
Conditioning of Feed Material Prior to Feeding: Approaches for a Sustainable Phosphorus Utilization

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Background
2.1. P Uptake by Plants
2.2. P in Animal Feed Sources
2.3. P-Digestion in Monogastric Animals
2.4. P Occurrence in Animal Manure
2.5. P in Soils and Run-Off
2.6. P Losses in Agriculture
3. State-of-the-Art: Existing Approaches for Improved P Digestibility in Animal Feed and Their Respective Limitations
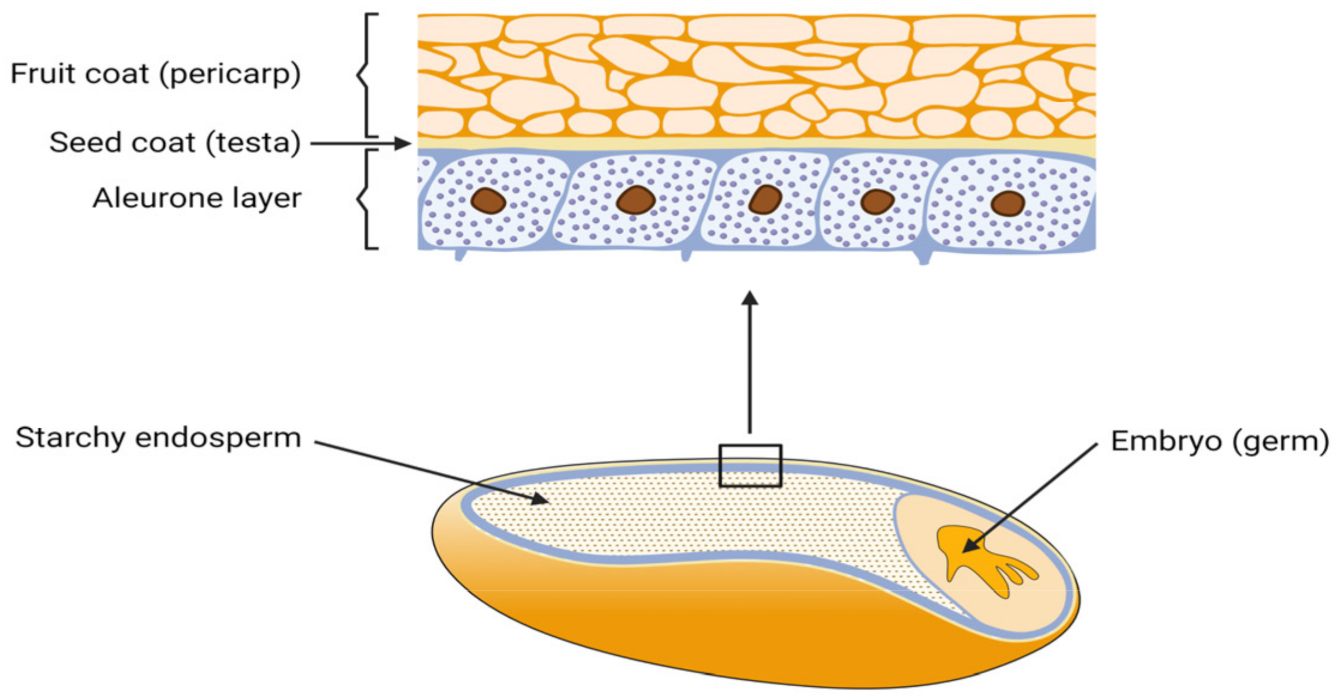
3.1. Mechanical P Separation from Whole Grains
3.1.1. Debranning
3.1.2. Specific Layer Exclusion
3.2. Enzymatic Feed Treatment
3.2.1. Feeding of Phytase-Rich Feed
3.2.2. Germination of Phytate-Rich Feed
3.2.3. Phytase as Feed Additive
3.2.4. Enzymatic Hydrolysis
3.3. Chemical Feed Treatment
3.3.1. Hydrothermal Treatment
- Since phytate is water-soluble, a certain proportion is dissolved into the liquid phase and, therefore, eliminated from the substrate [31].
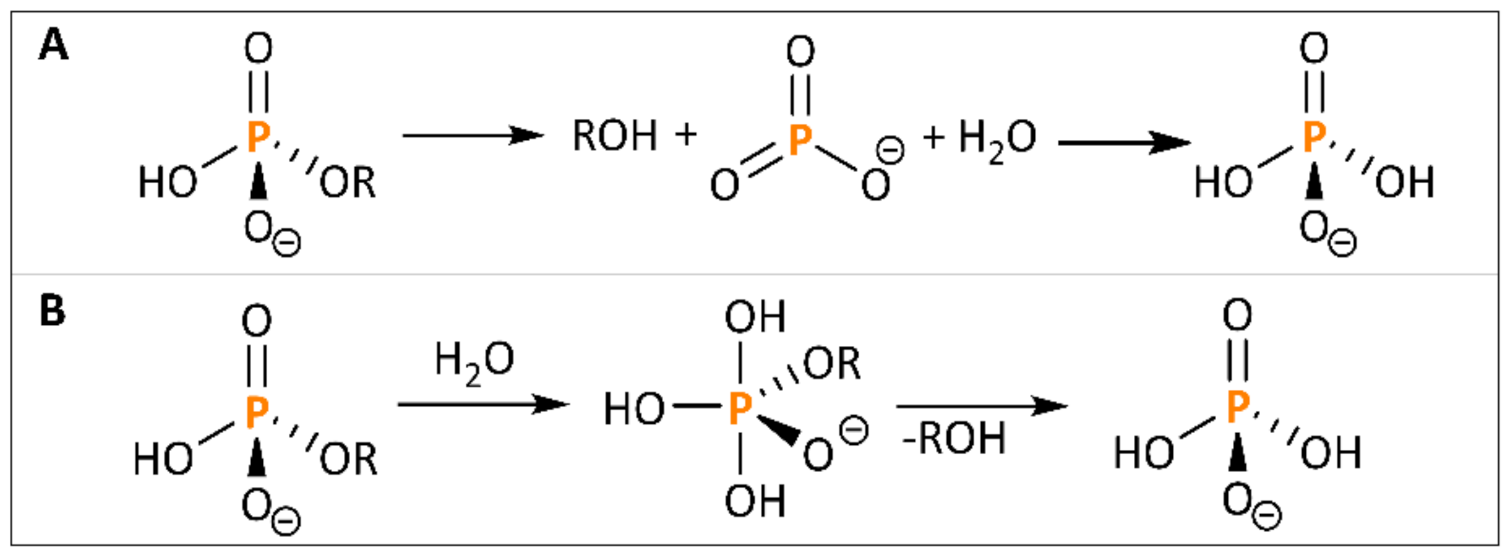
3.3.2. Acid Hydrolysis
4. Conceptual Design of a Feed Material Conditioning Process
4.1. Process Options for Feed Material Conditioning
- Option i: → Mechanical pre-separation & enzymatic hydrolysis
- Option ii: → Mechanical pre-separation & chemical hydrolysis
- Option iii: → Enzymatic hydrolysis
- Option iv: → Chemical hydrolysis
4.2. Qualitative Evaluation of the Process Options
- The process complexity refers to the number of individual process steps and therefore the requirement for, for example, additional tanks, pumps, and process peripherals.
