
Influence of Soil Moisture in Semi-Fixed Sand Dunes of the Tengger Desert, China, Based on PLS-SEM and SHAP Models


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Area
2.2. Field Study Design
2.3. Data Analysis Methods
2.3.1. Division of Soil Moisture
2.3.2. Calculate the Topographic Factors
2.3.3. Partial Least Squares Structural Equation Modeling Analysis
2.3.4. Machine Learning Algorithms
3. Results and Analysis
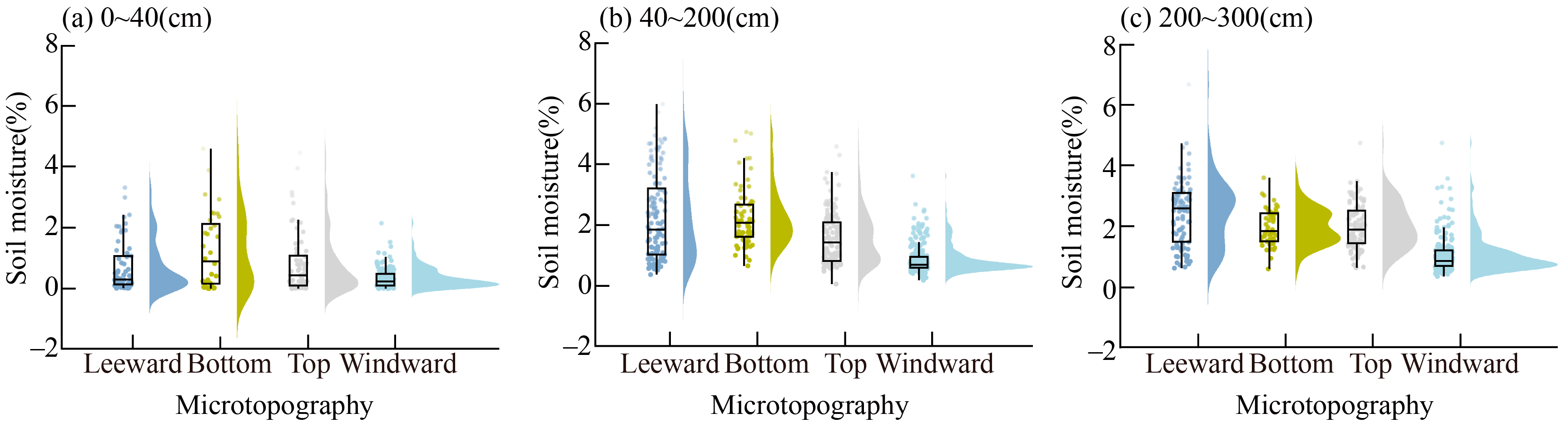
3.1. Patterns of Soil Moisture Distribution among Four Microtopography Types
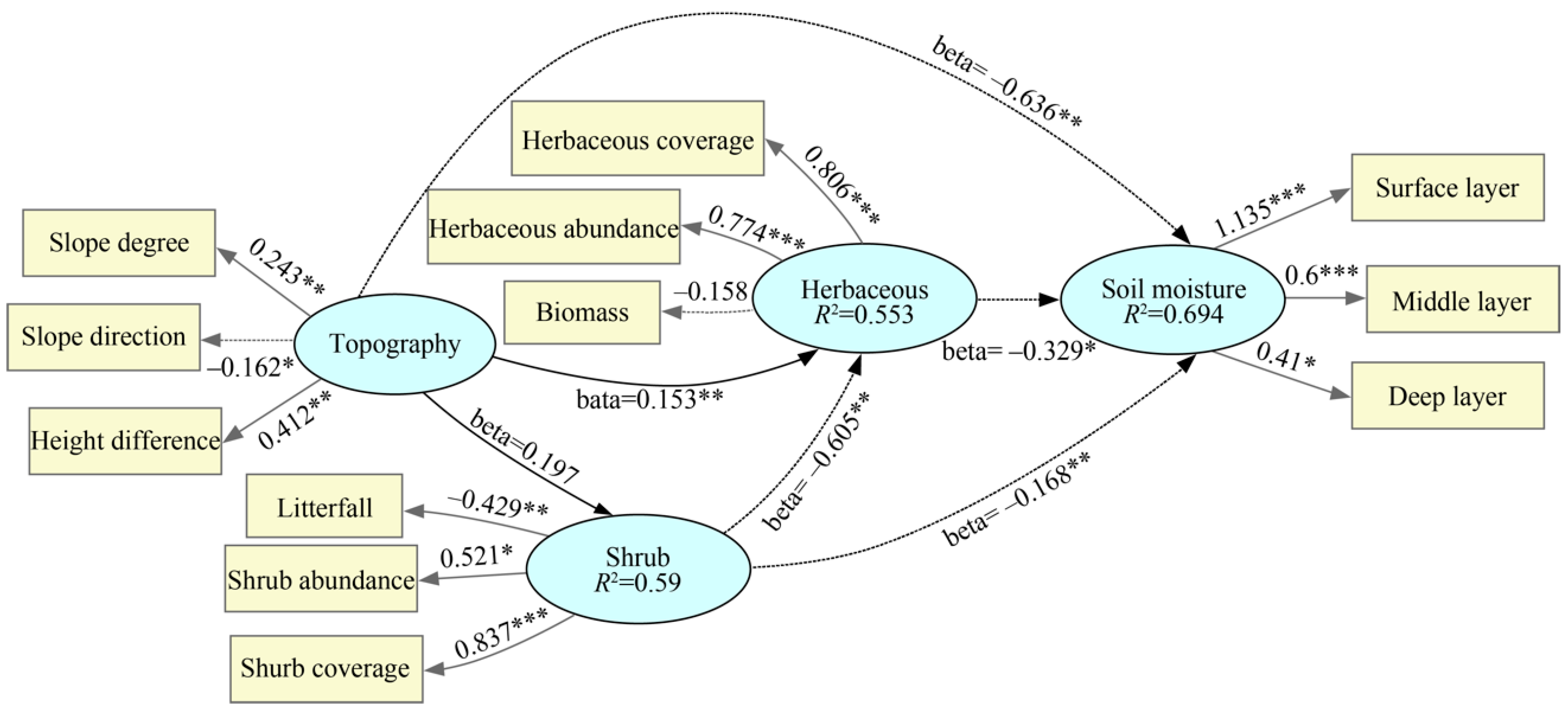
3.2. Influence Pathways of Topographical and Vegetative Factors
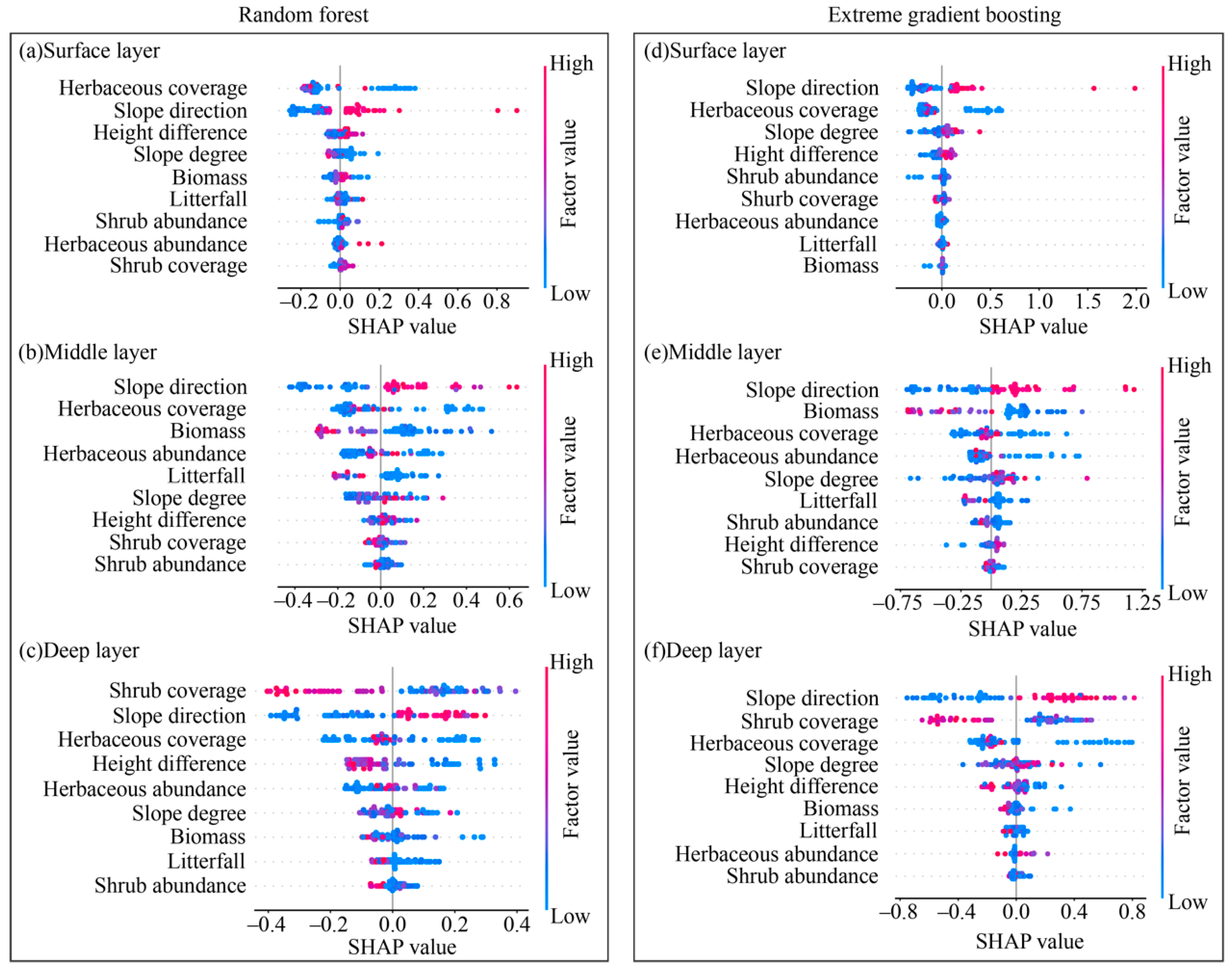
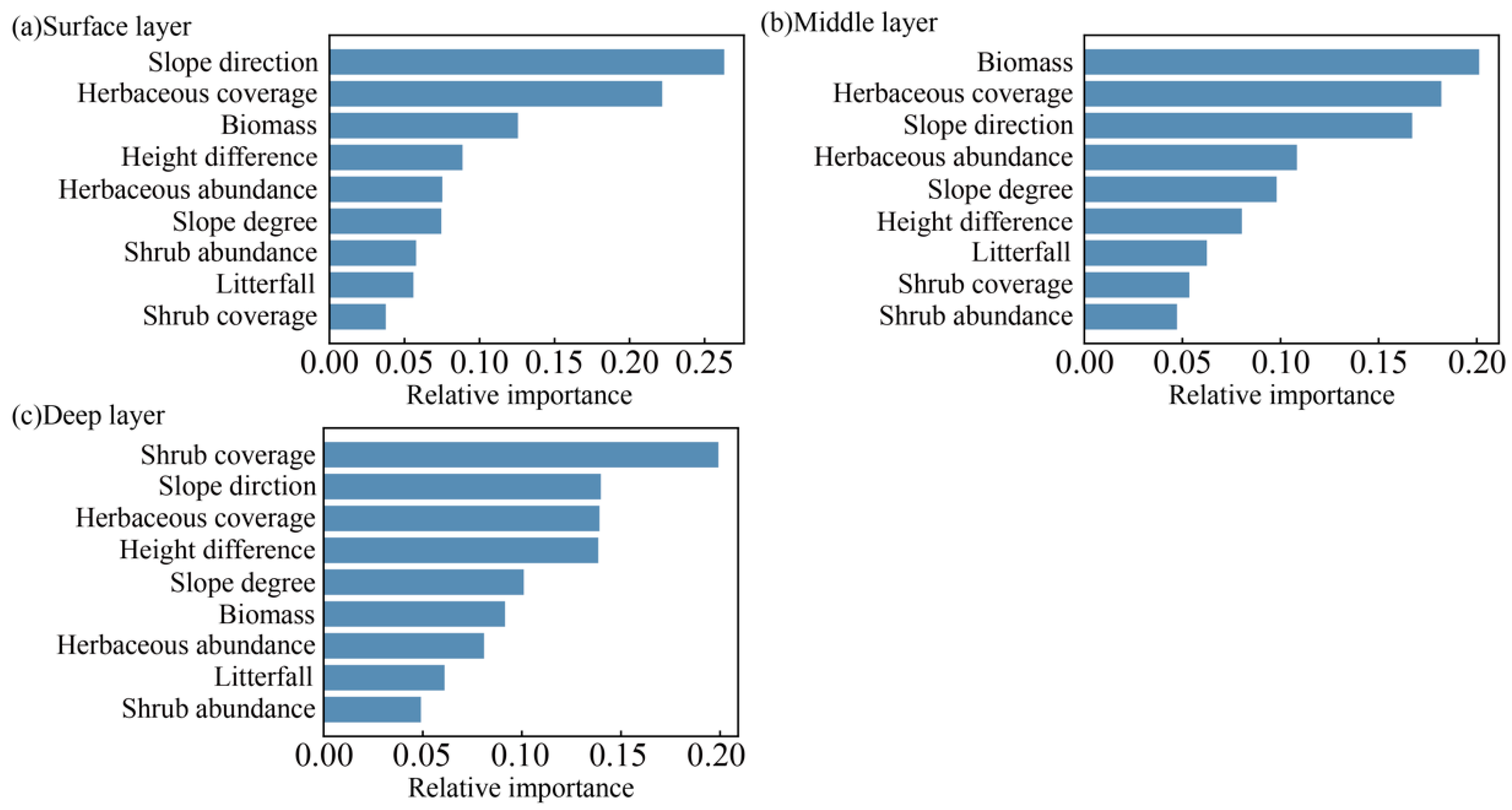
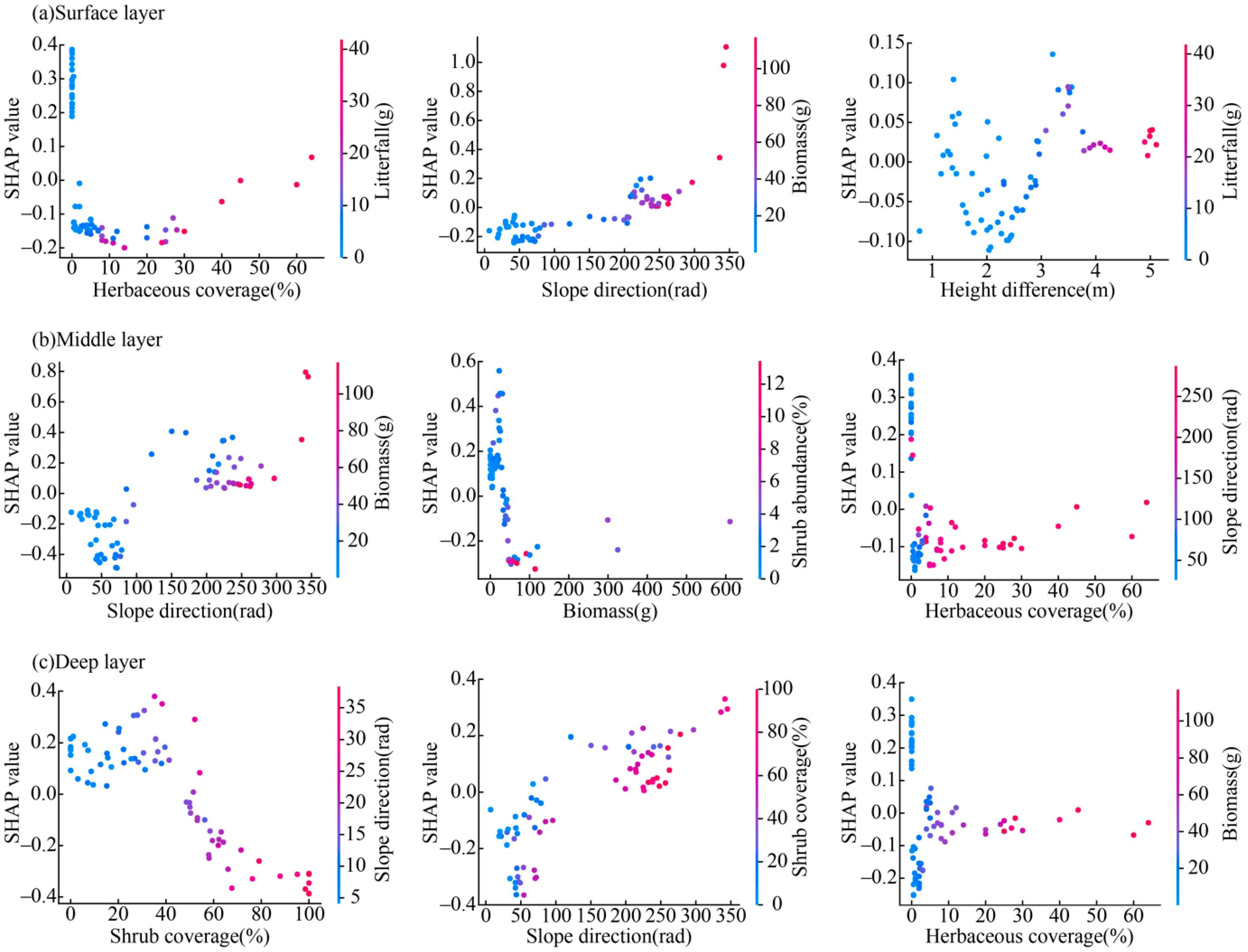
3.3. Analysis of Factors Influencing Soil Moisture Based on the SHAP Model
4. Discussion
4.1. Examination of Soil Moisture Distribution Patterns among Four Microtopography Types
4.2. Analysis of the Direct and Indirect Effects of Topography–Vegetation Factors
4.3. The Mechanism of Topography–Vegetation Factors Affecting Soil Moisture at Different Depths
4.4. Factors Influencing Soil Moisture Dynamics in Desert Ecosystems and the Potential Effects of Climate Change
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, Q.; Ma, H.; Jiang, X.; Wang, X.; Cao, S. What has caused desertification in China? Sci. Rep. 2015, 5, 15998. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, J. Characteristics of precipitation and desertification in arid and semi–arid regions of China. Environ. Sci. Pollut. Res. 2021, 28, 19800–19810. [Google Scholar]
- Zhao, H.; Jiang, H. Dynamics of fixed and semi-fixed dunes in response to vegetation cover and aeolian processes. Geomorphology 2019, 333, 27–38. [Google Scholar]
- Rasheed, M.W.; Tang, J.; Sarwar, A.; Shah, S.; Saddique, N.; Khan, M.U.; Khan, M.I.; Nawaz, S.; Shamshiri, R.R.; Aziz, M.; et al. Soil moisture measuring techniques and factors affecting the moisture dynamics: A comprehensive review. Sustainability 2022, 14, 11538. [Google Scholar] [CrossRef]
- Bar Kutiel, P.; Dorman, M. The importance of annual plants and multi–scalar analysis for understanding Coastal Dune stabilization process in the Mediterranean. Appl. Sci. 2021, 11, 2821. [Google Scholar] [CrossRef]
- Zhou, W.; Li, C.; Wang, S.; Ren, Z.; Stringer, L.C. Effects of vegetation restoration on soil properties and vegetation attributes in the arid and semi–arid regions of China. J. Environ. Manag. 2023, 343, 118186. [Google Scholar] [CrossRef]
- Guo, X.; Fu, Q.; Hang, Y.; Lu, H.; Gao, F.; Si, J. Spatial variability of soil moisture in relation to land use types and topographic features on hillslopes in the black soil (mollisols) area of northeast China. Sustainability 2020, 12, 3552. [Google Scholar] [CrossRef]
- Svoray, T.; Karnieli, A. Rainfall, topography and primary production relationships in a semiarid ecosystem. Ecohydrology 2011, 4, 56–66. [Google Scholar] [CrossRef]
- Pockman, W.T.; Small, E. The influence of spatial patterns of soil moisture on the grass and shrub responses to a summer rainstorm in a Chihuahuan Desert ecotone. Ecosystems 2010, 13, 511–525. [Google Scholar] [CrossRef]
- Liu, C.; Wang, F.G.; Xue, Q.; Li, L.; Wang, Z. Pattern formation of a spatial vegetation system with root hydrotropism. Appl. Math. Comput. 2022, 420, 126913. [Google Scholar] [CrossRef]
