
In the Spotlight: Bat Activity Shifts in Response to Intense Lighting of a Large Railway Construction Site



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bat Monitoring
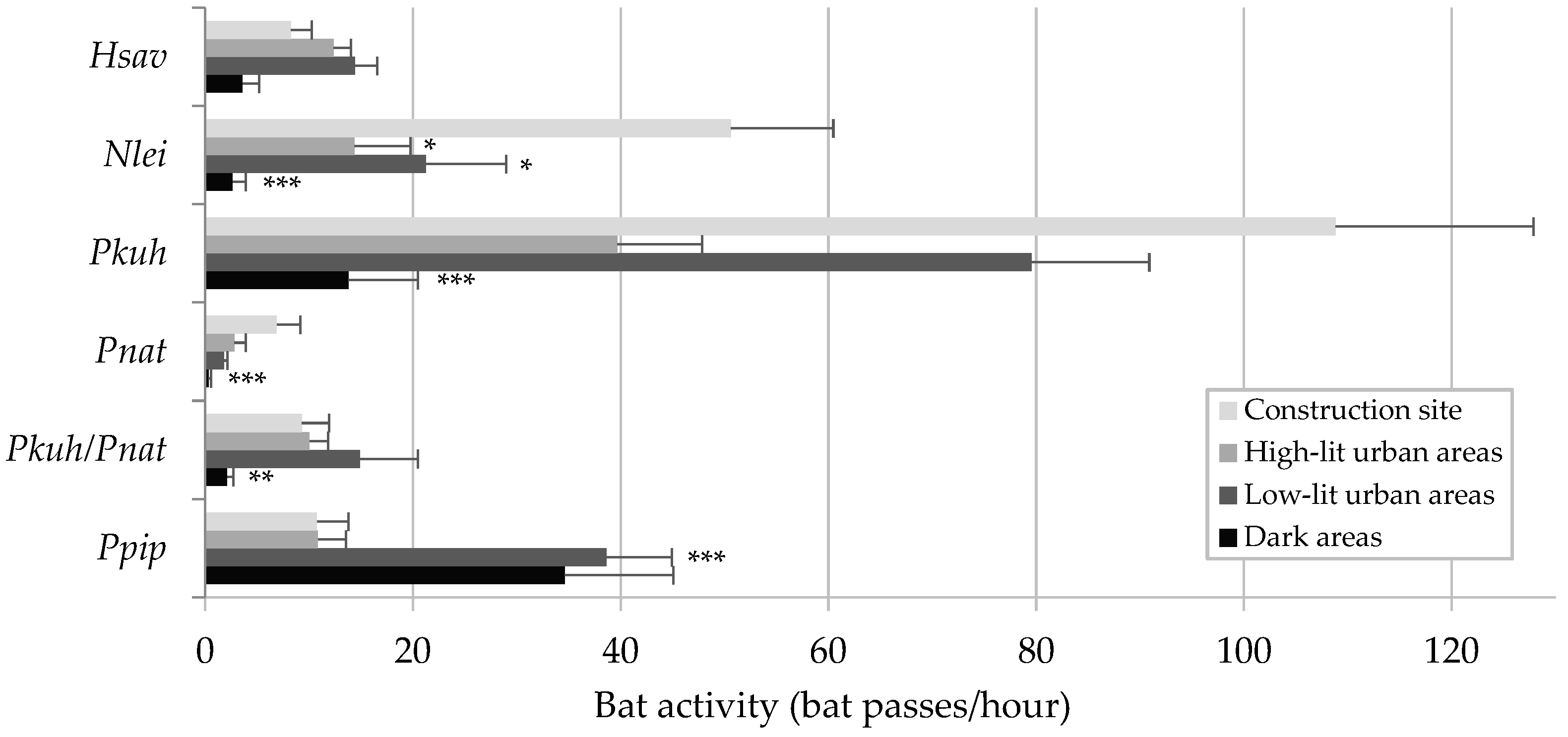
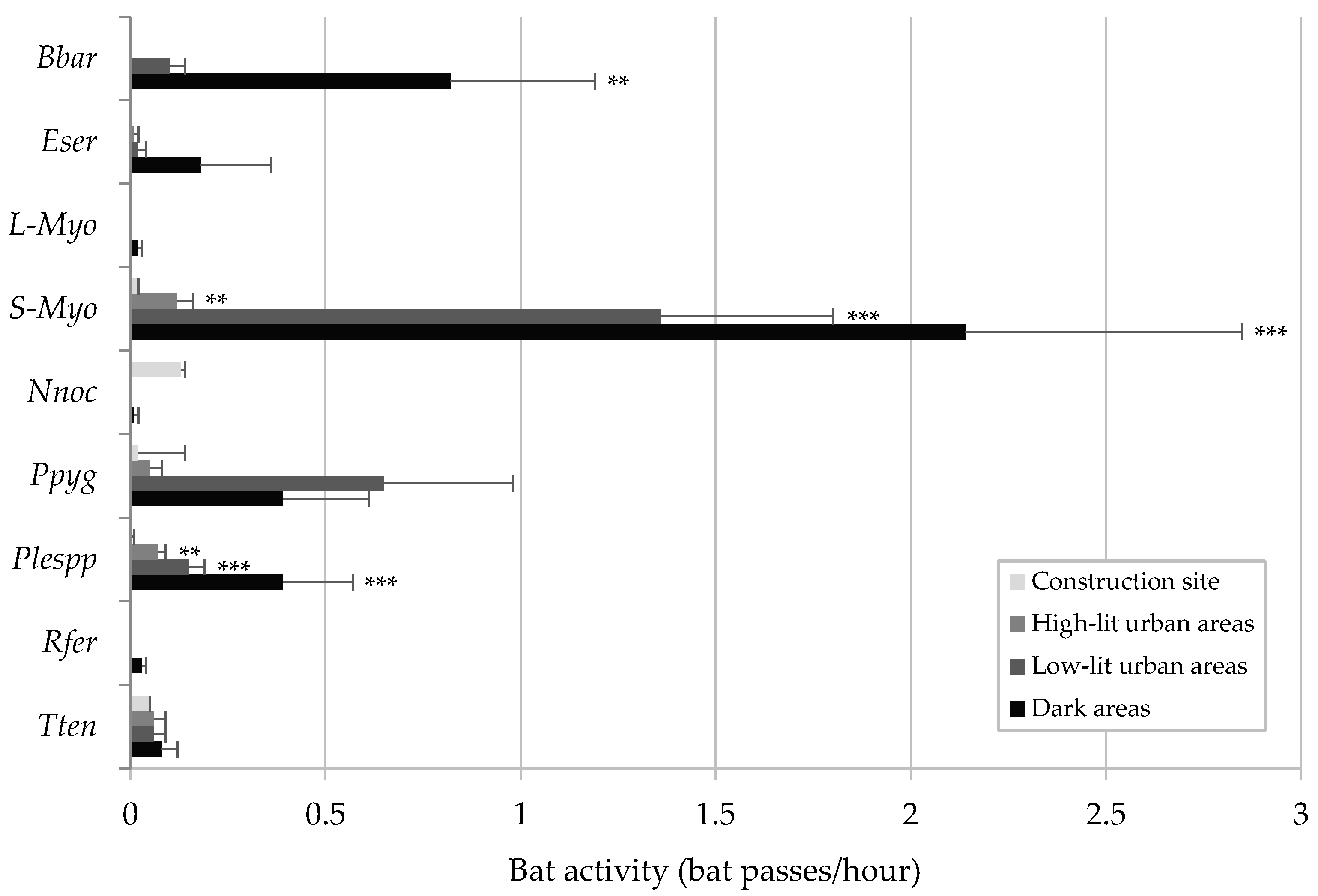
- Brightly lit area within the construction site: This area features strong and continuous artificial lighting every night. Three monitoring sites were inside the construction site, while two were outside the construction site but within the outermost fence of the area. The maximum light intensity was 146.58 ± 75.23 lux.
