
Current Utilization and Further Application of Zooplankton Indices for Ecosystem Health Assessment of Lake Ecosystems

 ,
,  and
and
Abstract
1. Introduction
1.1. Aquatic Ecosystem Health and Sustainable Aquatic Ecosystem Management
1.2. Health Assessment of Aquatic Ecosystems and Application of Biotic Indices


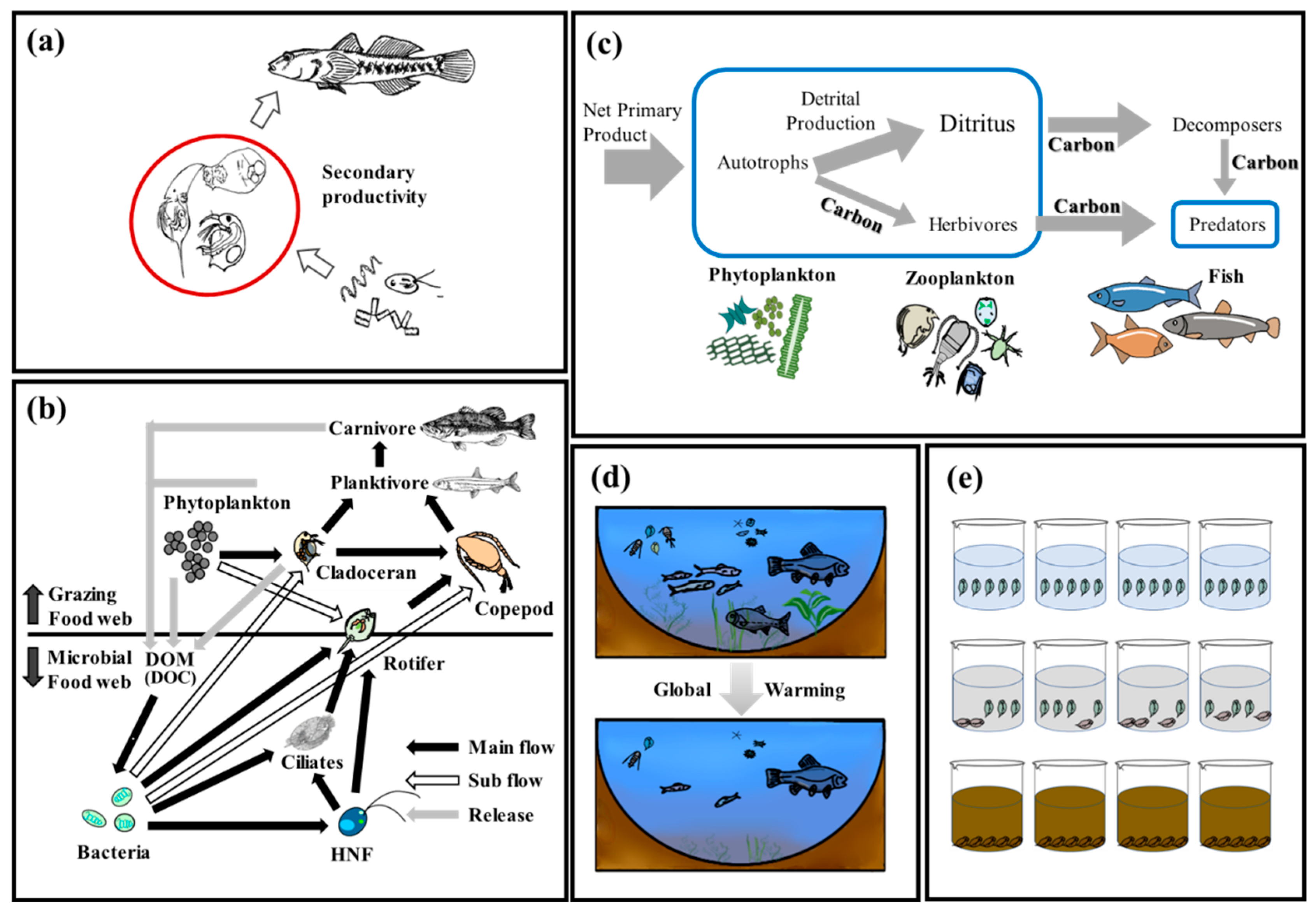
2. Zooplankton as Bioindicators for Monitoring and Assessing Lake Ecosystem Health
2.1. The Role of Zooplankton in Lake Ecosystems
2.2. Zooplankton Communities’ Response Patterns to Environmental Change

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zooplankton Indicators | Related Environment Factors | Response to Increase of Factors | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Eutrophication | Chl-a | TP | Water Hardness | Productive Land Use | Fish Biomass | Water Temperature/Latitude | |||
| Total Zooplankton *** | ● | - | ● | - | - | - | - | Decrease | [43,60] |
| Mean body size | ● | - | - | - | - | - | - | Decrease | [43] |
| Rotifera *,** | - | ● | ● | - | ● | - | - | Increase | [46,62,64] |
| Small Rotifera * (<0.2 mm) | - | - | - | ● | - | - | - | Decrease | [61] |
| Small Cladocera ** | - | ● | ● | - | ● | - | - | Increase | [46,62,63] |
| Large Cladocera *,*** | - | - | - | ● | - | - | - | Increase | [61] |
| - | ● | ● | - | ● | - | - | Decrease | [46,62,63,64] | |
| Cladocera body weight | - | - | ● | - | - | ● | - | Decrease | [60] |
| Cladocera mean body size | - | - | - | - | - | - | ● | Increase | [61] |
| Daphnia spp./ Cladocera ** | - | - | ● | - | - | - | - | Decrease | [60] |
| Cyclpoida *,** | - | ● | ● | - | ● | - | - | Increase | [46,62,64] |
| Cyclopoida/ Copepoda *,** | - | - | ● | - | - | - | - | Increase | [60] |
| Calanoida *** | - | ● | ● | - | ● | - | - | Decrease | [46,62,64] |
| Calanoida/ Copepoda *,** | - | - | ● | - | - | - | - | Decrease | [60] |
2.3. Zooplankton Indices for Freshwater Ecosystem Health Assessment
| Zooplankton Metrics | Description | Parameter | Reference |
|---|---|---|---|
| Ratio of large Cladocera frequently appearing in healthy lake | x = Large Cladocera (>0.5 mm) individual number y = Cladocera individual number | [12] | |
| Effects of zooplankton predation on phytoplankton | a = Cladocera and copepod biomass b = Chlorophyll a concentration | ||
| Eutrophic < 3 3 ≤ Mesotrophic ≤ 4 4 < Oligotrophic | = June biomass of edible algae taxa metric score = June% Mycrocystis, Anabaena, Aphanizomenon of total phytoplankton biomass metric score = June zooplankton ratio (Calanoida/(Cladocera + Cyclopoida)) metric score = July Limnocalanus macrurus density metric score = August zooplankton ratio (Calanoida/(Cladocera + Cyclopoida)) metric score = August Crustacea zooplankton biomass metric score M = Number of metrics S = Number of sites (within a basin) B = Number of basins | [47] | |
| A lake trophic state evaluation index using the Rotifera community | N = Rotifera numbers (ind./L) B = Total biomass (mg w.wt./L) BAC = Percentage of bacterivores in total numbers (%) TECTA = Percentage of form tecta in the population of Keratella cochlearis (%) B:N = Ratio of biomass to numbers (mg w.wt./ind.) IHT = Percentage of species indicative of high trophy in the indicative group’s numbers (%) | [34] | |
