
The Large-Scale Effect of Forest Cover on Long-Term Streamflow Variations in Mediterranean Catchments of Central Chile


 ,
,  ,
,  ,
,
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Rainy Season | Hydrologic Effect | Source |
|---|---|---|---|
| Oak, pine | All year round | Streamflow reduction | [13] |
| Sequoia serpervirens | Winter | Streamflow reduction | [34] |
| Eucalyptus sp. | All year round | Streamflow reduction | [45] |
| Eucalyptus grandis | Mostly summer | Streamflow reduction | [46] |
| Eucalyptus globulus | Mostly summer | Streamflow reduction | [47] |
| Pinus patula | All year round | Streamflow reduction | [48] |
| Eucalyptus globulus | All year round | Streamflow reduction | [49] |
| Pinus radiata | All year round | Streamflow reduction | [50] |
| Pseudotsuga menziesii | All year round | Streamflow reduction | [51] |
| Mixed conifers | All year round | Streamflow reduction | [52] |
| Picea sitchensis | All year round | Streamflow reduction | [53] |
| Cryptomeria japonica | All year round | Streamflow reduction | [54] |
| Eucalyptus grandis | Mostly summer | Streamflow reduction | [55] |
| Picea, Abies, Larix ssp. | Summer | Streamflow reduction | [56] |
| Pinus caribaea | Summer | Streamflow reduction | [57] |
| Pinus, Fagus, Quercus ssp. | Mostly spring | Streamflow reduction | [58] |
2. Materials and Methods
2.1. Mediterranean-Type Ecosystems of Central Chile and Catchments under Study
2.2. Land Use of the Mediterranean-Climate (MC) Catchments
