
Direct and Indirect Infection Effects of Four Potent Fungal Isolates on the Survival and Performance of Fall Armyworm Larval Parasitoid Cotesia icipe

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fall Armyworm Rearing
2.2. Cotesia icipe Parasitoid Rearing
2.3. Fungal Isolates Culture and Conidia Viability Assessment
2.4. Direct Infection Effects of Metarhizium anisopliae and Beauveria bassiana Isolates on Cotesia icipe
2.5. Indirect Infection Effects of Metarhizium anisopliae and Beauveria bassiana Isolates on Cotesia icipe
2.6. Effect of Entomopathogenic fungi on Cotesia icipe Emergence and Sex Ratio
2.7. Data Analysis
3. Results
3.1. Direct Effects of Entomopathogenic fungal Isolates on Cotesia icipe and FAW Larvae
3.2. Indirect Effects of Entomopathogenic fungal Isolates on Cotesia icipe Adults
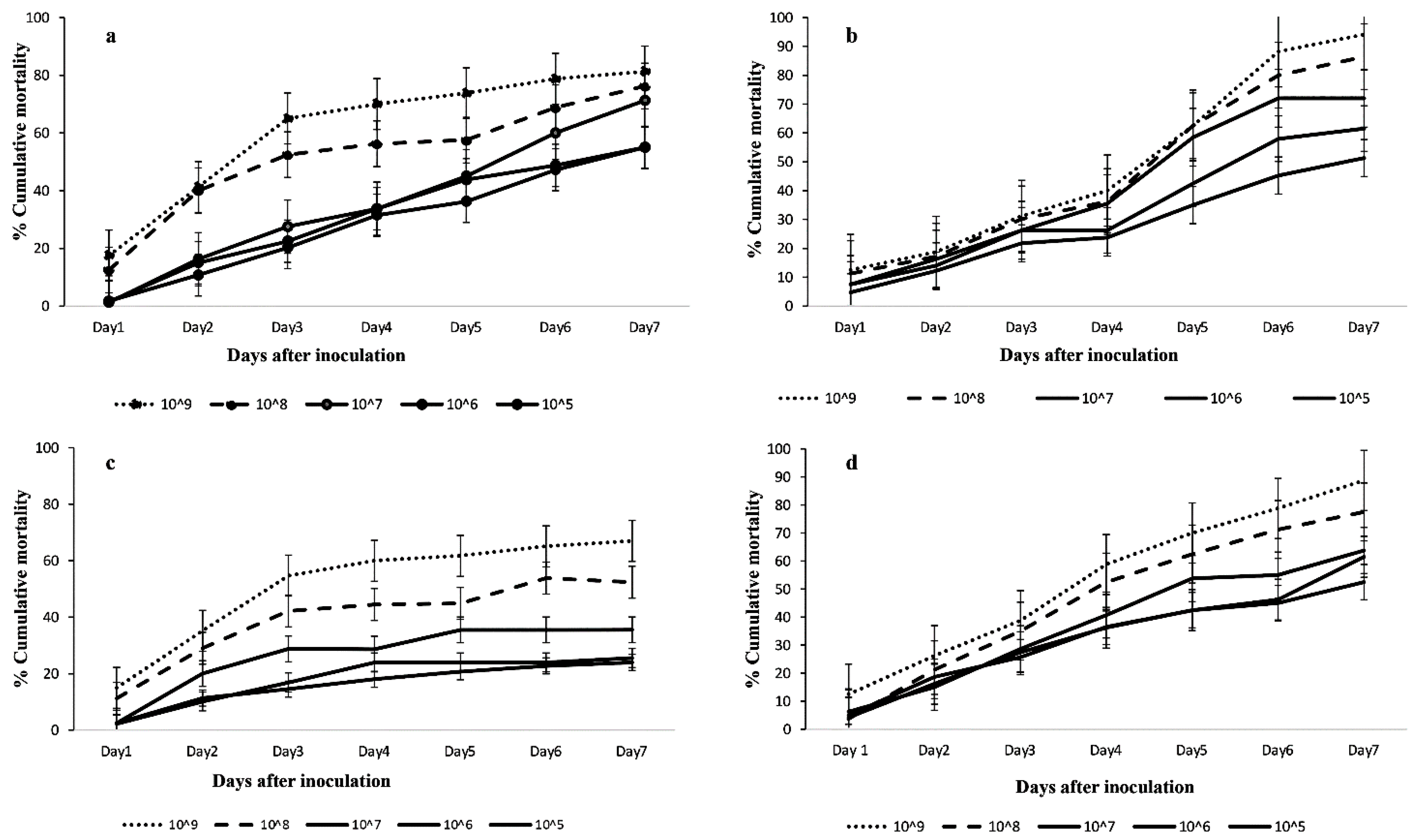
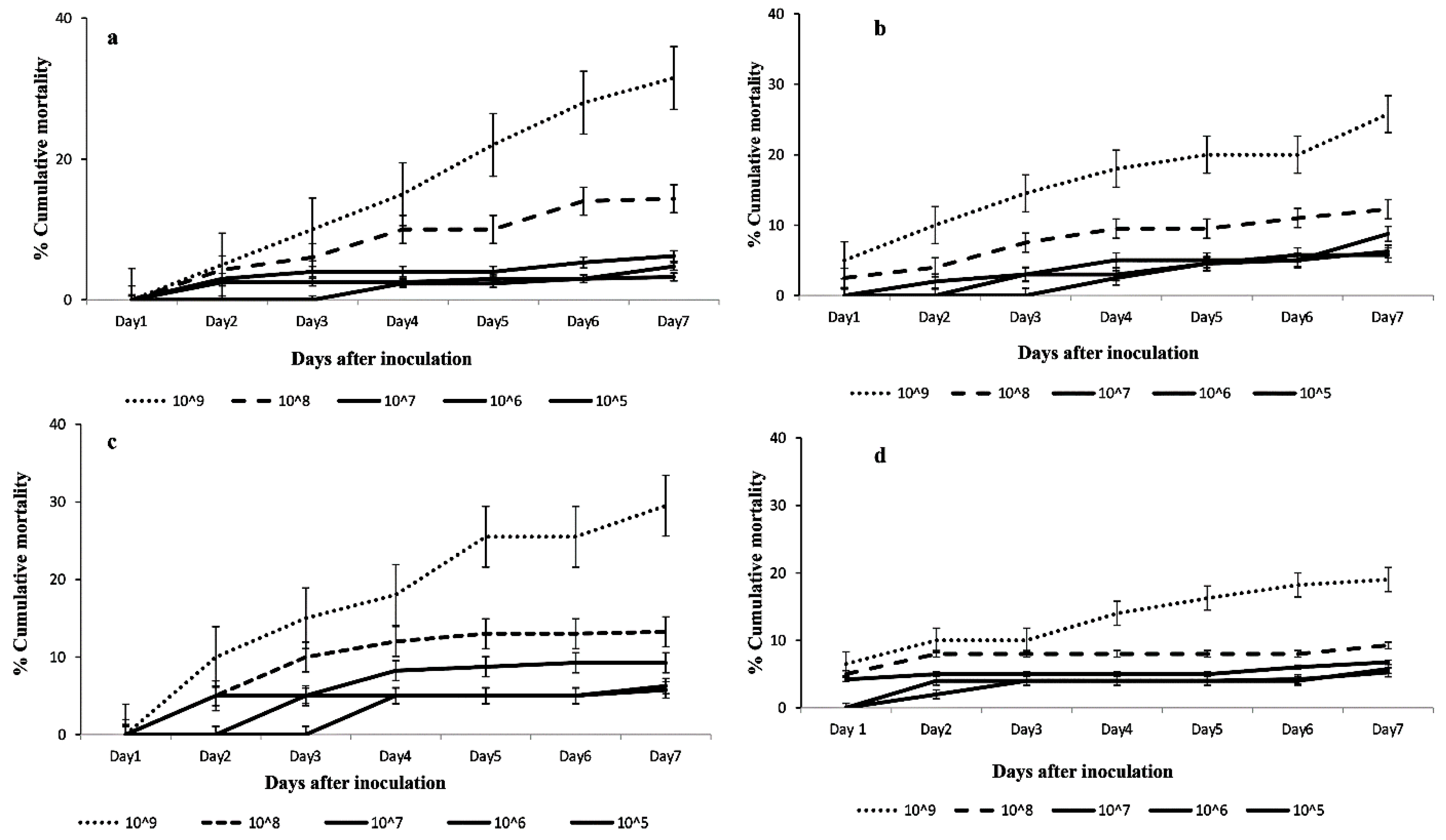
3.2.1. Mortality Rates of Cotesia icipe Adults and 2nd Instar Larvae of FAW after Indirect Exposure to FAW Larvae infected with Different Fungal Isolate Concentrations
3.2.2. Effect of Entomopathogenic fungal Isolate Concentrations on Cotesia icipe Emergence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Escamilla, R. Food security and the 2015–2030 sustainable development goals: From human to planetary health: Perspectives and opinions. Curr. Dev. Nutr. 2017, 1, e000513. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Goergen, G.; Lava, K.P.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae): A New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Tindo, M.; Tigui, A.; Kengni, F.; Atanga, J.; Bila, S.; Doumtsop, A.; Abega, R. First report of the fall army worm, Spodoptera frugiperda (Lepidoptera, Noctuidae) in Cameroon. Cameroon J. Biol. Biochem. Sci. 2017, 25, 30–32. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Sustainable Management of the Fall Armyworm in Africa; FAO Programme for Action; FAO: Rome, Italy, 2017; Available online: http://www.fao.org/3/a-bt417e.pdf (accessed on 19 July 2019).