| A lake trophic state evaluation index using the Crustacea community | N = Numbers of Crustacea (ind./L) B = Biomass of Cyclopoida (mg w.wt./L) CB = Percentage of Cyclopoida biomass in total biomass of Crustacea (%) CY/CL = Ratio of the Cyclopoida biomass to the biomass of Cladocera CY/CA = Ratio of Cyclopoida to Calanoida numbers IHT = Percentage of species indicative of high trophy in the indicative group’s numbers (%) | [38] | |
| An index that measures the ecological water quality of a lake by combining the dry biomass of plankton Low GP values: high zooplankton biomass dominated High GP values: increased phytoplankton biomass | B = dry biomass(mg/L) ROT = Rotifera CLAD = Cladocera COP = Copepoda CYANO = Cyanobacteria CHLORO = Chlorophyta CHRYSO = Chrysophyta CHYPTO = Cryptophyta PRYMNESIO = Prymnesiophyta DIATOMS = Bacillariophyta DINO = Dinophyta CONJ = Conjugatophyta | [48] | |
| bad ≤ 0.189 0.189 < poor ≤ 0.376 0.377 ≤ moderate ≤ 0.565 0.566 ≤ good ≤ 0.754 0.755 ≤ High | CA/CY = Ratio of Calanoida to Cyclopoida individual numbers(ind./L) NZOL = Zooplankton abundance(ind./L) TECTA = Percentage of form tecta in the population of Keratella cochlearis(%) IHTROT = Percentage of species indicative of high trophy in the indicative group’s number (%) D = Margalef’s diversity index | [21] | |
| Bad ≤ 6 6 < Poor ≤ 10 10 < Moderate ≤ 14 14 < Good ≤ 18 18 < High | = Abundance (ind./L) = Biomass (μg/L) = Mean body size (ind./μg) = Cladocera ratio | [49] |
3. Proposing Perspectives for Advancing Zooplankton Indices
3.1. Application of Biomass in Calculating Zooplankton Index

| Classification | Formula | ||
|---|---|---|---|
| Cladocera | ln W = ln a + b × ln L | ln a, b = species specific constants | |
| Copepoda * | |||
| Rotifera | Basic | W = {(L3 × FF) + (%BV × L3 × FF)} × 10−6 × WW:DW | w = the width measurement (μm) FF = species specific formula factor %BV = a percent of the volume of appendages to biovolume 10−6 = conversion to wet weight; assuming a density of 1) WW:DW ** = conversion to dry weight from wet weight |
| Collotheca | W = (w3 × FF) × 10−6 × WW:DW | ||
| Filinia Trichocerca Conochilus Conochiloides | W = {(L × w2 × FF) + (%BV × L × w2 × FF)} × 10−6 × WW:DW | ||
3.2. Application of eDNA to the Development of Zooplankton Indices
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schaeffer, D.J.; Herricks, E.E.; Kerster, H.W. Ecosystem health: I. Measuring ecosystem health. Environ. Manag. 1988, 12, 445–455. [Google Scholar] [CrossRef]
- Rapport, D.J. What constitutes ecosystem health? Perspect. Biol. Med. 1989, 33, 120–132. [Google Scholar] [CrossRef]
- Ulanowicz, R.E. Ecosystem health and trophic flow networks. In Ecosystem Health: New Goals for Environmental Management; Island Press: Washington, DC, USA, 1992; pp. 190–206. [Google Scholar]
- Rapport, D.J.; Costanza, R.; McMichael, A.J. Assessing ecosystem health. Trends Ecol. Evol. 1998, 13, 397–402. [Google Scholar] [CrossRef]
- Xu, F.L.; Tao, S.; Dawson, R.W.; Li, P.G.; Cao, J. Lake ecosystem health assessment: Indicators and methods. Water Res. 2001, 35, 3157–3167. [Google Scholar] [CrossRef] [PubMed]
- Rapport, D.J. The health of ecology and the ecology of health. Hum. Ecol. Risk Assess. 2002, 8, 205–213. [Google Scholar] [CrossRef]