- Zhang, G.; Biradar, C.M.; Xiao, X.; Dong, J.; Zhou, Y.; Qin, Y.; Zhang, Y.; Liu, F.; Ding, M.; Thomas, R.J. Exacerbated grassland degradation and desertification in Central Asia during 2000–2014. Ecol. Appl. 2018, 28, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Holdo, R.M.; Nippert, J.B. Linking resource-and disturbance-based models to explain tree–grass coexistence in savannas. New Phytol. 2023, 237, 1966–1979. [Google Scholar] [CrossRef] [PubMed]
- Demir, G.; Michalzik, B.; Filipzik, J.; Metzger, J.C.; Hildebrandt, A. Spatial variation of grassland canopy affects soil wetting patterns and preferential flow. Hydrol. Process. 2022, 36, e14760. [Google Scholar] [CrossRef]
- Cai, G.; Vanderborght, J.; Couvreur, V.; Mboh, C.M.; Vereecken, H. Parameterization of root water uptake models considering dynamic root distributions and water uptake compensation. Vadose Zone J. 2018, 17, 1–21. [Google Scholar] [CrossRef]
- Butz, P.; Hölscher, D.; Cueva, E.; Graefe, S. Tree water use patterns as influenced by phenology in a dry forest of Southern Ecuador. Front. Plant Sci. 2018, 9, 945. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, S.C.; Poot, P.; Veneklaas, E.J. Shallow soils negatively affect water relations and photosynthesis in two semi–arid Eucalyptus species. Environ. Exp. Bot. 2018, 155, 239–250. [Google Scholar] [CrossRef]
- Özkan, U.; Gökbulak, F. Effect of vegetation change from forest to herbaceous vegetation cover on soil moisture and temperature regimes and soil water chemistry. Catena 2017, 149, 158–166. [Google Scholar] [CrossRef]
- Wang, X.P.; Zhang, Y.F.; Wang, Z.N.; Pan, Y.X.; Hu, R.; Li, X.J.; Zhang, H. Influence of shrub canopy morphology and rainfall characteristics on stemflow within a revegetated sand dune in the Tengger Desert, NW China. Hydrol. Process. 2013, 27, 1501–1509. [Google Scholar] [CrossRef]
- Guderle, M.; Bachmann, D.; Milcu, A.; Gockele, A.; Bechmann, M.; Fischer, C.; Roscher, C.; Landais, D.; Ravel, O.; Devidal, S.; et al. Dynamic niche partitioning in root water uptake facilitates efficient water use in more diverse grassland plant communities. Funct. Ecol. 2018, 32, 214–227. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Mommer, L.; Sabatini, F.M.; Bruelheide, H.; Kuyper, T.W.; McCormack, M.L.; Bergmann, J.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Iversen, C.M.; et al. Root traits explain plant species distributions along climatic gradients yet challenge the nature of ecological trade–offs. Nat. Ecol. Evol. 2021, 5, 1123–1134. [Google Scholar] [CrossRef]
- Moeslund, J.E.; Arge, L.; Bøcher, P.K.; Dalgaard, T.; Svenning, J.C. Topography as a driver of local terrestrial vascular plant diversity patterns. Nord. J. Bot. 2013, 31, 129–144. [Google Scholar] [CrossRef]
- Schaefer, M.L.; Bogacki, W.; Lopez Caceres, M.L.; Kirschbauer, L.; Kato, C.; Kikuchi, S.I. Influence of Slope aspect and vegetation on the soil moisture response to snowmelt in the German Alps. Hydrology 2024, 11, 101. [Google Scholar] [CrossRef]
- Caviedes-Voullième, D.; Ahmadinia, E.; Hinz, C. Interactions of microtopography, slope and infiltration cause complex rainfall-runoff behavior at the hillslope scale for single rainfall events. Water Resour. Res. 2021, 57, e2020WR028127. [Google Scholar] [CrossRef]
- Yang, J.; El-Kassaby, Y.A.; Guan, W. The effect of slope aspect on vegetation attributes in a mountainous dry valley, Southwest China. Sci. Rep. 2020, 10, 16465. [Google Scholar] [CrossRef] [PubMed]
- Biederman, L.A.; Whisenant, S.G. Using mounds to create microtopography alters plant community development early in restoration. Restor. Ecol. 2011, 19, 53–61. [Google Scholar] [CrossRef]
