3.1. Lanthanide Phosphate Precipitates
High-resolution X-ray photoelectron spectroscopy (XPS) was used to analyze the interactions between lanthanides (Ln) and phosphorus (P), focusing on the shifts in binding energies that indicate chemical bonding. In the La matrix, peaks corresponding to the La 3d
5/2 and La 3d
3/2 orbitals—sublevels within the 3d electron shell—were observed. These orbitals represent core-level electrons, and their binding energies are sensitive indicators of the chemical environment. The La 3d
5/2 and La 3d
3/2 peaks appeared at 836.0 eV and 853.0 eV, respectively, consistent with inner-sphere complex formation [
29]. Similarly, in the Ce system, binding energies for the Ce 3d
3/2 (902.4 eV) and Ce 3d
5/2 (883.8 eV) orbitals—also core electron states—were identified, which play key roles in Ce-P interactions. For phosphorus, the most reactive electrons reside in the 2p
1/2 and 2p
3/2 subshells, with binding energies at 136 eV and 135 eV, respectively [
29].
The formation of inner-sphere bonds between phosphorus and lanthanides altered these valence electron energies.
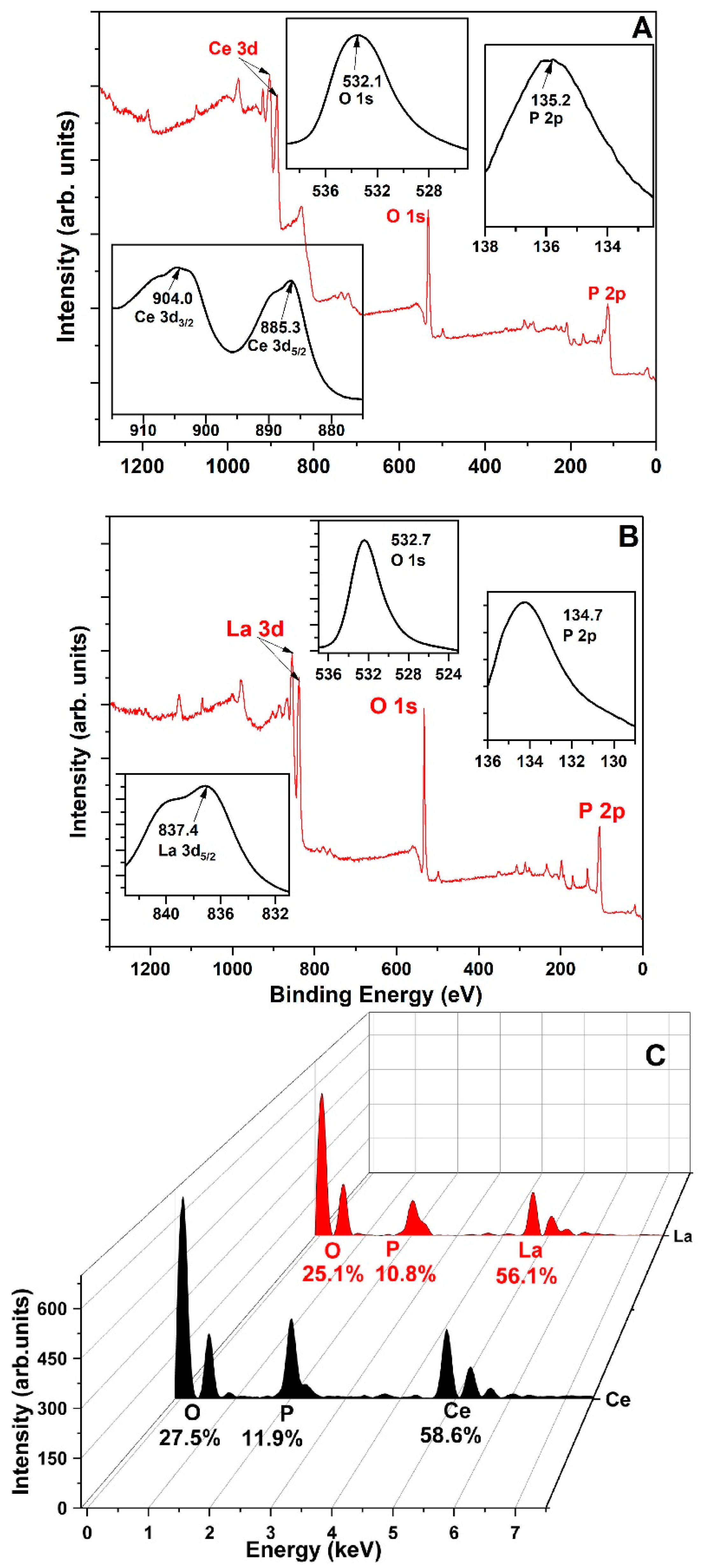
Figure 1A,B present the XPS spectra of Ln-P, showing clear peaks for Ce, La, P, and O. In the Ce-PO
4 complex, the Ce 3d
3/2 binding energy shifted to 904.0 eV—an increase of 1.6 eV. Likewise, in the La-PO
4 system, the La 3d
5/2 peak shifted by 1.4 eV. These shifts confirm the successful formation of La-O-P and Ce-O-P bonding structures.