- Davis, W.S.; Simon, T.P. (Eds.) Biological Assessment and Criteria: Tools for Water Resource Planning and Decision Making; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Barbour, M.T. Rapid Bioassessment Protocols for Use in Wadeable Streams and Rivers: Periphyton, Benthic Macroinvertebrates and Fish; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. [Google Scholar]
- Klemm, D.J.; Blocksom, K.A.; Fulk, F.A.; Herlihy, A.T.; Hughes, R.M.; Kaufmann, P.R.; Peck, P.R.; Stoddard, J.L.; Thoeny, W.T.; Griffith, M.B.; et al. Development and Evaluation of a Macroinvertebrate Biotic Integrity Index(MBII) for Regionally Assessing Mid-Atlantic Highlands Streams. Environ. Manag. 2003, 31, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Kolpin, D.W.; Barbash, J.E.; Gilliom, R.J. Occurrence of pesticides in shallow groundwater of the United States: Initial results from the National Water-Quality Assessment Program. Environ. Sci. Technol. 1998, 32, 558–566. [Google Scholar] [CrossRef]
- Directive, W.F. Water framework directive. J. Ref. OJL 2000, 327, 1–73. [Google Scholar]
- Moss, B.; Stephen, D.; Alvarez, C.; Becares, E.; Bund, W.V.D.; Collings, S.E.; Donk, E.V.; Eyto, E.D.; Feldmann, T.; Fernández-Aláez, C.; et al. The determination of ecological status in shallow lakes—A tested system (ECOFRAME) for implementation of the European Water Framework Directive. Aquat. Conserv. Mar. Freshw. 2003, 13, 507–549. [Google Scholar] [CrossRef]
- US EPA. National Lakes Assessment: A Collaborative Survey of the Nation’s Lakes; Environmental Protection Agency, Office of Water and Office of Research and Development: Washington, DC, USA, 2009; p. 118. [Google Scholar]
- US EPA. National Wetland Condition Assessment 2001: Field Operations Manual; Environmental Protection Agency: Washington, DC, USA, 2011. [Google Scholar]
- US EPA. National Coastal Condition Assessment 2010; Environmental Protection Agency, Office of Water and Office of Research and Development: Washington, DC, USA, 2015; p. 129. [Google Scholar]
- US EPA. National Rivers and Streams Assessment 2008–2009: A Collaborative Survey; Environmental Protection Agency, Office of Water and Office of Research and Development: Washington, DC, USA, 2016; p. 131. [Google Scholar]
- Minshall, G.W. Towards a theory of macroinvertebrate community structure in stream ecosystems. Archiv. Hydrobiol. 1985, 104, 49–76. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342, 151–164. [Google Scholar] [CrossRef]
- Zaghloul, A.; Saber, M.; Gadow, S.; Awad, F. Biological indicators for pollution detection in terrestrial and aquatic ecosystems. Bull. Natl. Res. Cent. 2020, 44, 1–11. [Google Scholar] [CrossRef]
- Karr, J.R. Assessing Biological Integrity in Running Waters: A Method and Its Rationale; Illinois Natural History Survey Special Publication No. 05; Illinois Natural History Survey: Champaign, IL, USA, 1986. [Google Scholar]
- Ochocka, A. ZIPLAS: Zooplankton Index for Polish Lakes’ Assessment: A new method to assess the ecological status of stratified lakes. Environ. Monit. Assess. 2021, 193, 664. [Google Scholar] [CrossRef]
- Blew, R.D. On the definition of ecosystem. Bull. Ecol. Soc. Am. 1996, 77, 171–173. Available online: https://www.jstor.org/stable/20168066 (accessed on 9 July 2023).