- High-lit urban area: This included four monitoring sites in the centre of the village of Chiomonte and one in the hamlet of Ramats, where artificial lighting levels are relatively high. The maximum light intensity was 66.94 ± 48.95 lux.
- Low-lit urban area: Four monitoring sites were established in the suburbs of Chiomonte and one in the hamlet of Ramats, where artificial lighting is reduced. The maximum light intensity was 20.26 ± 13.31 lux.
- Dark areas: These sites were in unlit natural areas near the construction site, where artificial lighting is nearly absent. Specifically, two sites were in forest environments, two in ecotonal environments, and one in an open environment. The maximum light intensity was 0.48 ± 0.86 lux.
2.3. Acoustic Analysis
2.4. Statistical Analysis
- Maximum light intensity;
- Elevation, measured in the field via GPS device (GPSMAP 66s, Garmin, Shaffhouse, Switzerland);
- Linear distance from the construction site;
- Linear distance from the nearest watercourse, using the regional vector dataset from Arpa Piemonte based on the European Water Framework Directive (WFD) 2000/60/EC (https://www.geoportale.piemonte.it/geonetwork/srv/api/records/arlpa_to:01.01.04-D_2011-06-14-16:09; accessed on 10 December 2023);
- Total built-up surface area within a 500 m radius, from the 2015 Imperviousness Density dataset by the European Copernicus Land Monitoring Service (CLMS; https://land.copernicus.eu/en/dataset-catalog; accessed on 10 December 2023);
- Total forest area within 500 m, from the 2015 Tree Cover Density dataset by CLMS;
- Total area occupied by meadows and pastures within 500 m, from the 2015 Grassland dataset by CLMS;
- Overall length of ecotonal strips within 500 m, based on the third-level classification of the 2010 Land Cover Piemonte vector dataset (www.regione.piemonte.it/sit; accessed on 10 December 2023).
3. Results
3.1. Comparative Analysis across Areas at Different Lighting Levels
3.2. Statistical Models
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Falchi, F.; Cinzano, P.; Duriscoe, D.M.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef]
- Bruce-White, C.; Shardlow, M. A Review of the Impact of Artificial Light on Invertebrates; Buglife-the Invertebrate Conservation Trust: Buglife, UK, 2011. [Google Scholar]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Shivanna, K.R. Impact of light pollution on nocturnal pollinators and their pollination services. Proc. Natl. Acad. Sci. India–Phys. Sci. 2022, 88, 626–633. [Google Scholar] [CrossRef]
- Russart, K.L.; Nelson, R.J. Artificial light at night alters behavior in laboratory and wild animals. J. Exp. Zoöl. Part A Ecol. Integr. Physiol. 2018, 329, 401–408. [Google Scholar] [CrossRef]
- Stone, E.L.; Harris, S.; Jones, G. Impacts of artificial lighting on bats: A review of challenges and solutions. Mamm. Biol. 2015, 80, 213–219. [Google Scholar] [CrossRef]
- Voigt, C.C.; Dekker, J.; Fritze, M.; Gazaryan, S.; Hölker, F.; Jones, G.; Lewanzik, D.; Limpens, H.J.G.A.; Mathews, F.; Rydell, J.; et al. The Impact of Light Pollution on Bats Varies According to Foraging Guild and Habitat Context. BioScience 2021, 71, 1103–1109. [Google Scholar] [CrossRef]