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Akutse, K.S.; Subramanian, S.; Maniania, K.N.; Dubois, T.; Ekesi, S. Biopesticide Research and Product Development in Africa for Sustainable Agriculture and Food Security—Experiences from the International Centre of Insect Physiology and Ecology (icipe). Front. Sustain. Food Syst. 2020, 4, 563016. [Google Scholar] [CrossRef]
- Akutse, K.S.; Kimemia, J.W.; Ekesi, S.; Khamis, F.M.; Ombura, O.L.; Subramanian, S. Ovicidal effects of entomopathogenic fungal isolates on the invasive Fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2019, 143, 626–634. [Google Scholar] [CrossRef]
- Akutse, K.S.; Khamis, F.M.; Ambele, F.C.; Kimemia, J.W.; Ekesi, S.; Subramanian, S. Combining insect pathogenic fungi and a pheromone trap for sustainable management of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2020, 177, 107477. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Wamalwa, M.; Obala, F.; Tonnang, H.E.Z.; Tefera, T.; Calatayud, P.A.; Subramanian, S.; Ekesi, S. A deadly encounter: Alien invasive Spodoptera frugiperda in Africa and indigenous natural enemy, Cotesia icipe (Hymenoptera, Braconidae). PLoS ONE 2021, 16, e0253122. [Google Scholar] [CrossRef]
- Fiaboe, K.R.; Agboka, K.; Agboyi, L.K.; Koffi, D.; Ofoe, R.; Kpadonou, G.E.; Agnamba, A.O.; Assogba, K.; Adjevi, M.K.A.; Zanou, K.T.; et al. First report and distribution of the South American tomato pinworm, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in Togo. Phytoparasitica 2021, 49, 167–177. [Google Scholar] [CrossRef]
- Akutse, K.S.; Maniania, N.K.; Fiaboe, K.K.M.; Van den Berg, J.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Goettel, M.S.; Inglis, G.D. Fungi: Hyphomycetes. In Manual of Techniques in Insect Pathology; Academic Press: Cambridge, MA, USA, 1997; pp. 213–249. [Google Scholar]
- Migiro, L.N.; Maniania, N.K.; Chabi-Olaye, A.; Vandenberg, J. Pathogenicity of entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana (Hypocreales: Clavicipitaceae) isolates to the adult pea leafminer (Diptera: Agromyzidae) and prospects of an autoinoculation device for infection in the field. Environ. Entomol. 2010, 39, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Opisa, S.; du Plessis, H.; Akutse, K.S.; Fiaboe, K.K.M.; Ekesi, S. Horizontal transmission of Metarhizium anisopliae between Spoladea recurvalis (Lepidoptera: Crambidae) adults and compatibility of the fungus with the attractant phenylacetaldehyde. Microb. Pathog. 2019, 131, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Burgerjon, A. An improved laboratory apparatus for applying direct sprays and surface films, with data on the electrostatic charge on atomized fluids. Ann. Appl. Biol. 1956, 39, 1–28. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 23 June 2021).
- Maina, U.M.; Galadima, I.B.; Gambo, F.M.; Zakaria, D. A review on the use of entomopathogenic fungi in the management of insect pests of field crops. J. Entomol. Zool. Stud. 2018, 6, 27–32. Available online: https://www.entomoljournal.com/archives/?year=2018&vol=6&issue=1&ArticleId=2930 (accessed on 18 November 2021).