Figure 1C displays EDS results, showing elemental ratios of La, Ce, O, and P that closely match the theoretical stoichiometry for pure Ce-/La-PO
4. Impurity levels were minimal, with CePO
4 containing up to 1.9% impurities and LaPO
4 up to 7.9%. Given their purity, these precipitates are suitable for use as phosphorus sources in plant growth applications.
Table 4 summarizes the characteristics of the centrate wastewater used for lanthanide treatment, including the initial concentrations and the percentage removal of phosphorus (PO
43−-P) and other key parameters following treatment with CeCl
3 and LaCl
3. The results highlight the high efficiency of both lanthanides in phosphorus removal (>98%) with moderate effects on other water quality indicators.
3.2. Germination Rate and Hydrogen Peroxide
Seed germination begins with water uptake, which activates metabolic processes essential for transforming a dormant seed into a growing plant. These metabolic activities not only determine the likelihood of germination but also influence the future health of the plant. To assess the effect of metal salts on germination, corn kernels (
n = 5 per treatment) were grown in Petri dishes with metal solutions at concentrations of 0, 50, 100, 500, 1000, 2000, and 5000 mg M
3+/L. After five days of incubation in the dark, the seeds were assessed for germination rate, root and shoot lengths, and hydrogen peroxide (H
2O
2) production. A seed was considered germinated if it developed a visible embryonic axis, regardless of root or shoot length. As shown in
Table A1 and
Table A2, no clear correlation was observed between metal concentration and germination rate. This inconsistency could be attributed to seed dormancy at the time of planting, which may have hindered germination even under otherwise optimal conditions (i.e., proper moisture, light, oxygen, pH, and temperature). However, root and shoot lengths showed a more consistent trend. At concentrations below 1000 mg M
3+/L, the root and shoot lengths were comparable to those observed in the control group treated with deionized water. At concentrations above 1000 mg M
3+/L, both root and shoot lengths decreased significantly (
Table A1 and
Table A3).
To further investigate these changes, H
2O
2 levels were measured in the germinated kernels, as H
2O
2 is a key reactive oxygen species (ROS) involved in seed metabolism. Water uptake during germination triggers a cascade of biochemical reactions, including the production of ROS such as H
2O
2 [
30]. It is estimated that 1−5% of the oxygen consumed during germination is converted into H
2O
2 [
31]. Most H
2O
2 in seeds is generated from the dismutation of superoxide anions (O
2•⁻), which are produced via electron leakage from mitochondrial complexes I, II, and III [
32,
33].
Another significant source of H
2O
2 is NADPH oxidase, located on the plasma membrane, which is especially active during biotic or abiotic stress. This process—known as the “oxidative burst” or respiratory burst oxidase homologs (RBOHs)—is triggered by melatonin and modulated by S-nitrosylation and phosphorylation, which enhance O
2•⁻ and, subsequently, H
2O
2 production [
31,
34,
35]. Additional contributors to ROS production during germination include polyamine oxidase [
35,
36] and peroxisomal metabolism [
37,
38]. Unlike other ROS, H
2O
2 is relatively stable and can migrate throughout the seed and even into the surrounding environment [
39]. While moderate levels of H
2O
2 are beneficial and function as signaling molecules, excessive accumulation can damage DNA, proteins, and lipids, thereby inhibiting germination [
40].