- Morse, J.C.; Bae, Y.J.; Munkhjargal, G.; Sangpradub, N.; Tanida, K.; Vshivkova, T.S.; Wang, B.; Yang, L.; Yule, C.M. Freshwater biomonitoring with macroinvertebrates in East Asia. Front. Ecol. Environ. 2007, 5, 33–42. [Google Scholar] [CrossRef]
- Allen, H.L. Primary productivity, chemo-organotrophy, and nutritional interactions of epiphytic algae and bacteria on macrophytes in the littoral of a lake. Ecol. Monogr. 1971, 41, 97–127. [Google Scholar] [CrossRef]
- Welti, N.; Striebel, M.; Ulseth, A.J.; Cross, W.F.; DeVilbiss, S.; Glibert, P.M.; Guo, L.; Hirst, A.G.; Hood, J.; Kominoski, J.S.; et al. Bridging food webs, ecosystem metabolism, and biogeochemistry using ecological stoichiometry theory. Front. Microbiol. 2017, 8, 1298. [Google Scholar] [CrossRef]
- Jeppesen, E.; Noges, P.; Davidson, T.A.; Haberman, J.; Noges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes: A scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Mehner, T.; Keeling, C.; Emmrich, M.; Holmgren, K.; Argillier, C.; Volta, P.; Winfield, I.J.; Brucet, S. Effects of fish predation on density and size spectra of prey fish communities in lakes. Can. J. Fish. Aquat. Sci. 2016, 73, 506–518. [Google Scholar] [CrossRef]
- Park, K.S.; Shin, H.W. Studies on phyto-and-zooplankton composition and its relation to fish productivity in a west coast fish pond ecosystem. J. Environ. Biol. 2007, 28, 415. [Google Scholar]
- Bruce, L.C.; Hamilton, D.; Imberger, J.; Gal, G.; Gophen, M.; Zohary, T.; Hambright, K.D. A numerical simulation of the role of zooplankton in C, N and P cycling in Lake Kinneret, Israel. Ecol. Model. 2006, 193, 412–436. [Google Scholar] [CrossRef]
- Korponai, J.; Braun, M.; Forró, L.; Gyulai, I.; Kövér, C.; Nédli, J.; Urák, I.; Buczkó, K. Taxonomic, functional and phylogenetic diversity: How subfossil cladocerans mirror contemporary community for ecosystem functioning: A comparative study in two oxbows= Sesgos en la diversidad taxonómica, funcional y filogenética de las comunidades vivas y subfósiles de cladóceros. Limnetica 2019, 38, 431–456. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G. Limnological Analyses; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000. [Google Scholar]
- May, L.; O’Hare, M. Changes in rotifer species composition and abundance along a trophic gradient in Loch Lomond, Scotland, UK. In Rotifera X: Rotifer Research: Trends, New Tools and Recent Advances, Proceedings of the Xth International Rotifer Symposium, held in Illmitz, Austria, 7–13 June 2003; Springer: Dordrecht, The Netherlands, 2005; pp. 397–404. [Google Scholar] [CrossRef]
- Duggan, I.C.; Green, J.D.; Shiel, R.J. Distribution of rotifers in North Island, New Zealand, and their potential use as bioindicators of lake trophic state. In Rotifera IX: Proceedings of the IXth International Rotifer Symposium, Khon Kaen, Thailand, 16–23 January 2000; Springer: Dordrecht, The Netherlands, 2001; pp. 155–164. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. The usefulness of zooplankton as lake ecosystem indicators: Rotifer trophic state index. Pol. J. Ecol. 2012, 60, 339–350. [Google Scholar]
- Ichise, S.; Wakabayashi, T. Illustrated Handbook of Freshwater Plankton in Japan; Godo Shuppan: Tokyo, Japan, 2005; pp. 116–117, 125. [Google Scholar]
- Dodson, S.I.; Frey, D.G. Chapter 22 Cladocera and other Branchiopoda. In Ecology and Classification of North American Freshwater-Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Chen, G.; Dalton, C.; Taylor, D. Cladocera as indicators of trophic state in Irish lakes. J. Paleolimnol. 2010, 44, 465–481. [Google Scholar] [CrossRef]
- Tumurtogoo, U.; Figler, A.; Korponai, J.; Sajtos, Z.; Grigorszky, I.; Berta, C.; Gyulai, I. Density and Diversity Differences of Contemporary and Subfossil Cladocera Assemblages: A Case Study in an Oxbow Lake. Water 2022, 14, 2149. [Google Scholar] [CrossRef]
