
Exploring the Structure of Static Net Fisheries in a Highly Invaded Region: The Case of Rhodes Island (Eastern Mediterranean)

 ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
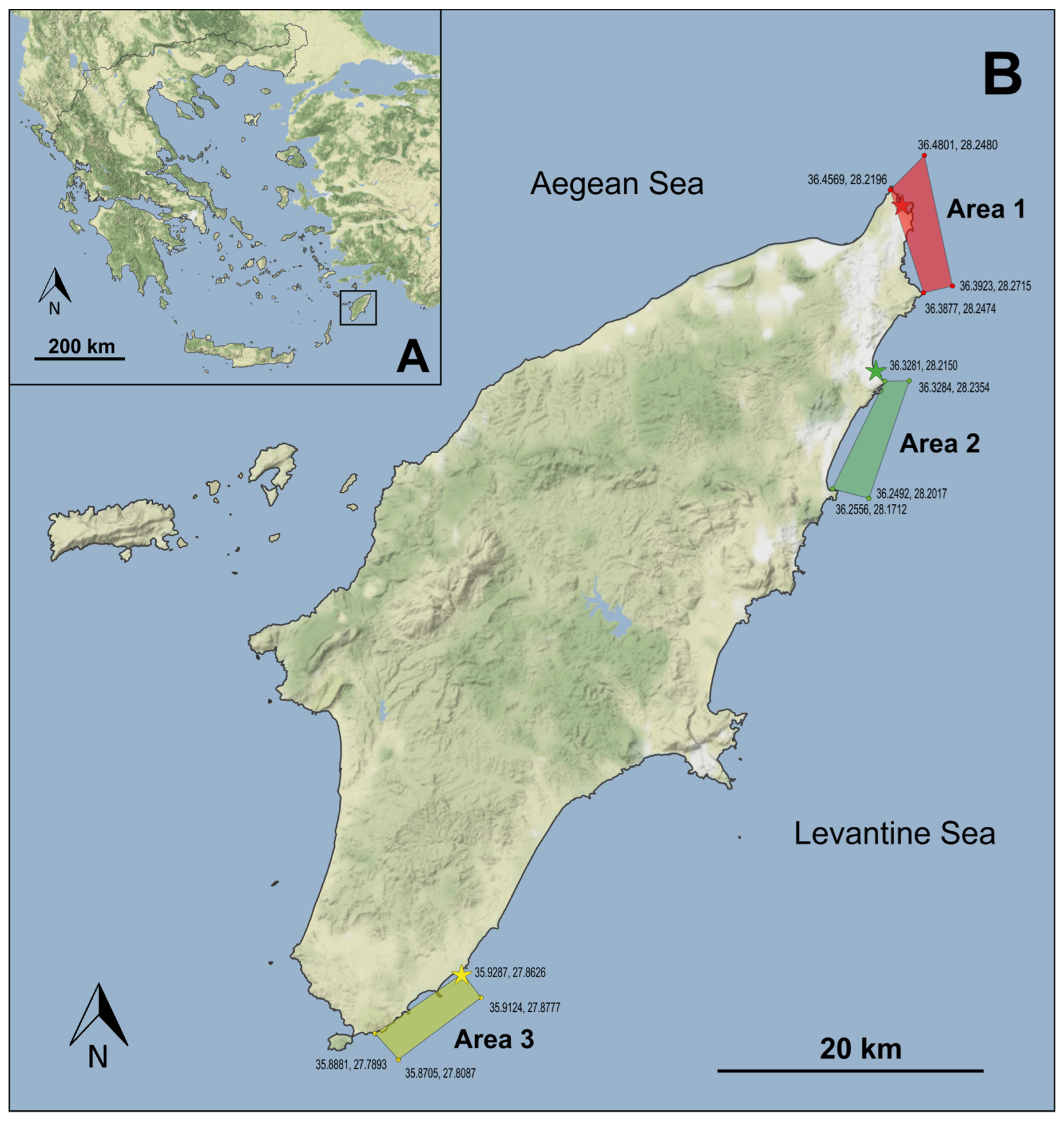
2.1. Study Area
2.2. Fishing Gear and Species Identification
2.3. Statistical Analyses
3. Results
3.1. Study Area
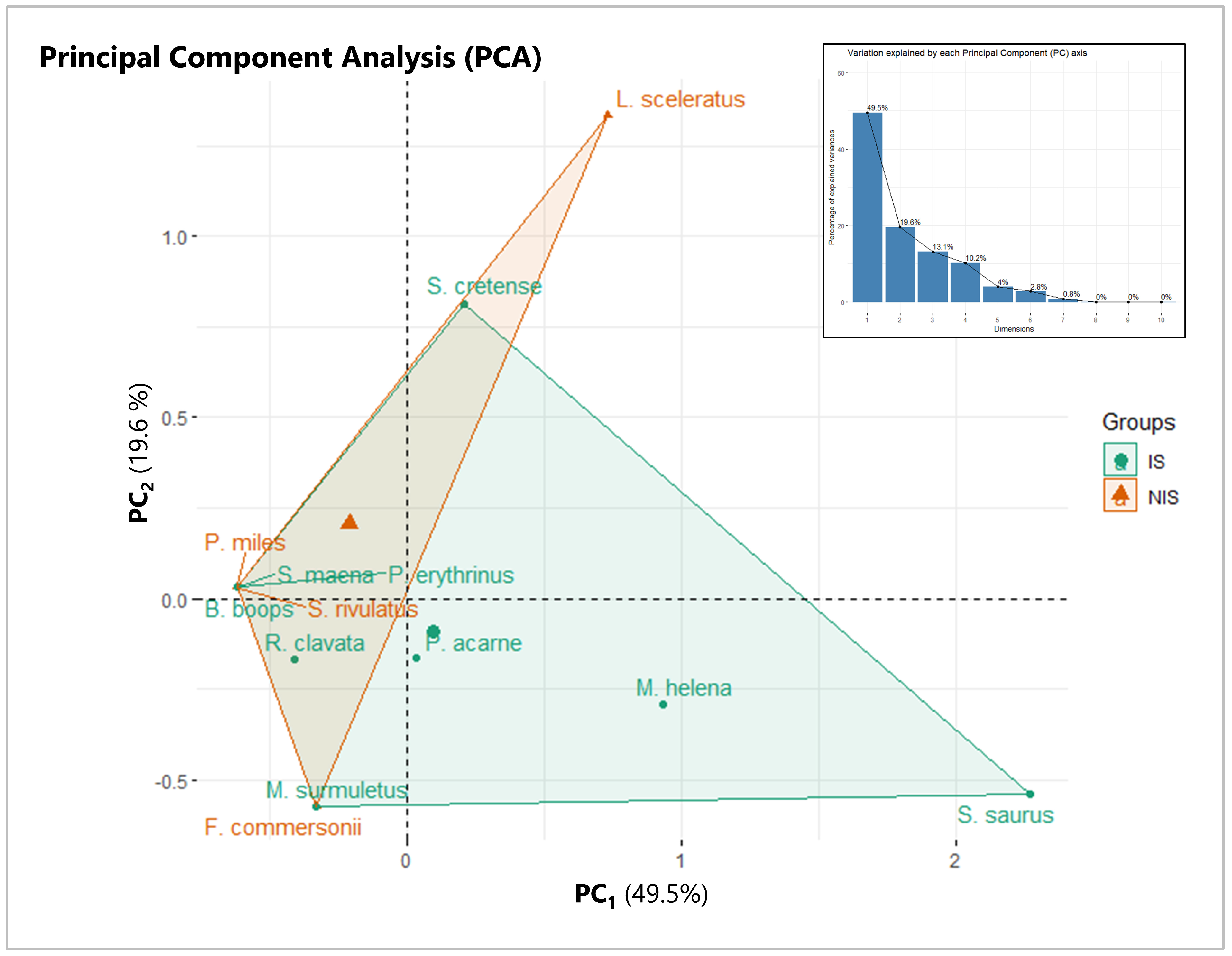
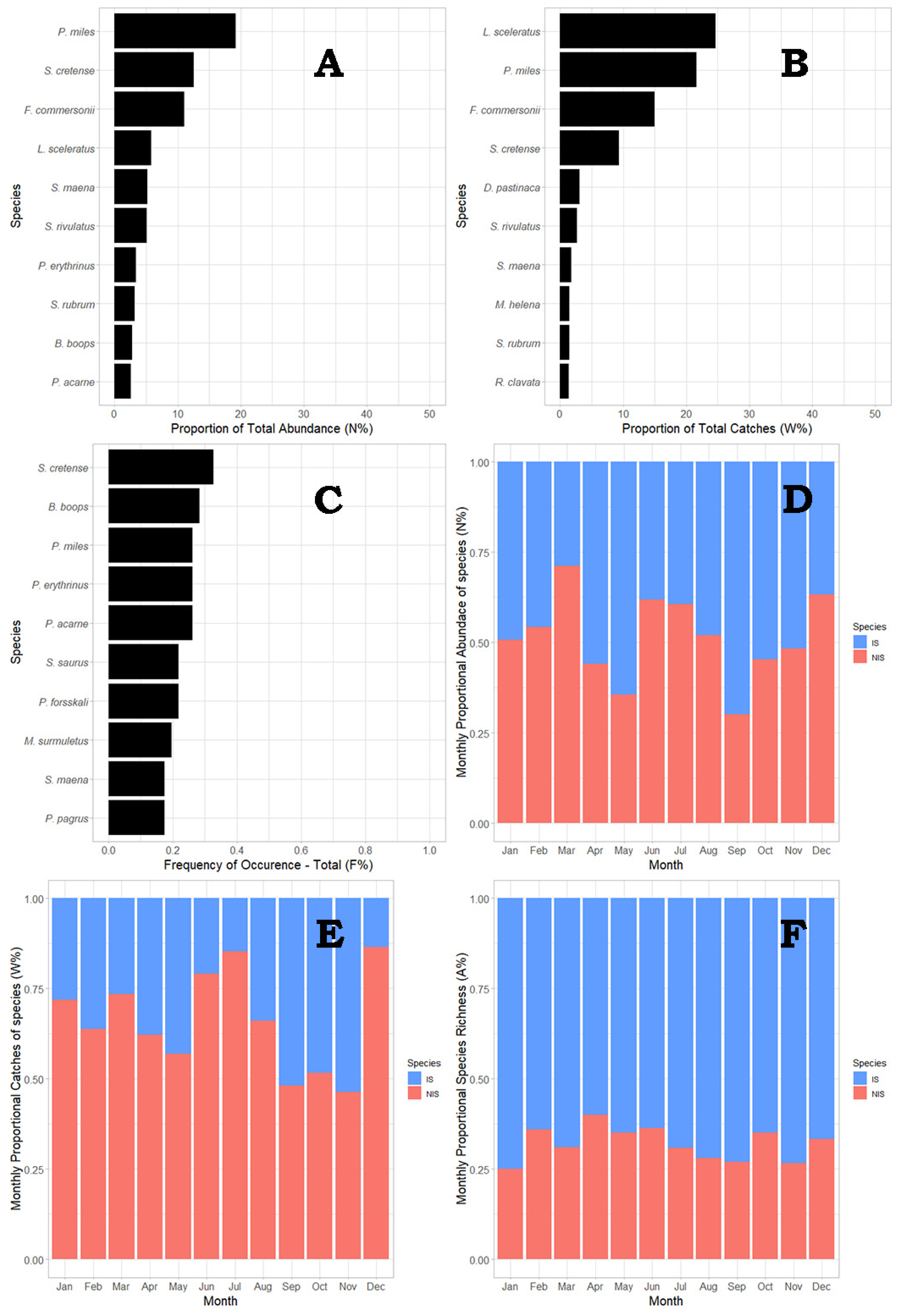
3.2. Population Structure
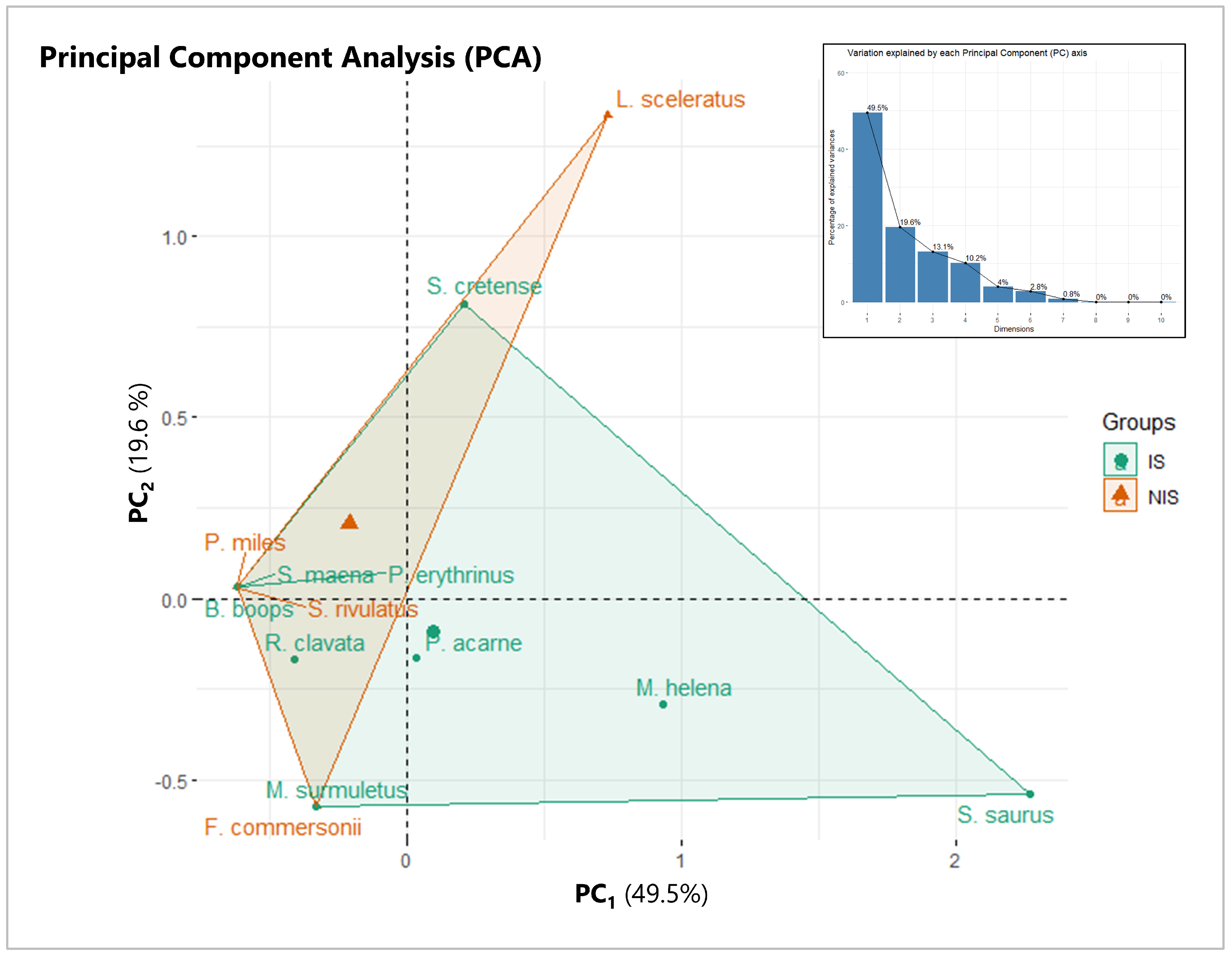
3.3. Species Prevalence Structure
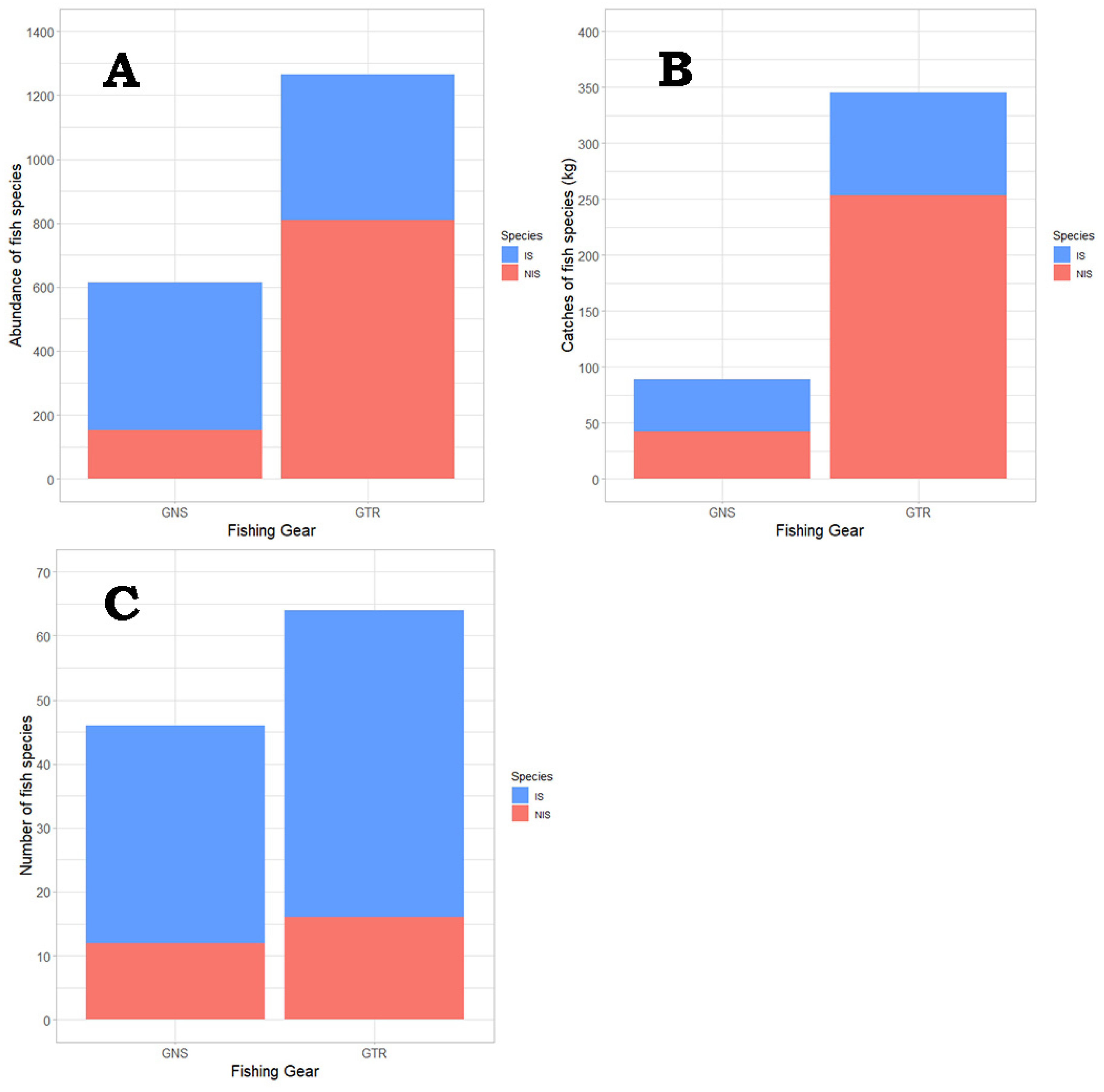
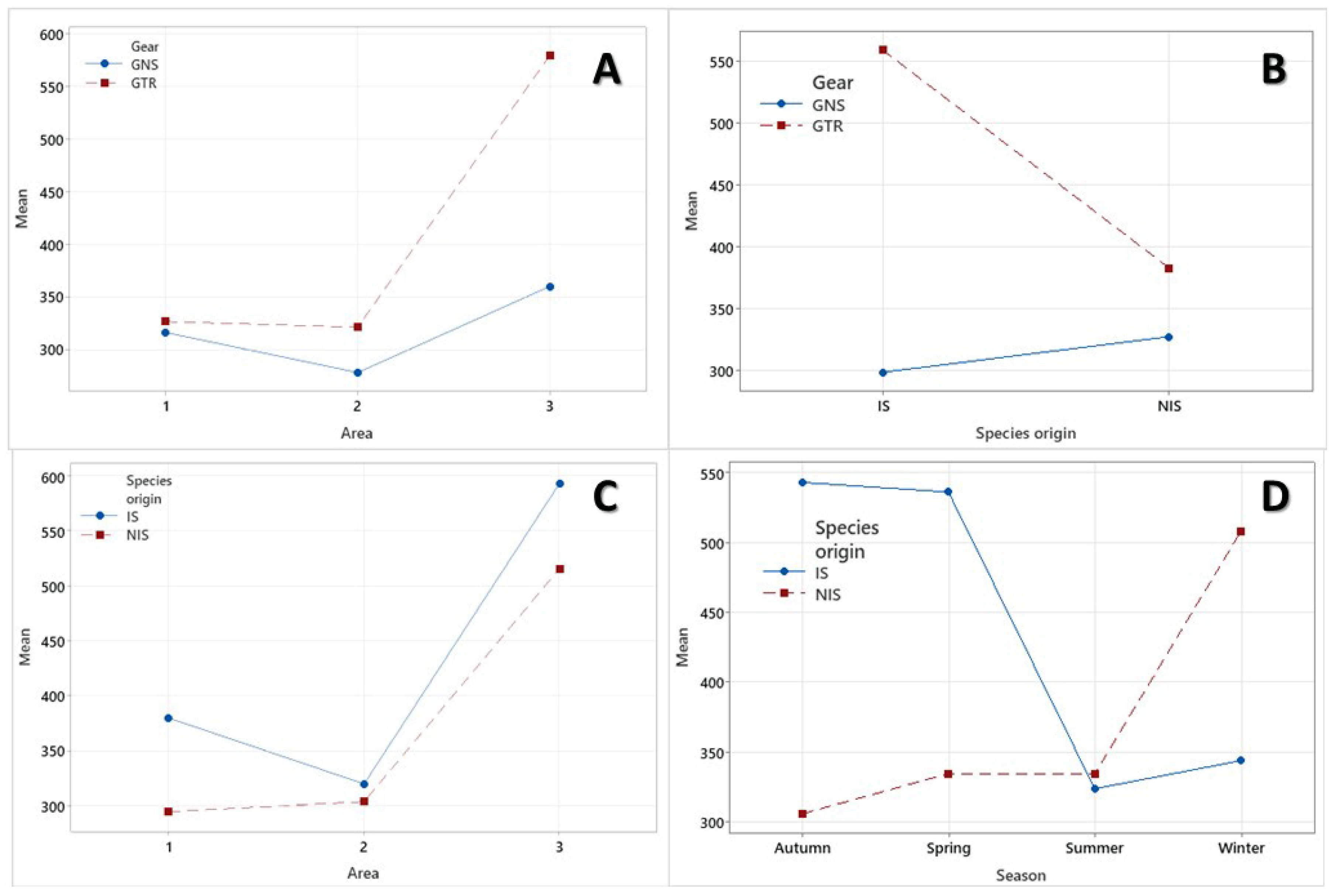
3.4. Static Gear Type Structure
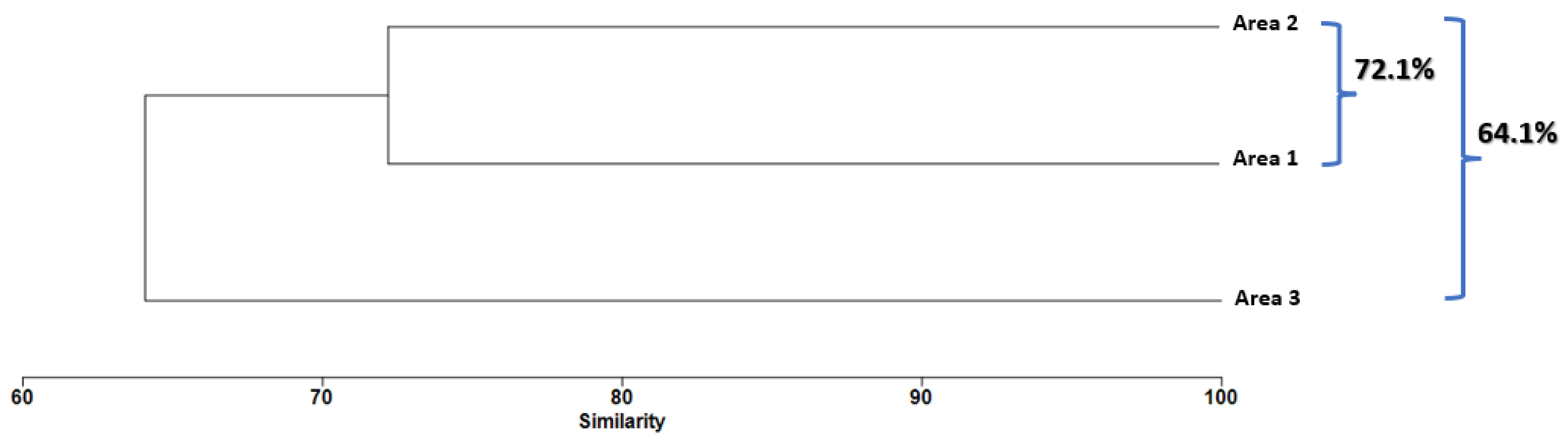
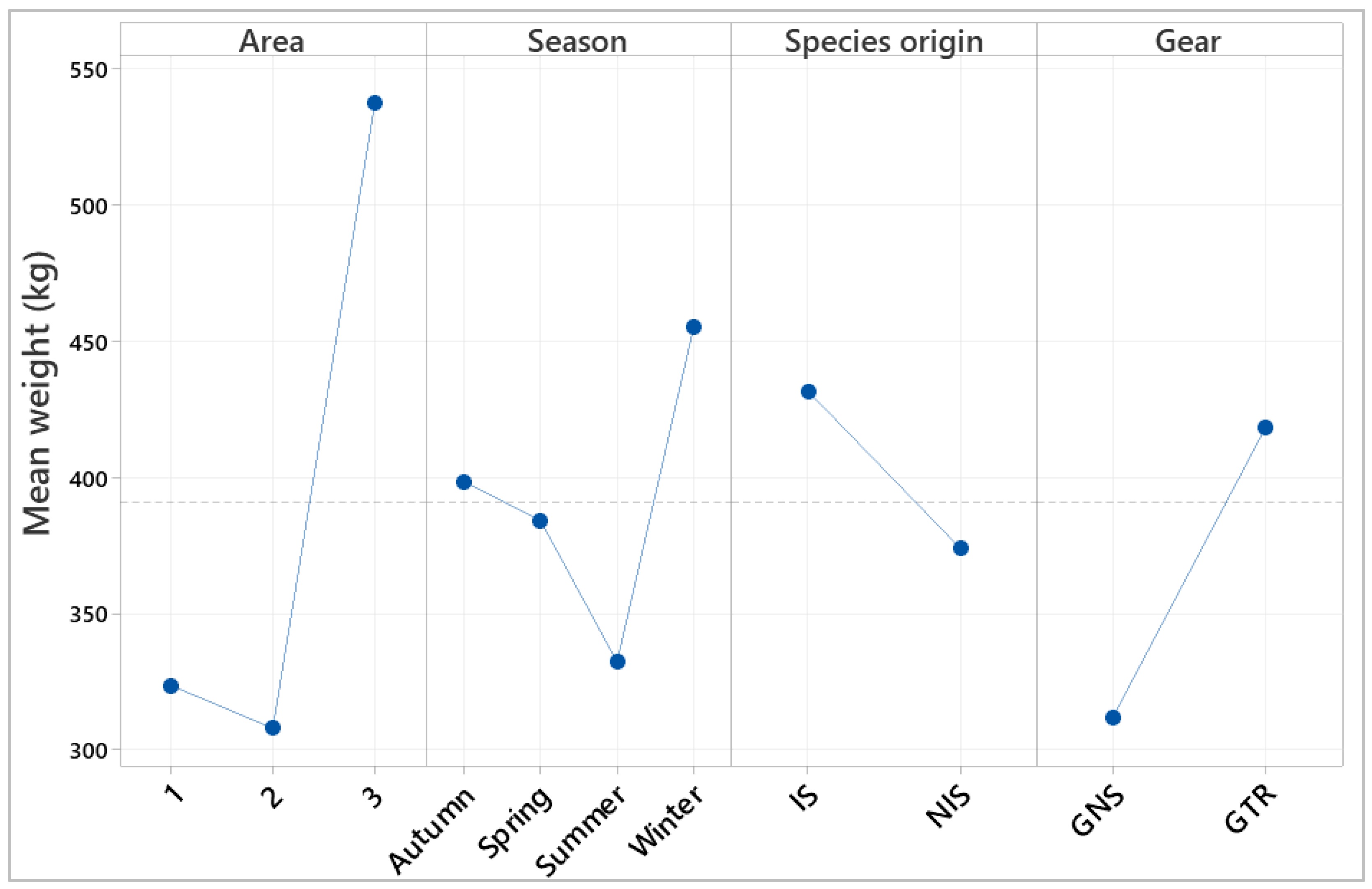
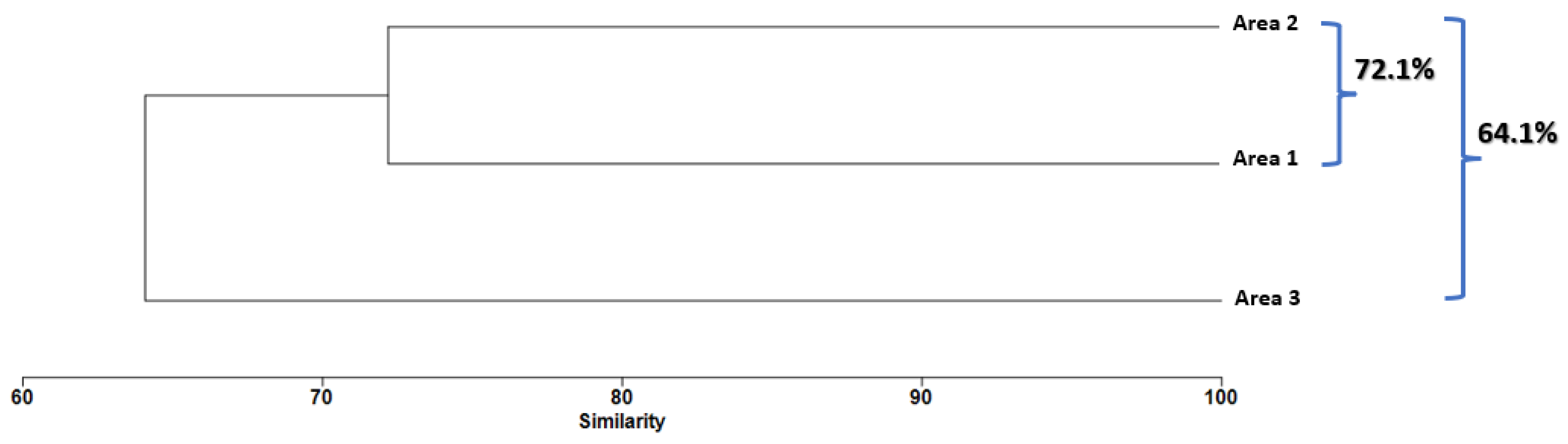
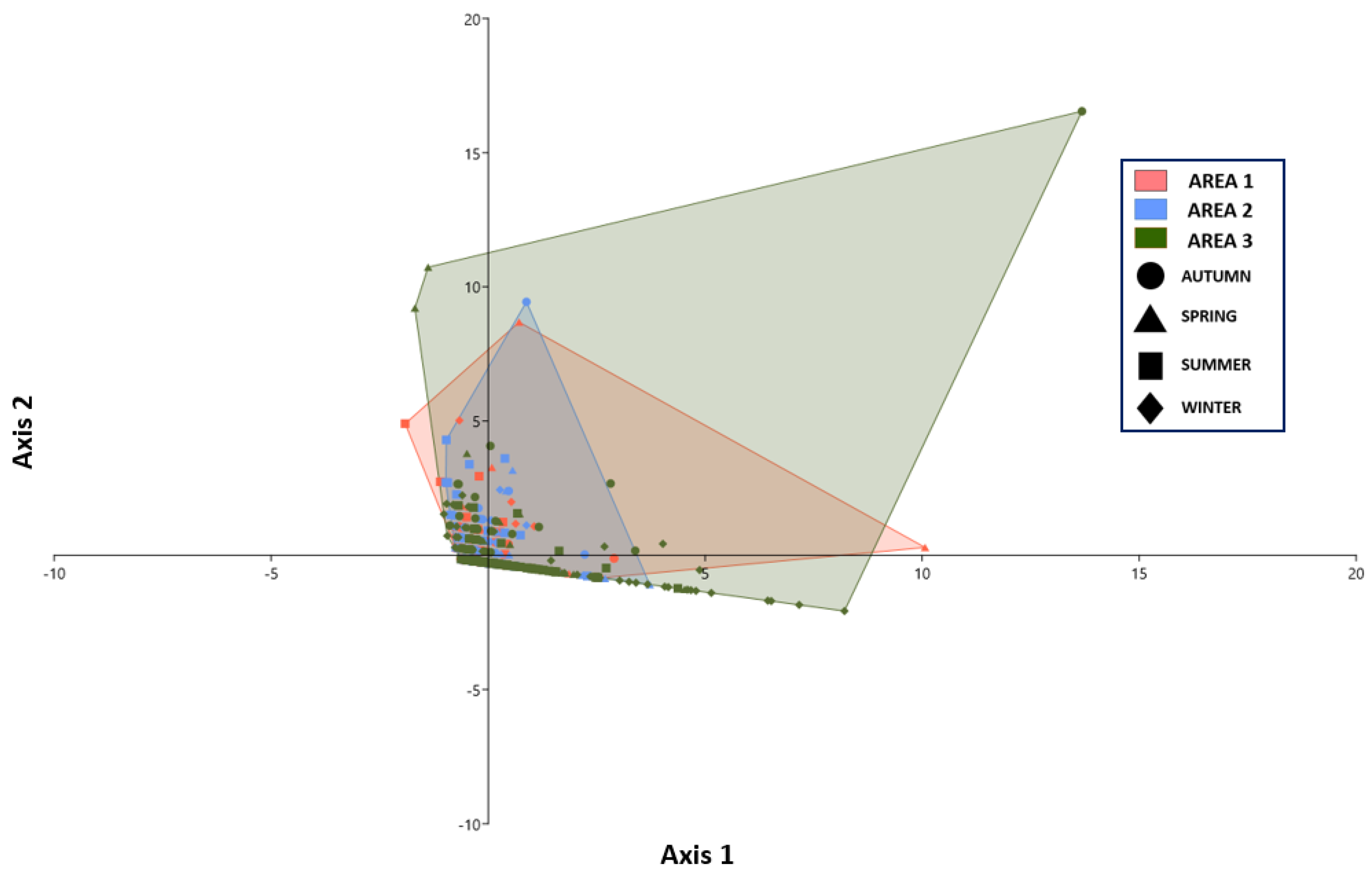
3.5. Spatial Effect Structure
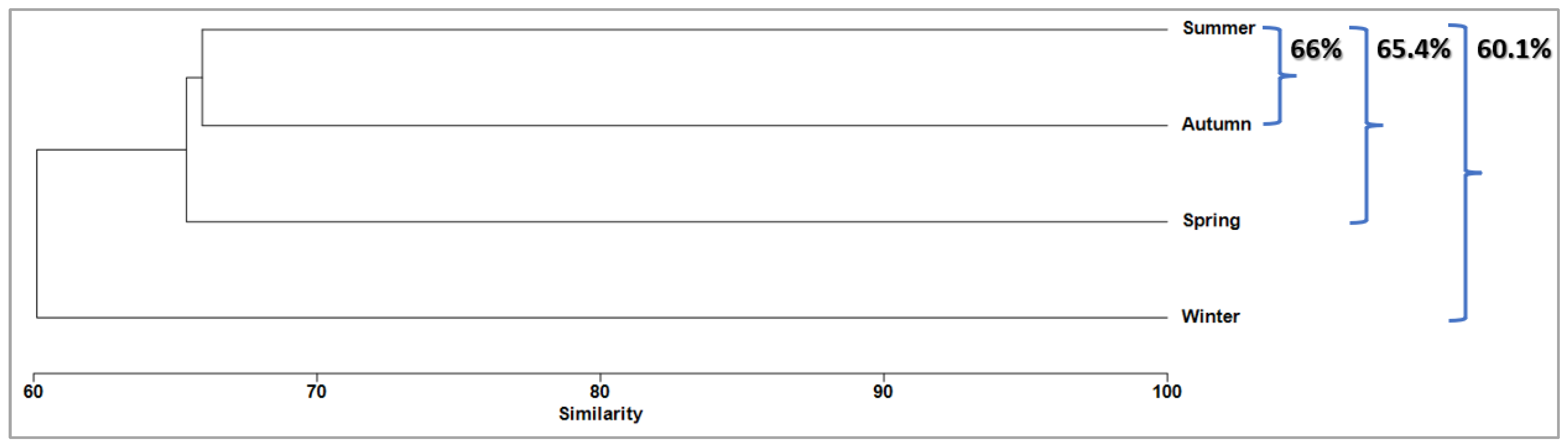
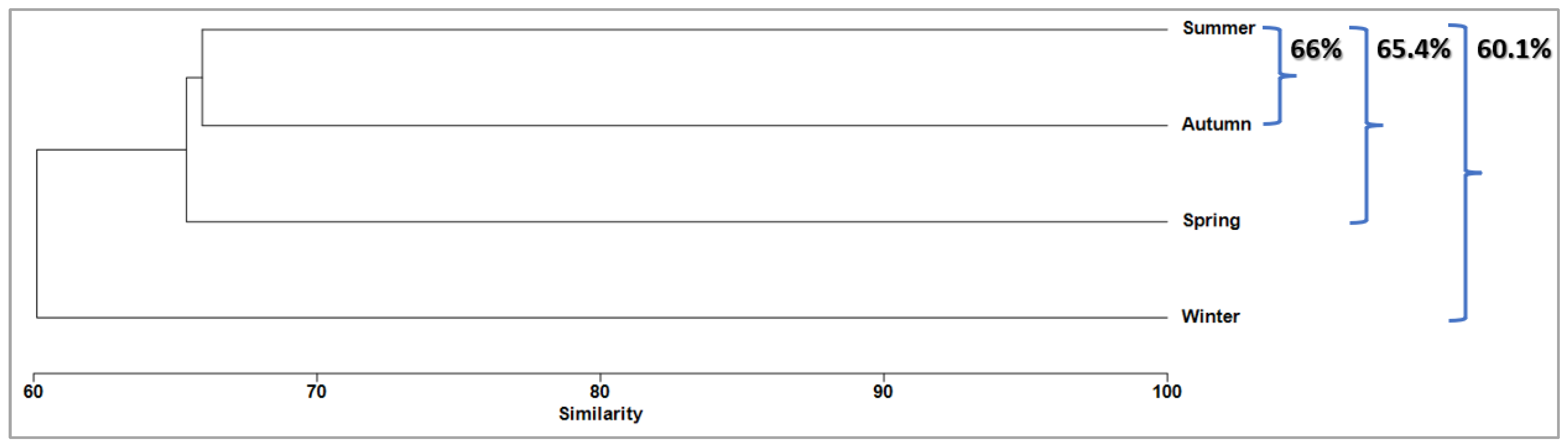
3.6. Temporal Structure
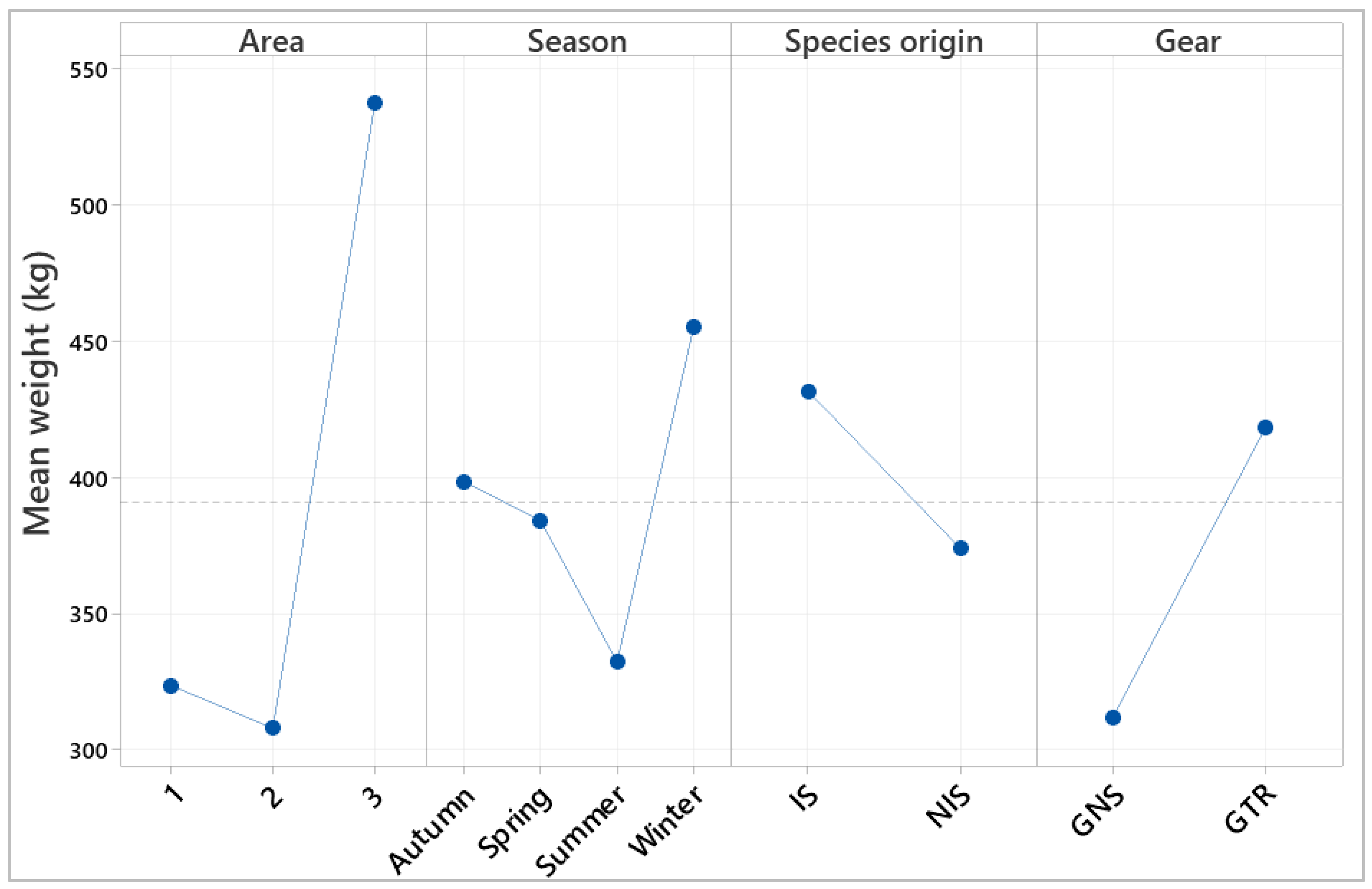
3.7. Factor Interaction
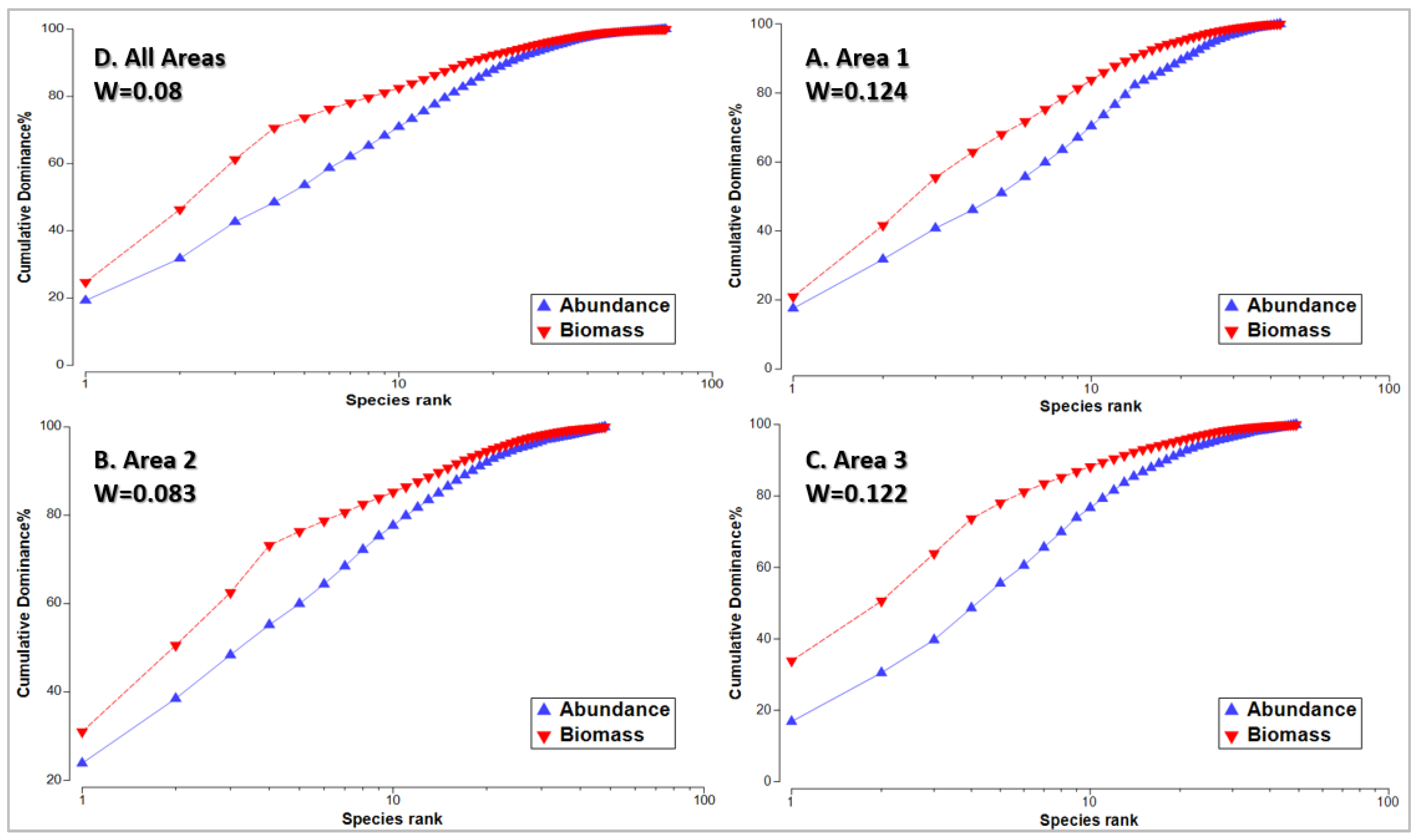
3.8. Ecological Condition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. Life Mediterr. Sea A Look Habitat Changes 2012, 1, 55. [Google Scholar]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C. Management priorities for Marine Invasive Species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Coll, M.; Piroddi, C.; Steenbeek, J.; Ben Rais Lasram, F.; Zenetos, A.; Cardoso, A.C. Invading the Mediterranean Sea: Biodiversity patterns shaped by human activities. Front. Mar. Sci. 2014, 1, 32. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of Invasive Alien Marine Species on Ecosystem Services and Biodiversity: A Pan-European Review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A. Biopollution by Invasive Marine Non-Indigenous Species: A Review of Potential Adverse Ecological Effects in a Changing Climate. Int. J. Environ. Res. Public Health 2021, 18, 4268. [Google Scholar] [CrossRef] [PubMed]