- Wakefield, A.; Stone, E.L.; Jones, G.; Harris, S. Light-emitting diode street lights reduce last-ditch evasive manoeuvres by moths to bat echolocation calls. R. Soc. Open Sci. 2015, 2, 150291. [Google Scholar] [CrossRef] [PubMed]
- Minnaar, C.; Boyles, J.G.; Minnaar, I.A.; Sole, C.L.; McKechnie, A.E. Stacking the odds: Light pollution may shift the balance in an ancient predator–prey arms race. J. Appl. Ecol. 2014, 52, 522–531. [Google Scholar] [CrossRef]
- Falcón, J.; Torriglia, A.; Attia, D.; Viénot, F.; Gronfier, C.; Behar-Cohen, F.; Martinsons, C.; Hicks, D.G. Exposure to Artificial Light at Night and the Consequences for Flora, Fauna, and Ecosystems. Front. Neurosci. 2020, 14, 602796. [Google Scholar] [CrossRef] [PubMed]
- Briolat, E.S.; Gaston, K.J.; Bennie, J.; Rosenfeld, E.J.; Troscianko, J. Artificial nighttime lighting impacts visual ecology links between flowers, pollinators and predators. Nat. Commun. 2021, 12, 4163. [Google Scholar] [CrossRef] [PubMed]
- Barré, K.; Kerbiriou, C.; Ing, R.-K.; Bas, Y.; Azam, C.; Le Viol, I.; Spoelstra, K. Bats seek refuge in cluttered environment when exposed to white and red lights at night. Mov. Ecol. 2021, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Camerano, P.; Giannetti, F.; Terzuolo, P.G.; Guiot, E. La Carta Forestale Del Piemonte—Aggiornamento 2016. IPLA S.p.A.; Regione Piemonte 2017. Available online: https://www.geoportale.piemonte.it/geonetwork/srv/ita/catalog.search#/metadata/r_piemon:812c28a8-763b-4c74-81a3-c5fe1ed99c68 (accessed on 10 December 2023).
- Istat. Il Censimento Permanente della Popolazione in Piemonte—Anno 2020. 2022. Available online: https://www.istat.it/it/archivio/268878 (accessed on 10 December 2023).
- CIPE. Delibera 21 Marzo 2018. Nuova Linea Ferroviaria Torino-Lione—Sezione Internazionale—Parte Comune Italo-Francese. Sezione Transfrontaliera. Parte in Territorio Italiano. Approvazione Progetto di Variante in Ottemperanza alla Prescrizione 235 della Delibera CIPE 19 del 2015. (CUP: C11J05000030001). (Delibera n. 30/2018). G.U.R.I. 2018, Serie Generale n. 185, 34–36. Available online: www.gazzettaufficiale.it/eli/id/2018/08/10/18A05313/sg (accessed on 10 December 2023).
- Bas, Y.; Bas, D.; Julien, J.-F. Tadarida: A Toolbox for Animal Detection on Acoustic Recordings. J. Open Res. Softw. 2017, 5, 6. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats. In Species Identification, Study of Their Habitats and Foraging Behaviour, 2nd ed.; (Inventaires & biodiversité series); Biotope éditions: Méze, France; Muséum National d’Histoire Naturelle: Paris, France, 2020. [Google Scholar]
- Russ, J. Bat Calls of Britain and Europe: A Guide to Species Identification; Pelagic Publishing: Exeter, UK, 2021. [Google Scholar]
- Middleton, N.; Froud, A.; French, K. Social Calls of the Bats of Britain and Ireland, 2nd ed.; Pelagic Publishing: London, UK, 2022. [Google Scholar]
- Toffoli, R.; Rughetti, M. Bat activity in rice paddies: Organic and conventional farms compared to unmanaged habitat. Agric. Ecosyst. Environ. 2017, 249, 123–129. [Google Scholar] [CrossRef]
- Kerbiriou, C.; Bas, Y.; Le Viol, I.; Lorrilliere, R.; Mougnot, J.; Julien, J.F. Potential of bat pass duration measures for studies of bat activity. Bioacoustics 2018, 28, 177–192. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 December 2023).
