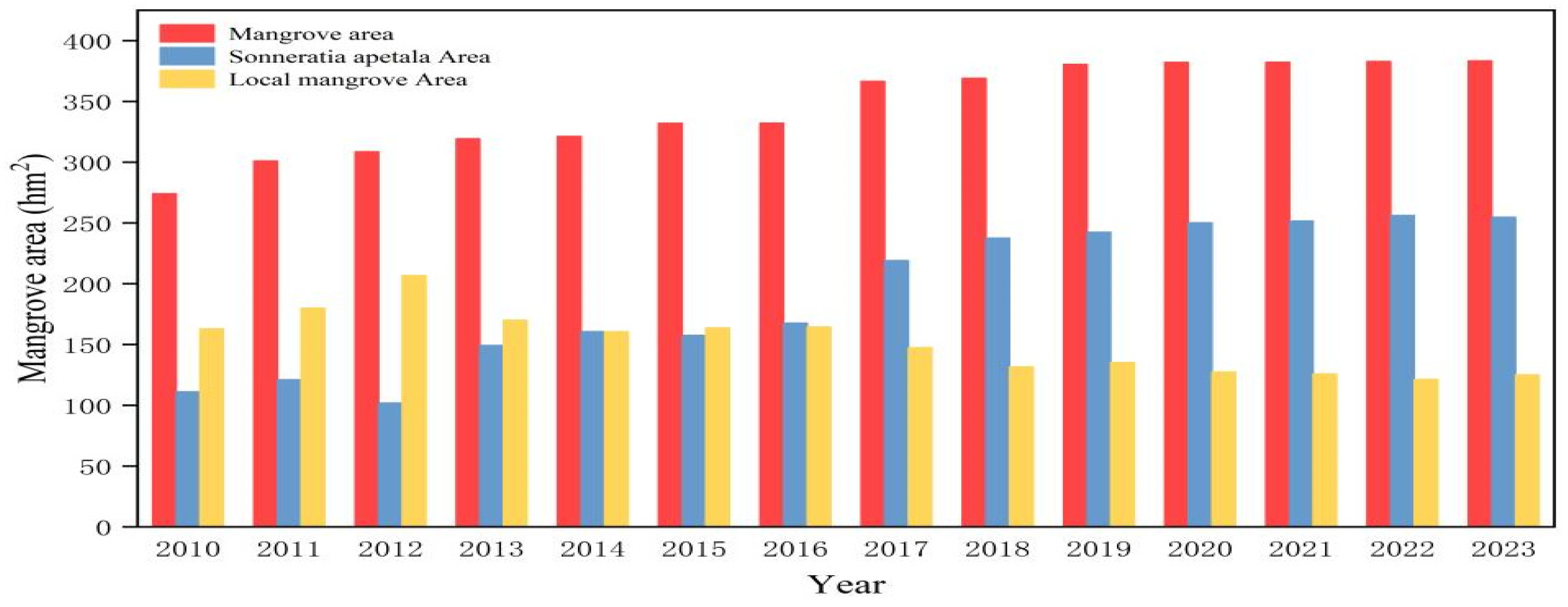
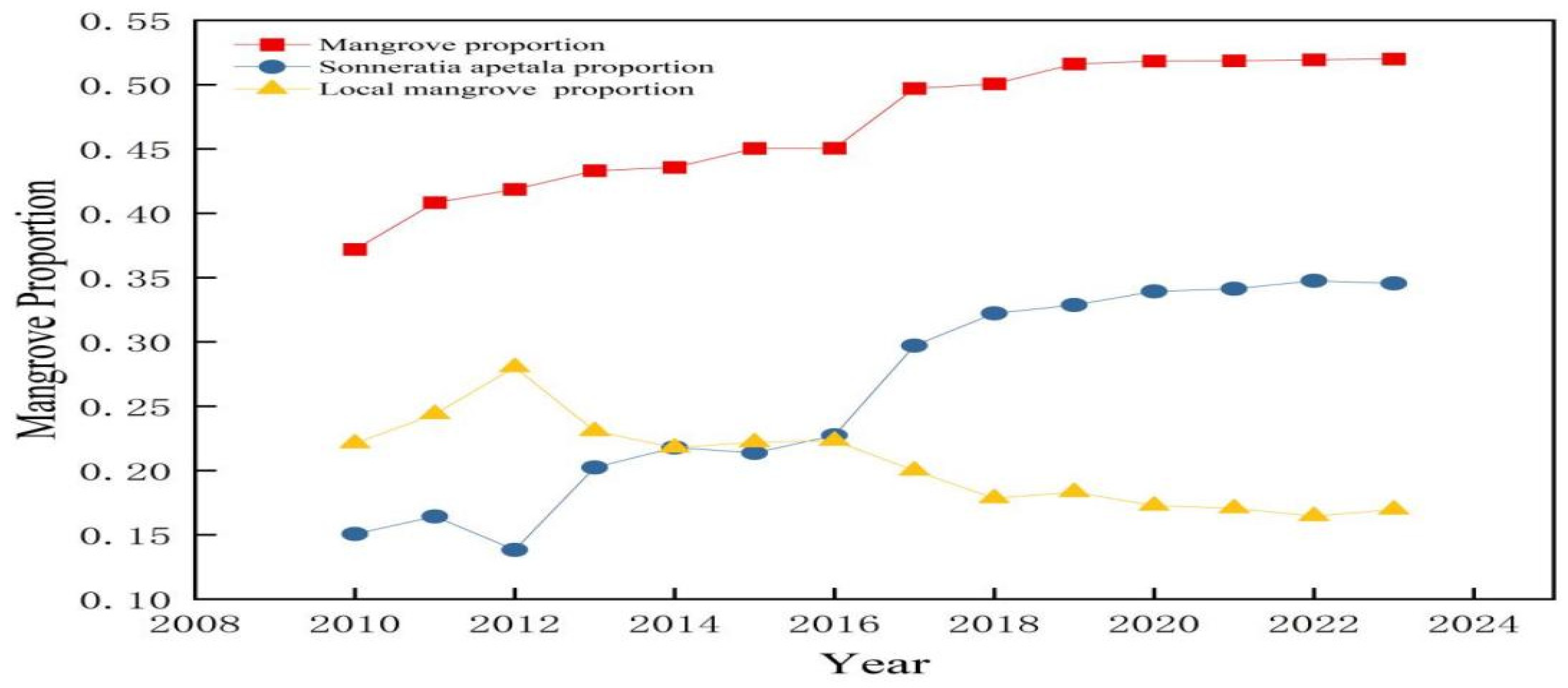
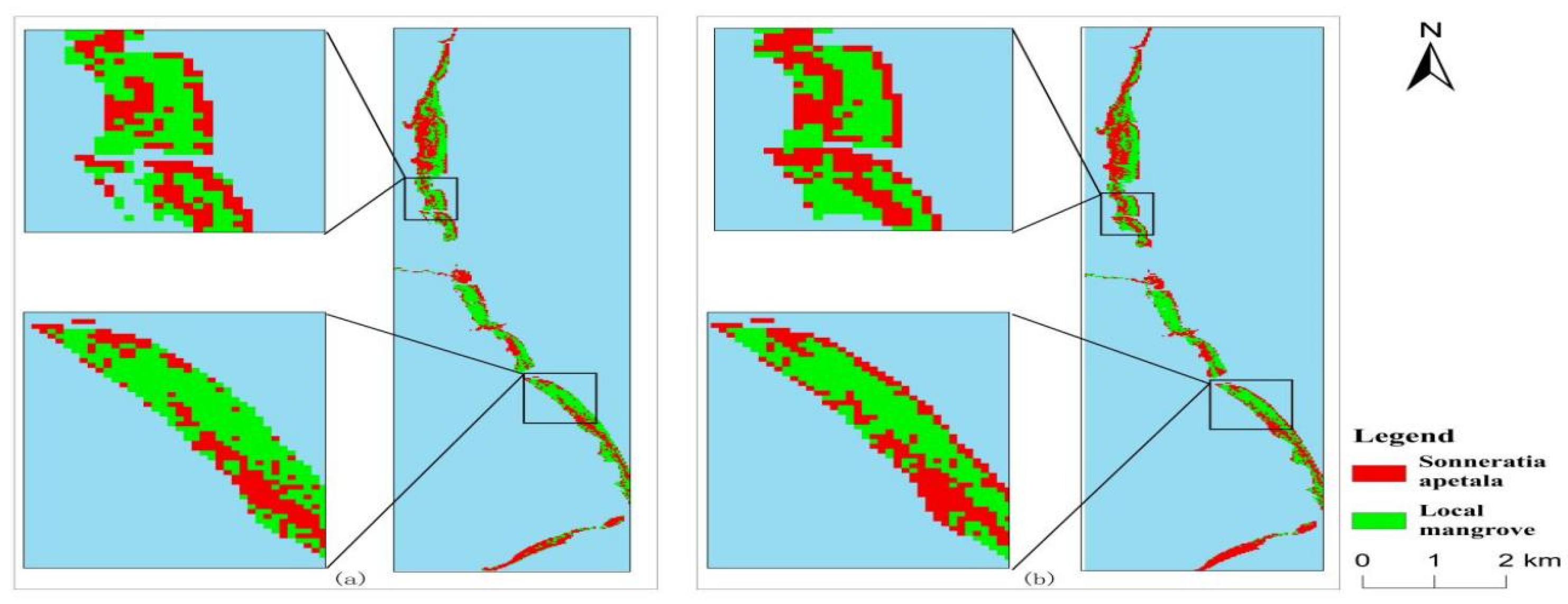
3.2. Changes in Mangrove Forests from 2010 to 2023
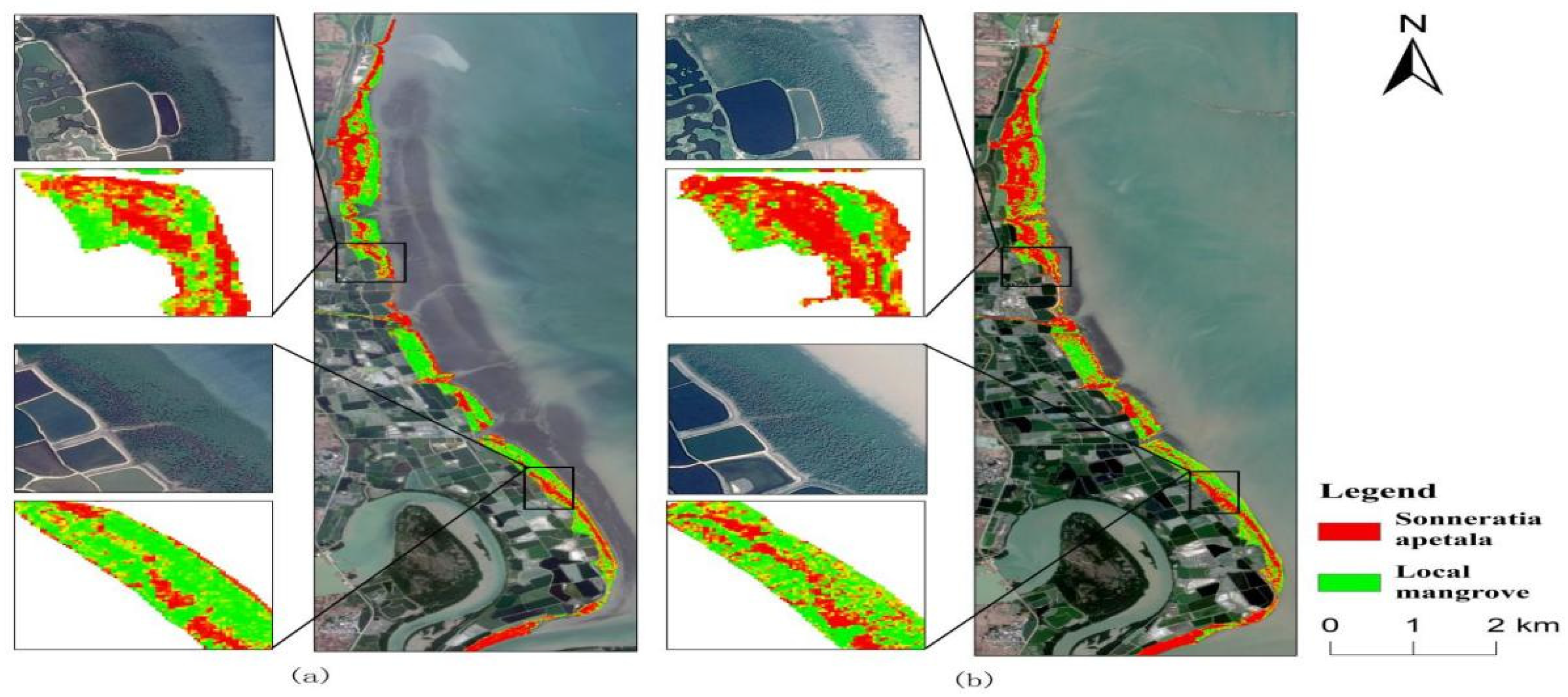
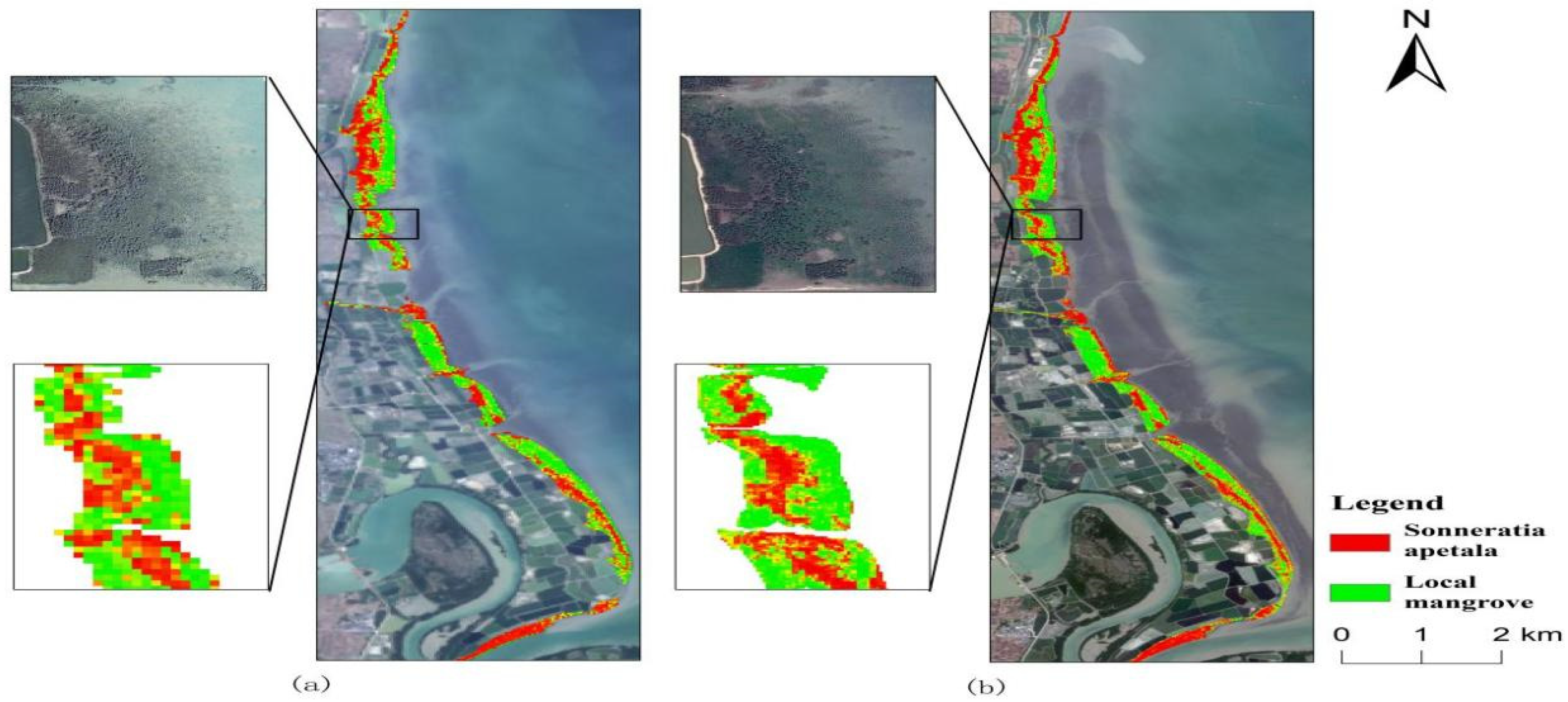
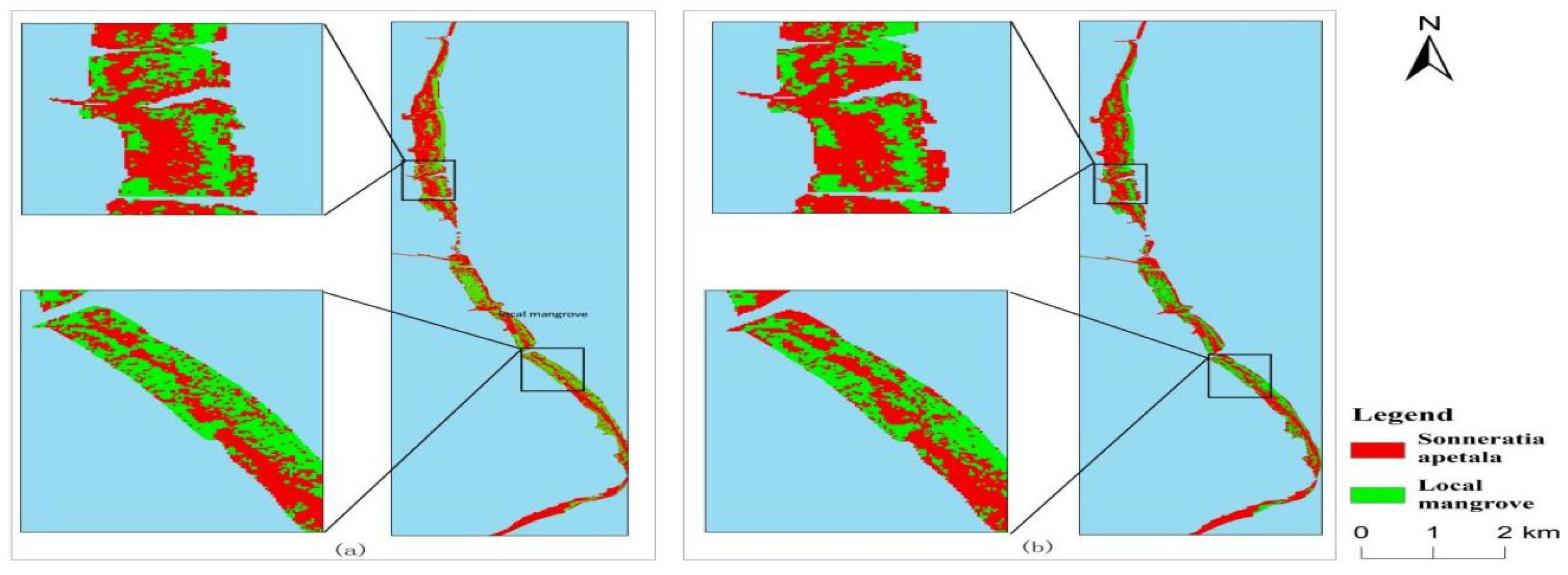