- Korponai, J.L.; Kövér, C.; López-Blanco, C.; Gyulai, I.; Forró, L.; Katalinic, A.; Ketola, M.; Nevalainen, L.; Lioto, T.P.; Sarmaja-Korjonen, K.; et al. Effect of temperature on the size of sedimentary remains of littoral chydorids. Water 2020, 12, 1309. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Defaye, D. Global diversity of copepods (Crustacea: Copepoda) in freshwater. Hydrobiologia 2008, 595, 195–207. [Google Scholar] [CrossRef]
- Perbiche-Neves, G.; Pomari, J.; Serafim-Júnior, M.; Nogueira, M.G. Cyclopoid copepods as indicators of trophic level in South American reservoirs: A new perspective at species level based on a wide spatial-temporal scale. Ecol. Indic. 2021, 127, 107744. [Google Scholar] [CrossRef]
- Gannon, J.E.; Stemberger, R.S. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. Am. Microsc. Soc. 1978, 97, 16–35. [Google Scholar] [CrossRef]
- Pinto-Coelho, R.; Pinel-Alloul, B.; Méthot, G.; Havens, K.E. Crustacean zooplankton in lakes and reservoirs of temperate and tropical regions: Variation with trophic status. Can. J. Aquat. Sci. 2005, 62, 348–361. [Google Scholar] [CrossRef]
- Sendacz, S.; Caleffi, S.; Santos-Soares, J. Zooplankton biomass of reservoirs in different trophic conditions in the State of São Paulo, Brazil. Braz. J. Biol. 2006, 66, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Van Egeren, S.J.; Dodson, S.I.; Torke, B.; Maxted, J.T. The relative significance of environmental and anthropogenic factors affecting zooplankton community structure in Southeast Wisconsin Till Plain lakes. Hydrobiologia 2011, 668, 137–146. [Google Scholar] [CrossRef]
- Kane, D.D.; Gordon, S.I.; Munawar, M.; Charlton, M.N.; Culver, D.A. The Planktonic Index of Biotic Integrity (P-IBI): An approach for assessing lake ecosystem health. Ecol. Indic. 2009, 9, 1234–1247. [Google Scholar] [CrossRef]
- Stamou, G.; Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E. Grazing potential—A functional plankton food web metric for ecological water quality assessment in Mediterranean lakes. Water 2019, 11, 1274. [Google Scholar] [CrossRef]
- Stamou, G.; Mazaris, A.D.; Moustaka-Gouni, M.; Špoljar, M.; Ternjej, I.; Dražina, T.; Dorak, Z.; Michaloudi, E. Introducing a zooplanktonic index for assessing water quality of natural lakes in the Mediterranean region. Ecol. Inform. 2022, 69, 101616. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Karabin, A. The suitability of zooplankton as lake ecosystem indicators: Crustacean trophic state index. Pol. J. Ecol. 2013, 61, 561–573. [Google Scholar]
- Dumont, H.J.; Van de Velde, I.; Dumont, S. The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R. Cascading trophic interactions and lake productivity. BioScience 1985, 35, 634–639. [Google Scholar] [CrossRef]
- Wickham, S.A. The direct and indirect impact of Daphnia and Cyclops on a freshwater microbial food web. J. Plankton Res. 1998, 20, 739–755. [Google Scholar] [CrossRef]
- Perga, M.E.; Kainz, M.; Matthews, B.; Mazumder, A. Carbon pathways to zooplankton: Insights from the combined use of stable isotope and fatty acid biomarkers. Freshw. Biol. 2006, 51, 2041–2051. [Google Scholar] [CrossRef]
- Khan, Q.; Khan, M. Effect of temperature on waterflea Daphnia magna (Crustacea: Cladocera). Nat. Preced. 2008, 1. [Google Scholar] [CrossRef]
- Korponai, J.; Magyari, E.K.; Buczkó, K.; Iepure, S.; Namiotko, T.; Czakó, D.; Kövér, C.; Braun, M. Cladocera response to Late Glacial to Early Holocene climate change in a South Carpathian mountain lake. Hydrobiologia 2011, 676, 223–235. [Google Scholar] [CrossRef]
- Ekvall, M.K.; Urrutia-Cordero, P.; Hansson, L.A. Linking cascading effects of fish predation and zooplankton grazing to reduced cyanobacterial biomass and toxin levels following biomanipulation. PLoS ONE 2014, 9, e112956. [Google Scholar] [CrossRef] [PubMed]