- Arndt, E.; Givan, O.; Edelist, D.; Sonin, O.; Belmaker, J. Shifts in Eastern Mediterranean Fish Communities: Abundance Changes, Trait Overlap, and Possible Competition between Native and Non-Native Species. Fishes 2018, 3, 19. [Google Scholar] [CrossRef]
- Kleitou, P.; Moutopoulos, D.K.; Giovos, I.; Kletou, D.; Savva, I.; Cai, L.L.; Hall-Spencer, J.M.; Charitou, A.; Elia, M.; Katselis, G. Conflicting Interests and Growing Importance of Non-indigenous Species in Commercial and Recreational Fisheries of the Mediterranean Sea. Fish. Manag. Ecol. 2022, 29, 169–182. [Google Scholar] [CrossRef]
- Galanidi, M.; Zenetos, A.; Bacher, S. Assessing the Socio-Economic Impacts of Priority Marine Invasive Fishes in the Mediterranean with the Newly Proposed SEICAT Methodology. Mediterr. Mar. Sci. 2018, 19, 107. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Rilov, G.; Edelist, D. Impacts of Marine Invasive Alien Species on European Fisheries and Aquaculture—Plague or Boon? CIESM Monogr. 2018, 50, 125–132. [Google Scholar]
- Chaffin, B.C.; Garmestani, A.S.; Angeler, D.G.; Herrmann, D.L.; Stow, C.A.; Nyström, M.; Sendzimir, J.; Hopton, M.E.; Kolasa, J.; Allen, C.R. Biological Invasions, Ecological Resilience and Adaptive Governance. J. Environ. Manag. 2016, 183, 399–407. [Google Scholar] [CrossRef]
- Kleitou, P.; Crocetta, F.; Giakoumi, S.; Giovos, I.; Hall-Spencer, J.M.; Kalogirou, S.; Kletou, D.; Moutopoulos, D.K.; Rees, S. Fishery Reforms for the Management of Non-Indigenous Species. J. Environ. Manag. 2021, 280, 111690. [Google Scholar] [CrossRef]
- Ünal, V.; Bodur, H.G. The Socio-Economic Impacts of the Silver-Cheeked Toadfish on Small-Scale Fishers: A Comparative Study from the Turkish Coast. Su Ürünleri Derg. 2017, 34, 119–127. [Google Scholar] [CrossRef]
- Carpentieri, P.; Lelli, S.; Colloca, F.; Mohanna, C.; Bartolino, V.; Moubayed, S.; Ardizzone, G.D. Incidence of Lessepsian Migrants on Landings of the Artisanal Fishery of South Lebanon. Mar. Biodivers. Rec. 2009, 2, e71. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; Garcia, E.L.; Stern, N.; Tsiamis, K.; Galanidi, M. Established Non-Indigenous Species Increased by 40% in 11 Years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23, 196. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East Is East and West Is West? Management of Marine Bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning-Clode, J.; Castriota, L.; Chainho, P.; Comas-Gonzalez, R.; Costa, A.C. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Mavruk, S.; Avsar, D. Non-Native Fishes in the Mediterranean from the Red Sea, by Way of the Suez Canal. Rev. Fish Biol. Fish. 2008, 18, 251–262. [Google Scholar] [CrossRef]
- Korakaki, E.; Legakis, A.; Katsanevakis, S.; Koulelis, P.P.; Avramidou, E.V.; Soulioti, N.; Petrakis, P.V. Invasive Alien Species of Greece. Invasive Alien Species Obs. Issues Around World 2021, 3, 124–189. [Google Scholar]
- Galil, B.S.; Mienis, H.K.; Hoffman, R.; Goren, M. Non-Indigenous Species along the Israeli Mediterranean Coast: Tally, Policy, Outlook. Hydrobiologia 2021, 848, 2011–2029. [Google Scholar] [CrossRef]
- Papaconstantinou, C. Fauna Graeciae. An Updated Checklist of the Fishes in the Hellenic Seas. Monographs on Marine Sciences; Hellenic Centre for Marine Research: Athens, Greece, 2014; p. 340. [Google Scholar]
- Corsini-Foka, M.; Zenetos, A.; Crocetta, F.; Cinar, M.; Kocak, F.; Golani, D.; Katsanevakis, S.; Tsiamis, K.; Cottier-Cook, E.; Froglia, C. Inventory of Alien and Cryptogenic Species of the Dodecanese (Aegean Sea, Greece): Collaborations through COST Action Training School. Manag. Biol. Invasions 2015, 6, 351–366. [Google Scholar] [CrossRef]
- Por, F.D.E. Lessepsian Migration, the Influx of Red Sea Biota into the Mediterranean by Way of the Suez Canal; Springer: Berlin/Heidelberg, Germany, 1978. [Google Scholar]
- Galil, B.S. The Marine Caravan–the Suez Canal and the Erythrean Invasion. In Bridging Divides: Maritime Canals as Invasion Corridors; Springer: Berlin/Heidelberg, Germany, 2006; pp. 207–300. [Google Scholar]
- Golani, D. Colonization of the Mediterranean by Red Sea Fishes via the Suez Canal-Lessepsian Migration. Fish Invasions Mediterr. Sea Chang. Renew. 2010, 145, 188. [Google Scholar]