3.3. Spatial and Temporal Changes of Sonneratia apetala
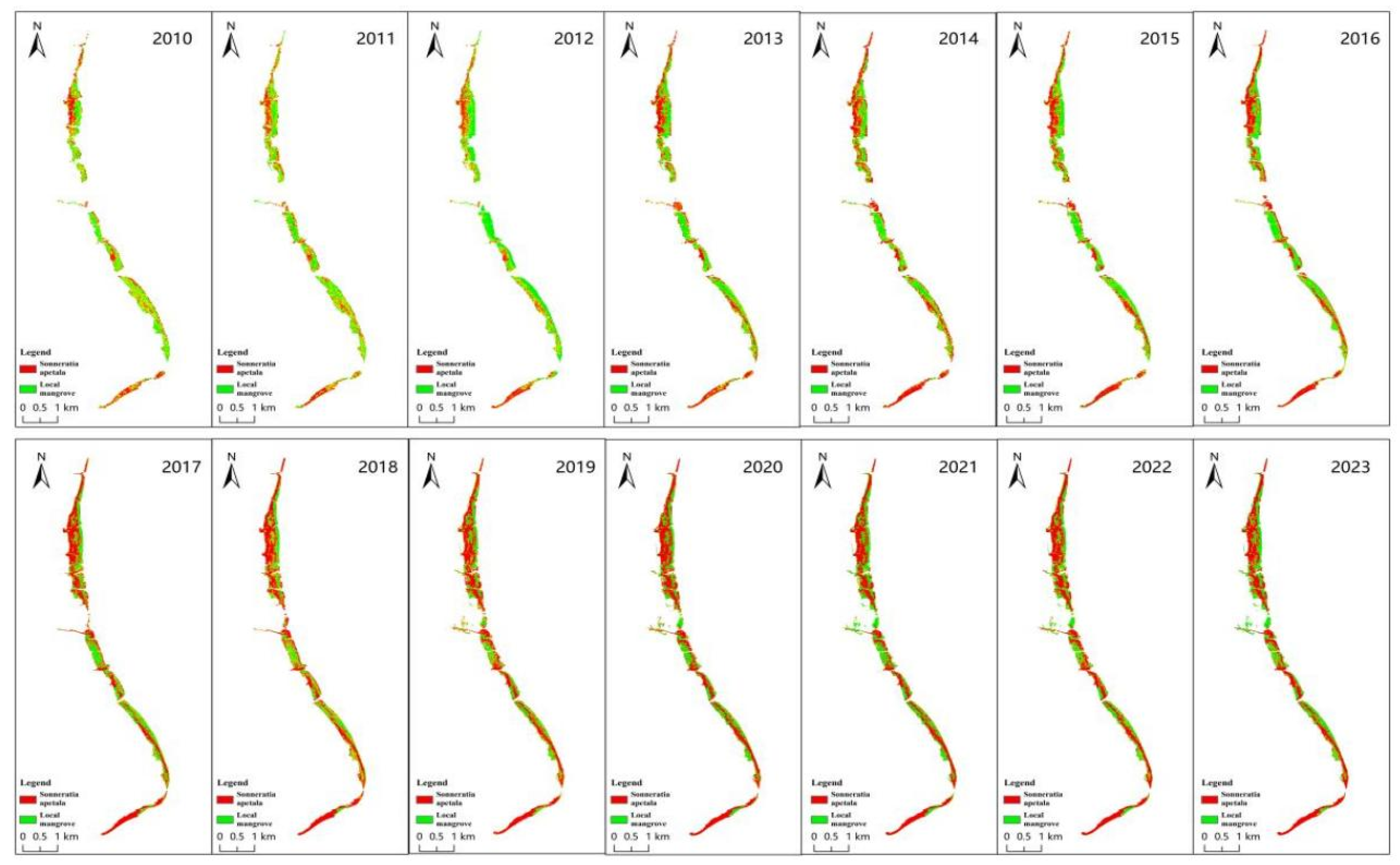
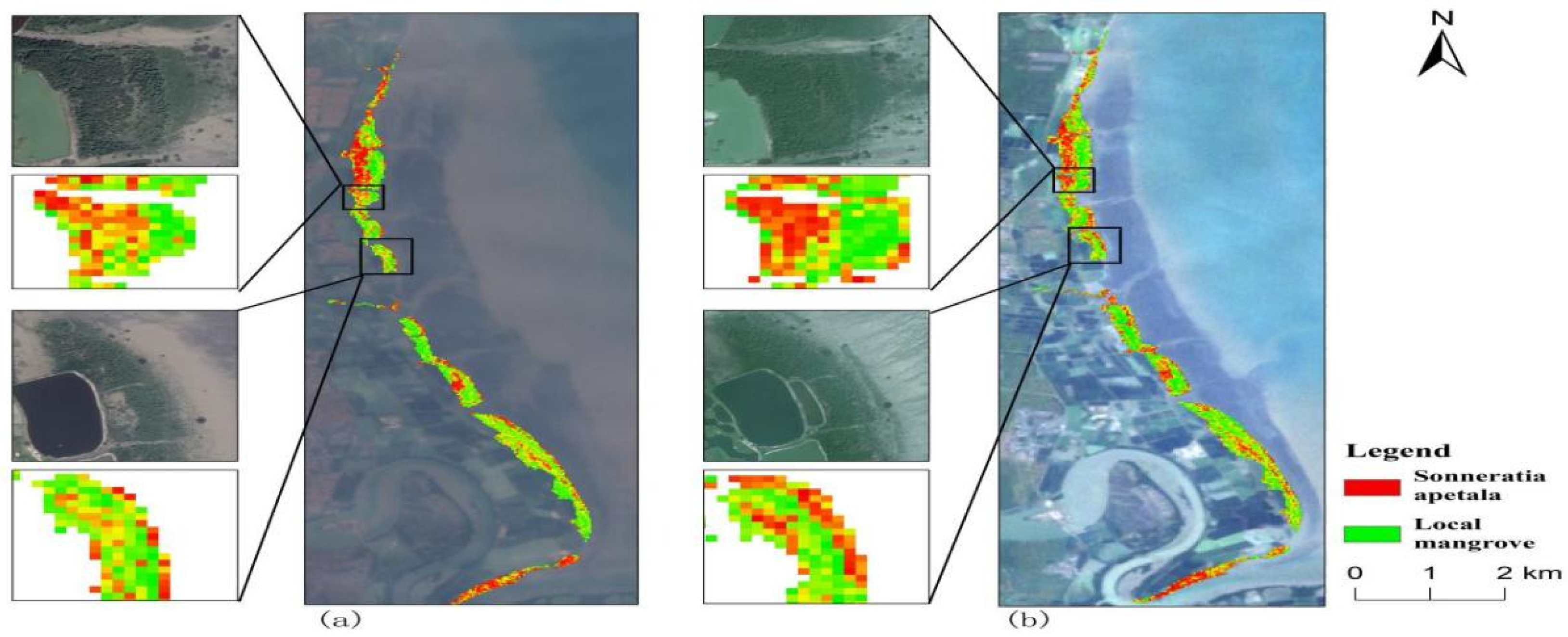
3.3.1. Spatial Variation of Mangroves from 2010 to 2023
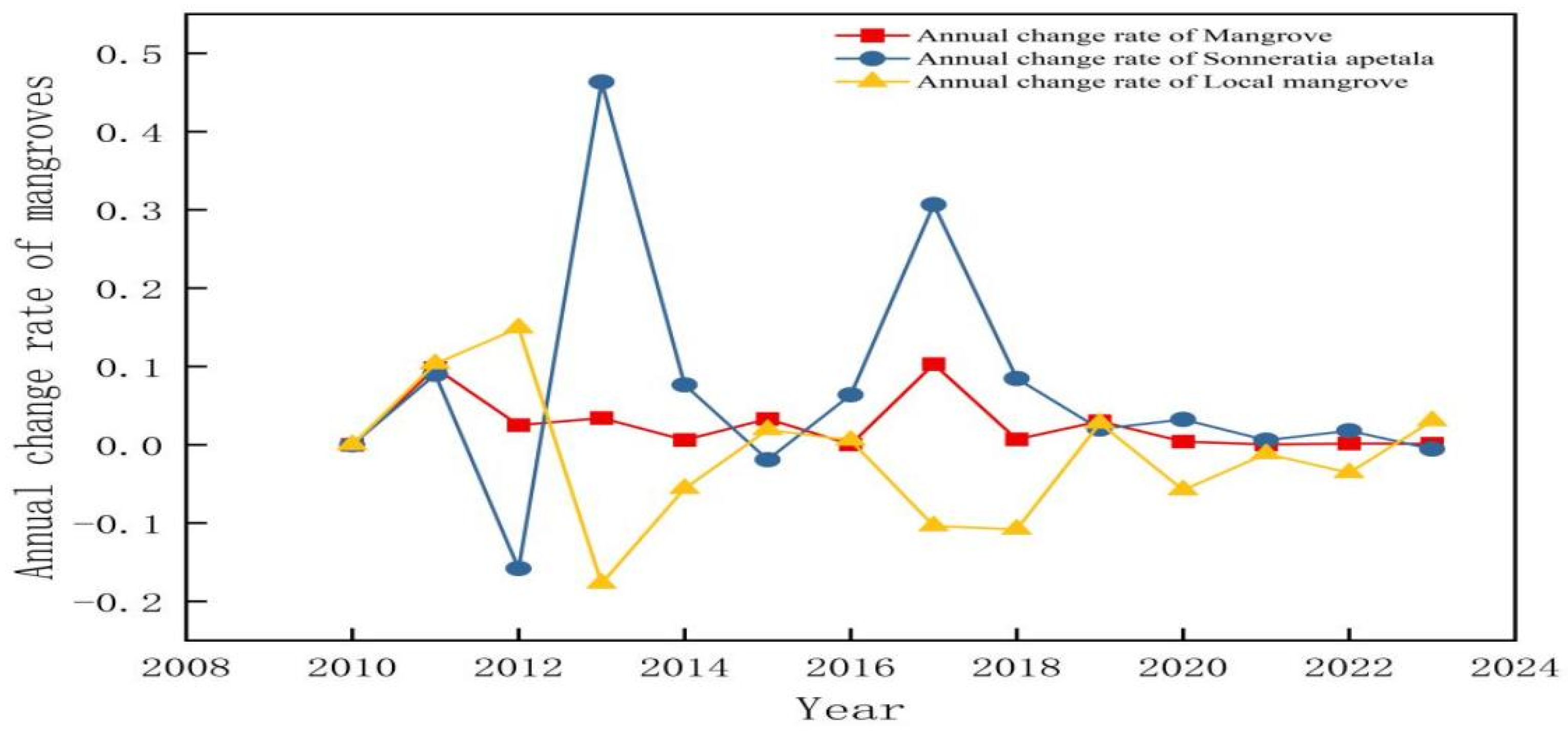
3.3.2. Temporal Changes of Sonneratia apetala from 2010 to 2023