- Brocca, L.; Ciabatta, L.; Massari, C.; Camici, S.; Tarpanelli, A. Soil moisture for hydrological applications: Open questions and new opportunities. Water 2017, 9, 140. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, Z.; Huang, Z.; López-Vicente, M.; Wu, G.-L. Influence of soil moisture and plant roots on the soil infiltration capacity at different stages in arid grasslands of China. Catena 2019, 182, 104147. [Google Scholar] [CrossRef]
- Srivastava, A.; Saco, P.M.; Rodriguez, J.F.; Kumari, N.; Chun, K.P.; Yetemen, O. The role of landscape morphology on soil moisture variability in semi-arid ecosystems. Hydrol. Process. 2021, 35, e13990. [Google Scholar] [CrossRef]
- Caracciolo, D.; Istanbulluoglu, E.; Noto, L.V.; Collins, S.L. Mechanisms of shrub encroachment into northern Chihuahuan Desert grasslands and impacts of climate change investigated using a cellular automata model. Adv. Water Resour. 2016, 91, 46–62. [Google Scholar] [CrossRef]
- Lawrence, J.E.; Hornberger, G.M. Soil moisture variability across climate zones. Geophys. Res. Lett. 2007, 34, 5. [Google Scholar] [CrossRef]
- Teuling, A.J.; Troch, P.A. Improved understanding of soil moisture variability dynamics. Geophys. Res. Lett. 2005, 32, L05404. [Google Scholar] [CrossRef]
- Jiang, L.M.; Sattar, K.; Lü, G.H.; Hu, D.; Zhang, J.; Yang, X.D. Different contributions of plant diversity and soil properties to the community stability in the arid desert ecosystem. Front. Plant Sci. 2022, 13, 969852. [Google Scholar] [CrossRef]
- Christin, S.; Hervet, É.; Lecomte, N. Applications for deep learning in ecology. Methods Ecol. Evol. 2019, 10, 1632–1644. [Google Scholar] [CrossRef]
- Olden, J.D.; Lawler, J.J.; Poff, N.L. Machine learning methods without tears: A primer for ecologists. Q. Rev. Biol. 2008, 83, 171–193. [Google Scholar] [CrossRef] [PubMed]
- Karpatne, A.; Ebert-Uphoff, I.; Ravela, S.; Babaie, H.A.; Kumar, V. Machine learning for the geosciences: Challenges and opportunities. IEEE Trans. Knowl. Data Eng. 2018, 31, 1544–1554. [Google Scholar] [CrossRef]
- Ryo, M.; Angelov, B.; Mammola, S.; Kass, J.M.; Benito, B.M.; Hartig, F. Explainable artificial intelligence enhances the ecological interpretability of black-box species distribution models. Ecography 2021, 44, 199–205. [Google Scholar] [CrossRef]
- Lundberg, S.M.; Lee, S.I. A Unified Approach to Interpreting Model Predictions. In Proceedings of the 31st International Conference on Neural Information Processing Systems, Long Beach, CA, USA, 4 December 2017; Volume 30. [Google Scholar]
- Zhang, S.; Wang, H.; Zhang, D.; Wu, X. Relationship between soil moisture and topography–vegetation factors on three types of sand dunes and four types of microgeomorphology in the southeastern margin of Tengger Desert in northern China. J. Gansu Agric. Univ. 2023, 58, 160–168+179. [Google Scholar]
- Li, X.; Zhang, Z.; Tan, H.; Gao, H.; Liu, L.; Wang, X. Ecological restoration and recovery in the wind–blown sand hazard areas of northern China: Relationship between soil water and carrying capacity for vegetation in the Tengger Desert. Sci. China Life Sci. 2014, 44, 257–266. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, D.; Zhang, Z. Relationship between soil moisture content and topography–vegetation factors in different types of dunes in the Tengger Desert. Arid Land Geogr. 2022, 45, 1570–1578. [Google Scholar]