- For energy demand, only a rough estimation considering temperature, electricity, and energy input for drying of the conditioned feed material is provided. However, energy demand is one of the primary economic factors and thus taken into account.
- The harm of reagents is evaluated in terms of impact on human health and the environment. It comprises toxicity as well as acidic and basic effects, among other types of contamination.
- The maturity of the process indicates how far the process is from industrial application in view of a potential feed material conditioning process. The current state of research and already established methods are taken into account.
- The rate of free P provision refers to the amount of recovered P and the conversion into industrial relevant compounds (e.g., feed, food or fertilizer). Here, the estimation is based on results derived from ongoing research (Section 3).
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| DNA1 | Deoxyribonucleic acid |
| inositol-P | inositol-phosphate |
| InsP1 | inositol-monophosphate |
| InsP3 | inositol-triphosphate |
| InsP4 | inositol-tetraphosphate |
| InsP6 | inositol-hexaphosphate |
| MtP/a | 106 metric tons phosphorus per year |
| P | phosphorus/ phosphate |
| phytate-P | phytate-bound phosphorus |
| PI | inorganic phosphate |
| PO | organic phosphate |
| polyP | polyphosphate |
| RNA | Ribonucleic acid |
| U | Unit [µmol/min] |
References
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannah, R.; Max, R. Meat and Dairy Production. Available online: https://ourworldindata.org/ (accessed on 26 November 2021).
- Herrmann, K.R.; Ruff, A.J.; Schwaneberg, U. Phytase-based phosphorus recovery process for 20 distinct press cakes. Sustain. Chem. Eng. 2020, 8, 3913–3921. [Google Scholar] [CrossRef]
- Reta, G.; Dong, X.; Li, Z.; Su, B.; Hu, X.; Bo, H.; Yu, D.; Wan, H.; Liu, J.; Li, Y.; et al. Environmental impact of phosphate mining and beneficiation: Review. IJH 2018, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Blengini, G.A.; El Latunussa, C.; Eynard, U.; Torres de Matos, C.; Wittmer, D.M.A.G.; Georgitzikis, K.; Pavel, C.C.; Carrara, S.; Mancini, L.; Unguru, M.; et al. Study on the EU’s List of Critical Raw Materials (2020): Final Report; Publications Office of the European Union: Luxembourg, 2020; ISBN 9789276210498.
- Schipanski, M.E.; Bennett, E.M. The Influence of Agricultural Trade and Livestock Production on the Global Phosphorus Cycle. Ecosystems 2012, 15, 256–268. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, S.; Sheng, H.; Liu, X.; Hua, H.; Liu, X.; Zhang, Y. Human Perturbation of the Global Phosphorus Cycle: Changes and Consequences. Environ. Sci. Technol. 2018, 52, 2438–2450. [Google Scholar] [CrossRef] [PubMed]
- Smil, V. Phosphorus in the environment: Natural Flows and Human Interferences. Annu. Rev. Energy Environ. 2000, 25, 53–88. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, K.R.; Ruff, A.J.; Infanzón, B.; Schwaneberg, U. Engineered phytases for emerging biotechnological applications beyond animal feeding. Appl. Microbiol. Biotechnol. 2019, 103, 6435–6448. [Google Scholar] [CrossRef]
- Hirvonen, J.; Liljavirta, J.; Saarinen, M.T.; Lehtinen, M.J.; Ahonen, I.; Nurminen, P. Effect of Phytase on in Vitro Hydrolysis of Phytate and the Formation of myo-Inositol Phosphate Esters in Various Feed Materials. J. Agric. Food Chem. 2019, 67, 11396–11402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; He, Z.; Jaisi, D.P. Role of metal complexation on the solubility and enzymatic hydrolysis of phytate. PLoS ONE 2021, 16, e0255787. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Chung, T.K.; Moughan, P.J. Effect of microbial phytase on phytate P degradation and apparent digestibility of total P and Ca throughout the gastrointestinal tract of the growing pig. J. Anim. Sci. 2014, 92, 189–197. [Google Scholar] [CrossRef]
- Shaddel, S.; Bakhtiary-Davijany, H.; Kabbe, C.; Dadgar, F.; Østerhus, S. Sustainable Sewage Sludge Management: From Current Practices to Emerging Nutrient Recovery Technologies. Sustainability 2019, 11, 3435. [Google Scholar] [CrossRef] [Green Version]
- Filippelli, G.M. The Global Phosphorus Cycle: Past, Present, and Future. Elements 2008, 4, 89–95. [Google Scholar] [CrossRef]