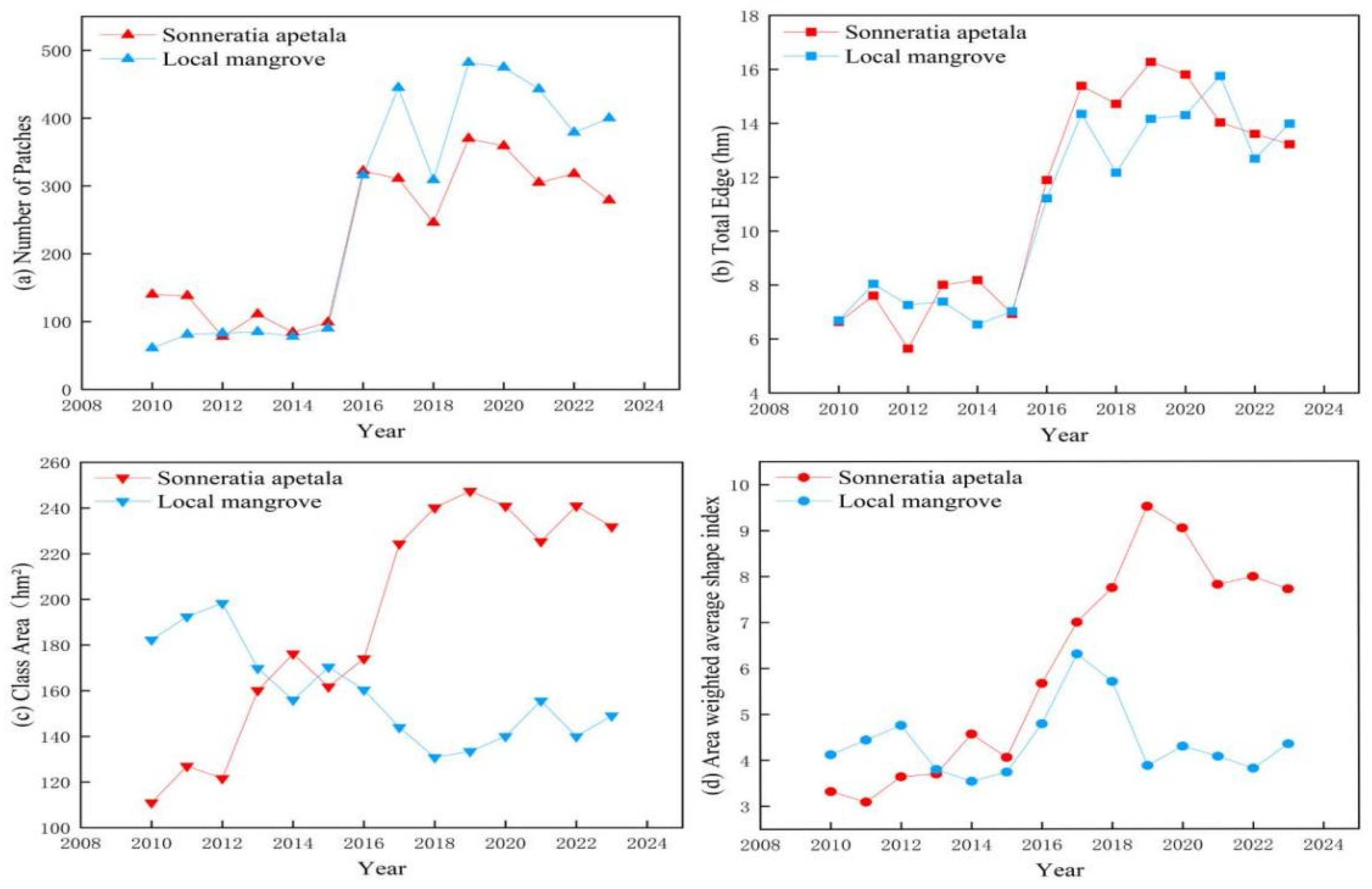
3.4. Changes in Landscape Pattern
4. Conclusions
- (1)
- Expansion of S. apetala and Decline of Local Mangroves: The area of mangrove forests increased, primarily driven by the expansion of S. apetala. While local mangroves showed a slow decline, S. apetala’s growth is largely attributed to its strong competitive ability and extensive planting efforts. This invasive species has significantly altered the distribution and coverage of local mangrove species.
- (2)
- Spatial and Temporal Trends: S. apetala has expanded along mudflats and the edges of mangrove forests, reducing the area of native mangroves. It is expected to continue spreading and may dominate the region within the next two to three decades. Thus, urgent measures to control S. apetala’s growth and protect local mangrove species are needed to maintain biodiversity and ecosystem balance.
