
The Climate Change Influence on Cedrela odorata L. Radial Growth in the Amazon

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
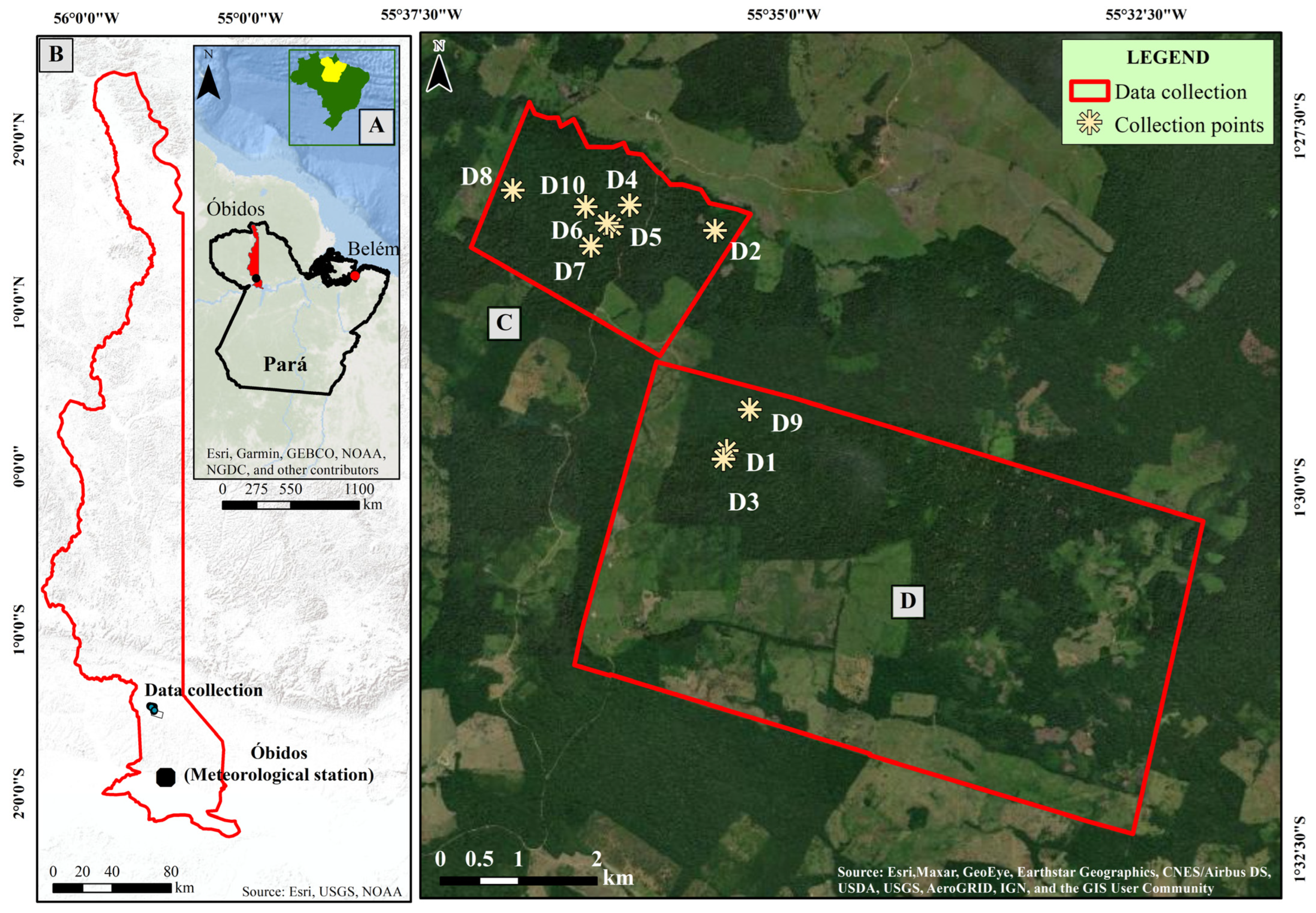
2.1. Study Area, Soil and Climate
2.2. Tree Ring Sample Collection and Chronology Establishment
2.3. Climate Data
2.4. Data Analysis
3. Results
3.1. Characterization and Measurement of C. odorata Growth Rings
3.2. Meteorological Data Analysis
3.3. Correlation between Growth and Local Climate
3.4. ENSO and Growth Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haghtalab, N.; Moore, N.; Heerspink, B.P.; Hyndman, D.W. Evaluating spatial patterns in precipitation trends across the Amazon basin driven by land cover and global scale forcings. Theor. Appl. Climatol. 2020, 140, 411–427. [Google Scholar] [CrossRef]
- Sorí, R.; Marengo, J.; Nieto, R.; Drumond, A.; Gimeno, L. Drought and wet episodes in Amazonia: The role of atmospheric moisture transport. In Proceedings of the 1st International Electronic Conference on Hydrological Cycle, Sciforum Electronic Conference Series, Basel, Switzerland, 12–16 November 2017; pp. 12–16. [Google Scholar] [CrossRef]
- Satyamurty, P.; Da Costa, C.P.W.; Manzi, A.O. Moisture source for the Amazon basin: A study of contrasting years. Theor. Appl. Climatol. 2013, 111, 195–209. [Google Scholar] [CrossRef]
- Jiménez-Muñoz, J.C.; Mattar, C.; Barichivich, J.; Santamaría-Artigas, A.; Takahashi, K.; Malhi, Y.; Van Der Schrier, G. Record-breaking warming and extreme drought in the Amazon rainforest during the course of El Niño 2015–2016. Sci. Rep. 2016, 6, 33130. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Trejo, F.; Barbosa, H.; Giovannettone, J.; Kumar, T.V.L.; Kumar, T.M.; De Oliveira, B.C. Drought variability and land degradation in the Amazon River basin. Front. Earth Sci. 2022, 10, 939908. [Google Scholar] [CrossRef]