- Berta, C.; Tóthmérész, B.; Wojewódka, M.; Augustyniuk, O.; Korponai, J.; Bertalan-Balázs, B.; Nagy, A.S.; Grigorszky, I.; Gyulai, I. Community response of Cladocera to trophic stress by biomanipulation in a shallow oxbow lake. Water 2019, 11, 929. [Google Scholar] [CrossRef]
- Duggan, I.C.; Green, J.D.; Shiel, R.J. Distribution of rotifer assemblages in North Island, New Zealand, lakes: Relationships to environmental and historical factors. Freshw. Biol. 2002, 47, 195–206. [Google Scholar] [CrossRef]
- Jeppesen, E.; Peder Jensen, J.; SØndergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Tessier, A.J.; Horwitz, R.J. Influence of water chemistry on size structure of zooplankton assemblages. Can. J. Aquat. Sci. 1990, 47, 1937–1943. [Google Scholar] [CrossRef]
- Stemberger, R.S.; Lazorchak, J.M. Zooplankton assemblage responses to disturbance gradients. Can. J. Aquat. Sci. 1994, 51, 2435–2447. [Google Scholar] [CrossRef]
- Beaver, J.R.; Tausz, C.E.; Renicker, T.R.; Holdren, G.C.; Hosler, D.M.; Manis, E.E.; Scotese, K.C.; Teacher, C.E.; Vitanye, B.T.; Davidson, R.M. The late summer crustacean zooplankton in western U.S.A reservoirs reflects ecoregion, temperature and latitude. Freshw. Biol. 2014, 59, 1173–1186. [Google Scholar] [CrossRef]
- Du, X.; García-Berthou, E.; Wang, Q.; Liu, J.; Zhang, T.; Li, Z. Analyzing the importance of top-down and bottom-up controls in food webs of Chinese lakes through structural equation modeling. Aquat. Ecol. 2015, 49, 199–210. [Google Scholar] [CrossRef]
- Mäemets, A. Izmenenija zooplanktona. In Antropogennoe Vozdeistvije na Malye Ozera [Changes of Zooplankton.–Anthropogenic Impact to Small Lakes]; Nauka: Leningrad, Russia, 1980; pp. 54–64. [Google Scholar]
- Barbour, M.T.; Stribling, J.B.; Karr, J.R. Multimetric approach for establishing biocriteria. Biological assessment and criteria. In Tools for Water Resource Planning and Decision Making; CRC Press: Boca Raton, FL, USA, 1995; pp. 63–77. [Google Scholar]
- Hering, D.; Feld, C.K.; Moog, O.; Ofenböck, T. Cook book for the development of a Multimetric Index for biological condition of aquatic ecosystems: Experiences from the European AQEM and STAR projects and related initiatives. In The Ecological Status of European Rivers: Evaluation and Intercalibration of Assessment Methods; Springer: Dordrecht, Switzerland, 2006; pp. 311–324. [Google Scholar] [CrossRef]
- Chen, K.; Hughes, R.M.; Brito, J.G.; Leal, C.G.; Leitao, R.P.; de Oliveira-Junior, J.M.B.; de Oliveira, V.C.; Dias-Silva, K.; Ferraz, S.F.B.; Ferreira, J.; et al. A multi-assemblage, multi-metric biological condition index for eastern Amazonia streams. Ecol. Indic. 2017, 78, 48–61. [Google Scholar] [CrossRef]
- Kairesalo, T.; Tátrai, I.; Luokkanen, E. Impacts of waterweed (Elodea canadensis Michx) on fish-plankton interactions in the lake littoral. Int. Ver. Für Theor. Und Angew. Limnol. Verhandlungen 1998, 26, 1846–1851. [Google Scholar] [CrossRef]
- Mack, H.R.; Conroy, J.D.; Blocksom, K.A.; Stein, R.A.; Ludsin, S.A. A comparative analysis of zooplankton field collection and sample enumeration methods. Limnol. Oceanogr. Methods 2012, 10, 41–53. [Google Scholar] [CrossRef]
- Hillbricht-Ilkowska, A. Morphological variation of Keratella cochlearis (Gosse)(Rotatoria) in several Masurian lakes of different trophic level. Pol. Arch. Hydrobiol. 1972, 19, 253–264. [Google Scholar]
- Pejler, B.I.R.G.E.R. Taxonomic notes on some planktic Rotifers. Zool. Bidr. Från Upps. 1962, 35, 307–319. [Google Scholar]
- Andronikova, I.N. Zooplankton characteristics in monitoring of Lake Ladoga. Hydrobiologia 1996, 322, 173–179. [Google Scholar] [CrossRef]
- Caroni, R.; Irvine, K. The potential of zooplankton communities for ecological assessment of lakes: Redundant concept or political oversight? In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 2010; Volume 110, pp. 35–53. Available online: https://www.muse.jhu.edu/article/809710 (accessed on 9 July 2023).