- (3)
- Increased Fragmentation: Over the 14-year study period, S. apetala patches increased in number and complexity due to early-stage planting and subsequent spread. The fragmentation of S. apetala habitats, influenced by both natural and human factors, poses a high invasion risk, especially in areas with sparse local mangrove coverage.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lagomasino, D.; Price, R.M.; Whitman, D.; Campbell, P.K.; Melesse, A. Estimating major ion and nutrient concentrations in mangrove estuaries in Everglades National Park using leaf and satellite reflectance. Remote Sens. Environ. 2014, 154, 202–218. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Purbopuspito, J.; Kauffman, J.B.; Warren, M.W.; Sasmito, S.D.; Donato, D.C.; Manuri, S.; Krisnawati, H.; Taberima, S.; Kurnianto, S. The potential of Indonesian mangrove forests for global climate change mitigation. Nat. Clim. Change 2015, 5, 1089–1092. [Google Scholar] [CrossRef]
- He, B.Y.; Fan, H.Q.; Wang, M.; Lai, T.H.; Wang, W.Q. Species diversity in mangrove wetlands of China and its causation analyses. Acta Ecol. Sin. 2007, 27, 4859–4870. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Elfes, C.T.; Sanciangco, J.C.; Pippard, H.; Carpenter, K.E. Conservation status of marine biodiversity in oceania: An analysis of marine species on the iucn red list of threatened species. J. Mar. Biol. 2010, 2011, 247030. [Google Scholar] [CrossRef]
- Proisy, C.; Couteron, P.; Fromard, F. Predicting and mapping mangrove biomass from canopy grain analysis using Fourier-based textural ordination of IKONOS images. Remote Sens. Environ. 2007, 109, 379–392. [Google Scholar] [CrossRef]
- Al-Mur, B.A. Green zinc oxide (ZnO) nanoparticle synthesis using mangrove leaf extract from Avicenna marina: Properties and application for the removal of toxic metal ions (Cd2+ and Pb2+). Water 2023, 15, 455. [Google Scholar] [CrossRef]
- Rahman, A.F.; Dragoni, D.; Didan, K.; Barreto-Munoz, A.; Hutabarat, J.A. Detecting large scale conversion of mangroves to aquaculture with change point and mixed-pixel analyses of high-fidelity MODIS data. Remote Sens. Environ. 2013, 130, 96–107. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M. Mangrove conversion and aquaculture development in Vietnam: A remote sensing-based approach for evaluating the Ramsar Convention on Wetlands. Glob. Environ. Change 2007, 17, 486–500. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Smoak, J.M.; Smith, T.J.; Sanders, C.J.; Hoare, A. Organic carbon burial rates in mangrove sediments: Strengthening the global budget. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The loss of species: Mangrove extinction risk and geographic areas of global concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef] [PubMed]
- Blasco, F.; Aizpuru, M.; Gers, C. Depletion of the mangroves of Continental Asia. Wetl. Ecol. Manag. 2001, 9, 255–266. [Google Scholar] [CrossRef]
- Romañach, S.S.; DeAngelis, D.L.; Koh, H.L.; Li, Y.; Teh, S.Y.; Barizan, R.S.R.; Zhai, L. Conservation and restoration of mangroves: Global status, perspectives, and prognosis. Ocean Coast. Manag. 2018, 154, 72–82. [Google Scholar] [CrossRef]
- Sun, Z.; Sun, W.; Tong, C.; Zeng, C.; Yu, X.; Mou, X. China’s coastal wetlands: Conservation history, implementation efforts, existing issues and strategies for future improvement. Environ. Int. 2015, 79, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xiao, X.; Li, X.; Pan, L.; Doughty, R.; Ma, J.; Dong, J.; Qin, Y.; Zhao, B.; Wu, Z.; et al. A mangrove forest map of China in 2015: Analysis of time series Landsat 7/8 and Sentinel-1A imagery in Google Earth Engine cloud computing platform. J. Photogramm. Remote Sens. 2017, 131, 104–120. [Google Scholar] [CrossRef]
- Moberg, F.; Rönnbäck, P. Ecosystem services of the tropical seascape: Interactions, substitutions and restoration. Ocean Coast. Manag. 2003, 46, 27–46. [Google Scholar] [CrossRef]
- Sarker, S.K.; Matthiopoulos, J.; Mitchell, S.N.; Ahmed, Z.U.; Al Mamun, B.; Reeve, R. 1980s–2010s: The world’s largest mangrove ecosystem is becoming homogeneous. Biol. Conserv. 2019, 236, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Jian, S.; Lu, H.; Zhang, Q.; Shen, W.; Han, W.; Yin, Z.; Guo, Q. Restoration of mangrove plantations and colonisation by native species in Leizhou bay, South China. Ecol. Res. 2008, 23, 401–407. [Google Scholar] [CrossRef]
- Chen, L.; Peng, S.; Li, J.; Lin, Z.; Zeng, Y. Competitive control of an exotic mangrove species: Restoration of native mangrove forests by altering light availability. Restor. Ecol. 2013, 21, 215–223. [Google Scholar] [CrossRef]
- Tian, G.; Chen, L.; Peng, S.; Yang, X.; Chen, J.; Chen, M.; Li, J.; Zeng, Y.; Lei, Z. Ecological traits of invasiveness of alien mangrove species Sonneratia apetala. Ecol. Environ. 2010, 19, 3014. [Google Scholar] [CrossRef]
- Zeng, W.J.; Liao, B.W.; Chen, X.R.; Li, J.; Ma, S.Q.; Guan, W. The ecological effect of mangrove Sonneratia apetala mixed with three local mangrove species. Ecol. Sci. 2008, 27, 31–37. [Google Scholar] [CrossRef]
- Li, M.; Liao, B.; Zheng, S.; Chen, Y. Allelopathic effects of Sonneratia apetala aqueous extracts on growth performance of some indigenous mangroves. For. Res. 2004, 17, 641–645. [Google Scholar] [CrossRef]
- Gao, J.; Lundquist, C.J.; Schwendenmann, L. Characterizing landscape patterns in changing mangrove ecosystems at high latitudes using spatial metrics. Estuar. Coast. Shelf Sci. 2018, 215, 1–10. [Google Scholar] [CrossRef]
- Buitre, M.J.C.; Zhang, H.; Lin, H. The mangrove forests change and impacts from tropical cyclones in the philippines using time series satellite imagery. Remote Sens. 2019, 11, 688. [Google Scholar] [CrossRef]
- Chamberlain, D.; Phinn, S.; Possingham, H. Remote sensing of mangroves and estuarine communities in central Queensland, Australia. Remote Sens. 2020, 12, 197. [Google Scholar] [CrossRef]