- Corsini-Foka, M. Current Status of Alien Fishes in Greek Seas. In Fish Invasions of the Mediterranean Sea: Change and Renewal; Pensoft Publishers: Sofia, Bulgaria, 2010; pp. 219–253. [Google Scholar]
- Raitsos, D.E.; Beaugrand, G.; Georgopoulos, D.; Zenetos, A.; Pancucci-Papadopoulou, A.M.; Theocharis, A.; Papathanassiou, E. Global Climate Change Amplifies the Entry of Tropical Species into the Eastern Mediterranean Sea. Limnol. Oceanogr. 2010, 55, 1478–1484. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Corsini-Foka, M.; Morri, C.; Zenetos, A. Thirty Years After-Dramatic Change in the Coastal Marine Habitats of Kos Island (Greece), 1981–2013. Mediterr. Mar. Sci. 2014, 15, 482–497. [Google Scholar] [CrossRef]
- Sisma-Ventura, G.; Yam, R.; Shemesh, A. Recent Unprecedented Warming and Oligotrophy of the Eastern Mediterranean Sea within the Last Millennium. Geophys. Res. Lett. 2014, 41, 5158–5166. [Google Scholar] [CrossRef]
- Zenetos, A.; Karachle, P.K.; Corsini-Foka, M.; Gerovasileiou, V.; Simboura, N.; Xentidis, N.J.; Tsiamis, K. Is the Trend in New Introductions of Marine Non-Indigenous Species a Reliable Criterion for Assessing Good Environmental Status? Τhe Case Study of Greece. Mediterr. Mar. Sci. 2020, 21, 775–793. [Google Scholar] [CrossRef]
- Dragičević, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Dulčić, J.; Edelist, D.; et al. New Mediterranean Biodiversity Records (December 2019). Mediterr. Mar. Sci. 2019, 20, 645–656. [Google Scholar] [CrossRef]
- Kousteni, V.; Bakiu, R.; Benhmida, A.; Crocetta, F.; Di Martino, V.; Dogrammatzi, A.; Doumpas, N.; Durmisha, S.; Giovos, I.; Gokoglu, M.; et al. New Mediterranean Biodiversity Records (April, 2019). Mediterr. Mar. Sci. 2019, 20, 230–247. [Google Scholar]
- Kampouris, T.E.; Doumpas, N.; Giovos, I.; Batjakas, I.E. First Record of the Lessepsian Nemipterus randalli Russell, 1986 (Perciformes, Nemipteridae) in Greece. Cah. Biol. Mar. 2019, 60, 559–561. [Google Scholar]