- Bivand, R.; Keitt, T.; Rowlingson, B. rgdal: Bindings for the ’Geospatial’ Data Abstraction Library, R Package Version 1.4-8. 2019. Available online: https://CRAN.R-project.org/package=rgdal (accessed on 10 December 2023).
- Bivand, R.; Rundel, C. RGEOS: Interface to Geometry Engine—Open Source (‘GEOS’); R Package Version 0.5-2. 2019. Available online: https://CRAN.R-project.org/package=rgeos (accessed on 10 December 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference; R Package Version 1.47.5. 2023. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 December 2023).
- Mathews, F.; Roche, N.; Aughney, T.; Jones, N.; Day, J.; Baker, J.A.; Langton, S. Barriers and benefits: Implications of artificial night-lighting for the distribution of common bats in Britain and Ireland. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140124. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Ancillotto, L.; Cistrone, L.; Nastasi, C.; Bosso, L.; Smeraldo, S.; Cordero, V.S.; Russo, D. Artificial illumination influences niche segregation in bats. Environ. Pollut. 2021, 284, 117187. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L.; Cistrone, L.; Libralato, N.; Domer, A.; Cohen, S.; Korine, C. Effects of artificial illumination on drinking bats: A field test in forest and desert habitats. Anim. Conserv. 2018, 22, 124–133. [Google Scholar] [CrossRef]
- Slough, B.G.; Reid, D.G.; Schultz, D.S.; Leung, M.C. Little brown bat activity patterns and conservation implications in agricultural landscapes in boreal Yukon, Canada. Ecosphere 2023, 14, e4446. [Google Scholar] [CrossRef]
- Węgiel, A.; Grzywiński, W.; Ciechanowski, M.; Jaros, R.; Kalcounis-Rüppell, M.; Kmiecik, A.; Kmiecik, P.; Węgiel, J. The foraging activity of bats in managed pine forests of different ages. Eur. J. For. Res. 2019, 138, 383–396. [Google Scholar] [CrossRef]
- Desouhant, E.; Gomes, E.; Mondy, N.; Amat, I. Mechanistic, ecological, and evolutionary consequences of artificial light at night for insects: Review and prospective. Èntomol. Exp. Appl. 2019, 167, 37–58. [Google Scholar] [CrossRef]
- Ancillotto, L.; Tomassini, A.; Russo, D. The fancy city life: Kuhl’s pipistrelle, Pipistrellus kuhlii, benefits from urbanisation. Wildl. Res. 2015, 42, 598. [Google Scholar] [CrossRef]
- Polak, T.; Korine, C.; Yair, S.; Holderied, M.W. Differential effects of artificial lighting on flight and foraging behaviour of two sympatric bat species in a desert. J. Zoöl. 2011, 285, 21–27. [Google Scholar] [CrossRef]
- Boston, E.S.M.; Dechmann, D.K.N.; Ruczyński, I. Leisler’s Noctule Nyctalus leisleri (Kuhl, 1817). In Handbook of the Mammals of Europe; Hackländer, K., Zachos, F.E., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2023; Volume Chiroptera, pp. 423–437. [Google Scholar]
- Barré, K.; Vernet, A.; Azam, C.; Le Viol, I.; Dumont, A.; Deana, T.; Vincent, S.; Challéat, S.; Kerbiriou, C. Landscape composition drives the impacts of artificial light at night on insectivorous bats. Environ. Pollut. 2022, 292, 118394. [Google Scholar] [CrossRef] [PubMed]
- Ancillotto, L.; Budinski, I.; Nardone, V.; Di Salvo, I.; Della Corte, M.; Bosso, L.; Conti, P.; Russo, D. What is driving range expansion in a common bat? Hints from thermoregulation and habitat selection. Behav. Process. 2018, 157, 540–546. [Google Scholar] [CrossRef]