- Xin, K.; Huang, X.; Hu, J.; Li, C.; Yang, X.; Arndt, S.K. Land use change impacts on heavy metal sedimentation in Mangrove wetlands—A case study in Dongzhai Harbor of Hainan, China. Wetlands 2014, 34, 1–8. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, X.; Wang, Z.; Zhang, T.; Liu, X. Remote sensing based spatial-temporal monitoring of the changes in coastline mangrove forests in China over the last 40 years. Remote Sens. 2021, 13, 1986. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, D.; Guo, W.; Gao, Y.; Su, X.; Wei, B. A review on the application of remote sensing in mangrove ecosystem monitoring. Acta Ecol. Sin. 2013, 33, 4523–4538. [Google Scholar] [CrossRef]
- Giri, C. Observation and monitoring of mangrove forests using remote sensing: Opportunities and challenges. Remote Sens. 2016, 8, 783. [Google Scholar] [CrossRef]
- Valderrama-Landeros, L.; Flores-De-Santiago, F.; Kovacs, J.M.; Flores-Verdugo, F. An assessment of commonly employed satellite-based remote sensors for mapping mangrove species in Mexico using an NDVI-based classification scheme. Environ. Monit. Assess. 2018, 190, 23. [Google Scholar] [CrossRef] [PubMed]
- Everitt, J.H.; Yang, C.; Sriharan, S.; Judd, F.W. Using high resolution satellite imagery to map black mangrove on the Texas Gulf Coast. J. Coast. Res. 2008, 246, 1582–1586. [Google Scholar] [CrossRef]
- Selvam, V. Environmental classification of mangrove wetlands of India. Curr. Sci. 2003, 84, 757–765. [Google Scholar] [CrossRef]
- Hai, P.M.; Tinh, P.H.; Son, N.P.; Van Thuy, T.; Hanh, N.T.H.; Sharma, S.; Hoai, D.T.; Duy, V.C. Mangrove health assessment using spatial metrics and multi-temporal remote sensing data. PLoS ONE 2022, 17, e0275928. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Qin, C.-Z.; Li, H.; Huang, C.; Su, F.-Z. Mapping mangrove forests based on multi-tidal high-resolution satellite imagery. Remote Sens. 2018, 10, 1343. [Google Scholar] [CrossRef]
- Hens, L.; Nierynck, E.; Van, Y.T.; Quyen, N.H.; Hien, L.T.T.; An, L.D. Land cover changes in the extended Ha long city area, north-eastern vietnam during the period 1988–1998. Environ. Dev. Sustain. 2000, 2, 235–252. [Google Scholar] [CrossRef]
- Zhao, C.; Qin, C.-Z.; Wang, Z.; Mao, D.; Wang, Y.; Jia, M. Decision surface optimization in mapping exotic mangrove species (Sonneratia apetala) across latitudinal coastal areas of China. J. Photogramm. Remote Sens. 2022, 193, 269–283. [Google Scholar] [CrossRef]
- Hu, Z.; Wu, J.; Wang, J.; Zhang, Y.; Zhou, H.; Gao, C.; Wang, J.; Wu, G. How exotic Sonneratia species affect the spatiotemporal dynamics of mangroves in Shenzhen Bay, China: A remote sensing perspective. Ecol. Indic. 2023, 153. [Google Scholar] [CrossRef]
- Yang, H.; Xu, B.; Han, C.; Huang, Z. Situation and benefit of mangrove resources in Leizhou peninsula. Ecol. Environ. 2004, 13, 222–224. [Google Scholar] [CrossRef]
- Gupta, K.; Mukhopadhyay, A.; Giri, S.; Chanda, A.; Majumdar, S.D.; Samanta, S.; Mitra, D.; Samal, R.N.; Pattnaik, A.K.; Hazra, S. An index for discrimination of mangroves from non-mangroves using LANDSAT 8 OLI imagery. MethodsX 2018, 5, 1129–1139. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, J.; Jiang, X.; Zhen, J.; Miao, J.; Wang, J.; Wu, G. Reflectance spectroscopy for assessing heavy metal pollution indices in mangrove sediments using XGBoost method and physicochemical properties. Catena 2022, 211, 105967. [Google Scholar] [CrossRef]
- Do, A.N.T.; Tran, H.D.; Ashley, M.; Nguyen, A.T. Monitoring landscape fragmentation and aboveground biomass estimation in Can Gio Mangrove Biosphere Reserve over the past 20 years. Ecol. Inform. 2022, 70. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Mao, D.; Huang, C.; Lu, C. Spatial-temporal changes of China’s mangrove forests over the past 50 years: An analysis towards the Sustainable Development Goals (SDGs). Chin. Sci. Bull. 2021, 66, 3886–3901. [Google Scholar] [CrossRef]
- Zhang, R.; Jia, M.; Wang, Z.; Zhou, Y.; Mao, D.; Ren, C.; Zhao, C.; Liu, X. Tracking annual dynamics of mangrove forests in mangrove National Nature Reserves of China based on time series Sentinel-2 imagery during 2016–2020. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102918. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, Q.; Peng, Y.; Pan, L.; Chen, Y.; Zhang, Y.; Chen, L. Distributions of the Non-Native Mangrove Sonneratia apetala in China: Based on Google Earth Imagery and Field Survey. Wetlands 2022, 42, 35. [Google Scholar] [CrossRef]
- Pimple, U.; Leadprathom, K.; Simonetti, D.; Sitthi, A.; Peters, R.; Pungkul, S.; Pravinvongvuthi, T.; Dessard, H.; Berger, U.; Siri-On, K.; et al. Assessing mangrove species diversity, zonation and functional indicators in response to natural, regenerated, and rehabilitated succession. J. Environ. Manag. 2022, 318, 115507. [Google Scholar] [CrossRef]
- Liao, W.B.; Lan, C.Y.; Zan, Q.J.; Wong, Y.S.; Tam, N.F.Y. Growth dynamics and self-thinning of the dominant populations in the mangrove community. Acta Bot. Sin. 2004, 46, 522–532. [Google Scholar]
- Peng, Z.; Wang, B.; Zhao, H. Population dynamics and spread characteristics of alien mangrove species Sonneratia apetala in Sanya River, China. Chin. J. Ecol. 2021, 40, 23. [Google Scholar] [CrossRef]
- Levine, J.M.; Vilà, M.; Antonio, C.M.D.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. B Biol. Sci. 2003, 270, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Gorchov, D.L.; Trisel, D.E. Competitive effects of the invasive shrub, Lonicera maackii (Rupr.) Herder (Caprifoliaceae), on the growth and survival of native tree seedlings. Plant Ecol. 2003, 166, 13–24. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Hardy, G.E.S.J.; Van Le, T.; Nguyen, H.Q.; Le, D.H.; Van Nguyen, T.; Dell, B. Mangrove dieback and leaf disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam. Forests 2021, 12, 1273. [Google Scholar] [CrossRef]
- Ren, H.; Lu, H.; Shen, W.; Huang, C.; Guo, Q.; Li, Z.; Jian, S. Sonneratia apetala Buch. Ham in the mangrove ecosystems of China: An invasive species or restoration species? Ecol. Eng. 2009, 35, 1243–1248. [Google Scholar] [CrossRef]