- Liu, L.; Bai, Y.; She, W.; Qiao, Y.; Qin, S.; Zhang, Y. A nurse shrub species helps associated herbaceous plants by preventing shade–induced evaporation in a desert ecosystem. Land Degrad. Dev. 2021, 32, 1796–1808. [Google Scholar] [CrossRef]
- Liu, R.; Li, Y.; Wang, Y.; Ma, J.; Cieraad, E. Variation of water use efficiency across seasons and years: Different role of herbaceous plants in desert ecosystem. Sci. Total Environ. 2019, 647, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.; Liu, L.; Jia, R.; Zhang, J.; Wang, T. Distribution, biomass, and dynamics of roots in a revegetated stand of Caragana korshinskii in the Tengger Desert, northwestern China. J. Plant Res. 2009, 122, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Fijałkowska, A. Analysis of the influence of DTM source data on the LS factors of the soil water erosion model values with the use of GIS technology. Remote Sens. 2021, 13, 678. [Google Scholar] [CrossRef]
- Hair, J.F., Jr.; Hult, G.T.; Ringle, C.M.; Sarstedt, M.; Danks, N.P.; Ray, S. Partial Least Squares Structural Equation Modeling (PLS–SEM) Using R: A Workbook; Springer: Cham, Switzerland, 2021; ISBN 978-3-030-80518-0. [Google Scholar]
- Hair, J.F., Jr.; Matthews, L.M.; Matthews, R.L.; Sarstedt, M. PLS–SEM or CB–SEM: Updated guidelines on which method to use. Int. J. Multivar. Data Anal. 2017, 1, 107–123. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Boulesteix, A.L.; Janitza, S.; Kruppa, J.; König, I.R. Overview of random forest methodology and practical guidance with emphasis on computational biology and bioinformatics. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2012, 2, 493–507. [Google Scholar] [CrossRef]
- Iban, M.C.; Bilgilioglu, S.S. Snow avalanche susceptibility mapping using novel tree–based machine learning algorithms (XGBoost, NGBoost, and LightGBM) with eXplainable Artificial Intelligence (XAI) approach. Stoch. Environ. Res. Risk Assess. 2023, 37, 2243–2270. [Google Scholar] [CrossRef]
- Gupta, A.; Gowda, S.; Tiwari, A.; Gupta, A.K. XGBoost–SHAP framework for asphalt pavement condition evaluation. Constr. Build. Mater. 2024, 426, 136182. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.M.; Tuleau-Malot, C. Variable selection using random forests. Pattern Recognit. Lett. 2010, 31, 2225–2236. [Google Scholar] [CrossRef]
- Altman, N.; Krzywinski, M. Ensemble methods: Bagging and random forests. Nat. Methods 2017, 14, 933–935. [Google Scholar] [CrossRef]
- Kotsiantis, S.B.; Pintelas, P.E. Combining bagging and boosting. Int. J. Math. Comput. Sci. 2007, 1, 372–381. [Google Scholar]
- González, S.; García, S.; Del Ser, J.; Rokach, L.; Herrera, F. A practical tutorial on bagging and boosting based ensembles for machine learning: Algorithms, software tools, performance study, practical perspectives and opportunities. Inf. Fusion 2020, 64, 205–237. [Google Scholar] [CrossRef]
- Rudin, C. Stop explaining black box machine learning models for high stakes decisions and use interpretable models instead. Nat. Mach. Intell. 2019, 1, 206–215. [Google Scholar] [CrossRef]
- Tavakol, M.; Dennick, R. Making sense of Cronbach’s alpha. Int. J. Med. Educ. 2011, 2, 53. [Google Scholar] [CrossRef] [PubMed]
- Mehmetoglu, M. Partial Least Squares Approach to Structural Equation Modeling for Tourism Research. In Advances in Hospitality and Leisure; Chen, J.S., Ed.; Emerald Group Publishing Limited: Leeds, UK, 2012; Volume 8, pp. 43–61. ISBN 978-1-78052-936-3. [Google Scholar]