- Bariche, M.; Al-Mabruk, S.; Ates, M.; Büyük, A.; Crocetta, F.; Dritsas, M.; Edde, D.; Fortic, A.; Gavriil, E.; Gerovasileiou, V. New Alien Mediterranean Biodiversity Records (March 2020). Mediterr. Mar. Sci. 2020, 21, 129–145. [Google Scholar] [CrossRef]
- Ragkousis, M.; Abdelali, N.; Azzurro, E.; Badreddine, A.; Bariche, M.; Bitar, G.; Crocetta, F.; Denitto, F.; Digenis, M.; El Zrelli, R.; et al. New Alien Mediterranean Biodiversity Records (October 2020). Mediterr. Mar. Sci. 2020, 21, 631–652. [Google Scholar]
- Corsini-Foka, M.; Mastis, S.; Kondylatos, G.; Batjakas, I.E. Alien and Native Fish in Gill Nets at Rhodes, Eastern Mediterranean (2014–2015). J. Mar. Biol. Assoc. UK 2017, 97, 635–642. [Google Scholar] [CrossRef]
- Ünal, V.; Göncüoğlu, H.; Durgun, D.; Tosunoğlu, Z.; Deval, C.; Turan, C. Silver-Cheeked Toadfish, Lagocephalus sceleratus (Actinopterygii: Tetraodontiformes: Tetraodontidae), Causes a Substantial Economic Losses in the Turkish Mediterranean Coast: A Call for Decision Makers. Acta Ichthyol. Piscat. 2015, 45, 231–237. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien Species as a Driver of Recent Extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Turan, C. The invasion and Establishment of Lionfish Species (Pterois spp.) as a Major Threat to Marine Biodiversity of Turkey. In Lionfish Invasion Its Management in the Mediterranean Sea; Turkish Marine Research Foundation: Istanbul, Turkey, 2018; pp. 10–16. [Google Scholar]
- Arianoutsou, M.; Adamopoulou, C.; Andriopoulos, P.; Bazos, I.; Christopoulou, A.; Galanidis, A.; Kalogianni, E.; Karachle, P.K.; Kokkoris, Y.; Martinou, A.F. HELLAS-ALIENS. The Invasive Alien Species of Greece: Time Trends, Origin and Pathways. NeoBiota 2023, 86, 45–79. [Google Scholar] [CrossRef]
- GFCM-UNEP/MAP Report of the Joint GFCM-UN Environment/ MAP Subregional Pilot Study for the Eastern Mediterranean on Non-Indigenous Species in Relation to Fisheries. Chania, Greece: Food and Agriculture Organization of the United Nations and General. 2018. Available online: https://www.fao.org/gfcm/technical-meetings/detail/fr/c/1127974/ (accessed on 12 September 2023).