- Smeraldo, S.; Bosso, L.; Salinas-Ramos, V.B.; Ancillotto, L.; Sánchez-Cordero, V.; Gazaryan, S.; Russo, D. Generalists yet different: Distributional responses to climate change may vary in opportunistic bat species sharing similar ecological traits. Mammal Rev. 2021, 51, 571–584. [Google Scholar] [CrossRef]
- Todd, V.L.G.; Williamson, L.D. Habitat usage of Daubenton’s bat (Myotis daubentonii), common pipistrelle (Pipistrellus pipistrellus), and soprano pipistrelle (Pipistrellus pygmaeus) in a North Wales upland river catchment. Ecol. Evol. 2019, 9, 4853–4863. [Google Scholar] [CrossRef] [PubMed]
- Mimet, A.; Kerbiriou, C.; Simon, L.; Julien, J.-F.; Raymond, R. Contribution of private gardens to habitat availability, connectivity and conservation of the common pipistrelle in Paris. Landsc. Urban Plan. 2020, 193, 103671. [Google Scholar] [CrossRef]
- Mendes, E.S.; Fonseca, C.; Marques, S.F.; Maia, D.M.S.; Pereira, M.J.R. Bat richness and activity in heterogeneous landscapes: Guild-specific and scale-dependent? Landsc. Ecol. 2016, 32, 295–311. [Google Scholar] [CrossRef]
- Degen, T.; Mitesser, O.; Perkin, E.K.; Weiß, N.; Oehlert, M.; Mattig, E.; Hölker, F. Street lighting: Sex-independent impacts on moth movement. J. Anim. Ecol. 2016, 85, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Eisenbeis, G. Artificial night lighting and insects: Attraction of insects to streetlamps in a rural setting in Germany. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006; pp. 281–304. [Google Scholar]
- Eisenbeis, G.; Hänel, A. Light pollution and the impact of artificial night lighting on insects. In Ecology of Cities and Towns; McDonnell, M.J., Hahs, A.H., Breuste, J.H., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 243–263. [Google Scholar]
- Kleinbaum, D.G.; Kupper, L.L.; Nizam, A.; Rosenberg, E.S. Applied Regression Analysis and Other Multivariable Methods; Cengage Learning: Boston, MA, USA, 2013. [Google Scholar]
- Azam, C.; Le Viol, I.; Bas, Y.; Zissis, G.; Vernet, A.; Julien, J.-F.; Kerbiriou, C. Evidence for distance and illuminance thresholds in the effects of artificial lighting on bat activity. Landsc. Urban Plan. 2018, 175, 123–135. [Google Scholar] [CrossRef]
- Voigt, C.C.; Azam, C.; Dekker, J.; Ferguson, J.; Fritze, M.; Gazaryan, S.; Hölker, F.; Jones, G.; Leader, N.; Lewanzik, D.; et al. EUROBATS Publication Series No. 8. Guidelines for Consideration of Bats in Lighting Projects; UNEP/EUROBATS Secretariat: Bonn, Germany, 2018. [Google Scholar]
- Barova, S.; Streit, A. Action Plan for the Conservation of All Bat Species in the European Union 2019–2024; European Commission, UNEP Eurobats: Bruxelles, Belgium, 2018. [Google Scholar]
- Straka, T.M.; Wolf, M.; Gras, P.; Buchholz, S.; Voigt, C.C. Tree Cover Mediates the Effect of Artificial Light on Urban Bats. Front. Ecol. Evol. 2019, 7, 91. [Google Scholar] [CrossRef]
- Laforge, A.; Pauwels, J.; Faure, B.; Bas, Y.; Kerbiriou, C.; Fonderflick, J.; Besnard, A. Reducing light pollution improves connectivity for bats in urban landscapes. Landsc. Ecol. 2019, 34, 793–809. [Google Scholar] [CrossRef]