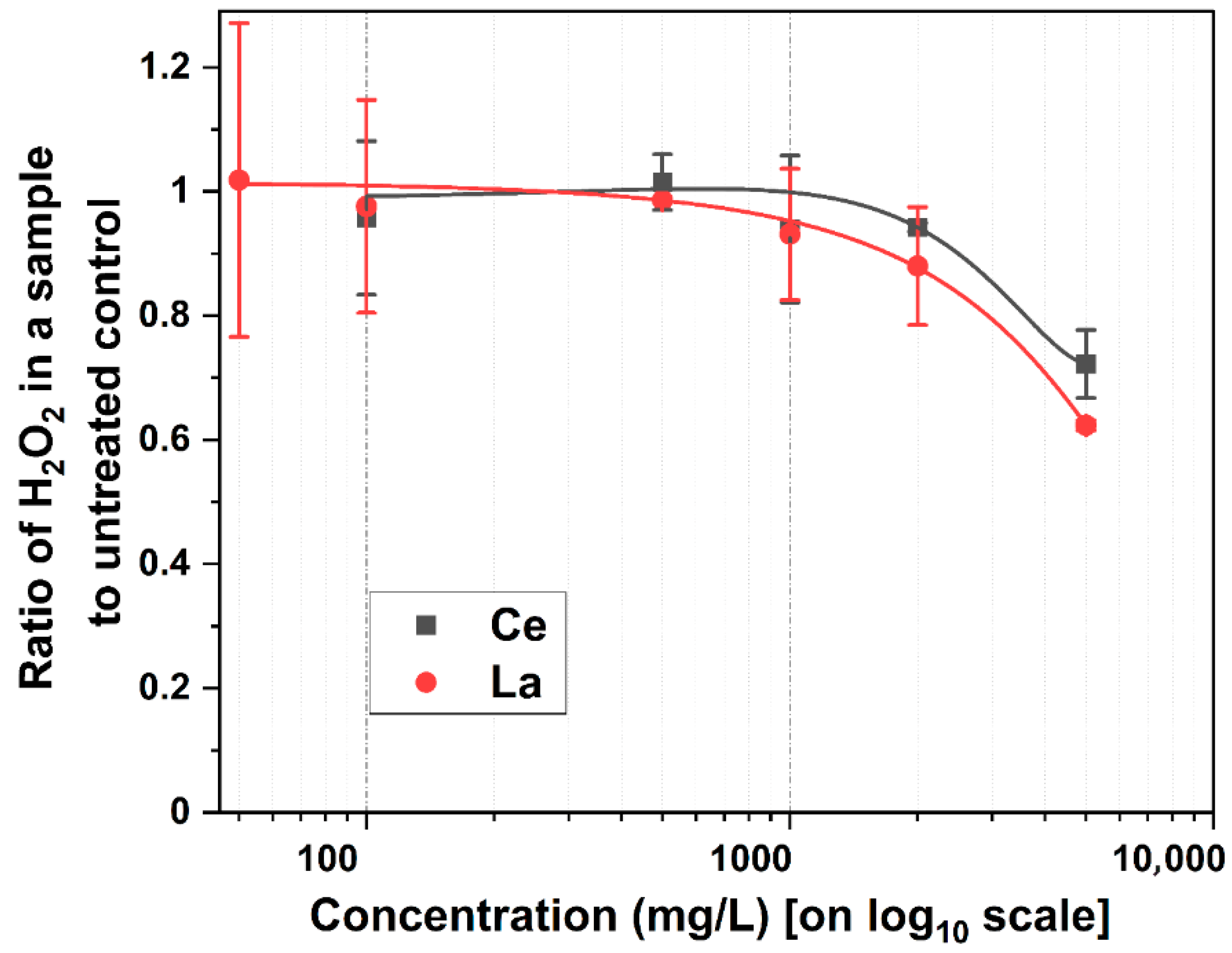
Figure 2 illustrates the ratio of H
2O
2 production relative to metal concentration, compared to the control. Despite increasing metal concentrations, H
2O
2 production remained relatively unchanged up to 1000 mg/L. This suggests that low-to-moderate metal levels did not significantly impact oxidative stress. This observation aligns with reports that exogenous H
2O
2 can promote germination by modulating the proteome, transcriptome, and hormone signaling pathways [
41]. Specifically, H
2O
2 activates mitogen-activated protein kinases (MAPKs) that suppress abscisic acid while also promoting protein carbonylation and activating the pentose phosphate pathway (PPP). This activation leads to increased NADPH production, which has antioxidant properties that support seed germination and early plant development [
30,
42]. However, at 2000 mg/L and above, a decline in H
2O
2 levels was observed. This reduction coincided with the suppression of root and shoot development, suggesting that excess metal concentrations may have impaired the physiological processes critical to germination and early growth.