2.3. Classification of Catchments According to Their Forest Cover
2.4. Calculation of Relative Changes in Forest Cover
2.5. Long-Term Streamflow Variations versus Forest Cover Dynamics
2.6. Intra-Catchment Relationships and Field Significance
2.7. Long-Term Variations of Other Hydrological Fluxes
3. Results
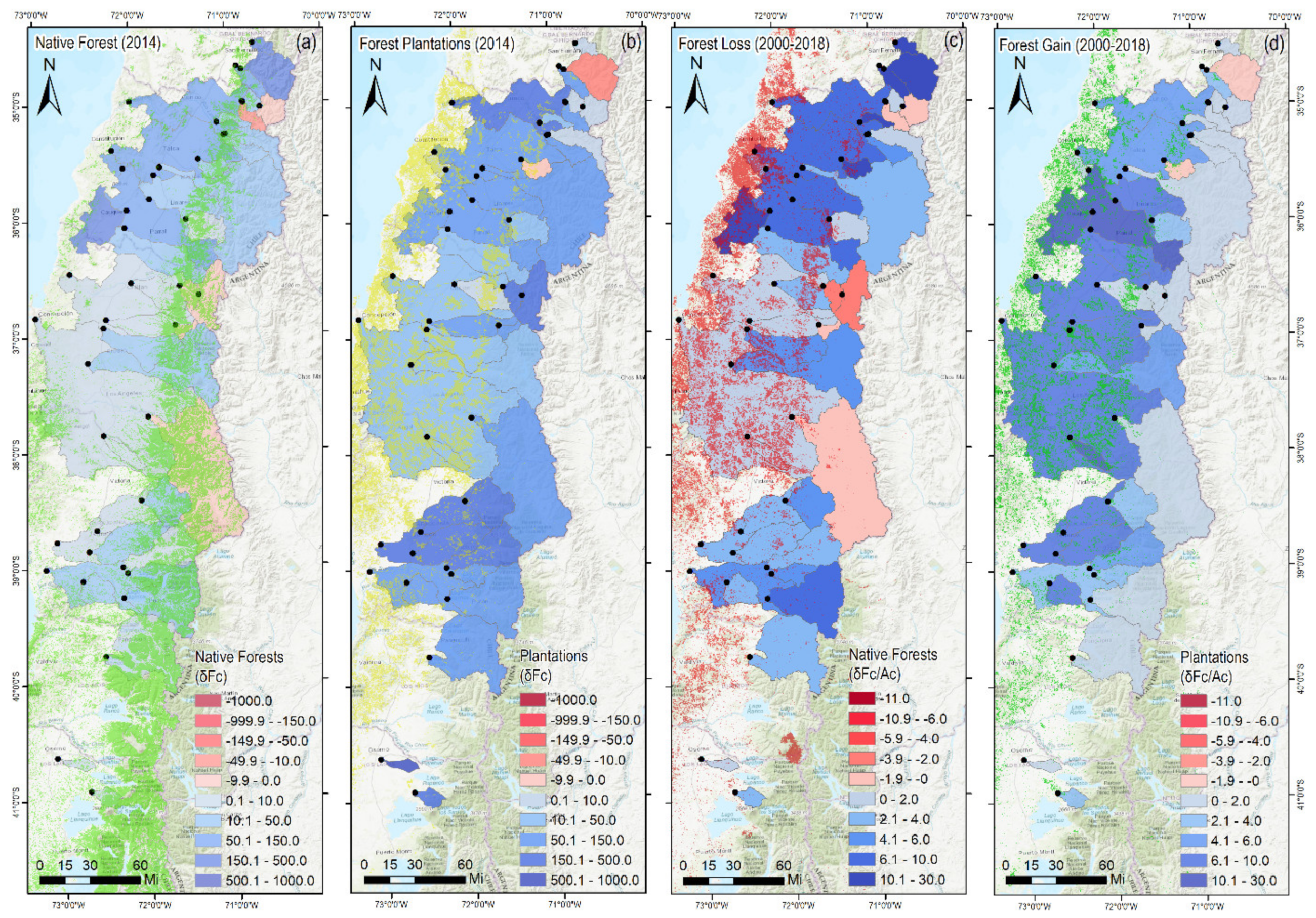
3.1. Mediterranean-Climate (MC) Ecosystems and Long-Term Forest Cover Changes
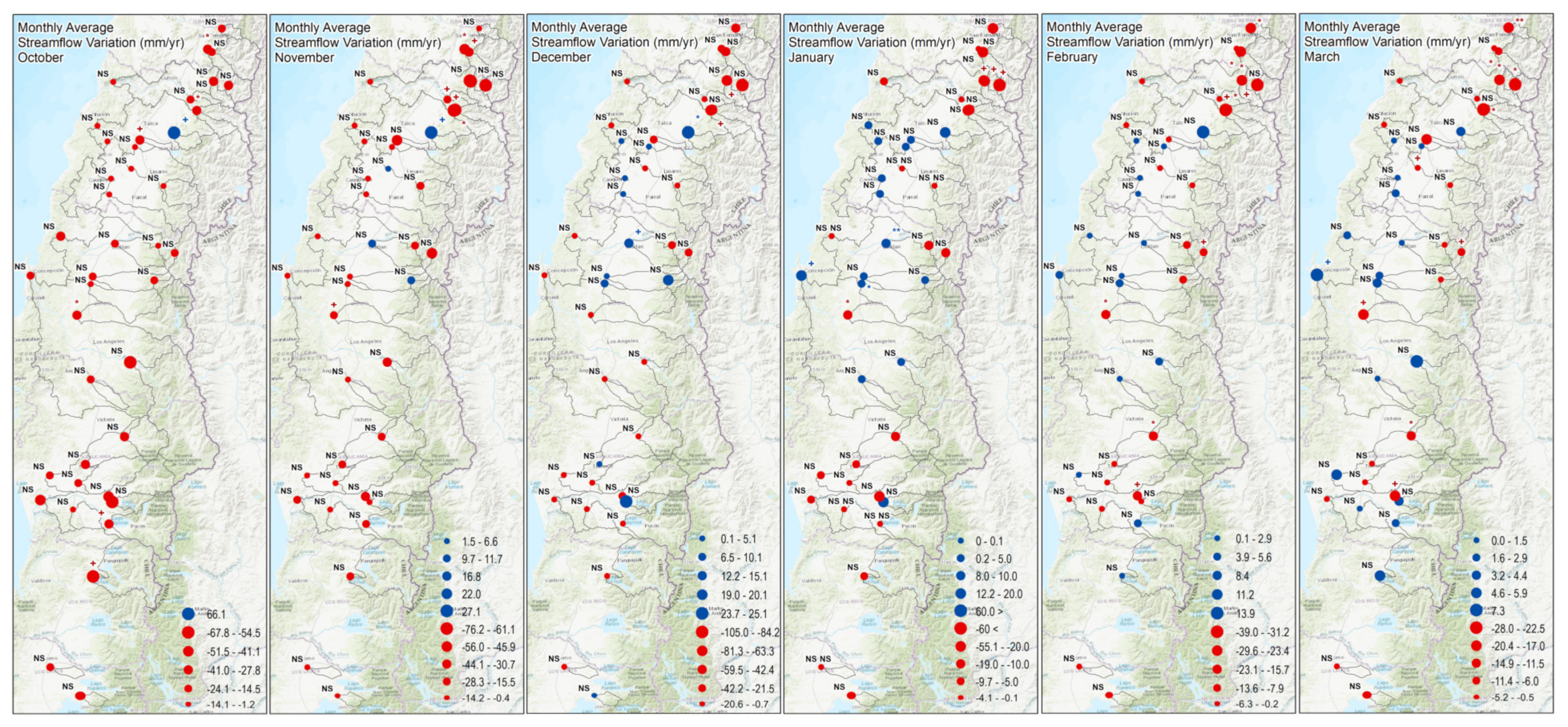
3.2. Long-Term Streamflow Variations within Mediterranean-Climate Catchments
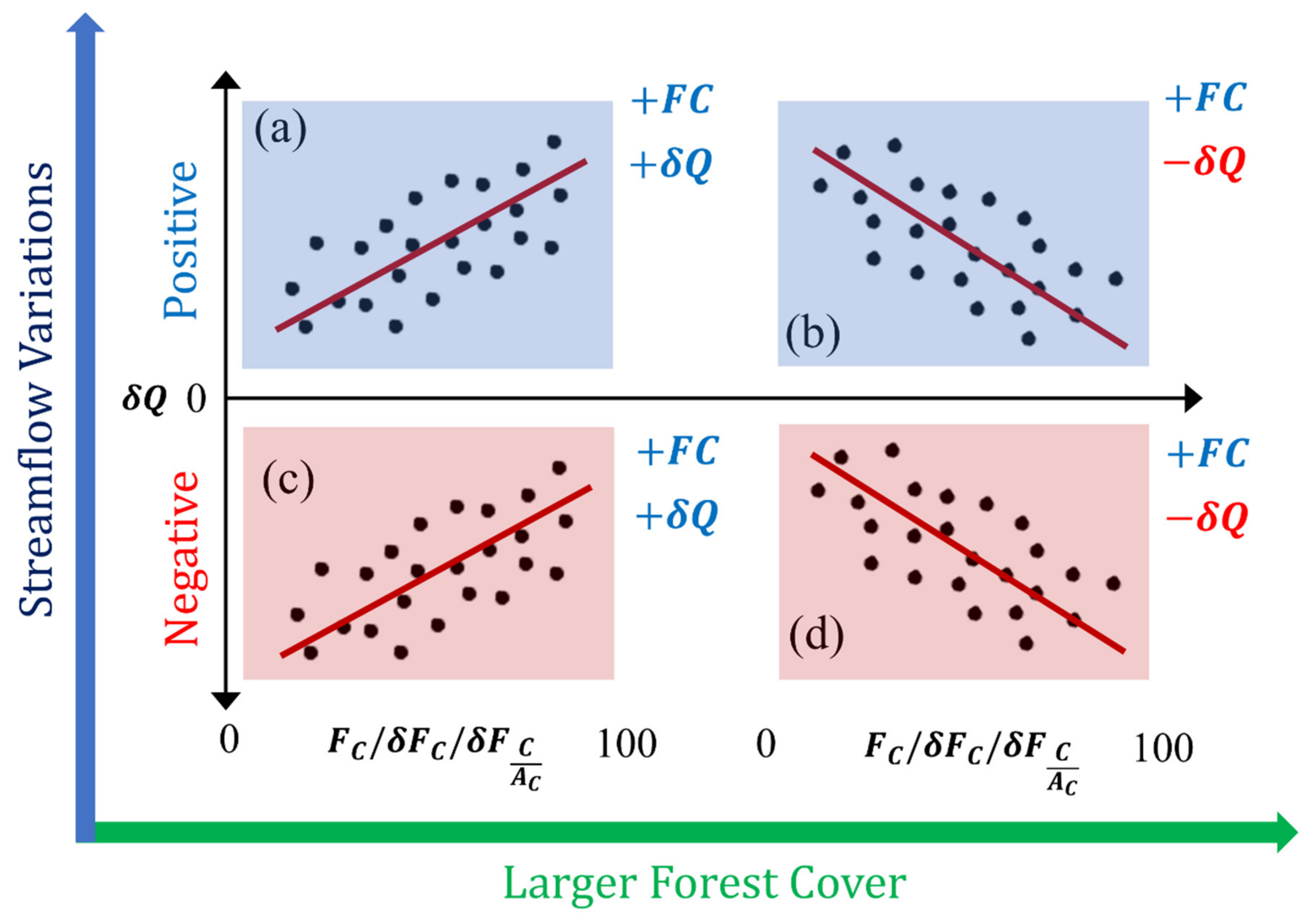
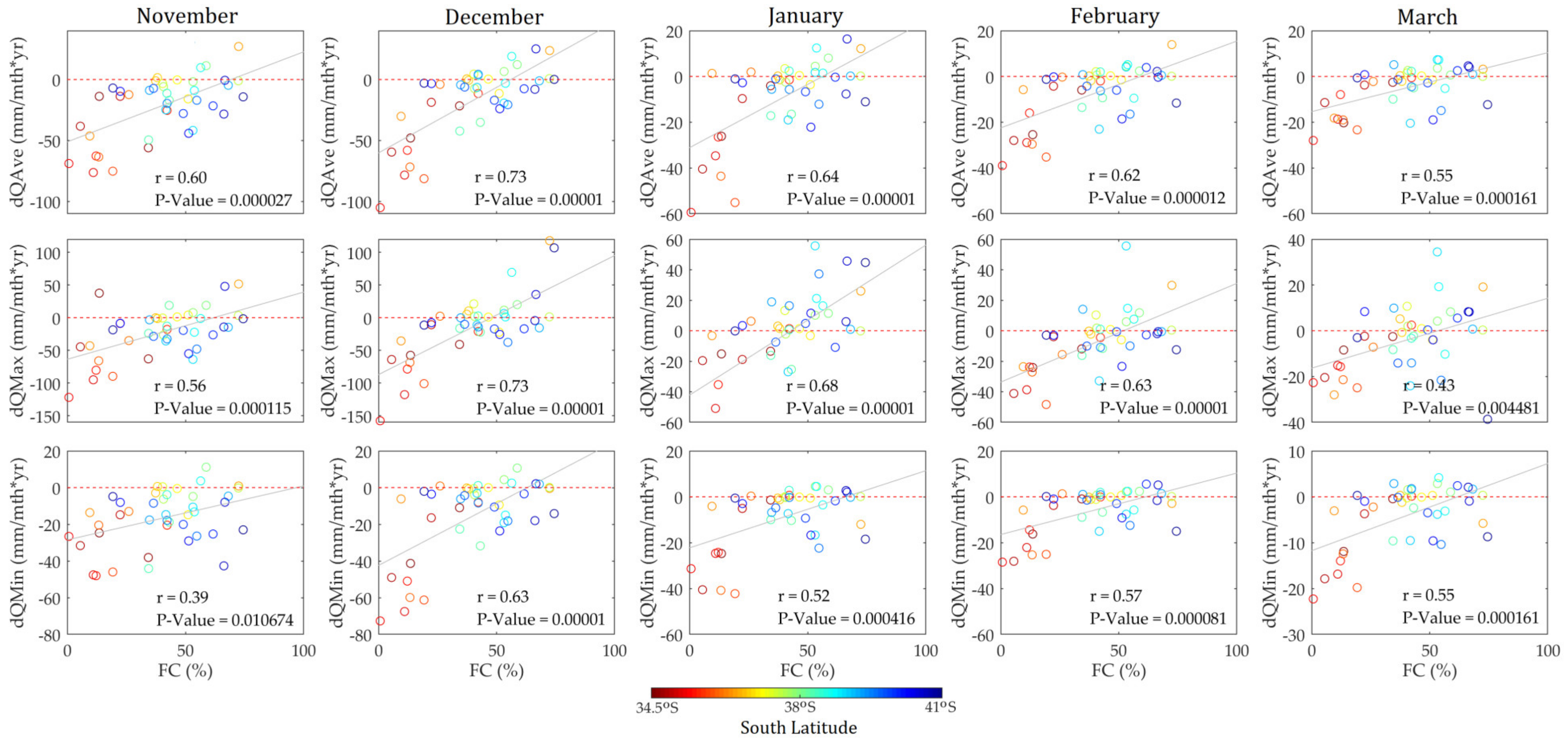
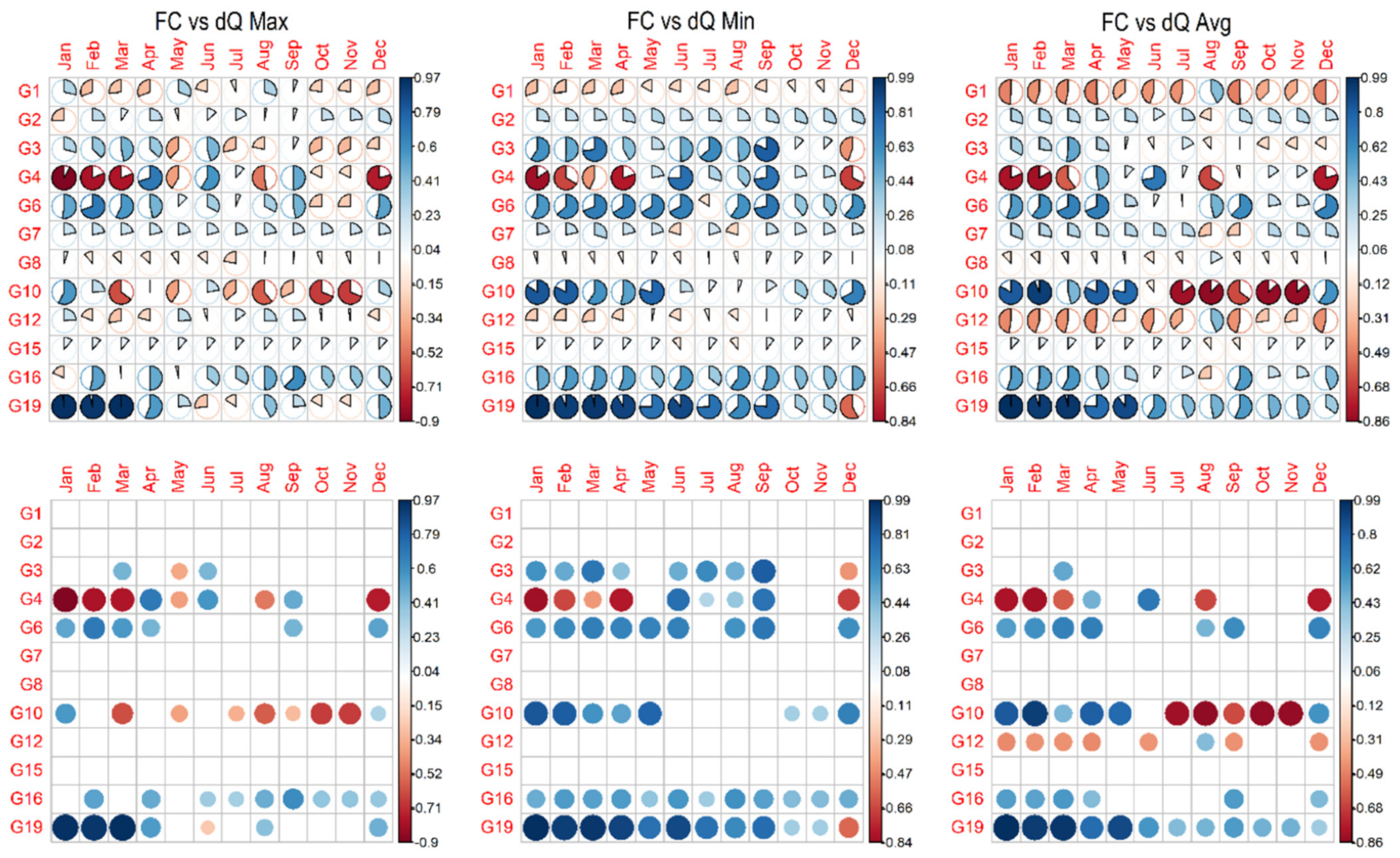
3.3. Forest Cover Dynamics versus Long-Term Streamflow Variations
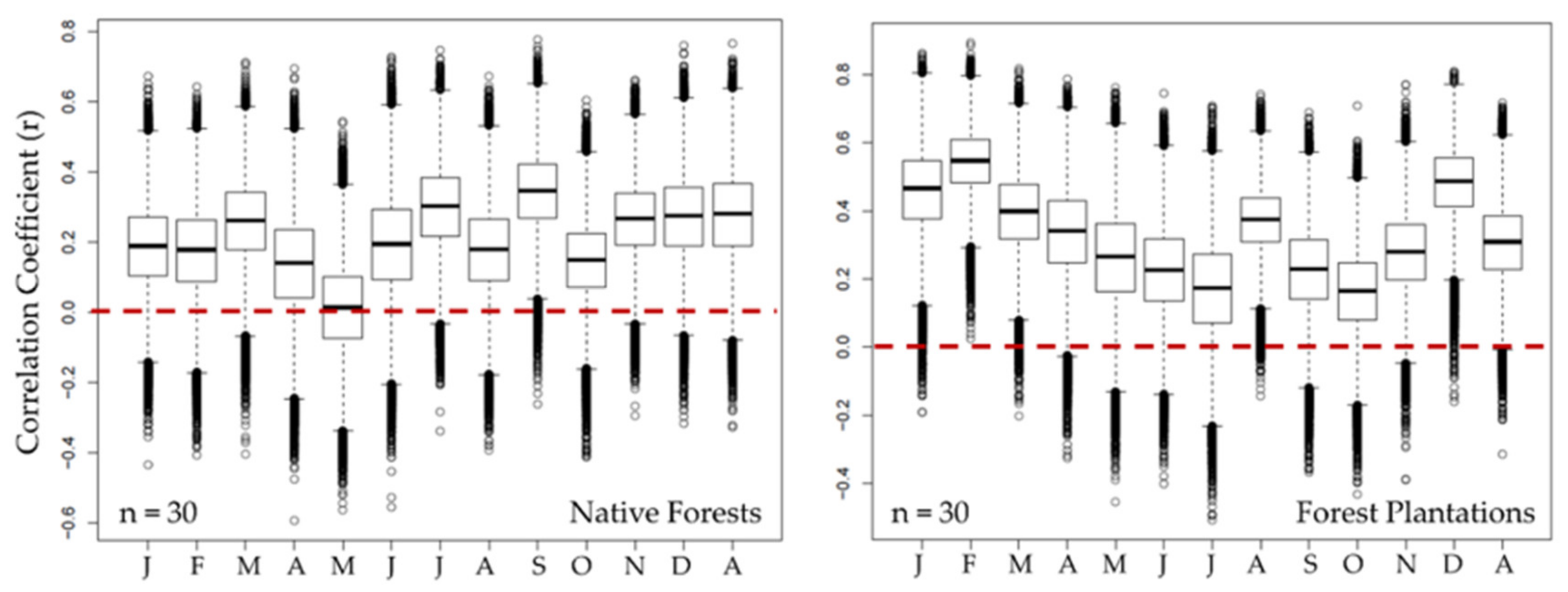
3.4. Disaggregation between Groups of Native Forests (NF) and Forest Plantations (FP)
3.5. Field significance: Bootstrapping Analysis
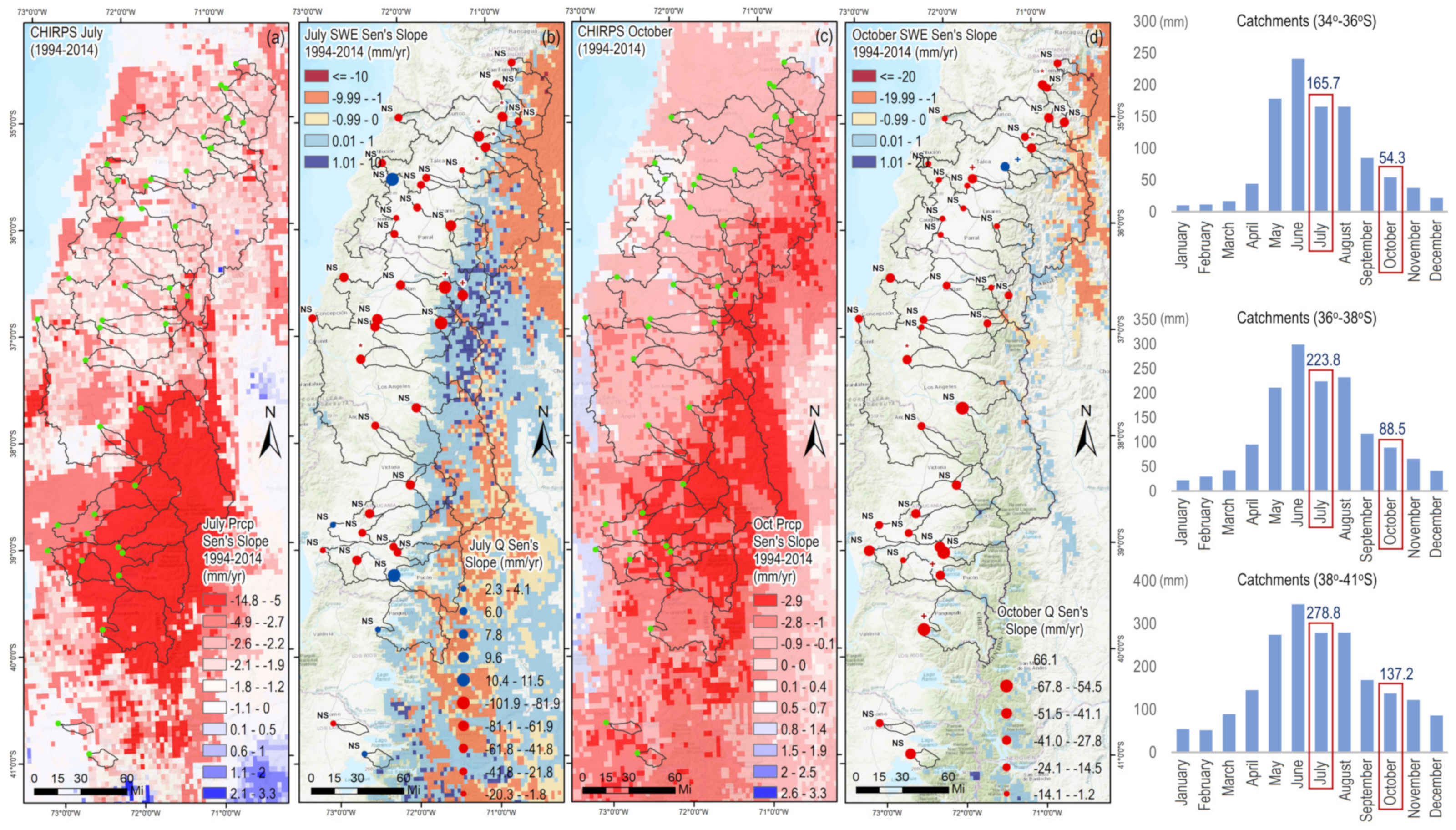
3.6. Influence of Precipitation and Snow Water Equivalent on Total Streamflow
4. Discussion
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, Q.; Dong, S.; Duan, B. The Effects of Land-Use Change/Conversion on Trade-Offs of Ecosystem Services in Three Precipitation Zones. Sustainability 2021, 13, 13306. [Google Scholar] [CrossRef]