- Stewart, W.M.; Dibb, D.W.; Johnston, A.E.; Smyth, T.J. The Contribution of Commercial Fertilizer Nutrients to Food Production. Agron. J. 2005, 97, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chien, S.H.; Prochnow, L.I.; Cantarella, H. Recent Developments of Fertilizer Production and Use to Improve Nutrient Efficiency and Minimize Environmental Impacts. In Recent Developments of Fertilizer Production and Use to Improve Nutrient Efficiency and Minimize Environmental Impacts; Elsevier: Amsterdam, The Netherlands, 2009; Chapter 8; pp. 267–322. ISBN 9780123748188. [Google Scholar]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus Uptake by Plant: From Soil to Cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.W. The phosphate uptake mechanism. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Springer: Dordrecht, The Netherlands, 2002; pp. 235–244. [Google Scholar] [CrossRef]
- King, K.W.; Williams, M.R.; Macrae, M.L.; Fausey, N.R.; Frankenberger, J.; Smith, D.R.; Kleinman, P.J.A.; Brown, L.C. Phosphorus transport in agricultural subsurface drainage: A review. J. Environ. Qual. 2015, 44, 467–485. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Xu, C.; Ridoutt, B.G.; Wang, X.; Ren, P. Nitrogen and phosphorus losses and eutrophication potential associated with fertilizer application to cropland in China. J. Clean. Prod. 2017, 159, 171–179. [Google Scholar] [CrossRef]
- Directive, E.U.W. Council Directive of 21 May 1991 concerning urban waste water treatment (91/271/EEC). J. Eur. Commun. 1991, 34, 40. [Google Scholar]
- Sengupta, S.; Pandit, A. Selective removal of phosphorus from wastewater combined with its recovery as a solid-phase fertilizer. Water Res. 2011, 45, 3318–3330. [Google Scholar] [CrossRef]
- Wang, X.-X.; Hoffland, E.; Feng, G.; Kuyper, T.W. Phosphate Uptake from Phytate Due to Hyphae-Mediated Phytase Activity by Arbuscular Mycorrhizal Maize. Front. Plant Sci. 2017, 8, 684. [Google Scholar] [CrossRef] [Green Version]
- Barrow, N.J. The effects of pH on phosphate uptake from the soil. Plant Soil 2017, 410, 401–410. [Google Scholar] [CrossRef]
- Kobae, Y. Dynamic Phosphate Uptake in Arbuscular Mycorrhizal Roots Under Field Conditions. Front. Environ. Sci. 2019, 6, 1216. [Google Scholar] [CrossRef] [Green Version]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sengupta, S.; Karmakar, A.; Sarkar, S.N.; Gangopadhyay, G.; Datta, K.; Datta, S.K. Genetically engineered rice with appA gene enhanced phosphorus and minerals. J. Plant Biochem. Biotechnol. 2019, 28, 470–482. [Google Scholar] [CrossRef]
- Eeckhout, W.; de Paepe, M. Total phosphorus, phytate-phosphorus and phytase activity in plant feedstuffs. Anim. Feed Sci. Technol. 1994, 47, 19–29. [Google Scholar] [CrossRef]
- Rodehutscord, M.; Rückert, C.; Maurer, H.P.; Schenkel, H.; Schipprack, W.; Bach Knudsen, K.E.; Schollenberger, M.; Laux, M.; Eklund, M.; Siegert, W.; et al. Variation in chemical composition and physical characteristics of cereal grains from different genotypes. Arch. Anim. Nutr. 2016, 70, 87–107. [Google Scholar] [CrossRef]
- Viveros, A.; Centeno, C.; Brenes, A.; Canales, R.; Lozano, A. Phytase and acid phosphatase activities in plant feedstuffs. J. Agric. Food Chem. 2000, 48, 4009–4013. [Google Scholar] [CrossRef] [Green Version]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Hendriks, W.; Moughan, P. Whole-body mineral composition of entire male and female pigs depositing protein at maximal rates. Livest. Prod. Sci. 1993, 33, 161–170. [Google Scholar] [CrossRef]
- Mahan, D.C.; Shields, R.G. Macro- and micromineral composition of pigs from birth to 145 kilograms of body weight. J. Anim. Sci. 1998, 76, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Liao, X.; Luo, X. Nutritional strategies for reducing nitrogen, phosphorus and trace mineral excretions of livestock and poultry. J. Integr. Agric. 2017, 16, 2815–2833. [Google Scholar] [CrossRef]
- Schulz, E.; Oslage, H.J. Untersuchungen zur intestinalen Hydrolyse von Inositphosphorsäureester und zur Absorption von Phytinphosphor beim Schwein. Z. Tierphysiol. Tierernährung Futterm. 1972, 30, 76–91. [Google Scholar] [CrossRef]
- Pallauf, J.; Rimbach, G. Nutritional significance of phytic acid and phytase. Arch. Tierernahr. 1997, 50, 301–319. [Google Scholar] [CrossRef]
- Walk, C.L. Phytate Destruction-Consequences for Precision Animal Nutrition, 1st ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; ISBN 9789086868360. [Google Scholar]
- Thompson, L.U.; Yoon, J.H. Starch Digestibility as Affected by Polyphenols and Phytic Acid. J. Food Sci. 1984, 49, 1228–1229. [Google Scholar] [CrossRef]