- Dijkstra, T.K.; Henseler, J. Linear indices in nonlinear structural equation models: Best fitting proper indices and other composites. Qual. Quant. 2011, 45, 1505–1518. [Google Scholar] [CrossRef]
- Wetzels, M.; Odekerken-Schröder, G.; Van Oppen, C. Using PLS path modeling for assessing hierarchical construct models: Guidelines and empirical illustration. MIS Q. 2009, 33, 177–195. [Google Scholar] [CrossRef]
- Zipper, S.C.; Soylu, M.E.; Booth, E.G.; Loheide, S.P. Untangling the effects of shallow groundwater and soil texture as drivers of subfield-scale yield variability. Water Resour. Res. 2015, 51, 6338–6358. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Z.; Huang, L.; Wang, X. Review of the ecohydrological processes and feedback mechanisms controlling sand–binding vegetation systems in sandy desert regions of China. Chin. Sci. Bull. 2013, 58, 397–410. [Google Scholar] [CrossRef]
- Song, G.; Li, X.; Hui, R. Biological soil crusts increase stability and invasion resistance of desert revegetation communities in northern China. Ecosphere 2020, 11, e03043. [Google Scholar] [CrossRef]
- Chamizo, S.; Canton, Y.; Lázaro, R.; Domingo, F. The role of biological soil crusts in soil moisture dynamics in two semiarid ecosystems with contrasting soil textures. J. Hydrol. 2013, 489, 74–84. [Google Scholar] [CrossRef]
- Meng, Z.; Dang, X.; Gao, Y.; Ren, X.; Ding, Y.; Wang, M. Interactive effects of wind speed, vegetation coverage and soil moisture in controlling wind erosion in a temperate desert steppe, Inner Mongolia of China. J. Arid Land 2018, 10, 534–547. [Google Scholar] [CrossRef]
- John, A.; Olden, J.D.; Oldfather, M.F.; Kling, M.M.; Ackerly, D.D. Topography influences diurnal and seasonal microclimate fluctuations in hilly terrain environments of coastal California. PLoS ONE 2024, 19, e0300378. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. Understanding the role of slope aspect in shaping the vegetation attributes and soil properties in Montane ecosystems. Trop. Ecol. 2018, 59, 417–430. [Google Scholar]
- Wunsch, M.; Betzler, C.; Eberli, G.P.; Lindhorst, S.; Lüdmann, T.; Reijmer, J.J. Sedimentary dynamics and high–frequency sequence stratigraphy of the southwestern slope of Great Bahama Bank. Sediment. Geol. 2018, 363, 96–117. [Google Scholar] [CrossRef]
- Brown, G.; Porembski, S. The maintenance of species diversity by miniature dunes in a sand–depleted Haloxylon salicornicum community in Kuwait. J. Arid Environ. 1997, 37, 461–473. [Google Scholar] [CrossRef]
- Gong, X.; Zhang, H.; Ren, C.; Sun, D.; Yang, J. Optimization allocation of irrigation water resources based on crop water requirement under considering effective precipitation and uncertainty. Agric. Water Manag. 2020, 239, 106264. [Google Scholar] [CrossRef]
- Liu, X.; Lu, K.; Li, P.; Xu, G.; Cheng, S.; Bai, L.; Wei, F. Research and simulation of soil water infiltration on slope under different rain–fall conditions. J. Arid Land Resour. Environ. 2018, 32, 114–118. [Google Scholar]
- Cheng, L.; Liu, W.; Li, Z. Soil water in deep layers under different land use patterns on the Loess Tableland. Acta Ecol. Sin. 2014, 34, 1975–1983. [Google Scholar]
- Zheng, L.; Wang, X.; Li, D.; Xu, G.; Guo, Y. Spatial heterogeneity of vegetation extent and the response to water level fluctuations and micro–topography in Poyang Lake, China. Ecol. Indic. 2021, 124, 107420. [Google Scholar] [CrossRef]
- Liu, Y.; Du, J.; Xu, X.; Kardol, P.; Hu, D. Microtopography–induced ecohydrological effects alter plant community structure. Geoderma 2020, 362, 114119. [Google Scholar] [CrossRef]