- Brouillard, B.M.; Dickenson, E.; Mikkelson, K.M.; Sharp, J.O. Water quality following extensive beetle-induced tree mortality: Interplay of aromatic carbon loading, disinfection byproducts, and hydrologic drivers. Sci. Total Environ. 2016, 572, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Mikkelson, K.; Bearup, L.; Navarre-Sitchler, A.; McCray, J.; Sharp, J. Changes in metal mobility associated with bark beetle-induced tree mortality. Environ. Sci. Processes Impacts 2014, 16, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Mikkelson, K.; Bearup, L.; Maxwell, R.; Stednick, J.; McCray, J.; Sharp, J. Bark beetle infestation impacts on nutrient cycling, water quality and interdependent hydrological effects. Biogeochemistry 2013, 115, 1–21. [Google Scholar] [CrossRef]
- Mikkelson, K.; Dickenson, E.; McCray, J.; Maxwell, R.; Sharp, J. Adverse water quality impacts from climate-induced forest die-off. Nat. Clim. Change 2013, 3, 218–222. [Google Scholar] [CrossRef]
- Pizarro, R.; Garcia-Chevesich, P.; Pino, J.; Ibáñez, A.; Pérez, F.; Flores, J.; Sharp, J.; Ingram, B.; Mendoza, R.; Neary, D.; et al. Stabilization of stage-discharge curves following the establishment of forest plantations: Implications for sediment production. River Res. Appl. 2020, 36, 1828–1837. [Google Scholar] [CrossRef]
- Reyna, T.; Garcia-Chevesich, P.; Neary, D.G.; Scott, D.F.; Benyon, R.G.; Reyna, S.M.; Lábaque, M.; Amani, C.; Pizarro, R.; Iroumé, A.; et al. Forest Management and the Impact on Water Resources: A Review of 13 Countries; United Nations Educational, Scientific and Cultural Organization (UNESCO): Paris, France, 2017; p. 204. [Google Scholar]