- Linares, L.B.; Broomhead, J.N.; Guaiume, E.A.; Ledoux, D.R.; Veum, T.L.; Raboy, V. Effects of low phytate barley (Hordeum vulgare L.) on zinc utilization in young broiler chicks. Poult. Sci. 2007, 86, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ru, Y.J.; Cowieson, A.J.; Li, F.D.; Cheng, X.C. Effects of phytate and phytase on the performance and immune function of broilers fed nutritionally marginal diets. Poult. Sci. 2008, 87, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ru, Y.J.; Di Li, F.; Wang, J.-P.; Lei, X.-Q. Effect of dietary phytate and phytase on proteolytic digestion and growth regulation of broilers. Arch. Anim. Nutr. 2009, 63, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Onyango, E.M.; Adeola, O. Dietary phytate (inositol hexaphosphate) regulates the activity of intestinal mucosa phytase. J. Anim. Physiol. Anim. Nutr. 2009, 93, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Weihrauch, D.; Nyachoti, C.M. Effect of dietary phytic acid on performance and nutrient uptake in the small intestine of piglets. J. Anim. Sci. 2012, 90, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Rieger, H. Untersuchungen zum Einfluss Einer Unterschiedlichen Phosphorversorgung auf die Entwicklung und Mineralisation Verschiedener Knochen Wachsender Schweine: Dissertation; University of Veterinary Medicine: Hannover, Germany, 2017. [Google Scholar]
- Kumaragamage, D.; Akinremi, O.O. Manure Phosphorus: Mobility in Soils and Management Strategies to Minimize Losses. Curr. Pollut. Rep. 2018, 4, 162–174. [Google Scholar] [CrossRef]
- Pagliari, P.H.; Laboski, C.A.M. Investigation of the inorganic and organic phosphorus forms in animal manure. J. Environ. Qual. 2012, 41, 901–910. [Google Scholar] [CrossRef]
- Liu, J.; Spargo, J.T.; Kleinman, P.J.A.; Meinen, R.; Moore, P.A.; Beegle, D.B. Water-Extractable Phosphorus in Animal Manure and Manure Compost: Quantities, Characteristics, and Temporal Changes. J. Environ. Qual. 2018, 47, 471–479. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Pagliari, P.H.; WALDRIP, H.M. Applied and Environmental Chemistry of Animal Manure: A Review. Pedosphere 2016, 26, 779–816. [Google Scholar] [CrossRef]
- Vadas, P.A.; Good, L.W.; Jokela, W.E.; Karthikeyan, K.G.; Arriaga, F.J.; Stock, M. Quantifying the Impact of Seasonal and Short-term Manure Application Decisions on Phosphorus Loss in Surface Runoff. J. Environ. Qual. 2017, 46, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajiboye, B.; Akinremi, O.O.; Racz, G.J. Laboratory Characterization of Phosphorus in Fresh and Oven-Dried Organic Amendments. Envorin. Qual. 2004, 33, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Darch, T.; Blackwell, M.S.A.; Hawkins, J.M.B.; Haygarth, P.M.; Chadwick, D. A Meta-Analysis of Organic and Inorganic Phosphorus in Organic Fertilizers, Soils, and Water: Implications for Water Quality. Crit. Rev. Environ. Sci. Technol. 2014, 44, 2172–2202. [Google Scholar] [CrossRef]
- Wang, F.; Deng, M.; Xu, J.; Zhu, X.; Mao, C. Molecular mechanisms of phosphate transport and signaling in higher plants. Semin. Cell Dev. Biol. 2018, 74, 114–122. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, T.Q.; Tan, C.S.; Vadas, P.; Qi, Z.M.; Wellen, C. Modeling phosphorus losses from soils amended with cattle manures and chemical fertilizers. Sci. Total Environ. 2018, 639, 580–587. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Schoumans, O.F.; Bouraoui, F.; Kabbe, C.; Oenema, O.; van Dijk, K.C. Phosphorus management in Europe in a changing world. Ambio 2015, 44 (Suppl. S2), S180–S192. [Google Scholar] [CrossRef] [Green Version]
- Finger, R.; Swinton, S.M.; El Benni, N.; Walter, A. Precision Farming at the Nexus of Agricultural Production and the Environment. Annu. Rev. Resour. Econ. 2019, 11, 313–335. [Google Scholar] [CrossRef] [Green Version]
- Möller, K.; Oberson, A.; Bünemann, E.K.; Cooper, J.; Friedel, J.K.; Glæsner, N.; Hörtenhuber, S.; Løes, A.-K.; Mäder, P.; Meyer, G.; et al. Improved Phosphorus Recycling in Organic Farming: Navigating Between Constraints; Elsevier: Amsterdam, The Netherlands, 2018; pp. 159–237. ISBN 9780128152836. [Google Scholar]
- Caballero, B.; Finglas, O.; Toldrá, F. Encyclopedia of food Sciences and Nutrition, 2nd ed.; Academic Press: Amsterdam, The Netherlands, 2003; ISBN 9780122270550. [Google Scholar]
- Prückler, M.; Siebenhandl-Ehn, S.; Apprich, S.; Höltinger, S.; Haas, C.; Schmid, E.; Kneifel, W. Wheat bran-based biorefinery 1: Composition of wheat bran and strategies of functionalization. LWT 2014, 56, 211–221. [Google Scholar] [CrossRef]
- Knight, S. Grain and Feed Annual_London_EU-28_4-15-2019. Available online: https://apps.fas.usda.gov/newgainapi/api/report/downloadreportbyfilename?filename=Grain%20and%20Feed%20Annual_London_EU-28_4-15-2019.pdf (accessed on 3 February 2022).