- Haberman, J.; Haldna, M. Indices of zooplankton community as valuable tools in assessing the trophic state and water quality of eutrophic lakes: Long term study of Lake Vőrtsjärv. J. Limnol. 2014, 73, 263–273. [Google Scholar] [CrossRef]
- Oh, H.J.; Seo, D.H.; Choi, M.; Jeong, H.G.; Kim, H.W.; Oh, J.M.; Chang, K.H. Calculation and applicability of rotifers biomass (Polyarthra spp.) based on length-weight relationship and spatial distribution of body length. KJEE 2018, 51, 205–211. [Google Scholar] [CrossRef]
- Ku, D.; Chae, Y.J.; Choi, Y.; Ji, C.W.; Park, Y.S.; Kwak, I.S.; Kim, Y.J.; Chang, K.H.; Oh, H.J. Optimal Method for Biomass Estimation in a Cladoceran Species, Daphnia Magna (Straus, 1820): Evaluating Length–Weight Regression Equations and Deriving Estimation Equations Using Body Length, Width and Lateral Area. Sustainability 2022, 14, 9216. [Google Scholar] [CrossRef]
- Chen, C.T.; Bănaru, D.; Carlotti, F.; Faucheux, M.; Harmelin-Vivien, M. Seasonal variation in biochemical and energy content of size-fractionated zooplankton in the Bay of Marseille (North-Western Mediterranean Sea). J. Mar. Syst. 2019, 199, 103223. [Google Scholar] [CrossRef]
- Choi, K.H.; Lee, C.R.; Kang, H.K.; Kang, K.A. Characteristics and variation of size-fractionated zooplankton biomass in the Northern East China Sea. Ocean Polar Res. 2011, 33, 135–147. [Google Scholar] [CrossRef]
- Böttger-Schnack, R.; Schnack, D. On the effect of formaldehyde fixation on the dry weight of copepods. In Meeresforschung Reports on Marine Research. Berichte der Deutschen Wissenschaftlichen Kommission für Meeresforschung; Hempel: Kiel, Germany, 1986; Volume 31, pp. 141–152. Available online: https://www.vliz.be/en/imis?refid=142288 (accessed on 9 July 2023).
- Wiebe, P.H. Relationships between zooplankton displacement volume, wet weight, dry weight and carbon. Fish. Bull. 1975, 73, 777–786. [Google Scholar]
- Burns, C.W. Relation between filtering rate, temperature, and body size in four species of Daphnia. Limnol. Oceanogr. 1969, 14, 693–700. [Google Scholar] [CrossRef]
- Ruttner-Kolisko, A. Suggestions for biomass calculations of planktonic rotifers. Arch. Hydrobiol. Beih. 1997, 21, 71–76. [Google Scholar]
- SOP. LG403. Revision 07; Standard Operating Procedure for Zooplankton Analysis. Great Lakes National Program Office, U.S. Environmental Protection Agency: Chicago, IL, USA, 2016.
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. A review of some problems in zooplankton production studies. Norw. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Rosen, R.A. Seasonal Cycles, Distribution, and Biomass of Crustacean Zooplankton, and Feeding and Growth of Young American Shad (Alosa sapidissima) in the Holyoke Pool, Connecticut River; University of Massachusetts Amherst: Amherst, MA, USA, 1981. [Google Scholar]
- Pace, M.L.; Orcutt, J.D., Jr. The relative importance of protozoans, rotifers, and crustaceans in a freshwater zooplankton community 1. Limnol. Oceanogr. 1981, 26, 822–830. [Google Scholar] [CrossRef]
- Hawkins, B.E.; Evans, M.S. Seasonal cycles of zooplankton biomass in Southeastern Lake Michigan. J. Great Lakes Res. 1979, 5, 256–263. [Google Scholar] [CrossRef]
- Doohan, M. An energy budget for adult Brachionus plicatilis Muller (Rotatoria). Oecologia 1973, 13, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.R.; Tausz, C.E.; Black, K.M.; Bolam, B.A. Cladoceran body size distributions along temperature and trophic gradients in the conterminous USA. J. Plankton Res. 2020, 42, 613–629. [Google Scholar] [CrossRef]
- Yebra, L.; Kobari, T.; Sastri, A.R.; Gusmão, F.; Hernández-León, S. Advances in biochemical indices of zooplankton production. Adv. Mar. Biol. 2017, 76, 157–240. [Google Scholar]
- Tao, Y.; Yu, J.; Liu, X.; Xue, B.; Wang, S. Factors affecting annual occurrence, bioaccumulation, and biomagnification of polycyclic aromatic hydrocarbons in plankton food webs of subtropical eutrophic lakes. Water Res. 2018, 132, 1–11. [Google Scholar] [CrossRef]
- Kwak, I.S.; Park, Y.S.; Chang, K.H. Application and Utilization of Environmental DNA Technology for Biodiversity in Water Ecosystems. KJEE 2021, 54, 151–155. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, X. eDNA metabarcoding in zooplankton improves the ecological status assessment of aquatic ecosystems. Environ. Int. 2020, 134, 105230. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jo, H.; Chang, M.H.; Woo, S.H.; Cho, Y.; Yoon, J.D. Application of Environmental DNA for Monitoring of Freshwater Fish in Korea. KJEE 2020, 53, 63–72. [Google Scholar] [CrossRef]