- Bosch, J.; Hewlett, J. A review of catchment experiments to determine the effect of vegetation change on water yield. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Andréassian, R. Water and forest: From historical controversy to scientific debate. J. Hydrol. 2004, 291, 1–27. [Google Scholar] [CrossRef]
- McCulloch, J.; Robinson, M. History of forest hydrology. J. Hydrol. 1993, 150, 189–216. [Google Scholar] [CrossRef]
- Zhang, L.; Dawes, W.; Walker, G. Response of mean annual evapotranspiration to vegetation changes at catchment scale. Water Resour. Res. 2001, 37, 701–708. [Google Scholar] [CrossRef]
- Xiao, Q.; Mcpherson, E.; Ustin, S. A new approach to modeling tree rainfall interception. J. Geophys. Res. 2000, 105, 29173–29188. [Google Scholar] [CrossRef]
- Calder, I.; Reid, I.; Nisbet, T.; Green, J. Impact of lowland forests in England on water resources: Application of the Hydrological Land Use Change (HYLUC) model. Water Resour. Res. 2003, 39, 1319. [Google Scholar] [CrossRef]
- Van Dijk, A.I.; Keenan, R.J. Planted forests and water in perspective. For. Ecol. Manag. 2007, 251, 1–9. [Google Scholar] [CrossRef]
- Benyon, R.; Theiveyanathan, S.; Doody, T. Impacts of tree plantations on groundwater in south-eastern Australia. Aust. J. Bot. 2006, 397, 181–192. [Google Scholar] [CrossRef]
- Keenan, R.; Gerrand, A.; Nambiar, S.; Parsons, M. Plantation and Water: Plantation Impacts on Streamflow; Science for Decision Makers, Bureau of Rural Sciences: Canberra, ACT, Australia, 2017.