- Loy, D.D.; Lundy, E.L. Nutritional Properties and Feeding Value of Corn and Its Coproducts. In Corn; AACC International Press: Oxford, UK, 2019; pp. 633–659. ISBN 9780128119716. [Google Scholar]
- Andersson, A.A.M.; Andersson, R.; Jonsäll, A.; Andersson, J.; Fredriksson, H. Effect of Different Extrusion Parameters on Dietary Fiber in Wheat Bran and Rye Bran. J Food Sci. 2017, 82, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.; Mosenthin, R.; Zimmermann, B.; Greiner, R.; Roth, S. Distribution of phytase activity, total phosphorus and phytate phosphorus in legume seeds, cereals and cereal by-products as influenced by harvest year and cultivar. Anim. Feed Sci. Technol. 2007, 133, 320–334. [Google Scholar] [CrossRef]
- Alam, S.A.; Järvinen, J.; Kirjoranta, S.; Jouppila, K.; Poutanen, K.; Sozer, N. Influence of Particle Size Reduction on Structural and Mechanical Properties of Extruded Rye Bran. Food Bioprocess Technol. 2014, 7, 2121–2133. [Google Scholar] [CrossRef]
- Ebringerová, A.; Hromádková, Z. Effect of ultrasound on the extractibility of corn bran hemicelluloses. Ultrason. Sonochem. 2002, 9, 225–229. [Google Scholar] [CrossRef]
- Belyea, R.L.; Rausch, K.D.; Tumbleson, M.E. Composition of corn and distillers dried grains with solubles from dry grind ethanol processing. Bioresour. Technol. 2004, 94, 293–298. [Google Scholar] [CrossRef]
- Fretzdorff, B.; Weipert, D. Phytinsäure in Getreide und Getreideerzeugnissen. Mitteilung 1: Phytinsäure und Phytase in Roggen und Roggenprodukten. Bundesforschungsanst. Getreide-Kartoff. 1986, 182, 287–293. [Google Scholar]
- Luithui, Y.; Baghya Nisha, R.; Meera, M.S. Cereal by-products as an important functional ingredient: Effect of processing. J. Food Sci. Technol. 2019, 56, 1–11. [Google Scholar] [CrossRef]
- Declour, A.J.; Poutanen, K. Fibre-Rich and Wholegrain Foods: Improving Quality; Woodhead Pub Ltd.: Oxford, UK, 2013; ISBN 9780857090386. [Google Scholar]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [Green Version]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat aleurone: Separation, composition, health aspects, and potential food use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef] [Green Version]
- Rausch, K.D.; Hummel, D.; Johnson, L.A.; May, J.B. Wet Milling: The Basis for Corn Biorefineries. In Corn; Elsevier: Amsterdam, The Netherlands, 2019; pp. 501–535. ISBN 9780128119716. [Google Scholar]
- Comm/Dg/Unit. Cereals Statistics. Available online: https://ec.europa.eu/info/food-farming-fisheries/farming/facts-and-figures/markets/overviews/market-observatories/crops/cereals-statistics_en (accessed on 15 October 2021).
- Anderson, B.; Almeida, H. Corn Dry Milling: Processes, Products, and Applications. In Corn; Elsevier: Amsterdam, The Netherlands, 2019; pp. 405–433. ISBN 9780128119716. [Google Scholar]
- Oloffs, K.; Cossa, J.; Jeroch, H. Die Bedeutung der korneigenen (nativen) Phytaseaktivität im Weizen für die Phosphor-Verwertung bei Broilern und Legehennen. Archiv. Geflugelkd. 2000, 64, 157–161. [Google Scholar]
- Pointillart, A. Enhancement of phosphorus utilization in growing pigs fed phytate-rich diets by using rye bran. J. Anim. Sci. 1991, 69, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Düngelhoef, M.; Rodehutscord, M.; Spiekers, H.; Pfeffer, H.S.E. Effects of supplemental microbial phytase on availability of phosphorus contained in maize, wheat and triticale to pigs. Anim. Feed Sci. Technol. 1994, 49, 1–10. [Google Scholar] [CrossRef]
- Hovenjürgen, M.; Pfeffer, E.; Rodehutscord, M. Effect of fertilization and variety on digestability of phosphorus from plant feedstuffs in pigs. J. Anim. Feed Sci. 2003, 12, 83–93. [Google Scholar] [CrossRef]
- Rodehutscord, M.; Faust, M.; Lorenz, H. Digestibility of phosphorus contained in soybean meal, barley, and different varieties of wheat, without and with supplemental phytase fed to pigs and additivity of digestibility in a wheatsoybean-meal diet. J. Anim. Physiol. Anim. Nutr. 1996, 75, 40–48. [Google Scholar] [CrossRef]
- Rodehutscord, M. Der gegenwärtige Stand der Phosphorbewertung für Nutztiere. Lohmann Inf. 2001, 1, 26–34. [Google Scholar]
- Jongbloed, A.W.; Kemme, P.A. Effect of pelleting mixed feeds on phytase activity and the apparent absorbability of phosphorus and calcium in pigs. Anim. Feed Sci. Technol. 1990, 28, 233–242. [Google Scholar] [CrossRef]
- Kemme, P.A.; Jongbloed, A.W.; Mroz, Z.; Beynen, A.C. Diurnal variation in degradation of phytic acid by plant phytase in the pig stomach. Livest. Prod. Sci. 1998, 54, 33–44. [Google Scholar] [CrossRef]
- Schlemmer, U.; Jany, K.D.; Berk, A.; Schulz, E.; Rechkemmer, G. Degradation of phytate in the gut of pigs-pathway of gastro-intestinal inositol phosphate hydrolysis and enzymes involved. Arch. Tierernahr. 2001, 55, 255–280. [Google Scholar] [CrossRef]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53 (Suppl. S2), S330–S375. [Google Scholar] [CrossRef]
- Greiner, R.; Konietzny, U. Phytase for Food Application. Food Technol. Biotechnol. 2006, 44, 125–140. [Google Scholar]