The reduction in root and shoot lengths at higher metal concentrations may be attributed to pH changes, particularly in environments treated with Al
3+ and Fe
3+. As the metal concentration increased, the pH of the Al
3+ and Fe
3+ solutions decreased significantly, as shown in
Table A4. These coagulants are typically formulated using strong acids such as sulfuric acid and hydrochloric acid [
6]. While a slight reduction in pH can help weaken the seed coat—facilitating water absorption into the endosperm—excessive acidification can lead to “water flooding.” This condition disrupts oxygen availability within the seed, thereby inhibiting the germination signaling pathways. In contrast, the pH in Ce
3+ and La
3+ treatments remained relatively stable, with Ce
3+ at 5.3 ± 0.1 and La
3+ at 5.2 ± 0.2 (
Table A5). In these cases, the decline in root and shoot development is more likely due to elevated total dissolved solids (TDSs), which increase electrical conductivity. Excessive TDSs create osmotic stress, limiting water uptake into the seed—a phenomenon often referred to as physiological drought [
43]. This water deficit prevents the initiation of germination or restricts early root development.
Figure 3 illustrates corn kernel germination in a Ce
3+ environment. At concentrations between 0 and 2000 mg/L, kernels were frequently covered by a fungal mat. The presence of this fungal growth is believed to aid in nutrient mobilization from more distant parts of the root system [
44], thereby improving germination and seedling development. However, at 5000 mg/L, no fungal development was observed. Without fungal assistance, the kernels likely depended solely on localized nutrients near the root surface, which may have limited their growth potential.
3.3. Corn Physical Characteristics
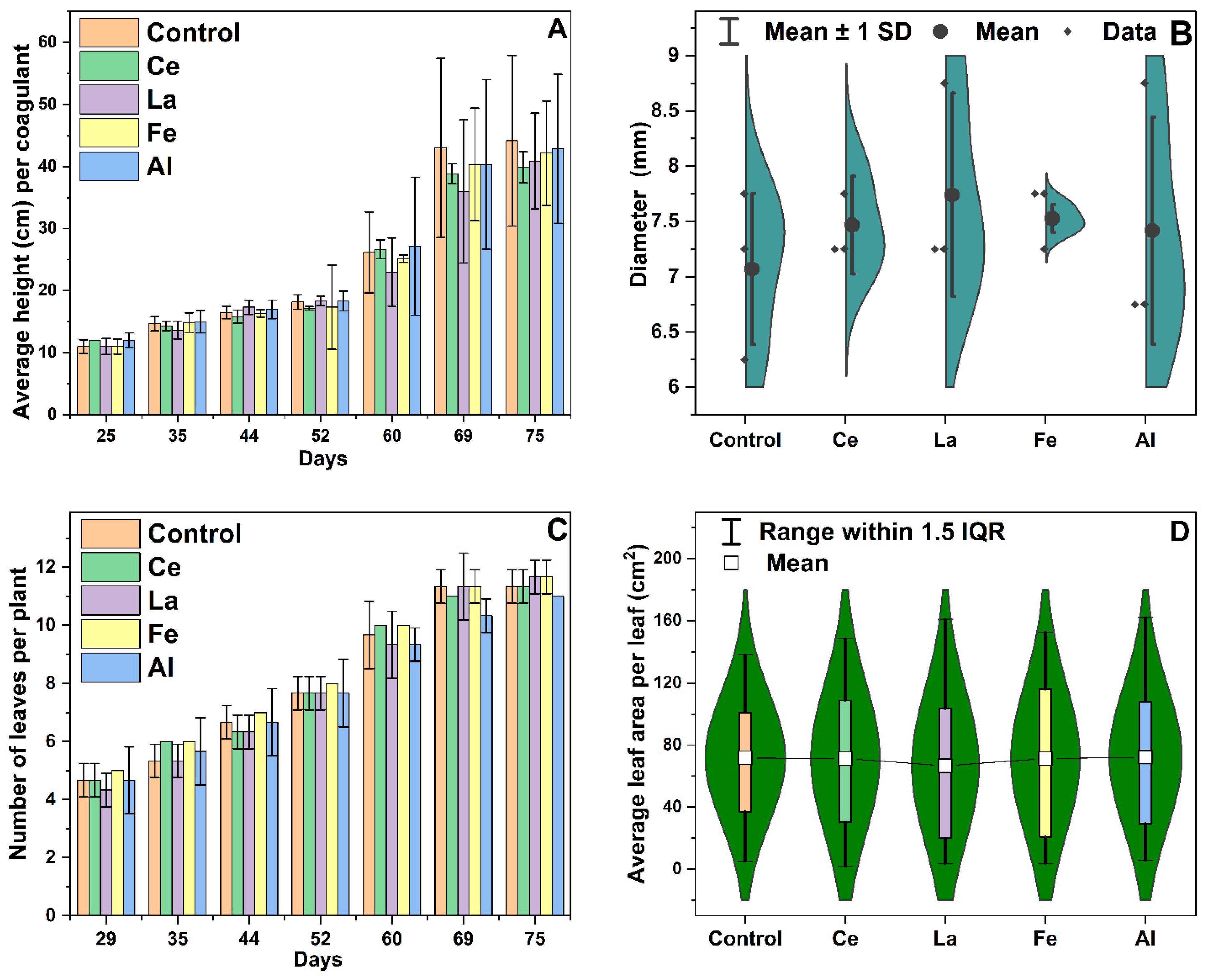