- Beets, P.; Oliver, G. Water use by managed stands of Pinus radiata, indigenous podocarp/hardwood forest, and improved pasture in the central north island of New Zealand. New Zealand J. For. Sci. 2006, 37, 306–323. [Google Scholar]
- Vertessy, R.; Watson, F.; O’Sullivan, S. Factor determining relations between stand age and catchment water balance in mountains ash forest. For. Ecol. Manag. 2001, 143, 13–26. [Google Scholar] [CrossRef]
- Brown, A.; Zhang, L.; McMahon, T.; Western, W.; Vertessy, R. A review of paired catchment studies for determining changes in water yield resulting from alterations in vegetation. J. Hydrol. 2005, 310, 28–61. [Google Scholar] [CrossRef]
- Cornish, P.; Vertessy, R. Forest age-induced changes in evapotranspiration and water yield in a eucalypt forest. J. Hydol. 2001, 242, 43–63. [Google Scholar] [CrossRef]
- Maier, C.; Albaugh, T.; Cook, R.; Hall, K.; McInnis, D.; Johnsen, K.; Johnson, J.; Rubilar, R.; Vose, J. Comparative water use in short-rotation Eucalyptus benthamii and Pinus taeda trees in the Southern United States. For. Ecol. Manag. 2017, 397, 126–138. [Google Scholar] [CrossRef]
- Whitehead, D.; Kelliher, F. Modeling the water balance of a samll Pinus radiata catchment. Tree Physiol. 1991, 9, 17–33. [Google Scholar] [CrossRef]
- Oyarzún, C.; Huber, A. Balance hídrico en plantaciones jóvenes de Eucalyptus globulus y Pinus radaiata en el sur de Chile. Terra 1999, 17, 35–44. [Google Scholar]
- Huber, A.; Iroumé, A.; Mohr, C.; Frêne, C. Efecto de las plantaciones de Pinus radiata y Eucalyptus globulus sobre el recurso agua en la cordillera de la costa región del Biobío, Chile. Bosque 2010, 31, 219–230. [Google Scholar] [CrossRef]
- Farley, K.; Jobbágy, E.; Jackson, R. Effects of afforestation on water yield: A global synthesis with implications for policy. Glob. Change Biol. 2005, 11, 1565–1576. [Google Scholar] [CrossRef]
- Scott, D. On the hydrology of industrial timber plantations. Hydrol. Processes 2005, 19, 4203–4206. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). Effects of Forest Practices on Peak Flows and Consequent Channel: A State-of- Science Report for Western Oregon and Washington; General Technical Report: PNW-GTR-760; United States Department of Agriculture (USDA): Washington, DC, USA, 2010; 76p.
- Lane, P.; Best, A.; Hickel, K.; Zhang, L. The response of flow duration curves to afforestation. J. Hydrol. 2005, 310, 253–265. [Google Scholar] [CrossRef]
- Best, A.; Zhang, L.; McMahon, T.; Western, A.; Vertessy, R. A Critical Review of Paired Catchment Studies with Reference to Seasonal Flows and Climatic Variability; CSIRO land and water technical report 25/03; CSIRO: Canberra, ACT, Australia, 2003. Available online: https://publications.csiro.au/rpr/download?pid=procite:fcdfd12f-dca1-41c0-85ce-d06d0c542e7c&dsid=DS1 (accessed on 3 March 2021).
- Cannel, M. Environmental impacts of forest monocultures: Water use, acidification, wildlife conservation, and carbon storage. New For. 1999, 17, 239–262. [Google Scholar] [CrossRef]
- Schwärzel, K.; Zhang, L.; Montanarella, L.; Wang, Y.; Sun, G. How afforestation affects the water cycle in drylands: A process-based comparative analysis. Glob. Change Biol. 2019, 26, 944–959. [Google Scholar] [CrossRef]
- Troch, P.; Lahmers, T.; Meira, A.; Mukherjee, R.; Pedersen, J.; Roy, T.; Valdés-Pineda, R. Catchment coevolution: A useful framework for improving predictions of hydrological change? Water Resour. Res. 2015, 51, 4903–4922. [Google Scholar] [CrossRef]
- Valdés-Pineda, R.; Pizarro, R.; Garcia-Chevesich, P.; Valdés, J.; Pérez, F.; Olivares, C.; Vera, M.; Balocchi, F.; Vallejos, C.; Fuentes, R.; et al. Water Governance in Chile: Availability, Management, and Climate Change. J. Hydrol. 2014, 519, 2538–2567. [Google Scholar] [CrossRef]
- Keppeler, E.; Ziemer, R. Logging effects on streamflow: Water yield and summer low flows at caspar creek in northwestern California. Water Resour. Res. 1990, 26, 1669–1679. [Google Scholar] [CrossRef]
- Iroumé, A.; Huber, A.; Schulz, K. Summer flows in experimental catchments with different forest covers, Chile. J. Hydrol. 2005, 300, 300–313. [Google Scholar] [CrossRef]
- Iroumé, A.; Mayen, O.; Huber, A. Runoff and peak response to timber harvest and forest age in southern Chile. Hydrol. Processes 2006, 20, 37–50. [Google Scholar] [CrossRef]
- Iroumé, A.; Palacios, H.; Bathurst, J.; Huber, A. Escorrentías y caudales máximos luego de la cosecha a tala rasa y del establecimiento de una nueva plantación en una cuenca experimental del sur de Chile. Bosque 2010, 31, 117–128. [Google Scholar] [CrossRef]
- Little, C.; Lara, A.; McPhee, J.; Urrutia, R. Revealing the impacts of forest exotic plantations on water yield in large scale watersheds in south-central Chile. J. Hydrol. 2009, 374, 162–170. [Google Scholar] [CrossRef]
- Pizarro, R.; Araya, S.; Jordán, C.; Farías, C.; Flores, J.; Bro, P. The effects of changes in vegetative cover on river flows in the Purapel river basin of central Chile. J. Hydrol. 2006, 327, 249–257. [Google Scholar] [CrossRef]
- Lara, A.; Little, C.; Urrutia, R.; McPhee, J.; Álvarez-Garretón, C.; Orazún, C.; Soto, D.; Donoso, P.; Nahuelhual, L.; Pino, M.; et al. Assessment of ecosystem services as an opportunity for the conservation and management of native forests in Chile. For. Ecol. Manag. 2009, 258, 415–424. [Google Scholar] [CrossRef]
- Birkinshaw, S.; Bathurst, J.; Iroumé, A.; Palacios, H. The effect of forest cover on peak flow and sediment discharge—An integrated field and modelling study in central–southern Chile. Hydrol. Processes 2011, 25, 1284–1297. [Google Scholar] [CrossRef]
- Iroumé, A.; Palacios, H. Afforestation and changes in forest composition affect runoff inlarge river basins with pluvial regime and Mediterranean climate, Chile. J. Hydrol. 2013, 15, 113–125. [Google Scholar] [CrossRef]
- Soto-Schönherr, S.; Iroumé, A. How much water do Chilean forests use? A review of interception losses in forest plot studies. Hydrol. Processes 2016, 30, 4674–4686. [Google Scholar] [CrossRef]
- Alvarez-Garreton, C.; Lara, A.; Boisier, J.; Galleguillos, M. The Impacts of Native Forests and Forest Plantation on Water Supply in Chile. Forests 2019, 10, 473. [Google Scholar] [CrossRef]
- Benyon, R.; Doody, T. Water Use by Tree Plantations in South East South Australia; Technical Report N°148; CSIRO Forestry and Forest Products: Mt. Gambier, SA, USA, 2004. Available online: https://publications.csiro.au/rpr/pub?pid=procite:b06215c8-60bb-4118-9045-fc2bd027d70f (accessed on 10 July 2021).