- Mata-González, R.; Averett, J.P.; Abdallah, M.A.; Martin, D.W. Variations in groundwater level and microtopography influence desert plant communities in shallow aquifer areas. Environ. Manag. 2022, 69, 45–60. [Google Scholar] [CrossRef] [PubMed]
- De Falco, N.; Tal-Berger, R.; Hjazin, A.; Yizhaq, H.; Stavi, I.; Rachmilevitch, S. Geodiversity impacts plant community structure in a semi–arid region. Sci. Rep. 2021, 11, 15259. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Deng, L.; Huang, C.; Chen, Y.; Peng, C. Effects of vegetation restoration on soil nutrients, plant diversity, and its spatiotemporal heterogeneity in a desert–oasis ecotone. Land Degrad. Dev. 2021, 32, 670–683. [Google Scholar] [CrossRef]
- Fan, B.; Tao, W.; Qin, G.; Hopkins, I.; Zhang, Y.; Wang, Q.; Lin, H.; Guo, L. Soil micro–climate variation in relation to slope aspect, position, and curvature in a forested catchment. Agric. For. Meteorol. 2020, 290, 107999. [Google Scholar] [CrossRef]
- Rita, A.; Bonanomi, G.; Allevato, E.; Borghetti, M.; Cesarano, G.; Mogavero, V.; Rossi, S.; Saulino, L.; Zotti, M.; Saracino, A. Topography modulates near–ground microclimate in the Mediterranean Fagus sylvatica treeline. Sci. Rep. 2021, 11, 8122. [Google Scholar] [CrossRef]
- Zhou, X.; Ke, T.; Li, S.; Deng, S.; An, X.; Ma, X.; De Philippis, R.; Chen, L. Induced biological soil crusts and soil properties varied between slope aspect, slope gradient and plant canopy in the Hobq desert of China. Catena 2020, 190, 104559. [Google Scholar] [CrossRef]
- Ramos, M.B.; Diniz, F.C.; de Almeida, H.A.; de Almeida, G.R.; Pinto, A.S.; Meave, J.A.; Lopes, S.D. The role of edaphic factors on plant species richness and diversity along altitudinal gradients in the Brazilian semi–arid region. J. Trop. Ecol. 2020, 36, 199–212. [Google Scholar] [CrossRef]
- Geng, Q.; Arif, M.; Yuan, Z.; Zheng, J.; He, X.; Ding, D.; Yin, F.; Li, C. Plant species composition and diversity along successional gradients in arid and semi–arid regions of China. For. Ecol. Manag. 2022, 524, 120542. [Google Scholar]
- Zhao, L.; Fang, Q.; Yang, Y.; Yang, H.; Yang, T.; Zheng, H. Stemflow contributions to soil erosion around the stem base under simulated maize–planted and rainfall conditions. Agric. For. Meteorol. 2020, 281, 107814. [Google Scholar] [CrossRef]
- Peters, R.; Walther, M.; Lovelock, C.; Jiang, J.; Berger, U. The interplay between vegetation and water in mangroves: New perspectives for mangrove stand modelling and ecological research. Wetl. Ecol. Manag. 2020, 28, 697–712. [Google Scholar] [CrossRef]
- Li, B.B.; Li, P.P.; Zhang, W.T.; Ji, J.Y.; Liu, G.B.; Xu, M.X. Deep soil moisture limits the sustainable vegetation restoration in arid and semi–arid Loess Plateau. Geoderma 2021, 399, 115122. [Google Scholar] [CrossRef]
- Na, L.; Na, R.; Bao, Y.; Zhang, J. Time–lagged correlation between soil moisture and intra–annual dynamics of vegetation on the Mongolian plateau. Remote Sens. 2021, 13, 1527. [Google Scholar] [CrossRef]
- Najafifar, A.; Hosseinzadeh, J.; Karamshahi, A. The role of hillshade, aspect, and toposhape in the woodland dieback of arid and semi–arid ecosystems: A case study in Zagros woodlands of Ilam province, Iran. J. Landsc. Ecol. 2019, 12, 79–91. [Google Scholar] [CrossRef]
- Jakšić, S.; Ninkov, J.; Milić, S.; Vasin, J.; Živanov, M.; Jakšić, D.; Komlen, V. Influence of slope gradient and aspect on soil organic carbon content in the region of Niš, Serbia. Sustainability 2021, 13, 8332. [Google Scholar] [CrossRef]