Throughout the corn growth cycle, key physical characteristics were monitored, including plant height, stem diameter, the number of leaves, leaf area, and dry biomass. These parameters serve as indicators of overall plant health and development. By day 75, each pot had received a total of 4.59 L of LRWW. The average plant height was 42.0 ± 1.7 cm, with corn grown under lanthanum treatment exhibiting the shortest average height at 39.9 ± 11.5 cm (
Figure 4A). The average stem diameter across all treatments was 7.44 ± 0.65 mm, with no statistically significant differences observed between treatments (
Figure 4B, ρ > 0.05). Leaf number increased steadily until day 75, at which point tassels (male flowers) had emerged on all plants. At maturity, plants produced between 11 and 12 leaves (
Figure 4C). Leaves play a crucial role in plant physiology, particularly in photosynthesis, where they facilitate CO
2 uptake and water vapor release. Consequently, leaf development influences sugar production and has been correlated with crop yield in several studies [
45,
46]. At maturity (day 75), the average leaf area per plant was 70.6 cm
2. Plants treated with aluminum showed the largest average leaf area, 72.2 cm
2 (
Figure 4D). However, no statistically significant differences (ρ > 0.05) were observed among treatments, including lanthanides, the control, and other coagulants. These results suggest that lanthanide treatments do not adversely affect corn leaf development.
3.4. Photosynthetic Parameters
The primary function of stomata is to regulate the exchange of gases—specifically, minimizing water loss while maximizing carbon dioxide (CO
2) uptake. While CO
2 uptake is essential for photosynthesis, excessive water loss poses a threat to plant survival. Plants mitigate this trade-off by optimizing photosynthesis to offset the cost of water loss. Stomatal conductance (gsw) reflects the rate at which CO
2 enters and water vapor exits the leaf. Throughout the corn growth cycle, gsw remained relatively consistent across all treatments involving different coagulants (
Figure 5A). As a result, transpiration rates also remained stable, showing no significant variation among treatments (