- Opisa, S.; du Plessis, H.; Akutse, K.S.; Fiaboe, K.K.M.; Ekesi, S. Effects of Entomopathogenic fungi and Bacillus thuringiensis -based biopesticides on Spoladea recurvalis (Lepidoptera: Crambidae). J. Appl. Entomol. 2018, 142, 617–626. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Ansari, M.A.; Pope, E.C.; Carpenter, S.; Scholte, E.J.; Butt, T.M. Entomopathogenic fungus as a biological control for an important vector of livestock disease: The culicoides biting midge. PLoS ONE 2011, 6, e16108. [Google Scholar] [CrossRef]
- Alves, R.T.; Bateman, R.P.; Gunn, J.; Prior, C.; Leather, S.R. Effects of different formulations on viability and medium-term storage of Metarhizium anisopliae conidia. Neotrop. Entomol. 2002, 31, 91–99. [Google Scholar] [CrossRef]
- Abbas, M.S.T. Interactions between Entomopathogenic Fungi and Entomophagous Insects. Adv. Entomol. 2020, 8, 130–146. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall armyworm, Spodoptera frugiperda infestations in East Africa: Assessment of damage and parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Uma Devi, K.; Padmavathi, J.; Uma Maheswara Rao, C.; Khan, A.A.P.; Mohan, M.C. A study of host specificity in the entomopathogenic fungus Beauveria bassiana (Hypocreales, Clavicipitaceae). Biocontrol Sci. Technol. 2008, 18, 975–989. [Google Scholar] [CrossRef]
- Montesinos, E. Development, registration and commercialization of microbial pesticides for plant protection. Int. Microbiol. 2003, 6, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Caballero, R.; Cyman, S.; Schuster, D.J.; Portillo, H.E.; Slater, R. Baseline susceptibility of Bemisia tabaci (Genn.) biotype B in southern Florida to cyantraniliprole. Crop. Prot. 2013, 44, 104–108. [Google Scholar] [CrossRef]
- Dimbi, S.; Maniania, N.K.; Lux, S.A.; Ekesi, S.; Mueke, J.K. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathologia 2003, 156, 375–382. [Google Scholar] [CrossRef]
- Förster, H.; Kanetis, L.; Adaskaveg, J.E. Spiral gradient dilution, a rapid method for determining growth responses and 50% effective concentration values in fungus-fungicide interactions. Phytopathology 2004, 94, 163–170. [Google Scholar] [CrossRef]
- Ramanujam, B.; Poornesha, B.; Shylesha, A.N. Effect of entomopathogenic fungi against invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in maize. Egypt J. Biol. Pest Control 2020, 30, 1–5. [Google Scholar] [CrossRef]
- Morales-Reyes, C.; Mascarin, G.M.; Jackson, M.A.; Hall, D.; Sánchez-Peña, S.R.; Arthurs, S.P. Comparison of aerial conidia and blastospores from two entomopathogenic fungi against Diaphorina citri (Hemiptera: Liviidae) under laboratory and greenhouse conditions. Biocontrol. Sci. Technol. 2018, 28, 737–749. [Google Scholar] [CrossRef]
- García-Munguía, A.M.; Garza-Hernndez, J.A.; Rebollar-Tellez, E.A.; Rodríguez-Pérez, M.A.; Reyes-Villanueva, F. Transmission of Beauveria bassiana from male to female Aedes aegypti mosquitoes. Parasites Vectors 2011, 4, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Mumo, L.; Yu, J.; Fang, K. Assessing impacts of seasonal climate variability on maize yield in Kenya. Int. J. Plant Prod. 2018, 12, 297–307. [Google Scholar] [CrossRef]