- Koutsouflakis, G.H. Fishing in the Greco-Roman World: Semiotics of Texts and Fishing Iconography, Fishing in the Greek Seas. Res. Noteb. 2010, 33, 55–83. (In Greek) [Google Scholar]
- FAO. GFCM. Available online: https://www.fao.org/gfcm/activities/fisheries/small-scale-fisheries/en/ (accessed on 12 September 2023).
- Gebremedhin, S.; Bruneel, S.; Getahun, A.; Anteneh, W.; Goethals, P. Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management. Water 2021, 13, 574. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture. Sustainability in Action. Rome. 2020. Available online: https://www.fao.org/3/ca9229en/ca9229en.pdf (accessed on 12 September 2023).
- Lloret, J.; Cowx, I.G.; Cabral, H.; Castro, M.; Font, T.; Gonçalves, J.M.S.; Gordoa, A.; Hoefnagel, E.; Matić-Skoko, S.; Mikkelsen, E. Small-Scale Coastal Fisheries in European Seas Are Not What They Were: Ecological, Social and Economic Changes. Mar. Policy 2018, 98, 176–186. [Google Scholar] [CrossRef]
- Tzanatos, E.; Georgiadis, M.; Peristeraki, P. Small-Scale Fisheries in Greece: Status, Problems, and Management. In Small-Scale Fisheries in Europe: Status, Resilience and Governance; Springer: Dodrecht, The Netherlands, 2020; pp. 125–150. [Google Scholar]
- Liontakis, A.; Tzouramani, I.; Mantziaris, S.; Sintori, A. Unravelling the Role of Gender in Fisheries’ Socio-Economic Performance: The Case of Greek Small-Scale Fisheries. Sustainability 2020, 12, 5304. [Google Scholar] [CrossRef]
- Ministry of Rural Development and Food. Annual Fleet Report 2020; Directorate General of Fishing: Athens, Greece, 2021.
- DG MARE. Fleet Register. Available online: https://webgate.ec.europa.eu/fleet-europa/results_en (accessed on 12 September 2023).
- Lucchetti, A.; Virgili, M.; Petetta, A.; Sartor, P. An Overview of Gill Net and Trammel Net Size Selectivity in the Mediterranean Sea. Fish. Res. 2020, 230, 105677. [Google Scholar] [CrossRef]
- Roditi, K.; Vafidis, D. Small-Scale Fisheries in the South Aegean Sea: Métiers and Associated Economics. Ocean Coast. Manag. 2022, 224, 106185. [Google Scholar] [CrossRef]
- Grati, F.; Azzurro, E.; Scanu, M.; Tassetti, A.N.; Bolognini, L.; Guicciardi, S.; Vitale, S.; Scannella, D.; Carbonara, P.; Dragičević, B. Mapping Small-scale Fisheries through a Coordinated Participatory Strategy. Fish Fish. 2022, 23, 773–785. [Google Scholar] [CrossRef]
- Adamidou, A. Commercial Fishing Gears and Methods Used in Hellas. In State of Hellenic Fisheries; HCMR: Athens, Greece, 2007; pp. 118–131. [Google Scholar]
- Olguner, M.T.; Deval, M.C. Catch and Selectivity of 40 and 44 Mm Trammel Nets in Small-Scale Fisheries in the Antalya Bay, Eastern Mediterranean. Ege J. Fish. Aquat. Sci. 2013, 30, 167–173. [Google Scholar]
- Petetta, A.; Herrmann, B.; Virgili, M.; De Marco, R.; Canduci, G.; Veli, D.L.; Bargione, G.; Vasapollo, C.; Lucchetti, A. Estimating Selectivity of Experimental Diamond (T0) and Turned Mesh (T90) Codends in Multi-Species Mediterranean Bottom trawl. Mediterr. Mar. Sci. 2020, 21, 545–557. [Google Scholar] [CrossRef]
- Jacquet, J.; Pauly, D. Funding Priorities: Big Barriers to Small-Scale Fisheries. Conserv. Biol. 2008, 22, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Tzanatos, E.; Castro, J.; Forcada, A.; Matić-Skoko, S.; Gaspar, M.; Koutsikopoulos, C. A Métier-Sustainability-Index (MSI25) to Evaluate Fisheries Components: Assessment of Cases from Data-Poor Fisheries from Southern Europe. ICES J. Mar. Sci. 2013, 70, 78–98. [Google Scholar] [CrossRef]
- Ponte, S.; Raakjær, J.; Campling, L. Swimming Upstream: Market Access for African Fish Exports in the Context of WTO and EU Negotiations and Regulation. Dev. Policy Rev. 2007, 25, 113–138. [Google Scholar] [CrossRef]
- Nilsson, J.A.; Fulton, E.A.; Johnson, C.R.; Haward, M. How to Sustain Fisheries: Expert Knowledge from 34 Nations. Water 2019, 11, 213. [Google Scholar] [CrossRef]
- Kyvelou, S.S.I.; Ierapetritis, D.G. Fisheries Sustainability through Soft Multi-Use Maritime Spatial Planning and Local Development Co-Management: Potentials and Challenges in Greece. Sustainability 2020, 12, 2026. [Google Scholar] [CrossRef]
- Calo, A.; Di Franco, A.; Quattrocchi, F.; Dimitriadis, C.; Ventura, P.; Milazzo, M.; Guidetti, P. Multi-specific Small-scale Fisheries Rely on Few, Locally Essential, Species: Evidence from a Multi-area Study in the Mediterranean. Fish Fish. 2022, 23, 1299–1312. [Google Scholar] [CrossRef]
- Corsini-Foka, M.; Pancucci-Papadopoulou, M.A. Προκαταρκτική Μελέτη της Aλλόχθονης Πανίδας στο Θαλάσσιο Oικοσύστημα του Νοτιοανατολικού Aιγαίου; Τελική Τεχνική Έκθεση: Ρόδος, Greece, 2010; p. 77. (In Greek) [Google Scholar]
- Stergiou, K.I.; Moutopoulos, D.K.; Soriguer, M.C.; Puente, E.; Lino, P.G.; Zabala, C.; Monteiro, P.; Errazkin, L.A.; Erzini, K. Trammel Net Catch Species Composition, Catch Rates and Métiers in Southern European Waters: A Multivariate Approach. Fish. Res. 2006, 79, 170–182. [Google Scholar] [CrossRef]
- Mastis, S.; Corsini-Foka, M.; Kondylatos, G.B.I. Alien and Native Fish in Gill Nets at Rhodes, Eastern Mediterranean (2014–2015). In Proceedings of the 51st European Marine Biology Symposium, Rhodes, Greece, 26–30 September 2016; p. 59. [Google Scholar]
- FAO. Voluntary Guidelines for Securing Sustainable Small-Scale Fisheries. Voluntary Guidelines for Securing Sustainable Small-Scale Fisheries in the Context of Food Security and Poverty Eradication. 2015. Available online: https://www.fao.org/voluntary-guidelines-small-scale-fisheries/en/ (accessed on 12 September 2023).
- Tassetti, A.N.; Galdelli, A.; Pulcinella, J.; Mancini, A.; Bolognini, L. Addressing Gaps in Small-Scale Fisheries: A Low-Cost Tracking System. Sensors 2022, 22, 839. [Google Scholar] [CrossRef] [PubMed]
- Commission Decision (EU) 2017/848 of May 2017 Laying down Criteria and Methodological Standards on Good Environmental Status of Marine Waters and Specifications and Standardised Methods for Monitoring and Assessment, and Repealing Decision 2010/477/EU. Off. J. Eur. Union 2017, 125, 43–74.