- Douville, H.; Raghavan, K.; Renwick, J.; Allan, R.P.; Arias, P.A.; Barlow, M.; Cerezo-Mota, R.; Cherchi, A.; Gan, T.Y.; Gergis, J.; et al. Water Cycle Changes [M/OL]//IPCC. In Climate Change 2021: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Moura, R.G.; Correia, F.W.S.; Veiga, J.A.P.; Capistrano, V.B.; Kobuta, P.Y. Avaliação do Brazilian Global Atmospheric Model na Simulação dos Componentes do Balanço de água na Bacia Amazônica. Rev. Bras. Meteorol. 2021, 36, 23–37. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef]
- Quintilhan, M.T.; Santini, L., Jr.; Rodriguez, D.R.O.; Guillemot, J.; Cesilio, G.H.M.; Chambi-Legoas, R.; Nouvellon, Y.; Tomazello-Filho, M. Growth-ring boundaries of tropical tree species: Aiding delimitation by long histological sections and wood density profiles. Dendrochronologia 2021, 69, 125878. [Google Scholar] [CrossRef]
- Silva, M.S.; Funch, L.S.; Silva, L.B. The growth ring concept: Seeking a broader and unambiguous approach covering tropical species. Biol. Rev. 2019, 94, 1161–1178. [Google Scholar] [CrossRef]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010; pp. 1–73. [Google Scholar]
- Brienem, R.J.W.; Zuidema, P.A.; Schöngart, J. Tree rings in the tropics: Insights into the ecology and climate sensitivity of tropical species. In Tropical Tree Physiology; Goldstein, G., Santiago, L.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 439–461. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program cofecha. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Albernaz, A.L.K.M. Estudo Preliminar da Anuidade dos Anéis de Crescimento em Árvores de Espécies Ocorrentes na Região do TAPAJÓS (PA). In Distrito Florestal Sustentável da BR 163: Dinâmicas Sociais e Mudanças Ambientais e Produção; Moutinho, V.H.P., Chagas, M.P., Tomazello Filho, M., Silva, R., Eds.; MPEG: Belém, Brazil, 2015; p. 464. [Google Scholar]
- FAO. Informe de la Quinta Reunión del Cuadro del Expertos de la FAO en Recursos Genéticos Forestales; FO:FGR/5/Rep; FAO: Roma, Italy, 1984. [Google Scholar]
- CITES 2015—Convention on international Trade in Endangered Species of Wild Fauna and Flora. Available online: https://cites.org/sites/default/files/eng/app/2015/E-Appendices-2015-02-05.pdf (accessed on 11 November 2023).