Figure 5B). To assess photosynthetic induction, the electron transport rate (ETR) and quantum efficiency (PhiPS2) were measured. ETR values were not statistically different from the control under any coagulant treatment throughout the experiment (
Figure 5C). Similarly, PhiPS2 values remained above 0.5 regardless of metal addition, indicating that the photosynthetic machinery of the plants was not adversely affected (
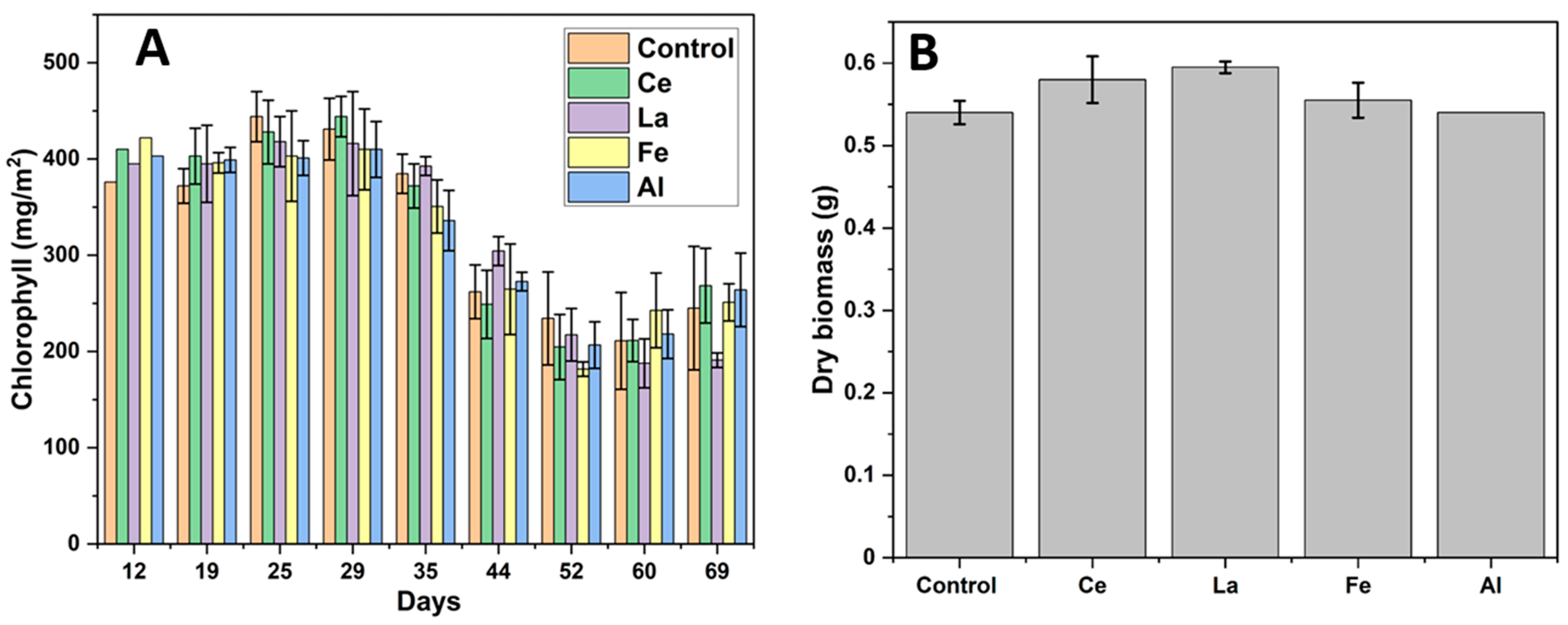
Figure 5D). Quantum efficiency values greater than 0.5 suggest that lanthanides did not negatively impact corn photosynthesis. This consistency is likely due to stable chlorophyll production throughout the experiment (
Figure 6A) [
47]. A slight decline in chlorophyll levels observed after day 35 can be attributed to reduced daylight hours during the winter season when the study was conducted.
3.5. Metals Uptake
Due to the strong bond formed between lanthanides and phosphate (Ln–P), it was initially anticipated that Ln
3+ might reduce phosphorus availability to corn by forming insoluble Ln–P precipitates. Metal uptake analysis was performed on 94-day-old corn plants. Each pot treated with cerium or lanthanum received 165.9 mg of Ce
3+ and 164.9 mg of La
3+, respectively.
Figure 7A,B show the uptake of Ce, La, P, Ca, and Mg in the roots of plants grown in Ce and La environments. Compared to the control (
Figure 7C), Ln
3+ treatments did not inhibit or enhance phosphorus uptake. If any interaction between Ln and phosphorus occurred, it is likely that soil fungi and/or mycorrhizae may have played a role in breaking down the Ln-O-P bond to release phosphorus [
48], although this mechanism remains unclear and warrants further research. Trace amounts of cerium and lanthanum were detected in the roots, but none were found in the stems or leaves. This absence suggests limited translocation of lanthanides within the plant, likely due to their moderate to high binding affinity with silica-based substrates like sand [
49,
50], which may immobilize them in the soil matrix.