- Frid, O.; Belmaker, J. Catch Dynamics of Set Net Fisheries in Israel. Fish. Res. 2019, 213, 1–11. [Google Scholar] [CrossRef]
- Whitehead, P.J.P.; Bauchot, M.L.; Hureau, J.C.; Nielsen, J.; Tortonese, E. Fishes of the North-eastern Atlantic and The Mediterranean. UNESCO 1986, 3, 1287–1293. [Google Scholar]
- Fischer, W.; Bauchot, M.L.; Schneider, M. Fiche FAO D’identification des Especes Pour les Besoins de la Peche, Rev. 1 Mediterranee et Mer Noire (Zone de Peche 37); FAO: Rome, Italy, 1987. [Google Scholar]
- Golani, D.; Orsi-Relini, L.; Massuti, E.; Quignard, J.-P. CIESM Atlas of Exotic Species in the Mediterranean–Vol. 1 Fishes; CIESM Publishers: Monte Carlo, Monaco, 2002; p. 256. [Google Scholar]
- Golani, D.; Öztürk, B.; Başusta, N. Fishes of the Eastern Mediterranean, 1st ed.; Turkish Marine Research Foundation: Istanbul, Turkey, 2006; p. 259. [Google Scholar]
- Golani, D.; Azzurro, E.; Dulčić, J.; Massutí, E.; Orsi-Relini, L. Atlas of Exotic Fishes in the Mediterranean Sea; Ciesm Publishers: Paris, France; Monte Carlo, Monaco, 2021. [Google Scholar]
- Louisy, P. Guide D’identification Des Poissons Marins: Europe De l’ouest et Méditerranée; Editions Eugen Ulmer: Paris, France, 2002. [Google Scholar]
- Şahin, M.; Aybek, E. Jamovi: An Easy to Use Statistical Software for the Social Scientists. Int. J. Assess. Tools Educ. 2019, 6, 670–692. [Google Scholar] [CrossRef]
- Rutherford, A. Introducing ANOVA And ANCOVA: A GLM Approach; Sage: Thousand Oaks, CA, USA, 2001; ISBN 9780761951612. [Google Scholar]
- Warwick, R.M. A New Method for Detecting Pollution Effects on Marine Macrobenthic Communities. Mar. Biol. 1986, 92, 557–562. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric Multivariate Analyses of Changes in Community Structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Shannon, C.; Weiner, W. The Mathematical Theory of Communication; Urban University Illinois Press: Champaign, IL, USA, 1963; 125p. [Google Scholar]
- Margalef, D. Information Theory in Ecology, General Systems. Transl. Mem. Real Acad. Cienc. Artes Barcelona 1958, 32, 373–449. [Google Scholar]
- Pielou, E.C. Mathematical Ecology, 2nd ed.; Wiley and Sons: New York, NY, USA, 1977. [Google Scholar]
- Mika, S.; Ratsch, G.; Weston, J.; Scholkopf, B.; Mullers, K.R. Fisher Discriminant Analysis with Kernels. In Neural Networks for Signal Processing IX, Proceedings of the 1999 IEEE Signal Processing Society Workshop (cat. no. 98th8468), Madison, WI, USA, 25 August 1999; IEEE: Piscataway, NJ, USA, 1999; pp. 41–48. [Google Scholar]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy. The Principles and Practice of Numerical Classification; Freeman W.H. and Company: San Francisco, CA, USA, 1973. [Google Scholar]
- The R Development Core Team. R A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean-the 100 ‘Worst Invasives’ and Their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- García-de-la-Fuente, L.; Fernández-Vázquez, E.; Ramos-Carvajal, C. A Methodology for Analyzing the Impact of the Artisanal Fishing Fleets on Regional Economies: An Application for the Case of Asturias (Spain). Mar. Policy 2016, 74, 165–176. [Google Scholar] [CrossRef]
- Tzanatos, E.; Dimitriou, E.; Katselis, G.; Georgiadis, M.; Koutsikopoulos, C. Composition, Temporal Dynamics and Regional Characteristics of Small-Scale Fisheries in Greece. Fish. Res. 2005, 73, 147–158. [Google Scholar] [CrossRef]
- Christidis, G.; Batziakas, E.; Peristeraki, P.; Kosoglou, I.; Tampakakis, K.; Somarakis, S.; Tserpes, G. The Impact of Lagocephalus sceleratus (Gmelin, 1789) on Small-Scale Fisheries in Crete: Preliminary Results. In Proceedings of the Marine and Inland Waters Research Symposium, Porto Heli, Argolida, Greece, 16–19 September 2022; pp. 357–361. [Google Scholar]
- Azzurro, E.; Stancanelli, B.; Di Martino, V.; Bariche, M. Range expansion of the common lionfish Pterois miles (Bennett, 1828) in the Mediterranean Sea: An unwanted new guest for Italian waters. BioInvasions Rec. 2017, 6, 95–98. [Google Scholar] [CrossRef]
- Batjakas, I.E.; Evangelopoulos, A.; Giannou, M.; Pappou, S.; Papanikola, E.; Atsikvasi, M.; Poursanidis, D.; Gubili, C. Lionfish Diet Composition at Three Study Sites in the Aegean Sea: An Invasive Generalist? Fishes 2023, 8, 314. [Google Scholar] [CrossRef]
- Vagenas, G.; Dogrammatzi, A.; Apostolopoulos, G.; Konida, K.; Nalmpanti, M.; Koufalis, E.; Tsikliras, A.C.; Margaritis, M.K.P. Fishers Perceptions on the Status and Impact of the Alien Fish Siganus luridus and S. rivulatus in the Aegean and Ionian Seas. In Proceedings of the Marine and Inland Waters Research Symposium, Porto Heli, Argolida, Greece, 16–19 September 2022; p. 363. [Google Scholar]
- Stamouli, C.; Akel, E.; Azzurro, E.; Bakiu, R.; Bas, A.; Bitar, G.; Boyaci, Y.; Cakalli, M.; Corsini-Foka, M.; Crocetta, F.; et al. New Mediterranean Biodiversity Records (December 2017). Mediterr. Mar. Sci. 2017, 18, 534–556. [Google Scholar]
- Evagelopoulos, A.; Nikolaou, A.; Michailidis, N.; Kampouris, T.E.; Batjakas, I.E. Progress of the Dispersal of the Alien Goatfish Parupeneus forsskali (Fourmanoir & Guézé, 1976) in the Mediterranean, with Preliminary Information on Its Diet Composition in Cyprus. BioInvasions Rec. 2020, 9, 209–222. [Google Scholar]
- Saemundsson, S.; Tsikliras, A.C.; Chartosia, N. Reproduction and Growth of the Red Sea Goatfish Parupeneus forsskali in Its New Environment (Cyprus, Eastern Mediterranean Sea). Sci. Mar. 2023, 87, e064. [Google Scholar] [CrossRef]
- Giakoumi, S. Distribution Patterns of the Invasive Herbivore Siganus luridus (Rüppell, 1829) and its Relation to Native Benthic Communities in the Central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 2014, 35, 96–105. [Google Scholar] [CrossRef]
- Pennington, A.; Shepperson, J.; Jeffery, A.; Miliou, A.; Anagnostou, V. Siganus Species in an Artisanal Fishery in the Eastern Aegean. Rapp. Comm. Int. Mer Mediterr. 2013, 40, 591. [Google Scholar]
- Stergiou, K.I.; Moutopoulos, D.K.; Erzini, K. Gill Net and Longlines Fisheries in Cyclades Waters (Aegean Sea): Species Composition and Gear Competition. Fish. Res. 2002, 57, 25–37. [Google Scholar] [CrossRef]
- Peristeraki, P.; Skarvelis, K.; Giannakaki, A.; Tambakakis, K.; Tserpes, G. Preliminary Results on the Abundance of Alien Species in the Coastal Fisheries Catches of Crete. In Proceedings of the 11th Panhellenic Symposium on Oceanography and Fisheries, Mytilene, Lesvos Island, Greece, 13–17 May 2015; pp. 149–152. [Google Scholar]
- Damalas, D.; Stamouli, C.; Fotiadis, N.; Kikeri, M.; Kousteni, V.; Mantopoulou-Palouka, D. The Gyaros Island Marine Reserve: A Biodiversity Hotspot in the Eastern Mediterranean Sea. PLoS ONE 2022, 17, e0262943. [Google Scholar] [CrossRef] [PubMed]
- Ragheb, E.; Kamal, R.M.; Hasan, M.W.A. Species Diversity of Gillnet Catches along the Egyptian Mediterranean Coast of Alexandria. Egypt. J. Aquat. Res. 2022, 48, 281–289. [Google Scholar] [CrossRef]
- Kosswig, C. Erythräische Fische Im Mittelmeer Und an Der Grenze Der Ä Gais. In Syllegomena Biol; von Jordans, A., Peus, F., Eds.; Academie Verlag: Festschrift Kleinschmidt, Leipzig, 1950; pp. 203–212. [Google Scholar]
- Bariche, M.; Kajajian, A.; Azzurro, E. Reproduction of the Invasive Bluespotted Cornetfish Fistularia commersonii (Teleostei, Fistulariidae) in the Mediterranean Sea. Mar. Biol. Res. 2013, 9, 169–180. [Google Scholar] [CrossRef]
- Turan, C.; Öztürk, B. First Record of the Lionfish Pterois miles (Bennett 1828) from the Aegean Sea. J. Black Sea/Mediterr. Environ. 2015, 20, 334–388. [Google Scholar]
- Gurlek, M.; Turan, C.; Yağlioğlu, D. Catch Composition of Non-Indigenous Species by Gill and Trammel Net Fishery in the Mediterranean Coast of Turkey. Fish. Aquat. Sci. 2018, 47, 265–269. [Google Scholar]
- Petetta, A.; Vasapollo, C.; Virgili, M.; Bargione, G.; Lucchetti, A. Pots vs Trammel Nets: A Catch Comparison Study in a Mediterranean Small-Scale Fishery. PeerJ 2020, 8, e9287. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of in-vasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Henry, M.; Leung, B.; Cuthbert, R.N.; Bodey, T.W.; Ahmed, D.A.; Angulo, E.; Bal-zani, P.; Briski, E.; Courchamp, F.; Hulme, P.E.; et al. Unveiling the hidden economic toll of biological invasions in the European Union. Environ. Sci. Eur. 2023, 35, 1–16. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Pattison, Z.; Taylor, N.G.; Verbrugge, L.; Diagne, C.; Ahmed, D.A.; Leroy, B.; Angulo, E.; Briski, E.; Capinha, C.; et al. Global economic costs of aquatic invasive alien species. Sci. Total Environ. 2021, 775, 145238. [Google Scholar] [CrossRef]
- Sini, M.; Vatikiotis, K.; Thanopoulou, Z.; Katsoupis, C.; Maina, I.; Kavadas, S.; Karachle, P.K.; Katsanevakis, S. Small-Scale Coastal Fishing Shapes the Structure of Shallow Rocky Reef Fish in the Aegean Sea. Front. Mar. Sci. 2019, 6, 599. [Google Scholar] [CrossRef]