- Paredes-Villanueva, K.; López, L.; Cerrillo, R.M.N. Regional chronologies of Cedrela fissilis and Cedrela angustifolia in three forest types and their relation to climate. Trees 2016, 30, 1581–1593. [Google Scholar] [CrossRef]
- Santos, G.M.; Granato-Souza, D.; Barbosa, A.C.; Oelkers, R.; Andreu-Hayles, L. Radiocarbon analysis confirms annual periodicity in Cedrela odorata tree rings from the equatorial Amazon. Quat. Geochronol. 2020, 58, 101079. [Google Scholar] [CrossRef]
- Pagotto, M.A.; Menezes, I.R.N.; Costa, C.M.; Lisi, C.S.; Bräuning, A. Oxygen isotopes in tree rings of Cedrela odorata L. as an indicator of hydroclimate variations in a seasonally dry tropical forest in northeastern Brazil. Trees 2021, 35, 1889–1903. [Google Scholar] [CrossRef]
- Menezes, I.R.N.; Aragão, J.R.V.; Pagotto, M.A.; Lisi, C.S. Teleconnections and edaphoclimatic effects on tree growth of Cedrela odorata L. in a seasonally dry tropical forest in Brazil. Dendrochronologia 2022, 72, 125932. [Google Scholar] [CrossRef]
- Callado, C.H.; Roig, F.A.; Tomazello-Filho, M.; Barros, C.F. Cambial growth periodicity studies of South American woody species-a review. IAWA J. 2013, 34, 213–230. [Google Scholar] [CrossRef]
- Santos, A.D.; Bechis, I.O.; Miranda, G.F.; Souto, J.I.O.; Beltrão, N.E.S. Variabilidade espaço-temporal da precipitação na microrregião de Óbidos, Pará: Um estudo utilizando a CPC Morphing Tecnique (2005–2017). Rev. De Geogr. 2019, 362, 152–167. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Arstan: Guide for Computer Program Arstan; Adapted from User’s Manual for Program Arstan, in Tree-Ring Chronologies of Western North America: California, Eastern Oregon and Northern Great Basin; 1996; pp. 50–65. Available online: https://www.ltrr.arizona.edu/~sheppard/DISC2019/arstan.txt (accessed on 8 August 2021).
- Trenberth, K.E. The definition of El Niño. Bull. Am. Meteorol. Soc. 1997, 78, 2771–2778. [Google Scholar] [CrossRef]
- Mondal, A.; Kundu, S.; Mukhopadhyay, A. Rainfall trend analysis by Mann-Kendall test: A case study of north-eastern part of Cuttack district, Orissa. Int. J. Geol. Earth Environ. Sci. 2012, 2, 70–78. [Google Scholar]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program Library in R. 2021. Available online: https://cran.r-project.org/web/packages/dplR/index.html/ (accessed on 8 August 2021).
- Coutinho, E.C.; Rocha, E.J.P.; Lima, A.M.M.; Ribeiro, H.M.C.; Gutierrez, L.A.C.L.; Barbosa, A.J.S.; Paes, G.K.A.A.; Bispo, C.J.C.; Tavares, P.A. Water Balance in the Brazilian Amazon Basin. Rev. Bras. Geogr. Física 2018, 11, 1926–1940. [Google Scholar] [CrossRef]
- Oliveira, V.G.; Santos, C.A.C. Trends in extreme climate índices for Pará State, Brazil. Rev. Bras. Meteorol. 2017, 32, 13–24. [Google Scholar] [CrossRef]
- Lisi, C.S.; Pagotto, M.A.; Anholeto, C.R., Jr.; Nogueira, F.C., Jr.; Santos, H.L.; Costa, C.M.; Menezes, I.R.N.; Juñet, F.A.R.; Tomazello-Filho, M. Dendroecological Studies with Cedrela odorata L. Northeastern Brazil. In Latin American Dendroecology; Pompa-García, M., Camarero, J.J., Eds.; Springer: Cham, Switerland, 2020; pp. 37–59. [Google Scholar]
- Granato-Souza, D.; Stahle, D.; Barbosa, M.C. Tree rings and rainfall in the equatorial Amazon. Clim. Dyn. 2019, 52, 1857–1869. [Google Scholar] [CrossRef]
- Dünisch, O.; Montoia, V.R.; Bauch, J. Dendroecological investigations on Swietenia macrophylla King and Cedrela odorata L. (Meliaceae) in the central Amazon. Trees 2003, 17, 244–250. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal; Artmed: Porto Alegre, Brazil, 2017; Volume 6, p. 888. [Google Scholar]
- Marengo, J.A.; Alves, L.M.; Beserra, E.A.; Lacerda, F.F. Variabilidade e mudanças climáticas no semiárido brasileiro. In Recursos Hídricos em Regiões Áridas e Semiáridas; Instituto Nacional do Semiárido-INSA: Campina Grande, Brazil, 2011; pp. 384–422. [Google Scholar]