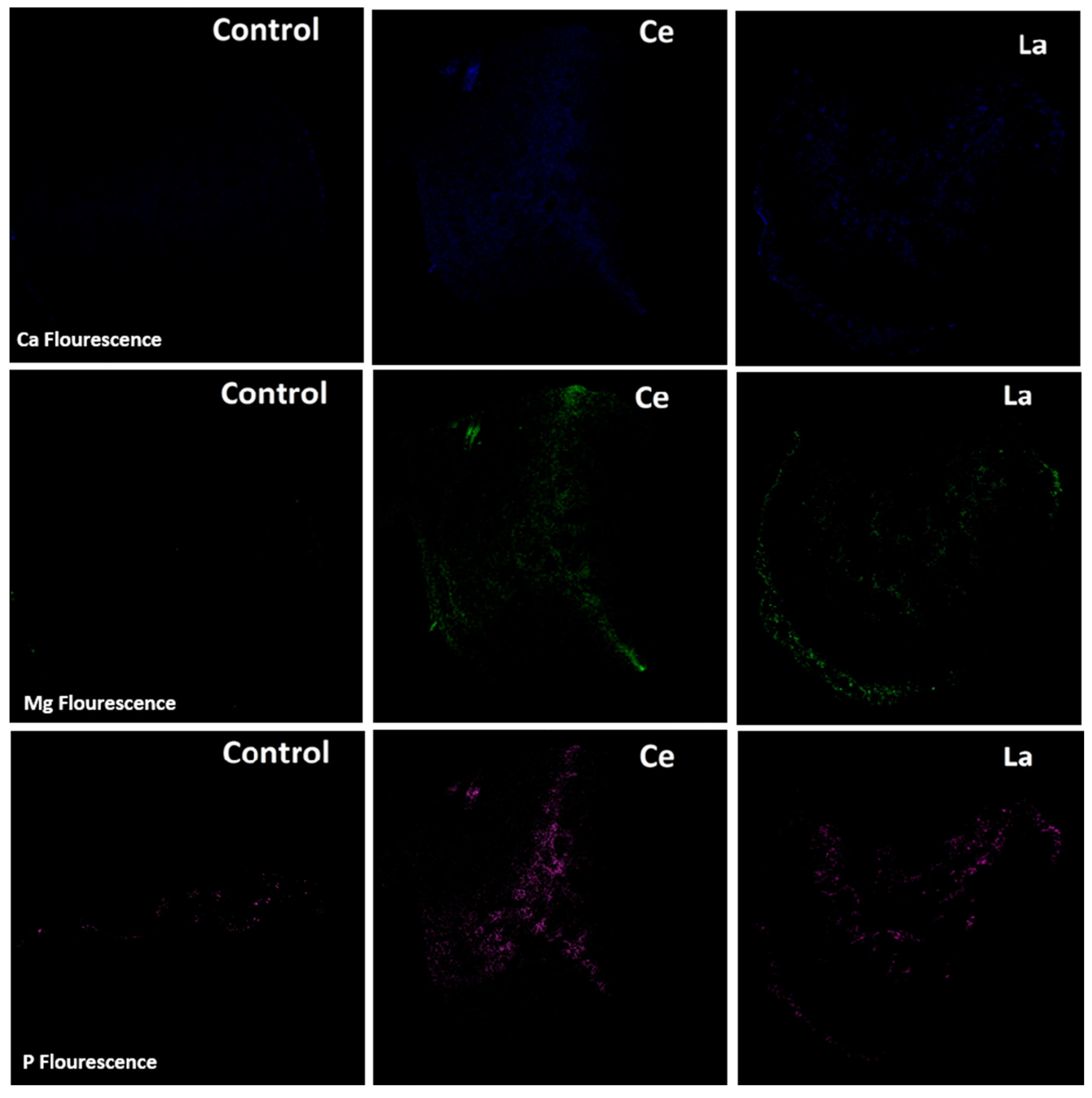
The presence of other metals (P, Ca, and Mg) in the stems and leaves likely resulted from capillary transport through the plant’s vascular system, as the irrigation method used (drip irrigation) avoided direct application of LRWW to aerial parts. Based on fluorescence intensity results (
Figure 8), the presence of rare earth elements (REEs) in the root zone appeared to enhance the uptake of P, Ca, and Mg compared to the control. Similar findings have been reported in soybeans, where exposure to lanthanum improved the uptake of metals such as Ca, K, and P [
51,
52]. Although the precise mechanism by which lanthanides enhance nutrient uptake is not fully understood, one hypothesis is that their interaction with soil silica may liberate micro- and macronutrients, increasing their bioavailability to plant roots and associated fungi. Additionally, as lanthanides enter root tissues, they may create micro-pores or “gates” that facilitate the entry of smaller ions such as Fe and P [
53]. This enhanced metal uptake may explain the slightly higher biomass observed in corn plants irrigated with LRWW (
Figure 6B). These findings support the potential benefits of using Ln-P and LRWW as phosphorus sources in crop production.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}