- Cole, M.; DeJohn, M.; McClure, L.; Rogers, S.M. Led Lighting. Minimizing Ecological Impact without Compromising Human Safety. In Proceedings of the 2021 IEEE IAS Petroleum and Chemical Industry Technical Conference (PCIC), San Antonio, TX, USA, 13–16 September 2021. [Google Scholar]
- Andraši, P.; Radišić, T.; Muštra, M.; Ivošević, J. Night-time Detection of UAVs using Thermal Infrared Camera. Transp. Res. Procedia 2017, 28, 183–190. [Google Scholar] [CrossRef]
- Zeale, M.R.K.; Stone, E.L.; Zeale, E.; Browne, W.J.; Harris, S.; Jones, G. Experimentally manipulating light spectra reveals the importance of dark corridors for commuting bats. Glob. Chang. Biol. 2018, 24, 5909–5918. [Google Scholar] [CrossRef]
- Kerbiriou, C.; Barré, K.; Mariton, L.; Pauwels, J.; Zissis, G.; Robert, A.; Le Viol, I. Switching LPS to LED Streetlight May Dramatically Reduce Activity and Foraging of Bats. Diversity 2020, 12, 165. [Google Scholar] [CrossRef]
- Bjelajac, D.; Đerčan, B.; Kovačić, S. Dark skies and dark screens as a precondition for astronomy tourism and general well-being. Inf. Technol. Tour. 2020, 23, 19–43. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Species | Elevation | Maximum Light Intensity | Distance from Construction Site | Total Built-Up Area | Total Forest Area | Total Area of Meadows and Pastures | Average Temperature | Julian Day | Hours after Sunset | Variance Explained (R2m) | Variance Explained (R2c) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Common species | |||||||||||
| Hsav | * | ** | NS | *** | ** | 0.35 | 0.49 | ||||
| + | + | + |  | ||||||||
| Nlei | ** | NS | NS | NS | NS | NS | *** | 0.25 | 0.47 | ||
| + | - | ||||||||||
| Pkuh and Pnat | NS | ** | * | NS | NS | NS | *** | *** | 0.38 | 0.60 | |
| + | + | + |  | ||||||||
| Ppip | NS | NS | NS | NS | *** | 0.22 | 0.54 | ||||
 | |||||||||||
| Rare species | |||||||||||
| Bbar | NS | * | NS | 0.54 | 0.59 | ||||||
| - | |||||||||||
| S-Myo | *** | * | * | 0.48 | 0.73 | ||||||
| - | - | + | |||||||||
| Plespp | ** | ** | * | ** | ** | *** | 0.56 | 0.63 | |||
| + | + | + | + | - | + | ||||||
| Tten | *** | 0.15 | 0.15 | ||||||||
| + | |||||||||||
| Species richness | |||||||||||
| SR | * | * | 0.09 | 0.09 | |||||||
| - | + | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gili, F.; Fassone, C.; Rolando, A.; Bertolino, S. In the Spotlight: Bat Activity Shifts in Response to Intense Lighting of a Large Railway Construction Site. Sustainability 2024, 16, 2337. https://doi.org/10.3390/su16062337
Gili F, Fassone C, Rolando A, Bertolino S. In the Spotlight: Bat Activity Shifts in Response to Intense Lighting of a Large Railway Construction Site. Sustainability. 2024; 16(6):2337. https://doi.org/10.3390/su16062337
Chicago/Turabian StyleGili, Fabrizio, Carlotta Fassone, Antonio Rolando, and Sandro Bertolino. 2024. "In the Spotlight: Bat Activity Shifts in Response to Intense Lighting of a Large Railway Construction Site" Sustainability 16, no. 6: 2337. https://doi.org/10.3390/su16062337
APA StyleGili, F., Fassone, C., Rolando, A., & Bertolino, S. (2024). In the Spotlight: Bat Activity Shifts in Response to Intense Lighting of a Large Railway Construction Site. Sustainability, 16(6), 2337. https://doi.org/10.3390/su16062337