- Bosch, J.; Smith, J. A review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration. J. Hydrol. 1989, 55, 3–23. [Google Scholar] [CrossRef]
- Sikka, A.; Samra, J.; Sharda, V.; Samraj, P.; Lakshmanan, V. Low flow and high flow responses to converting natural grassland into bluegum (Eucalyptus globulus) in Nilgiris watersheds of South India. J. Hydrol. 2003, 270, 12–26. [Google Scholar] [CrossRef]
- Buytaert, W.; Iñiguez, V.; Bièvre, B. The effects of afforestation and cultivation on water yield in the andean páramo. For. Ecol. Manag. 2007, 251, 22–30. [Google Scholar] [CrossRef]
- Cornish, P. The effects of logging ad forest regeneration on water yields in a moist eucalypt forest in New South Wales, Australia. J. Hydrol. 1993, 150, 301–322. [Google Scholar] [CrossRef]
- Fahey, B.; Jackson, R. Hydrological impacts of converting native forests and grasslands to pine plantations, South Island, New Zealand. Agric. For. Meteorol. 1997, 84, 69–82. [Google Scholar] [CrossRef]
- Harr, R.; Levno, A.; Mersereau, R. Streamflow changes after logging 130-year-old Douglas fir in two small watershed. Water Resour. Res. 1982, 18, 637–644. [Google Scholar] [CrossRef]
- Hicks, B.; Beschta, R.; Harr, D. Long-term changes in streamflow following logging in western Oregon and associated fisheries implications. Water Resour. Bull. 1991, 27, 217–226. [Google Scholar] [CrossRef]
- Hudson, J.; Crane, S.; Blackie, J. The Plynlimon water balance 1969–1995: The impact of forest moorland vegetation on evaporation and streamflow in upland catchment. Hydrol. Earth Syst. Sci. 1997, 1, 409–427. [Google Scholar] [CrossRef]
- Komatsu, H.; Tanaka, N.; Kume, T. Do coniferous forest evaporate more water than broad-leaved forest in Japan? J. Hydrol. 2007, 336, 361–375. [Google Scholar] [CrossRef]
- Scott, D.; Lesch, W. Streamflow responses to afforestation with Eucalyptus grandis and Pinus patula and to felling in the Mokobulaan experimental catchments, South Africa. J. Hydrol. 1997, 199, 360–377. [Google Scholar] [CrossRef]
- Swanson, R.; Golding, D.; Rothwell, R.; Bernier, P. Hydrologic Effects Bof Clear-Cutting at Marmot Creek and Streeter Watershed; Alberta Information Report NOR-X-278; Canadian Forestry Service, Northern Forestry Centre: Edmonton, AB, Canada, 1986. Available online: https://research-groups.usask.ca/hydrology/documents/pubs/marmot/swanson_et_al_1986a.pdf (accessed on 3 March 2021).
- Waterloo, M.; Schellekens, J.; Bruijnzeel, L.; Rawaqa, T. Changes in catchment runoff after harvesting and burning of Pinus caribaea plantation in Viti Levu, Fiji. For. Ecol. Manag. 2007, 251, 31–44. [Google Scholar] [CrossRef]
- Serrano-Muela, P.; Regüés, D.; Lana-Renault, N.; Nadal, E. 2008 Estudio de la trascolación bajo diferentes tipos de cubierta forestal durante el periodo fenológico con hojas en el Pirineo Central Español. In Proceedings of the Trabajos de Geomorfología en España, 2006–2008: X Reunión Nacional de Geomorfología, Cádiz, Spain, 14–19 September 2008. [Google Scholar]
- Santoro, C.; Capriles, J.; Gayo, E.; De Porras, M.; Maldonado, A.; Standen, V.; Latorre, C.; Castro, V.; Angelo, D.; McRostie, V.; et al. Continuities and discontinuities in the socio-environmental systems of the Atacama Desert during the last 13,000 years. J. Anthropol. Archeol. 2017, 46, 28–39. [Google Scholar] [CrossRef]
- Rech, J.; Quade, J.; Betancourt, J. Late quaternary paleohydrology of the central Atacama desert (lat 22°–24°), Chile. Geol. Soc. Am. Bull. 2002, 114, 334–348. [Google Scholar] [CrossRef]
- Carro, L.; Castro, J.; Razmilic, V.; Noiovi, I.; Pan, C.; Igual, J.; Jaspars, M.; Goodfellow, M.; Bull, A.; Asenjo, J.; et al. Uncovering the potential of novel micromonosporae isolated from an extreme hyper-arid Atacama Desert soil. Nat. Sci. Rep. 2019, 9, 4678. [Google Scholar] [CrossRef]
- Cowling, R.; Rundel, P.; Lamont, B.; Arroyo, M.; Arianoutsou, M. Plant diversity in Mediterranean-climate regions. Trend Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Armesto, J.; Arroyo, M.; Hinojosa, O. The mediterranean environment of central Chile. In The Physical Geography of South America; Oxford Scholarship Online: Oxford, UK, 2007; pp. 184–199. ISBN 9780195313413. [Google Scholar]
- Corporación Nacional Forestal de Chole (Conaf). Catastro Vegetacional- Superficies de uso de Suelo Regional. 2017. Available online: https://www.conaf.cl/nuestros-bosques/bosques-en-chile/catastro-vegetacional/ (accessed on 10 May 2021).