- Pal, R.S.; Bhartiya, A.; Yadav, P.; Kant, L.; Mishra, K.K.; Aditya, J.P.; Pattanayak, A. Effect of dehulling, germination and cooking on nutrients, anti-nutrients, fatty acid composition and antioxidant properties in lentil (Lens culinaris). J. Food Sci. Technol. 2017, 54, 909–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, R.; Alminger, M.L. Purification and characterization of a phytate-degrading enzyme from germinated oat (Avena sativa). J. Sci. Food Agric. 1999, 79, 1453–1460. [Google Scholar] [CrossRef]
- Pakfetrat, S.; Amiri, S.; Radi, M.; Abedi, E.; Torri, L. Reduction of phytic acid, aflatoxins and other mycotoxins in wheat during germination. J. Sci. Food Agric. 2019, 99, 4695–4701. [Google Scholar] [CrossRef] [PubMed]
- Reddy, N.R.; Sathe, S.K.; Salunkhe, D.K. Phytates in Legumes and Cereals. In Phytates in Legumes and Cereals; Advances in Food Research, Ed.; Elsevier: Amsterdam, The Netherlands, 1982; pp. 1–92. ISBN 9780120164288. [Google Scholar]
- Singh, B.; Satyanarayana, T. Fungal phytases: Characteristics and amelioration of nutritional quality and growth of non-ruminants. J. Anim. Physiol. Anim. Nutr. 2015, 99, 646–660. [Google Scholar] [CrossRef] [PubMed]
- Ariza, A.; Moroz, O.V.; Blagova, E.V.; Turkenburg, J.P.; Waterman, J.; Roberts, S.M.; Vind, J.; Sjøholm, C.; Lassen, S.F.; de Maria, L.; et al. Degradation of phytate by the 6-phytase from Hafnia alvei: A combined structural and solution study. PLoS ONE 2013, 8, e65062. [Google Scholar] [CrossRef] [Green Version]
- Haefner, S.; Knietsch, A.; Scholten, E.; Braun, J.; Lohscheidt, M.; Zelder, O. Biotechnological production and applications of phytases. Appl. Microbiol. Biotechnol. 2005, 68, 588–597. [Google Scholar] [CrossRef]
- Lassen, S.F.; Breinholt, J.; Østergaard, P.R.; Brugger, R.; Bischoff, A.; Wyss, M.; Fuglsang, C.C. Expression, gene cloning, and characterization of five novel phytases from four basidiomycete fungi: Peniophora lycii, Agrocybe pediades, a Ceriporia sp., and Trametes pubescens. Appl. Environ. Microbiol. 2001, 67, 4701–4707. [Google Scholar] [CrossRef] [Green Version]
- Greiner, R.; Konietzny, U.; Jany, K.D. Purification and characterization of two phytases from Escherichia coli. Arch. Biochem. Biophys. 1993, 303, 107–113. [Google Scholar] [CrossRef]
- Rodriguez, E.; Han, Y.; Lei, X.G. Cloning, sequencing, and expression of an Escherichia coli acid phosphatase/phytase gene (appA2) isolated from pig colon. Biochem. Biophys. Res. Commun. 1999, 257, 117–123. [Google Scholar] [CrossRef]
- Augspurger, N.I.L.; Webel, D.M.; Lei, X.G.; Baker, D.H. Efficacy of an E. coli phytase expressed in yeast for releasing phytate-bound phosphorus in young chicks and pigs. J. Anim. Sci. 2003, 81, 474–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Luo, H.; Wang, Y.; Fu, D.; Shao, N.; Wang, G.; Yang, P.; Yao, B. A novel phytase from Yersinia rohdei with high phytate hydrolysis activity under low pH and strong pepsin conditions. Appl. Microbiol. Biotechnol. 2008, 80, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Hang, H.; Zhu, T.; Zhuang, Y.; Chu, J.; Zhang, S. Effect of glycosylation on biochemical characterization of recombinant phytase expressed in Pichia pastoris. Enzyme Microb. Technol. 2008, 42, 340–345. [Google Scholar] [CrossRef]
- Scheers, N.; Sandberg, A.-S. Usefulness of microbial phytases to improve zinc and iron bioavailability. In Proceedings of the International Phytase Summit 2012, Rome, Italy, 11–13 December 2012. [Google Scholar]
- Wu, T.-H.; Chen, C.-C.; Cheng, Y.-S.; Ko, T.-P.; Lin, C.-Y.; Lai, H.-L.; Huang, T.-Y.; Liu, J.-R.; Guo, R.-T. Improving specific activity and thermostability of Escherichia coli phytase by structure-based rational design. J. Biotechnol. 2014, 175, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rebello, S.; Jose, L.; Sindhu, R.; Aneesh, E.M. Molecular advancements in the development of thermostable phytases. Appl. Microbiol. Biotechnol. 2017, 101, 2677–2689. [Google Scholar] [CrossRef]
- Wang, X.; Du, J.; Zhang, Z.-Y.; Fu, Y.-J.; Wang, W.-M.; Liang, A.-H. A rational design to enhance the resistance of Escherichia coli phytase appA to trypsin. Appl. Microbiol. Biotechnol. 2018, 102, 9647–9656. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Yang, P.; Luo, H.; Huang, H.; Wang, Y.; Yao, B. Engineering of Yersinia Phytases to Improve Pepsin and Trypsin Resistance and Thermostability and Application Potential in the Food and Feed Industry. J. Agric. Food Chem. 2017, 65, 7337–7344. [Google Scholar] [CrossRef]
- Han, N.; Miao, H.; Yu, T.; Xu, B.; Yang, Y.; Wu, Q.; Zhang, R.; Huang, Z. Enhancing thermal tolerance of Aspergillus niger PhyA phytase directed by structural comparison and computational simulation. BMC Biotechnol. 2018, 18, 36. [Google Scholar] [CrossRef] [Green Version]
- Shivange, A.V.; Schwaneberg, U. Recent Advances in Directed Phytase Evolution and Rational Phytase Engineering. In Directed Enzyme Evolution: Advances and Applications; Alcalde, M., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 145–172. ISBN 978-3-319-50411-7. [Google Scholar]
- Tan, H.; Wu, X.; Xie, L.; Huang, Z.; Peng, W.; Gan, B. Identification and characterization of a mesophilic phytase highly resilient to high-temperatures from a fungus-garden associated metagenome. Appl. Microbiol. Biotechnol. 2016, 100, 2225–2241. [Google Scholar] [CrossRef]
- Ushasree, M.V.; Shyam, K.; Vidya, J.; Pandey, A. Microbial phytase: Impact of advances in genetic engineering in revolutionizing its properties and applications. Bioresour. Technol. 2017, 245, 1790–1799. [Google Scholar] [CrossRef]