- Yang, L.; Wei, W.; Chen, L.; Jia, F.; Mo, B. Spatial variations of shallow and deep soil moisture in the semi–arid Loess Plateau, China. Hydrol. Earth Syst. Sci. 2012, 16, 3199–3217. [Google Scholar] [CrossRef]
- Thomas, A.; Yadav, B.K.; Šimůnek, J. Root water uptake under heterogeneous soil moisture conditions: An experimental study for unraveling compensatory root water uptake and hydraulic redistribution. Plant Soil 2020, 457, 421–435. [Google Scholar] [CrossRef]
- D’Odorico, P.; Caylor, K.; Okin, G.S.; Scanlon, T.M. On soil moisture–vegetation feedbacks and their possible effects on the dynamics of dryland ecosystems. J. Geophys. Res. Biogeosci. 2007, 112, G04010. [Google Scholar] [CrossRef]
- Cheng, M.; Jiao, X.; Liu, Y.; Shao, M.; Yu, X.; Bai, Y.; Wang, Z.; Wang, S.; Tuohuti, N.; Liu, S.; et al. Estimation of soil moisture content under high maize canopy coverage from UAV multimodal data and machine learning. Agric. Water Manag. 2022, 264, 107530. [Google Scholar] [CrossRef]
- Cheng, R.R.; Chen, Q.W.; Zhang, J.G.; Shi, W.Y.; Li, G.; Du, S. Soil moisture variations in response to precipitation in different vegetation types: A multi-year study in the loess hilly region in China. Ecohydrology 2020, 13, e2196. [Google Scholar] [CrossRef]
- Chen, N.; Zhang, Y.; Zhao, C. On the importance of stemflow to the woody plants in drylands: Individual vs. ecosystem scales. J. Hydrol. 2021, 601, 126591. [Google Scholar] [CrossRef]
- Yue, K.; De Frenne, P.; Fornara, D.A.; Van Meerbeek, K.; Li, W.; Peng, X.; Ni, X.; Peng, Y.; Wu, F.; Yang, Y.; et al. Global patterns and drivers of rainfall partitioning by trees and shrubs. Glob. Chang. Biol. 2021, 27, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Ankenbauer, K.J.; Loheide, S.P. The effects of soil organic matter on soil water retention and plant water use in a meadow of the Sierra Nevada, CA. Hydrol. Process. 2017, 31, 891–901. [Google Scholar] [CrossRef]
- D’odorico, P.; He, Y.; Collins, S.; De Wekker, S.F.; Engel, V.; Fuentes, J.D. Vegetation–microclimate feedbacks in woodland–grassland ecotones. Glob. Ecol. Biogeogr. 2013, 22, 364–379. [Google Scholar] [CrossRef]
- Jaroszynska, F.; Althuizen, I.; Halbritter, A.H.; Klanderud, K.; Lee, H.; Telford, R.J.; Vandvik, V. Bryophytes dominate plant regulation of soil microclimate in alpine grasslands. Oikos 2023, 12, e10091. [Google Scholar] [CrossRef]
- Sharma, S.; Carlson, J.D.; Krueger, E.S.; Engle, D.M.; Twidwell, D.; Fuhlendorf, S.D.; Patrignani, A.; Feng, L.; Ochsner, T.E. Soil moisture as an indicator of growing–season herbaceous fuel moisture and curing rate in grasslands. Int. J. Wildland Fire 2020, 30, 57–69. [Google Scholar] [CrossRef]
- Kim, Y.; Eltahir, E.A. Role of topography in facilitating coexistence of trees and grasses within savannas. Water Resour. Res. 2004, 40, W07505. [Google Scholar] [CrossRef]
- Rodríguez-Moreno, V.M.; Bullock, S.H. Vegetation response to rainfall pulses in the Sonoran Desert as modelled through remotely sensed imageries. Int. J. Climatol. 2014, 34, 3967–3976. [Google Scholar] [CrossRef]
- Nano, C.E.; Pavey, C.R. Refining the ‘pulse-reserve’model for arid central A ustralia: Seasonal rainfall, soil moisture and plant productivity in sand ridge and stony plain habitats of the Simpson Desert. Austral Ecol. 2013, 38, 741–753. [Google Scholar] [CrossRef]
- Hassan, W.H. Climate change impact on groundwater recharge of Ummer Radhuma unconfined aquifer Western Desert, Iraq. Int. J. Hydrol. Sci. Technol. 2020, 10, 392–412. [Google Scholar] [CrossRef]