- Tzanatos, E.; Dimitriou, E.; Papaharisis, L.; Roussi, A.; Somarakis, S.; Koutsikopoulos, C. Principal Socio-Economic Characteristics of the Greek Small-Scale Coastal Fishermen. Ocean Coast. Manag. 2006, 49, 511–527. [Google Scholar] [CrossRef]
- Frid, O.; Gavriel, T.; Ben-Ari, Y.; Weinberger, A.; Yancovich-Shalom, H.; Belmaker, J. Catch Estimates and Species Composition of Recreational Fishing in Israel. Fishes 2023, 8, 69. [Google Scholar] [CrossRef]
- Font, T.; Lloret, J. Biological and Ecological Impacts Derived from Recreational Fishing in Mediterranean Coastal Areas. Rev. Fish. Sci. Aquac. 2014, 22, 73–85. [Google Scholar] [CrossRef]
- Tsafoutis, D.; Metaxas, T. Fishing Tourism in Greece: Defining Possibilities and Prospects. Sustainability 2021, 13, 13847. [Google Scholar] [CrossRef]
- González, R.C.L.; Piñeiro Antelo, M. de los Á. Fishing Tourism as an Opportunity for Sustainable Rural Development—The Case of Galicia, Spain. Land 2020, 9, 437. [Google Scholar] [CrossRef]
- Golani, D. First Record of the Bluespotted Cornetfish from the Mediterranean Sea. J. Fish Biol. 2000, 56, 1545–1547. [Google Scholar] [CrossRef]
- Akyol, O.; Ünal, V.; Ceyhan, T.; Bilecenoglu, M. First Confirmed Record of Lagocephalus sceleratus (Gmelin, 1789) in the Mediterranean Sea. J. Fish Biol. 2005, 66, 1183–1186. [Google Scholar] [CrossRef]
- Golani, D.; Sonin, O. New Records of the Red Sea Fishes, Pterois miles (Scorpaenidae) and Pteragogus pelycus (Labridae) from the Eastern Mediterranean Sea. Jpn. J. Ichthyol. 1992, 39, 167–169. [Google Scholar] [CrossRef]
- Spanier, E.; Pisanty, S.; Tom, M.; Almog-Shtayer, G. The Fish Assemblage on a Coralligenous Shallow Shelf off the Mediterranean Coast of Northern Israel. J. Fish Biol. 1989, 35, 641–649. [Google Scholar] [CrossRef]
- Shakman, E.A.; Kinzelbach, R. Distribution and characterization of Lessepsian migrant fishes along the coast of Libya. Acta Ichthyol. Piscat. 2007, 37, 7–15. [Google Scholar] [CrossRef]
- Shakman, E.; Kinzelbach, R. Commercial Fishery and Fish Species Composition in Coastal Waters of Libya. Rostocker Meeresbiol. Beiträge 2007, 18, 63–78. [Google Scholar]
- Kavalakis, G. Composition of Fish Catch in Dodecanese, Technical Report; Fishery Office of the Dodecanese: Rhodes, Greece, 1964; 19p. (In Greek) [Google Scholar]
- Tortonese, E. Biological Investigations in the Aegean Sea. Nature 1947, 159, 887–888. [Google Scholar] [CrossRef]
- Maldura, L.M. La Pesca Nelle Isole Italiane dell’Egeo. Boll. Pesca Piscic. Idrobiol. 1938, 14, 460–480. [Google Scholar]
- Papaconstantinou, C.; Mytilineou, C.; Panos, T. Aspects of the Life History and Fishery of Red Pandora, Pagellus erythrinus (Sparidae) off Western Greece. Cybium 1988, 12, 267–280. [Google Scholar]
- Kalogirou, S.; Wennhage, H.; Pihl, L. Non-Indigenous Species in Mediterranean Fish Assemblages: Contrasting Feeding Guilds of Posidonia oceanica Meadows and Sandy Habitats. Estuar. Coast. Shelf Sci. 2012, 96, 209–218. [Google Scholar] [CrossRef]
- Tzanatos, E.; Somarakis, S.; Tserpes, G.; Koutsikopoulos, C. Identifying and Classifying Small-Scale Fisheries Métiers in the Mediterranean: A Case Study in the Patraikos Gulf, Greece. Fish. Res. 2006, 81, 158–168. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area 1 | Area 2 | Average Dissimilarity: 59.19 | ||
|---|---|---|---|---|
| Species | Average Abundance | Average Abundance | Contribution % | Cumulative Contribution % |
| Lagocephalus sceleratus | 35.93 | 39.18 | 9.70 | 9.70 |
| Fistularia commersonii | 42.97 | 28.82 | 8.90 | 18.60 |
| Pterois miles | 46.88 | 55.58 | 7.28 | 25.88 |
| Sparisoma cretense | 31.19 | 32.33 | 4.60 | 30.48 |
| Siganus rivulatus | 17.9 | 14.66 | 4.19 | 34.67 |
| Area 1 | Area 3 | Average Dissimilarity: 63.68 | ||
| Lagocephalus sceleratus | 35.93 | 63.41 | 11.05 | 11.05 |
| Fistularia commersonii | 42.97 | 42.69 | 8.46 | 19.51 |
| Pterois miles | 46.88 | 51.97 | 7.11 | 26.62 |
| Sparisoma cretense | 31.19 | 32.54 | 4.98 | 31.60 |
| Dasyatis pastinaca | 10.29 | 22.61 | 4.27 | 35.87 |
| Area 2 | Area 3 | Average Dissimilarity: 62.43 | ||
| Lagocephalus sceleratus | 39.18 | 63.41 | 10.74 | 10.74 |
| Fistularia commersonii | 28.82 | 42.69 | 8.19 | 18.93 |
| Pterois miles | 55.58 | 51.97 | 7.93 | 26.85 |
| Sparisoma cretense | 32.33 | 32.54 | 5.99 | 32.84 |
| Dasyatis pastinaca | 0 | 22.61 | 3.89 | 36.73 |
| Species | Average Abundance | Average Similarity | Contribution % | Cumulative Contribution % |
|---|---|---|---|---|
| Autumn | Average Similarity: 26.75 | |||
| Sparisoma cretense | 51.73 | 7.01 | 26.21 | 26.21 |
| Lagocephalus sceleratus | 20.52 | 4.15 | 15.52 | 41.73 |
| Parupeneus forsskali | 13.68 | 2.94 | 10.99 | 52.72 |
| Sargocentron rubrum | 15.03 | 2.04 | 7.62 | 60.34 |
| Pagellus erythrinus | 12.64 | 1.88 | 7.01 | 67.35 |
| Dasyatis pastinaca | 22.04 | 1.69 | 6.3 | 73.65 |
| Spring | Average Similarity: 44.89 | |||
| Lagocephalus sceleratus | 53.25 | 9.8 | 21.83 | 21.83 |
| Pterois miles | 54.5 | 9.33 | 20.79 | 42.63 |
| Spicara maena | 28.91 | 5.92 | 13.19 | 55.82 |
| Sparisoma cretense | 29.2 | 5.18 | 11.55 | 67.37 |
| Fistularia commersonii | 38.85 | 2.48 | 5.53 | 72.89 |
| Summer | Average Similarity: 33.69 | |||
| Pterois miles | 83.57 | 16.44 | 48.8 | 48.8 |
| Lagocephalus sceleratus | 43.1 | 4.87 | 14.46 | 63.26 |
| Sargocentron rubrum | 20.54 | 3.35 | 9.95 | 73.2 |
| Winter | Average Similarity: 24.12 | |||
| Fistularia commersonii | 50.47 | 4.2 | 17.42 | 17.42 |
| Trachinus araneus | 17.51 | 3.33 | 13.81 | 31.23 |
| Lagocephalus sceleratus | 67.81 | 2.39 | 9.9 | 41.13 |
| Synodus saurus | 11.61 | 2.07 | 8.57 | 49.7 |
| Oblada melanura | 7.46 | 1.65 | 6.84 | 56.55 |
| Pterois miles | 31.98 | 1.56 | 6.48 | 63.02 |
| Diplodus vulgaris | 10.89 | 1.41 | 5.83 | 68.85 |
| Sparisoma cretense | 20.63 | 1.27 | 5.28 | 74.13 |
| A. Gear–Area Interaction | SS | DFs | F | p |
|---|---|---|---|---|
| Model | 15,249,493.09 | 5 | 8.74 | *** |
| Gear | 1,703,481.15 | 1 | 4.88 | * |
| Area | 4,359,783.85 | 2 | 6.24 | ** |
| Gear × area | 1,588,092.45 | 2 | 2.27 | ns |
| Residuals | 385,064,808.8 | 1103 | ||
| Total | 400,314,301.9 | 1108 | ||
| B. Gear–Provenance Interaction | ||||
| Model | 6,460,827.76 | 3 | 6.04 | *** |
| Gear | 4,551,604.13 | 1 | 12.77 | *** |
| Species origin | 989,154.66 | 1 | 2.77 | ns |
| Gear × species origin | 1,912,391.91 | 1 | 5.36 | * |
| Residuals | 393,584,959 | 1104 | ||
| Total | 400,045,786.7 | 1107 | ||
| C. Provenance–Area Interaction | ||||
| Model | 13,408,652.74 | 5 | 7.64 | *** |
| Species origin | 804,105.5 | 1 | 2.29 | ns |
| Area | 10,734,151.97 | 2 | 15.3 | *** |
| Species origin × area | 212,249.94 | 2 | 0.3 | ns |
| Residuals | 386,637,134 | 1102 | ||
| Total | 400,045,786.7 | 1107 | ||
| D. Provenance–Season Interaction | ||||
| Model | 9,067,715.72 | 7 | 3.64 | *** |
| Species origin | 955,362.57 | 1 | 2.69 | ns |
| Season | 1,656,432.88 | 3 | 1.55 | ns |
| Species origin × season | 6,172,448.55 | 3 | 5.79 | *** |
| Residuals | 390,978,071 | 1100 | ||
| Total | 400,045,786.7 | 1107 |
| Group | Species Number | Numerical Abundance | Species Richness Margalef (d) | Evenness (J) | Shannon Diversity Index (H′) | Simpson 1-Lambda′ |
|---|---|---|---|---|---|---|
| Area 1 | 43 | 598 | 6.569 | 0.8009 | 3.012 | 0.9244 |
| Area 2 | 48 | 587 | 7.373 | 0.7301 | 2.826 | 0.8974 |
| Area 3 | 49 | 695 | 7.335 | 0.7565 | 2.944 | 0.9208 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondylatos, G.; Vagenas, G.; Kalaentzis, K.; Mavrouleas, D.; Conides, A.; Karachle, P.K.; Corsini-Foka, M.; Klaoudatos, D. Exploring the Structure of Static Net Fisheries in a Highly Invaded Region: The Case of Rhodes Island (Eastern Mediterranean). Sustainability 2023, 15, 14976. https://doi.org/10.3390/su152014976
Kondylatos G, Vagenas G, Kalaentzis K, Mavrouleas D, Conides A, Karachle PK, Corsini-Foka M, Klaoudatos D. Exploring the Structure of Static Net Fisheries in a Highly Invaded Region: The Case of Rhodes Island (Eastern Mediterranean). Sustainability. 2023; 15(20):14976. https://doi.org/10.3390/su152014976
Chicago/Turabian StyleKondylatos, Gerasimos, Georgios Vagenas, Konstantinos Kalaentzis, Dimitris Mavrouleas, Alexios Conides, Paraskevi K. Karachle, Maria Corsini-Foka, and Dimitris Klaoudatos. 2023. "Exploring the Structure of Static Net Fisheries in a Highly Invaded Region: The Case of Rhodes Island (Eastern Mediterranean)" Sustainability 15, no. 20: 14976. https://doi.org/10.3390/su152014976
APA StyleKondylatos, G., Vagenas, G., Kalaentzis, K., Mavrouleas, D., Conides, A., Karachle, P. K., Corsini-Foka, M., & Klaoudatos, D. (2023). Exploring the Structure of Static Net Fisheries in a Highly Invaded Region: The Case of Rhodes Island (Eastern Mediterranean). Sustainability, 15(20), 14976. https://doi.org/10.3390/su152014976