- Polanczyk, R.A.; Pratissoli, D.; Dalvi, L.P.; Grecco, E.D.; Franco, C.R. Efeito de Beauveria bassiana (Bals.) Vuillemin e Metarhizium anisopliae (Metsch.) Sorokin nos parâmetros biológicos de Trichogramma atopovirilia Oatman & Platner, 1983 (Hymenoptera: Trichogrammatidade). Cienc. Agrotec. 2010, 34, 1412–1416. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fungal Species | Isolates | Source | Location/Country | Year of Isolation | % Germination ± SE |
|---|---|---|---|---|---|
| Metarhizium anisopliae | ICIPE 7 | Rhipicephalus appendiculatus | Rusinga island (Kenya) | 1996 | 93.0 ± 1.3 a |
| ICIPE 78 | Temnoschoita nigroplagiata | Ungoe (Kenya) | 1990 | 94.2 ± 1.7 a | |
| ICIPE 41 | Soil | Kenya | 1990 | 87.3 ± 2.3 b | |
| Beauveria bassiana | ICIPE 621 | Soil | Kericho (Kenya) | 2008 | 91.2 ± 1.04 ab |
| Fungal Isolates | Mean Adult Mortality (%) | FAW Larval Mortality (%) | Parasitism Rates (%) | Sex Ratios (F:M) | Cotesia icipe LT50 (days) | % Mycosis of C. icipe Cadavers |
|---|---|---|---|---|---|---|
| ICIPE 7 | 73.95 ± 7.49 a | 55.25 ± 6.74 a | 35.75 ± 4.80 b | 2:1 | 2.3 ± 0.0 a | 65.75 ± 6.69 a |
| ICIPE 78 | 33.63 ± 6.63 b | 28.25 ± 4.41 b | 62.00 ± 5.02 a | 2:1 | 5.2 ± 0.0 c | 43.75 ± 5.91 ab |
| ICIPE 41 | 66.33 ± 7.29 a | 53.75 ± 3.91 a | 36.75 ± 3.82 b | 2:1 | 2.8 ± 0.0 b | 57.25 ± 6.84 ab |
| ICIPE 621 | 36.59 ± 6.65 b | 31.00 ± 5.29 b | 58.00 ± 5.24 a | 2:1 | 5.0 ± 0.0 d | 35.00 ± 4.56 b |
| Concentration | M. anisopliae ICIPE 7 | M. anisopliae ICIPE 41 | M. anisopliae ICIPE 78 | B. bassiana ICIPE 621 |
|---|---|---|---|---|
| 1 × 109 | 54.75 ± 3.33 bA (2:1) | 46.50 ± 7.27 cA (1:1) | 44.75 ± 2.90 dA (1:1) | 56.25 ± 3.40 aA (2:1) |
| 1 × 108 | 60.25 ± 5.14 bA (2:1) | 51.25 ± 5.84 bcA (2:1) | 56.00 ± 6.56 cdA (1:1) | 59.25 ± 3.74 aA (2:1) |
| 1 × 107 | 64.75 ± 3.84 abA (2:1) | 61.50±5.58 abcA (2:1) | 65.75 ± 4.53 bcA (2:1) | 61.25 ± 4.60 aA (2:1) |
| 1 × 106 | 66.50 ± 8.91 abA (2:1) | 67.25±4.73 abcA (2:1) | 71.50 ± 4.33 abA (2:1) | 66.75 ± 6.14 aA (2:1) |
| 1 × 105 | 70.50 ± 7.37 abA (2:1) | 74.25±10.05 abA (2:1) | 76.75 ± 3.66 abA (2:1) | 69.50 ± 8.68 aA (2:1) |
| Control | 85.75 ± 4.46 aA (2:1) | 83.50 ± 2.47 aA (2:1) | 82.75 ± 3.47 aA (2:1) | 77.25 ± 3.33 aA (2:1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chepkemoi, J.; Fening, K.O.; Ambele, F.C.; Munywoki, J.; Akutse, K.S. Direct and Indirect Infection Effects of Four Potent Fungal Isolates on the Survival and Performance of Fall Armyworm Larval Parasitoid Cotesia icipe. Sustainability 2023, 15, 3250. https://doi.org/10.3390/su15043250
Chepkemoi J, Fening KO, Ambele FC, Munywoki J, Akutse KS. Direct and Indirect Infection Effects of Four Potent Fungal Isolates on the Survival and Performance of Fall Armyworm Larval Parasitoid Cotesia icipe. Sustainability. 2023; 15(4):3250. https://doi.org/10.3390/su15043250
Chicago/Turabian StyleChepkemoi, Junitor, Ken Okwae Fening, Felicitas Chaba Ambele, Joseph Munywoki, and Komivi Senyo Akutse. 2023. "Direct and Indirect Infection Effects of Four Potent Fungal Isolates on the Survival and Performance of Fall Armyworm Larval Parasitoid Cotesia icipe" Sustainability 15, no. 4: 3250. https://doi.org/10.3390/su15043250
APA StyleChepkemoi, J., Fening, K. O., Ambele, F. C., Munywoki, J., & Akutse, K. S. (2023). Direct and Indirect Infection Effects of Four Potent Fungal Isolates on the Survival and Performance of Fall Armyworm Larval Parasitoid Cotesia icipe. Sustainability, 15(4), 3250. https://doi.org/10.3390/su15043250