- Elizalde Mac-Clure, R. La Sobrevivencia de Chile. La Conservación de Sus Recursos Naturales Renovables; Servicio Agrícola y Ganadero, Ministerio de Agricultura: Santiago, Chile, 1958; 492. [Google Scholar]
- Zamorano-Elgueta, C.; Rey, M.; Cayuela, L.; Hantson, S.; Armenteras, D. Native forest replacement by exotic plantations in southern Chile (1985–2011) and partial compensation by natural regeneration. For. Ecol. Manag. 2015, 345, 10–20. [Google Scholar] [CrossRef]
- Uribe, S.; Estades, C.; Radeloff, V. Pine plantations and five decades of land use change in central Chile. PLoS ONE 2020, 15, e0230193. [Google Scholar] [CrossRef]
- Durán, A.; Barbosa, O. Seeing Chile’s forest for the tree plantations. Science 2019, 365, 1388. [Google Scholar] [CrossRef]
- Heilmayr, R.; Echeverría, C.; Lambin, E. Impacts of Chilean forest subsidies on forest cover, carbon and biodiversity. Nat. Sustain. 2020, 3, 701–709. [Google Scholar] [CrossRef]
- Heymar, R.; Echeverría, C.; Fuentes, R.; Lambin, E. A plantation-dominated forest transition in Chile. Appl. Geogr. 2016, 75, 71–82. [Google Scholar]
- Valdés-Pineda, R.; Pizarro, R.; Valdés, J.; Carrasco, J.; García-Chevesich, P.; Olivares, C. Spatio-temporal trends of precipitation, its aggressiveness and concentration, along the Pacific coast of South America (36–49°S). Hydrol. Sci. J. 2016, 61, 2110–2132. [Google Scholar] [CrossRef]
- Quintana, J.; Aceituno, P. Changes in the rainfall regime along the extratropical west coast of South America (Chile): 30–43° S. Atmósfera 2012, 25, 1–22. [Google Scholar]
- Montecinos, A.; Aceituno, P. Seasonality of the ENSO-Related rainfall variability in Central Chile and associated circulation anomalies. J. Clim. 2003, 16, 281–296. [Google Scholar] [CrossRef]
- Falvey, M.; Garreaud, R. Wintertime precipitation episodes in central Chile: Associated meteorological condition and Orographic influences. J. Hydrometeorol. 2007, 8, 171–193. [Google Scholar] [CrossRef]
- Viale, M.; Garreaud, R. Orographic effects of the subtropical and extratropical Andes on upwind precipitating clouds. J. Geophys. Res. Atmos. 2015, 120, 4962–4974. [Google Scholar] [CrossRef]
- Pizarro, R.; Vera, M.; Valdés, R.; Helwig, B.; Olivares, C. Multi-decadal variations in annual maximum peak flows in semi-arid and temperate regions of Chile. Hydrol. Sci. J. 2014, 59, 300–311. [Google Scholar] [CrossRef][Green Version]
- Shadmani, M.; Marofi, S.; Roknian, M. Trend analysis in reference evapotranspiration using Mann-Kendall and Spearman’s Rho test in arid regions of Iran. Water Resour. Manag. 2012, 26, 211–224. [Google Scholar] [CrossRef]
- Valdés-Pineda, R.; Valdés, J.; Diaz, H.; Pizarro-Tapia, R. Analysis of spatio-temporal changes and seasonal precipitation variability in South America-Chile and related ocean-atmosphere circulation patterns. Int. J. Climatol. 2016, 36, 2979–3001. [Google Scholar] [CrossRef]
- Sangüesa, C.; Pizarro, R.; Ibañez, A.; Pino, J.; Rivera, D.; García-Chevesich, P.; Ingram, B. Spatial and Temporal Analysis of Rainfall Concentration Using the Gini Index and PCI. Water 2018, 10, 112. [Google Scholar] [CrossRef]
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The climate hazards infrared precipitation with stations—A new environmental record for monitoring extremes. Nat. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef]
- Abatzoglou, J.; Dobrowski, S.; Parks, S.; Hegewisch, K. Terraclimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Contesse, D. El Desarrollo Forestal Chileno: Una Realidad Sustentable; SEREPSERVI: Santiago, Chile, 1990; 144p. [Google Scholar]
- Guardiola-Claramonte, M.; Troch, P.; Breshears, D.; Huxman, T.; Switanek, M.; Durcik, M.; Cobb, N. Decreased streamflow in semi-arid basins following drought-induced tree die-off: A counter-intuitive and indirect climate impact on hydrology. J. Hydrol. 2011, 406, 225–233. [Google Scholar] [CrossRef]
- Nobert, J.; Jeremiah, J. Hydrological response of watershed system to land use/cover change. A case of Wami River basin. Open Hydrol. J. 2012, 6, 78–87. [Google Scholar] [CrossRef]
- Balocchi, F.; Flores, N.; Neary, D.; White, D.; Silberstein, R.; Ramírez de Arellano, P. The effect of the ‘Las Maquinas’ wildfire of 2017 on the hydrologic balance of a high conservation value Hualo (Nothofagus glauca (Phil.) Krasser) forest in central Chile. For. Ecol. Manag. 2020, 477, 118482. [Google Scholar] [CrossRef]
- Balocchi, F.; Rivera, D.; Arumi, J.; Morgenstern, U.; White, D.; Silberstein, R.; Ramírez de Arellano, P. An Analysis of the Effects of Large Wildfires on the Hydrology of Three Small Catchments in Central Chile Using Tritium-Based Measurements and Hydrological Metrics. Hydrology 2022, 9, 45. [Google Scholar] [CrossRef]
- Bravo, C.; Loriaux, T.; Rivera, A.; Brock, B. Assesing glacier melt contribution to streamflow at Universidad Glacier, central Andes of Chile. Hydrol. Earth Syst. Sci. 2017, 21, 3249–3266. [Google Scholar] [CrossRef]
- Bown, F.; Rivera, A.; Acuña, C. Recent glacier variations at the Aconcagua basin, central Chilean Andes. Ann. Glaciol. 2008, 48, 43–48. [Google Scholar] [CrossRef]
- Le Quesne, C.; Acuña, C.; Boninsegna, J.; Rivera, A.; Barichivich, J. Long-term glacier variations in the Central Andes of Argentina and Chile, inferred from historical records and tree-ring reconstructed precipitation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 281, 334–344. [Google Scholar] [CrossRef]
- Azócar, G.; Brenning, A. Hydrological and geomorphological significance of rock glaciers in the dry Andes, Chile (27–33 S). Permafr. Periglac. Processes 2010, 21, 42–53. [Google Scholar] [CrossRef]
- Barcaza, G.; Nussbaumer, S.; Tapia, G.; Valdés, J.; García, J.; Videla, Y.; Albornoz, A.; Arias, V. Glacier inventory and recent glacier variations in the Andes of Chile, South America. Ann. Glaciol. 2017, 58, 166–180. [Google Scholar] [CrossRef]
- Ayala, Á.; Farías-Barahona, D.; Huss, M.; Pellicciotti, F.; McPhee, J.; Farinotti, D. Glacier runoff variations since 1955 in the Maipo River basin, in the semiarid Andes of central Chile. Cryosphere 2020, 14, 2005–2027. [Google Scholar] [CrossRef]
- López, F. Restauración Hidrológico Forestal de Cuencas y Control de la Erosión; Ediciones Mundi-Prensa; Tragsa y Tragsatec: Madrid, España, 1994; 902p. [Google Scholar]
- Vicente, E.; Vilagrosa, A.; Ruiz, S.; Manrique, À.; González, M.; Moutahir, H.; Chirino, E.; del Campo, A.; Bellot, J. Water balance of mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in semiarid climates: A review in a climate change context. Forests 2018, 9, 426. [Google Scholar] [CrossRef]