- Logan, J.; Kirstin, E. Real-Time PCR: Current Technology and Applications; Caister Academic Press: Poole, UK, 2009; p. vii-284. [Google Scholar]
- Walsh, J.R.; Spear, M.J.; Shannon, T.P.; Krysan, P.J.; Vander Zanden, M.J. Using eDNA, sediment subfossils, and zooplankton nets to detect invasive spiny water flea (Bythotrephes longimanus). Biol. Invasions 2019, 21, 377–389. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, X.; Xu, M.; Mao, Y.; Huang, Y. DNA metabarcoding of zooplankton communities: Species diversity and seasonal variation revealed by 18S rRNA and COI. PeerJ 2021, 9, e11057. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.L.D.; Lin, Y.Y.; Sato, M.; Hsieh, C.H.; Shiah, F.K.; Machida, R.J. Using metatranscriptomics to estimate the diversity and composition of zooplankton communities. Mol. Ecol. Resour. 2022, 22, 638–652. [Google Scholar] [CrossRef]
- Monchamp, M.È.; Walsh, D.A.; Garner, R.E.; Kraemer, S.A.; Beisner, B.E.; Cristescu, M.E.; Gregory-Eaves, I. Comparative analysis of zooplankton diversity in freshwaters: What can we gain from metagenomic analysis? Environ. DNA 2022, 4, 1250–1264. [Google Scholar] [CrossRef]
- Hinlo, R.; Furlan, E.; Suitor, L.; Gleeson, D. Environmental DNA monitoring and management of invasive fish: Comparison of eDNA and fyke netting. Manag. Biol. Invasions 2017, 8, 89. [Google Scholar] [CrossRef]
- Wang, S.; Yan, Z.; Hänfling, B.; Zheng, X.; Wang, P.; Fan, J.; Li, J. Methodology of fish eDNA and its applications in ecology and environment. Sci. Total Environ. 2021, 755, 142622. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, X.; Chen, Y.; Fu, R.; Du, X.; Chen, X.; Zhan, A. Zooplankton biodiversity monitoring in polluted freshwater ecosystems: A technical review. Environ. Sci. Ecotechnol. 2020, 1, 100008. [Google Scholar] [CrossRef]
- Oh, H.J.; Chae, Y.J.; Choi, Y.; Ku, D.; Heo, Y.J.; Kwak, I.S.; Jo, H.; Park, Y.S.; Chang, K.H.; Kim, H.W. Review and suggestions for applying DNA sequencing to zooplankton researches: From taxonomic approaches to biological interaction analysis. KJEE 2021, 54, 156–169. [Google Scholar] [CrossRef]
- Cordier, T.; Lanzén, A.; Apothéloz-Perret-Gentil, L.; Stoeck, T.; Pawlowski, J. Embracing Environmental Genomics and Machine Learning for Routine Biomonitoring. Trends Microbiol. 2019, 27, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Lu, Q.; Jia, C.; Dai, Y.; Ouyang, S.; Wu, X. The Effects of Water Level Fluctuation on Zooplankton Communities in Shahu Lake Based on DNA Metabarcoding and Morphological Methods. Animals 2022, 12, 950. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.W.; Oh, H.J.; Chang, K.H.; Park, Y.S.; Kwak, I.S. A Comparative Analyzing of Zooplankton Community Diversity in Surface Layer Water of Reservoir Via eDNA Metabarcoding and Microscopy. Divers 2022, 14, 797. [Google Scholar] [CrossRef]
- Harvey, J.B.; Johnson, S.B.; Fisher, J.L.; Peterson, W.T.; Vrijenhoek, R.C. Comparison of morphological and next generation DNA sequencing methods for assessing zooplankton assemblages. J. Exp. Mar. Biol. Ecol. 2017, 487, 113–126. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.; Oh, H.-J.; Lee, D.-H.; Jang, M.-H.; Lee, K.-L.; Chang, K.-H.; Kim, H.-W. Current Utilization and Further Application of Zooplankton Indices for Ecosystem Health Assessment of Lake Ecosystems. Sustainability 2023, 15, 10950. https://doi.org/10.3390/su151410950
Choi Y, Oh H-J, Lee D-H, Jang M-H, Lee K-L, Chang K-H, Kim H-W. Current Utilization and Further Application of Zooplankton Indices for Ecosystem Health Assessment of Lake Ecosystems. Sustainability. 2023; 15(14):10950. https://doi.org/10.3390/su151410950
Chicago/Turabian StyleChoi, Yerim, Hye-Ji Oh, Dae-Hee Lee, Min-Ho Jang, Kyung-Lak Lee, Kwang-Hyeon Chang, and Hyun-Woo Kim. 2023. "Current Utilization and Further Application of Zooplankton Indices for Ecosystem Health Assessment of Lake Ecosystems" Sustainability 15, no. 14: 10950. https://doi.org/10.3390/su151410950
APA StyleChoi, Y., Oh, H.-J., Lee, D.-H., Jang, M.-H., Lee, K.-L., Chang, K.-H., & Kim, H.-W. (2023). Current Utilization and Further Application of Zooplankton Indices for Ecosystem Health Assessment of Lake Ecosystems. Sustainability, 15(14), 10950. https://doi.org/10.3390/su151410950