- Stringer, L.C.; Mirzabaev, A.; Benjaminsen, T.A.; Harris, R.M.; Jafari, M.; Lissner, T.K.; Stevens, N.; Der Pahlen, C.T.-V. Climate change impacts on water security in global drylands. One Earth 2021, 4, 851–864. [Google Scholar] [CrossRef]
- Zuo, X.; Cheng, H.; Zhao, S.; Yue, P.; Liu, X.; Wang, S.; Liu, L.; Xu, C.; Luo, W.; Knops, J.M.; et al. Observational and experimental evidence for the effect of altered precipitation on desert and steppe communities. Glob. Ecol. Conserv. 2020, 21, e00864. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Factors | Units |
|---|---|---|
| Topographic factors | Slope degree | rad |
| Slope direction | rad | |
| Height difference | m | |
| Shrub factors | Shrub coverage | % |
| Shrub abundance | % | |
| Litterfall | g | |
| Herbaceous factors | Herbaceous coverage | % |
| Herbaceous abundance | % | |
| Biomass | g | |
| Soil moisture | Surface layer | % |
| Middle layer | % | |
| Deep layer | % |
| Microtopography | 0~40 cm | 40~200 cm | 200~300 cm | |||
|---|---|---|---|---|---|---|
| Average Soil Moisture (%) | Coefficient of Variation | Average Soil Moisture (%) | Coefficient of Variation | Average Soil Moisture (%) | Coefficient of Variation | |
| Leeward | 0.71 | 0.62 | 2.25 | 0.52 | 2.43 | 0.36 |
| Bottom | 1.21 | 0.41 | 2.23 | 0.25 | 1.98 | 0.23 |
| Top | 0.81 | 0.87 | 1.57 | 0.46 | 2.01 | 0.27 |
| Windward | 0.33 | 0.57 | 0.86 | 0.41 | 1.09 | 0.51 |
| Latent Variable | C. Alpha Coefficient | DG. Rho Coefficient | First Eigenvalue | Second Eigenvalue |
|---|---|---|---|---|
| Topography | 0.89 | 0.92 | 2.58 | 0.53 |
| Shrub | 0.61 | 0.84 | 1.43 | 0.56 |
| Herbaceous | 0.66 | 0.80 | 2.09 | 0.99 |
| Soil moisture | 0.79 | 0.88 | 2.14 | 0.70 |
| Parameter | Value |
|---|---|
| n_estimators | 49 |
| min_samples_split | 6 |
| min_samples_leaf | 1 |
| random_state | 42 |
| Parameter | Value |
|---|---|
| n_estimators | 87 |
| learning_rate | 0.1 |
| subsample | 0.05 |
| colsample_bytree | 0.8 |
| max_depth | 5 |
| reg_alpha | 0 |
| reg_lambda | 1 |
| Evaluation Metrics | Algorithm | Surface Layer | Middle Layer | Deep Layer |
|---|---|---|---|---|
| R2 | RF | 0.84 | 0.70 | 0.83 |
| XGBoost | 0.68 | 0.61 | 0.59 | |
| MSE | RF | 0.19 | 0.52 | 0.30 |
| XGBoost | 0.31 | 0.45 | 0.36 | |
| MAE | RF | 0.34 | 0.50 | 0.35 |
| XGBoost | 0.28 | 0.58 | 0.41 | |
| MAPE | RF | 5.25% | 13.5% | 5.47% |
| XGBoost | 7.11% | 13.7% | 11.59% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, H.; Zhang, D.; Zhang, Z.; Zhao, Y.; Shi, Z. Influence of Soil Moisture in Semi-Fixed Sand Dunes of the Tengger Desert, China, Based on PLS-SEM and SHAP Models. Sustainability 2024, 16, 6971. https://doi.org/10.3390/su16166971
Qi H, Zhang D, Zhang Z, Zhao Y, Shi Z. Influence of Soil Moisture in Semi-Fixed Sand Dunes of the Tengger Desert, China, Based on PLS-SEM and SHAP Models. Sustainability. 2024; 16(16):6971. https://doi.org/10.3390/su16166971
Chicago/Turabian StyleQi, Haidi, Dinghai Zhang, Zhishan Zhang, Youyi Zhao, and Zhanhong Shi. 2024. "Influence of Soil Moisture in Semi-Fixed Sand Dunes of the Tengger Desert, China, Based on PLS-SEM and SHAP Models" Sustainability 16, no. 16: 6971. https://doi.org/10.3390/su16166971
APA StyleQi, H., Zhang, D., Zhang, Z., Zhao, Y., & Shi, Z. (2024). Influence of Soil Moisture in Semi-Fixed Sand Dunes of the Tengger Desert, China, Based on PLS-SEM and SHAP Models. Sustainability, 16(16), 6971. https://doi.org/10.3390/su16166971