- Farias, N.; Almeida, I.; Meneses, C. New Bacterial Phytase through Metagenomic Prospection. Molecules 2018, 23, 448. [Google Scholar] [CrossRef] [Green Version]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Flachowsky, G.; Gropp, J.; Kolar, B.; et al. Safety and efficacy of Natuphos® E (6-phytase) as a feed additive for avian and porcine species. EFSA J. 2017, 15, e05024. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.; Hofmann, I.; Jungherz, D.; Wittwer, M.; Infanzón, B.; Hamer, S.N.; Davari, M.D.; Ruff, A.J.; Schwaneberg, U. Generation of phytase chimeras with low sequence identities and improved thermal stability. J. Biotechnol. 2021, 339, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Konietzny, U.; Greiner, R. Molecular and catalytic properties of phytate-degrading enzymes (phytases). Int. J. Food Sci. Technol. 2002, 37, 791–812. [Google Scholar] [CrossRef] [Green Version]
- Wyss, M.; Brugger, R.; Kronenberger, A.; Rémy, R.; Fimbel, R.; Oesterhelt, G.; Lehmann, M.; van Loon, A.P. Biochemical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): Catalytic properties. Appl. Environ. Microbiol. 1999, 65, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Zeller, E.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Hydrolysis of phytate and formation of inositol phosphate isomers without or with supplemented phytases in different segments of the digestive tract of broilers. J. Nutr. Sci. 2015, 4, e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontoppidan, K.; Glitsoe, V.; Guggenbuhl, P.; Quintana, A.P.; Nunes, C.S.; Pettersson, D.; Sandberg, A.-S. In vitro and in vivo degradation of myo-inositol hexakisphosphate by a phytase from Citrobacter braakii. Arch. Anim. Nutr. 2012, 66, 431–444. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Davin, R.; Christensen, T.; Kwakernaak, C. Effect of two phytases at two doses on performance and phytate degradation in broilers during 1–21 days of age. PLoS ONE 2021, 16, e0247420. [Google Scholar] [CrossRef]
- Infanzón, B.; Herrmann, K.R.; Hofmann, I.; Willbold, S.; Ruff, A.J.; Schwaneberg, U. Phytase blends for enhanced phosphorous mobilization of deoiled seeds. Enzym. Microb. Technol. 2021, 153, 109953. [Google Scholar] [CrossRef]
- Espinosa, C.D.; Oliveira, M.S.F.; Velayudhan, D.E.; Dersjant-Li, Y.; Stein, H.H. Influence of a novel consensus bacterial 6-phytase variant on mineral digestibility and bone ash in young growing pigs fed diets with different concentrations of phytate-bound phosphorus. J. Anim. Sci. 2021, 99, skab211. [Google Scholar] [CrossRef]
- Lee, S.A.; Febery, E.; Wilcock, P.; Bedford, M.R. Application of Creep Feed and Phytase Super-Dosing as Tools to Support Digestive Adaption and Feed Efficiency in Piglets at Weaning. Animals 2021, 11, 2080. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Cowieson, A.J.; Wilson, J.W.; Ajuwon, K.M.; Adeola, O. Extra-phosphoric effects of super dosing phytase on growth performance of pigs is not solely due to release of myo-inositol. J. Anim. Sci. 2019, 97, 3898–3906. [Google Scholar] [CrossRef] [PubMed]
- Noureddini, H.; Dang, J. Degradation of phytates in distillers’ grains and corn gluten feed by Aspergillus niger phytase. Appl. Biochem. Biotechnol. 2009, 159, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Christ, J.J.; Smith, S.A.; Willbold, S.; Morrissey, J.H.; Blank, L.M. Biotechnological synthesis of water-soluble food-grade polyphosphate with Saccharomyces cerevisiae. Biotechnol. Bioeng. 2020, 117, 2089–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitmaier, E.; Jung, G. Organische Chemie: Grundlagen, Verbindungsklassen, Reaktionen, Konzepte, Molekülstruktur, Naturstof; Georg Thieme Verlag: New York, NY, USA, 2009; ISBN 9783135415062. [Google Scholar]
- Sun, X.; Liu, Z.; Qu, Y.; Li, X. The Effects of Wheat bran Comosition on the Production of Biomass-Hydrolyzing Enzymes by Penicillium decumbens. In Biotechnology for Fuels and Chemicals; Humana Press: Totowa, NJ, USA, 2008; pp. 119–128. [Google Scholar] [CrossRef]
- Wu, X.; Fu, J.; Lu, X. Kinetics and Mechanism of Hydrothermal Decomposition of Lignin Model Compounds. Ind. Eng. Chem. Res. 2013, 52, 5016–5022. [Google Scholar] [CrossRef]
- Moriyoshi, T.; Sam, K.; Uosaki, Y. Hydrothermal decomposition of esters under high pressure. High Press. Res. 2001, 20, 491–505. [Google Scholar] [CrossRef]
- Vincent, J.B.; Crowder, M.W.; Averill, B.A. Hydrolysis of phosphate monoesters: A biological problem with multiple chemical solutions. Trends Biochem. Sci. 1992, 17, 105–110. [Google Scholar] [CrossRef]
- Humer, E.; Zebeli, Q. Phytate in feed ingredients and potentials for improving the utilization of phosphorus in ruminant nutrition. Anim. Feed Sci. Technol. 2015, 209, 1–15. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Sun, S.; Cao, X.; Sun, R. Effect of hydrothermal pretreatment on the structural changes of alkaline ethanol lignin from wheat straw. Sci. Rep. 2016, 6, 39354. [Google Scholar] [CrossRef]
- Ojo, M.A. Phytic acid in legumes: A review of nutritional importance and hydrothermal processing effect on underutilised species. Food Res. 2021, 5, 22–28. [Google Scholar] [CrossRef]
- Larsson, M.; Sandberg, A.-S. Phytate Reduction in Oats during Malting. J. Food Sci. 1992, 57, 994–997. [Google Scholar] [CrossRef]