- Yan, W.; Deng, X.; Chen, X.; Tian, D.; Wenhua, X.; Peng, Y. Long-term variation of rainfall interception in different growth stages of Chinese fir plantation. Hydrol. Sci. J. 2015, 60, 2178–2188. [Google Scholar] [CrossRef]
- Navar, J.; Méndez, J.; González, H. Intercepción de la lluvia en especies de leguminosas del nordeste de México. Terra Latinoam. 2008, 26, 61–68. [Google Scholar]
- Huber, A.; Iroume, A.; Bathurst, J. Effect of Pinus radiate on water balance in Chile. Hydrol. Processes 2008, 22, 142–148. [Google Scholar] [CrossRef]
- Sheng, H.; Cai, T. Influence of rainfall on canopy interception in mixed broad-leaved-korean pine forest in Xiaoxing’ an mountains, Northeastern China. Forest 2019, 10, 248. [Google Scholar]
- Garcia-Chevesich, P. Erosion Control and Land Restoration; Outskirts Press: Denver, CO, USA, 2016; 486p. [Google Scholar]
- Yu, P.; Wang, Y.; Coles, N.; Xiong, W.; Xu, L. Simulation of runoff changes caused by cropland to forest conversion in the upper Yangtze river region, SW China. PLoS ONE 2015, 10, e0132395. [Google Scholar] [CrossRef]
- United Nations Educational, Scientific and Cultural Organization (UNESCO). Antecedentes de la Relación Masa Forestal y Disponibilidad Hídrica en Chile; Regional Bureau for Science in Latin America and the Caribbean, Unesco Office Montevideo: Montevideo, Uruguay, 2019; 38p. [Google Scholar]
- Brooks, R.; Barnard, H.; Coulombe, R.; McDonnell, J. Ecohydrologic separation of water between trees and streams in a Mediterranean climate. Nat. Geosci. 2010, 3, 100–104. [Google Scholar] [CrossRef]
- McDonnell, J. The two-water world hypothesis: Ecohydrological separation of water between streams and trees? Wires Water 2014, 1, 323–329. [Google Scholar] [CrossRef]
- Hervé, P.; Oyarzún, C.; Brumbt, C.; Huygens, D.; Bodé, S.; Verhoest, N.; Boeckx, P. Assessing the ‘two water world’ hypothesis and water sources for native and exotic evergreen species in south-central Chile. Hydrol. Processes 2016, 30, 4227–4241. [Google Scholar]
- Van den Heuvel, D.; Troch, P.; Booij, M.; Niu, G.; Volkman, T.; Pangle, L. Effects of differential hillslope-scale water retention characteristics on rainfall-runoff response at the landscape evolution observatory. Hydrol. Processes 2018, 32, 2118–2127. [Google Scholar] [CrossRef]
- Balocchi, F.; Flores, N.; Arumí, J.; White, D.; Silberstein, R.; Ramírez de Arellano, P. Comparison of streamflow recession between plantations and native forests in small catchments in Central-Southern Chile. Hydrol. Processes 2021, 35, e14182. [Google Scholar] [CrossRef]








| Condition | Classification | Group | FP Cover (%) | NF Cover (%) | Number of Catchments | Forested Area (%) | Pass (n ≥ 5) |
|---|---|---|---|---|---|---|---|
| 1 | Very Low Forest Plantation Cover | G1 | 0 to 4.9 | Any | 17 | 40 | Yes |
| 2 | Low Forest Plantation Cover | G2 | 5 to 14.9 | Any | 12 | 29 | Yes |
| 3 | Mid Forest Plantation Cover | G3 | 15 to 29.9 | Any | 8 | 19 | Yes |
| 4 | High Forest Plantation Cover | G4 | ≥30 | Any | 5 | 12 | Yes |
| 5 | Very Low Native Forest Cover | G5 | any | 0 to 4.9 | 4 | 10 | No |
| 6 | Low Native Forest Cover | G6 | any | 5 to 14.9 | 9 | 26 | Yes |
| 7 | Mid Native Forest Cover | G7 | any | 15 to 29.9 | 14 | 29 | Yes |
| 8 | High Native Forest Cover | G8 | any | ≥30 | 15 | 36 | Yes |
| 1–5 | Very Low Forest Plantation Cover and Very Low Native Forest Cover | G9 | 0 to 4.9 | 0 to 4.9 | 2 | 5 | No |
| 1–6 | Very Low Forest Plantation Cover and Low Native Forest Cover | G10 | 0 to 4.9 | 5 to 14.9 | 5 | 12 | Yes |
| 1–7 | Very Low Forest Plantation Cover and Mid Native Forest Cover | G11 | 0 to 4.9 | 15 to 29.9 | 2 | 5 | No |
| 1–8 | Very Low Forest Plantation Cover and High Native Forest Cover | G12 | 0 to 4.9 | ≥30 | 8 | 19 | Yes |
| 2–5 | Low Forest Plantation Cover and Very Low Native Forest Cover | G13 | 5 to 14.9 | 0 to 4.9 | 0 | 0 | No |
| 2–6 | Low Forest Plantation Cover and Low Native Forest Cover | G14 | 5 to 14.9 | 5 to 14.9 | 1 | 2 | No |
| 2–7 | Low Forest Plantation Cover and Mid Native Forest Cover | G15 | 5 to 14.9 | 15 to 29.9 | 5 | 12 | Yes |
| 2–8 | Low Forest Plantation Cover and High Native Forest Cover | G16 | 5 to 14.9 | ≥30 | 6 | 14 | Yes |
| 3–5 | Mid Forest Plantation Cover and Very Low Native Forest Cover | G17 | 15 to 29.9 | 0 to 4.9 | 0 | 0 | No |
| 3–6 | Mid Forest Plantation Cover and Low Native Forest Cover | G18 | 15 to 29.9 | 5 to 14.9 | 2 | 5 | No |
| 3–7 | Mid Forest Plantation Cover and Mid Native Forest Cover | G19 | 15 to 29.9 | 15 to 29.9 | 5 | 12 | Yes |
| 3–8 | Mid Forest Plantation Cover and High Native Forest Cover | G20 | 15 to 29.9 | ≥30 | 1 | 2 | No |
| 4–5 | High Forest Plantation Cover and Very Low Native Forest Cover | G21 | ≥30 | 0 to 4.9 | 2 | 5 | No |
| 4–6 | High Forest Plantation Cover and Low Native Forest Cover | G22 | ≥30 | 5 to 14.9 | 1 | 2 | No |
| 4–7 | High Forest Plantation Cover and Mid Native Forest Cover | G23 | ≥30 | 15 to 29.9 | 2 | 5 | No |
| 4–8 | High Forest Plantation Cover and High Native Forest Cover | G24 | ≥30 | ≥30 | 0 | 0 | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pizarro, R.; Valdés-Pineda, R.; Garcia-Chevesich, P.A.; Ibáñez, A.; Pino, J.; Scott, D.F.; Neary, D.G.; McCray, J.E.; Castillo, M.; Ubilla, P. The Large-Scale Effect of Forest Cover on Long-Term Streamflow Variations in Mediterranean Catchments of Central Chile. Sustainability 2022, 14, 4443. https://doi.org/10.3390/su14084443
Pizarro R, Valdés-Pineda R, Garcia-Chevesich PA, Ibáñez A, Pino J, Scott DF, Neary DG, McCray JE, Castillo M, Ubilla P. The Large-Scale Effect of Forest Cover on Long-Term Streamflow Variations in Mediterranean Catchments of Central Chile. Sustainability. 2022; 14(8):4443. https://doi.org/10.3390/su14084443
Chicago/Turabian StylePizarro, Roberto, Rodrigo Valdés-Pineda, Pablo A. Garcia-Chevesich, Alfredo Ibáñez, Juan Pino, David F. Scott, Daniel G. Neary, John E. McCray, Miguel Castillo, and Patricio Ubilla. 2022. "The Large-Scale Effect of Forest Cover on Long-Term Streamflow Variations in Mediterranean Catchments of Central Chile" Sustainability 14, no. 8: 4443. https://doi.org/10.3390/su14084443
APA StylePizarro, R., Valdés-Pineda, R., Garcia-Chevesich, P. A., Ibáñez, A., Pino, J., Scott, D. F., Neary, D. G., McCray, J. E., Castillo, M., & Ubilla, P. (2022). The Large-Scale Effect of Forest Cover on Long-Term Streamflow Variations in Mediterranean Catchments of Central Chile. Sustainability, 14(8), 4443. https://doi.org/10.3390/su14084443