- Ng, S.; Corlett, R. The bad biodiversity: Alien plant species in Hong Kong. Biodivers. Sci. 2002, 10, 109–118. [Google Scholar] [CrossRef]
- Forman, R.T.T. Land Mosaics: The Ecology of Landscapes and Regions; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Li, C.; Liu, S.; Fan, H.; Dai, H. A patch-based method for mechanism analysis on spatial dynamics of mangrove distribution. Acta Ecol. Sin. 2012, 32, 4329–4342. [Google Scholar] [CrossRef]
- Li, M.; Liao, B.W.; Guan, W.; Zheng, S.F.; Chen, Y.J. Survey on cold damage of mangroves in Guangdong Province. Prot. For. Sci. Technol. 2009, 2, 29–31. [Google Scholar] [CrossRef]
- Huang, X.; Yuan, J.; Wang, X.; Kwan, K.; Zhang, Q. Dynamics of mangrove change: Insights from 30-year observations of Maowei Sea. J. Mar. Sci. 2022, 40, 132. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index Name | Equation | Description |
|---|---|---|
| Normalized difference water index (NDWI) | The index was used to determine the threshold between vegetation and water. |
| Index Name | Formulas | Descriptive |
|---|---|---|
| Number of patches (NP) | NP = N | Total number of patches in the landscape |
| Patch area (CA) | CA = A | The sum of the areas of all patches of a given class in a landscape. |
| Total edge length (TE) | TE = E | Total perimeter of patches of a given patch type. |
| Area-weighted mean shape index (AWMSI) | The perimeter-to-area ratio of area-weighted patches was calculated by dividing the perimeter of each patch by the square root of the area and multiplying the result by a square correction factor. |
| Year | Mangrove Area/hm2 | Average Annual Rate of Change/% | Percentage of Mangroves/% | Sonneratia apetala Area/hm2 | Average Annual Rate of Change/% | Percentage of Sonneratia apetala/% | Local Mangrove Area/hm2 | Average Annual Rate of Change/% | Percentage of Local Mangroves/% |
|---|---|---|---|---|---|---|---|---|---|
| 2010 | 274.17 | 0.3718 | 111.15 | 0.1507 | 163.02 | 0.2211 | |||
| 2011 | 301.02 | 0.098 | 0.4082 | 121.14 | 0.0898 | 0.1642 | 179.88 | 0.1034 | 0.2439 |
| 2012 | 308.71 | 0.025 | 0.4186 | 101.97 | −0.1582 | 0.1382 | 206.73 | 0.1492 | 0.2803 |
| 2013 | 319.34 | 0.034 | 0.4331 | 149.22 | 0.4633 | 0.2023 | 170.11 | −0.1771 | 0.2307 |
| 2014 | 321.25 | 0.006 | 0.4356 | 160.65 | 0.07651 | 0.2178 | 160.60 | −0.0559 | 0.2177 |
| 2015 | 332.10 | 0.033 | 0.4503 | 157.59 | −0.0190 | 0.2137 | 163.66 | 0.0191 | 0.2219 |
| 2016 | 332.23 | 0.0003 | 0.4505 | 167.65 | 0.0638 | 0.2273 | 164.57 | 0.0055 | 0.2231 |
| 2017 | 366.55 | 0.103 | 0.4971 | 219.06 | 0.3066 | 0.2970 | 147.49 | −0.1038 | 0.2000 |
| 2018 | 369.15 | 0.007 | 0.5006 | 237.6 | 0.0846 | 0.3222 | 131.55 | −0.1081 | 0.1784 |
| 2019 | 380.58 | 0.030 | 0.5161 | 242.37 | 0.0201 | 0.3286 | 135.17 | 0.0275 | 0.1833 |
| 2020 | 382.20 | 0.004 | 0.5183 | 250.21 | 0.0323 | 0.3393 | 127.36 | −0.0578 | 0.1727 |
| 2021 | 382.37 | 0.0004 | 0.5185 | 251.7 | 0.0059 | 0.3413 | 125.87 | −0.0116 | 0.1707 |
| 2022 | 382.95 | 0.0015 | 0.5193 | 256.23 | 0.0179 | 0.3474 | 121.35 | −0.0359 | 0.1645 |
| 2023 | 383.42 | 0.0012 | 0.5199 | 254.81 | −0.0055 | 0.3455 | 125.06 | 0.0305 | 0.1696 |
| Aggregate change | 109.25 | 0.3985 | 0.1481 | 143.66 | 1.2924 | 0.1948 | −37.95 | −0.2328 | −0.0514 |
| Year | Number of Sonneratia apetala Patches | Number of Local Mangrove Patches | Area of Sonneratia apetala Patches/hm2 | Area of Local Mangrove Patches/hm2 | Total Edge Length of Sonneratia apetala /hm | Total Edge Length of Local Mangroves /hm | Area-Weighted Mean Shape Index of Sonneratia apetala | Area-Weighted Mean Shape Index of Local Mangroves |
|---|---|---|---|---|---|---|---|---|
| 2010 | 140 | 61 | 110.97 | 182.25 | 6.630 | 6.690 | 3.32 | 4.12 |
| 2011 | 138 | 81 | 126.99 | 192.42 | 7.608 | 8.046 | 3.09 | 4.44 |
| 2012 | 78 | 83 | 121.59 | 198.27 | 5.646 | 7.260 | 3.64 | 4.76 |
| 2013 | 111 | 85 | 160.11 | 169.83 | 8.010 | 7.386 | 3.7 | 3.8 |
| 2014 | 84 | 78 | 176.13 | 155.97 | 8.184 | 6.540 | 4.57 | 3.54 |
| 2015 | 99 | 90 | 161.73 | 170.37 | 6.930 | 7.026 | 4.06 | 3.74 |
| 2016 | 322 | 316 | 174.04 | 160.35 | 11.896 | 11.214 | 5.67 | 4.79 |
| 2017 | 311 | 445 | 224.32 | 143.98 | 15.390 | 14.348 | 7 | 6.31 |
| 2018 | 246 | 309 | 240.11 | 130.77 | 14.720 | 12.168 | 7.75 | 5.71 |
| 2019 | 370 | 482 | 247.42 | 133.45 | 16.278 | 14.166 | 9.52 | 3.88 |
| 2020 | 359 | 475 | 240.83 | 140.04 | 15.808 | 14.302 | 9.05 | 4.3 |
| 2021 | 305 | 443 | 225.31 | 155.56 | 14.030 | 15.756 | 7.82 | 4.08 |
| 2022 | 318 | 379 | 240.94 | 139.93 | 13.608 | 12.688 | 7.99 | 3.82 |
| 2023 | 279 | 400 | 231.82 | 149.05 | 13.222 | 13.988 | 7.72 | 4.35 |
| Aggregate change | 139 | 339 | 120.85 | −33.2 | 6.592 | 7.298 | 4.4 | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Deng, Y.; Zhong, W.; Xie, Z.; Liu, H.; Li, Z.; Jia, Y.; Li, X.; Chen, R.; Peng, X.; et al. Tracking the Expansion of Sonneratia apetala and Its Impact on Local Mangroves Using Time-Series Remote Sensing Data. Sustainability 2025, 17, 1069. https://doi.org/10.3390/su17031069
Feng X, Deng Y, Zhong W, Xie Z, Liu H, Li Z, Jia Y, Li X, Chen R, Peng X, et al. Tracking the Expansion of Sonneratia apetala and Its Impact on Local Mangroves Using Time-Series Remote Sensing Data. Sustainability. 2025; 17(3):1069. https://doi.org/10.3390/su17031069
Chicago/Turabian StyleFeng, Xuesong, Yingbin Deng, Weiping Zhong, Zhiyi Xie, Hua Liu, Zhao Li, Yiwen Jia, Xin Li, Renrong Chen, Xiaoyan Peng, and et al. 2025. "Tracking the Expansion of Sonneratia apetala and Its Impact on Local Mangroves Using Time-Series Remote Sensing Data" Sustainability 17, no. 3: 1069. https://doi.org/10.3390/su17031069
APA StyleFeng, X., Deng, Y., Zhong, W., Xie, Z., Liu, H., Li, Z., Jia, Y., Li, X., Chen, R., Peng, X., Deng, Y., Li, M., Li, M., & Guo, D. (2025). Tracking the Expansion of Sonneratia apetala and Its Impact on Local Mangroves Using Time-Series Remote Sensing Data. Sustainability, 17(3), 1069. https://doi.org/10.3390/su17031069