- Lemmens, E.; de Brier, N.; Spiers, K.M.; Ryan, C.; Garrevoet, J.; Falkenberg, G.; Goos, P.; Smolders, E.; Delcour, J.A. The impact of steeping, germination and hydrothermal processing of wheat (Triticum aestivum L.) grains on phytate hydrolysis and the distribution, speciation and bio-accessibility of iron and zinc elements. Food Chem. 2018, 264, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.L.; Papházy, M.J.; Haygarth, P.M.; McKelvie, I.D. Inositol phosphates in the environment. hilos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 449–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergman, E.-L.; Fredlund, K.; Reinikainen, P.; Sandberg, A.-S. Hydrothermal Processing of Barley (cv. Blenheim): Optimisation of Phytate Degradation and Increase of FreeMyo-inositol. J. Cereal Sci. 1999, 29, 261–272. [Google Scholar] [CrossRef]
- March, J.G.; Grases, F.; Salvador, A. Hydrolysis of Phytic Acid by Microwave Treatment: Application to Phytic Acid Analysis in Pharmaceutical Preparations. Microchem. J. 1998, 59, 413–416. [Google Scholar] [CrossRef]
- Hassan, S.; Ahmad, N.; Ahmad, T.; Imran, M.; Xu, C.; Khan, M.K. Microwave processing impact on the phytochemicals of sorghum seeds as food ingredient. J. Food Process. Preserv. 2019, 43, e13924. [Google Scholar] [CrossRef]
- Ali, N.; Sheeba, S.; Gillani, S.Q.; Hamid, M. Production, Purification and Application of Microbial Phytase: An Overview. Int. J. Biol. Biotechnol. 2017, 14, 503–511. [Google Scholar]
- Jupp, A.R.; Beijer, S.; Narain, G.C.; Schipper, W.; Slootweg, J.C. Phosphorus recovery and recycling-closing the loop. Chem. Soc. Rev. 2021, 50, 87–101. [Google Scholar] [CrossRef]
- White, R.E. Fossil Fuel and Food Security. In Fossil Fuel and the Environment; Khan, S., Ed.; InTech: London, UK, 2012; ISBN 978-953-51-0277-9. [Google Scholar]
- Lott, J.N.A.; Kolasa, J.; Batten, G.D.; Campbell, L.C. The critical role of phosphorus in world production of cereal grains and legume seeds. Food Sec. 2011, 3, 451–462. [Google Scholar] [CrossRef]
- Carraresi, L.; Berg, S.; Bröring, S. Emerging value chains within the bioeconomy: Structural changes in the case of phosphate recovery. J. Clean. Prod. 2018, 183, 87–101. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Total P (%) | Phytate-P (%) | (Phytate-P/ Total P) × 100 | Phytase Activity (U/kg) | |
|---|---|---|---|---|
| Cereals | 0.23–0.31 | 0.17–0.23 | 59–78 | 56–5147 |
| Legume seeds | 0.33–0.73 | 0.08–0.33 | 21–56 | 32–258 |
| Oilseeds | 0.6–1.05 | 0.34–0.76 | 57–72 | 73–295 |
| Cereal by-products (bran) | 0.83–1.16 | 0.68–0.88 | 76–82 | 25–7339 |
| Treatment | Process | Substrate | Phytate Reduction | Process/Conditions |
|---|---|---|---|---|
| Mechanical | debranning | cereal grains | up to 90% | removal of outer hull (debranning) [58] |
| layer exclusion | cereal grains | up to 90% | stepwise debranning/specific electrostatic separation; degermination [71,74] | |
| Enzymatic | co-feeding of phytase | cereal grains | 30–50% | phytase supplementation [85] |
| germination | cereal grains | up to 100% (rye) | 2 days pre-steeping, incubation in H2O, 25 °C, 5 days [89] | |
| biotechnological processing | deoiled seeds, bran | up to 90% | 200 to 400 U phytase, 7 volumes H2O, 37 °C, pH 4.5–6 [3] | |
| Chemical | hydrothermal treatment | brown beans | up to 65% | 140 °C, 90 min, H2O [84] |
| acidic hydrolysis | barley | up to 96% | 0.1 M lactic acid soaking, two-step heating at 48 and 50 °C for 5 h and 1 h, respectively [133] | |
| phytic acid solution | up to 99% | 0.44 M HCl, 6 × 2 min microwave heating at 650 W [134] |
| Mechanical & Enzymatic (Option i) | Enzymatic (Option ii) | Mechanical & Chemical (Option iii) | Chemical (Option iv) | |
|---|---|---|---|---|
| Process complexity | − | + | − | o |
| Estimated energy demand | + | o | + | − |
| Harm of regents | + | + | − | − |
| Process maturity | − | o | − | o |
| Free P provision | − | o | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widderich, N.; Mayer, N.; Ruff, A.J.; Reckels, B.; Lohkamp, F.; Visscher, C.; Schwaneberg, U.; Kaltschmitt, M.; Liese, A.; Bubenheim, P. Conditioning of Feed Material Prior to Feeding: Approaches for a Sustainable Phosphorus Utilization. Sustainability 2022, 14, 3998. https://doi.org/10.3390/su14073998
Widderich N, Mayer N, Ruff AJ, Reckels B, Lohkamp F, Visscher C, Schwaneberg U, Kaltschmitt M, Liese A, Bubenheim P. Conditioning of Feed Material Prior to Feeding: Approaches for a Sustainable Phosphorus Utilization. Sustainability. 2022; 14(7):3998. https://doi.org/10.3390/su14073998
Chicago/Turabian StyleWidderich, Niklas, Natalie Mayer, Anna Joelle Ruff, Bernd Reckels, Florian Lohkamp, Christian Visscher, Ulrich Schwaneberg, Martin Kaltschmitt, Andreas Liese, and Paul Bubenheim. 2022. "Conditioning of Feed Material Prior to Feeding: Approaches for a Sustainable Phosphorus Utilization" Sustainability 14, no. 7: 3998. https://doi.org/10.3390/su14073998
APA StyleWidderich, N., Mayer, N., Ruff, A. J., Reckels, B., Lohkamp, F., Visscher, C., Schwaneberg, U., Kaltschmitt, M., Liese, A., & Bubenheim, P. (2022). Conditioning of Feed Material Prior to Feeding: Approaches for a Sustainable Phosphorus Utilization. Sustainability, 14(7), 3998. https://doi.org/10.3390/su14073998