- Antezana-Vera, S.; Marenco, R.A. Intra-annual tree growth responds to micrometeorological variability in the central Amazon. iforest—Biogeosci. For. 2021, 14, 242–249. [Google Scholar] [CrossRef]
- Venegas-Gonzáles, A.; Roig, F.A.; Lisi, C.S.; Albiero-Junior, A.; Alvares, C.A.; Tomazello-Filho, M. Drought and climate change incidence on hotspot Cedrela forests from the Mata Atlântica biome in southeastern Brazil. Glob. Ecol. Conserv. 2018, 15, e00408. [Google Scholar] [CrossRef]
- Bräuning, A.; Arsalani, M.; Grießinger, J. Tree-ring-based seasonal temperature reconstructions and ecological implications of recent warming on oak forest health in the Zagros Mountains, Iran. Int J Biometeorol. 2022, 66, 2553–2565. [Google Scholar] [CrossRef]
- Aragão, J.R.V.; Zuidema, P.A.; Groenendijk, P. Climate-growth relations of congeneric tree species vary across a tropical vegetation gradient in Brazil. Dendrochronologia 2022, 71, 125913. [Google Scholar] [CrossRef]
- Granato-Souza, D.; Stahle, D.W.; Torbenson, M.C.A.; Howard, I.M.; Barbosa, A.C.; Feng, S.; Fernandes, K.; Schöngart, J. Multidecadal changes in wet season precipitation totalsover the Eastern Amazon. Geophys. Res. Lett. 2020, 47, e2020GL087478. [Google Scholar] [CrossRef]
- Do Carmo, F.H.D.; Latorraca, J.V.F.; Volpato, M.; Santos, G.C.V.; Souza, P.P.; Takizawa, F.H. Climate variations affect the growth period of young Tectona grandis Linn F. in the Amazon. Acta Bot. Bras. 2022, 36, e2020abb0525. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Series | Interval | Year Average | Max. Segments | Flags | Master Correlation | Sensibility |
|---|---|---|---|---|---|---|
| D1 | 1844–2017 | 174 | 7 | 3 | 0.227 | 0.386 |
| D2 | 1908–2017 | 110 | 4 | 1 | 0.433 | 0.326 |
| D3 | 1914–2017 | 104 | 4 | 1 | 0.318 | 0.439 |
| D4 | 1904–2017 | 112 | 4 | 0 | 0.521 | 0.286 |
| D5 | 1941–2017 | 77 | 3 | 0 | 0.505 | 0.254 |
| D6 | 1904–2017 | 114 | 4 | 0 | 0.575 | 0.426 |
| D7 | 1812–2017 | 206 | 7 | 4 | 0.210 | 0.371 |
| D8 | 1956–2017 | 62 | 2 | 2 | 0.235 | 0.283 |
| D9 | 1911–2017 | 107 | 4 | 1 | 0.396 | 0.406 |
| D10 | 1853–2017 | 165 | 6 | 2 | 0.201 | 0.237 |
| Total | - | 1.231 | 45 | 14 | - | - |
| Critical correlation | 0.342 | 0.346 | ||||
| Meteorological Variable | Correlation Coefficient |
|---|---|
| Relative humidity (%) | −0.100 |
| Insolation | 0.048 ns |
| Precipitation | 0.003 ns |
| Mean temperature (°C) | 0.236 |
| Max. temperature (°C) | 0.062 ns |
| Min. temperature (°C) | 0.467 |
| Cloudiness | 0.353 |
| Evapotranspiration | −0.259 |
| Thermic Amplitude (°C) | −0.028 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Costa, W.P.L.B.; Pinheiro, É.F.M.; Latorraca, J.V.F.; Moutinho, V.H.P.; Carmo, F.H.D.J.; Ataíde, G.C.V.S.; Volpato, M.; Aguiar, D.L.; Andrade, F.W.C. The Climate Change Influence on Cedrela odorata L. Radial Growth in the Amazon. Sustainability 2023, 15, 16755. https://doi.org/10.3390/su152416755
Da Costa WPLB, Pinheiro ÉFM, Latorraca JVF, Moutinho VHP, Carmo FHDJ, Ataíde GCVS, Volpato M, Aguiar DL, Andrade FWC. The Climate Change Influence on Cedrela odorata L. Radial Growth in the Amazon. Sustainability. 2023; 15(24):16755. https://doi.org/10.3390/su152416755
Chicago/Turabian StyleDa Costa, Wayka P. L. B., Érika F. M. Pinheiro, João V. F. Latorraca, Victor H. P. Moutinho, Fábio H. D. J. Carmo, Glaycianne C. V. S. Ataíde, Maristela Volpato, Diego L. Aguiar, and Fernando W. C. Andrade. 2023. "The Climate Change Influence on Cedrela odorata L. Radial Growth in the Amazon" Sustainability 15, no. 24: 16755. https://doi.org/10.3390/su152416755
APA StyleDa Costa, W. P. L. B., Pinheiro, É. F. M., Latorraca, J. V. F., Moutinho, V. H. P., Carmo, F. H. D. J., Ataíde, G. C. V. S., Volpato, M., Aguiar, D. L., & Andrade, F. W. C. (2023). The Climate Change Influence on Cedrela odorata L. Radial Growth in the Amazon. Sustainability, 15(24), 16755. https://doi.org/10.3390/su152416755