
The Conservation and Restoration of Riparian Forests along Caribbean Riverbanks Using Legume Trees



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Selection and Description
2.2. Seed Collection
2.3. Experimental Conditions of Cultivation
2.4. Seed Mass, Germination, Herbivory and Survival
2.5. Biotechnical Traits of Seedlings
2.6. Statistical Analysis
3. Results
3.1. Germination, Herbivory and Survival
3.2. Biotechnical Traits of Seedlings
4. Discussion
4.1. Species Traits and Strategies
4.2. Which Species, When and Where?
4.3. Using Legume Species in Soil Bioengineering
4.4. Caribbean Legume Tree Species for Restoring and Conserving Riparian Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Environment Program. Becoming #GenerationRestoration: Ecosystem Restoration for People, Nature and Climate. Nairobi, Kenya. 2021, p. 56. Available online: https://wedocs.unep.org/bitstream/handle/20.500.11822/36251/ERPNC.pdf (accessed on 15 January 2022).
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef] [Green Version]
- González, E.; Felipe-Lucia, M.R.; Bourgeois, B.; Boz, B.; Nilsson, C.; Palmer, G.; Sher, A.A. Integrative conservation of riparian zones. Biol. Conserv. 2017, 211, 20–29. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J.A. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feld, C.K.; Birk, S.; Bradley, D.C.; Hering, D.; Kail, J.; Marzin, A.; Melcher, A.; Nemitz, D.; Pedersen, M.L.; Pletterbauer, F.; et al. From natural to degraded rivers and back again: A test ofnrestoration ecology theory and practice. Adv. Ecol. Res. 2011, 44, 119–209. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Okada, K.I.; Mori, A.S. Reconsidering biodiversity hotspots based on the rate of historical land-use change. Biol. Conserv. 2019, 233, 268–275. [Google Scholar] [CrossRef]
- UICN. Liste Rouge de la Flore Vasculaire de Guadeloupe; UICN: Paris, France, 2019. [Google Scholar]
- Rollet, B. Description des espèces. In Arbres des Petites Antilles Tome 2; O.N.F.: Paris, France, 2010. [Google Scholar]
- Rousteau, A.; Portecop, J.; Rolle, B. Carte Écologique de la Guadeloupe; ONF, UAG, PNG, CGG: Jarry, Guadeloupe, 1996. [Google Scholar]
- Gayot, M.; Procopio, L.; Conjard, S.; Boulange, E.; Bernus, J. Étude de la Typologie des Ripisylves de Guadeloupe et Proposition D’espèces Utilisables en Génie Végétal sur les Berges; ONF PNG: Jarry, Guadeloupe, 2018. [Google Scholar]
- Cohen-Shacham, E.; Waters, G.; Janzen, C.; Maginnis, S. (Eds.) Nature Based Solutions to Address Global Societal Challenges; IUCN: Gland, Switzerland, 2016. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.; Hellin, J. Bioengineering for Effective Road Maintenance in the Caribbean; Natural Resources Institute: Chatham, UK, 1996. [Google Scholar]
- Stangl, R.; Hochbichler, E.; Bellos, P.N.; Florineth, F. Allometric estimation of the above-ground biomass components of Alnus incana (L.) Moench used for landslide stabilisation at Bad Goisern (Austria). Plant Soil 2009, 324, 115–129. [Google Scholar] [CrossRef]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Wu, T.H.; McKinnell, W.P.; Swanston, D.N. Strength of tree roots and landslides on Prince of Wales Island, Alaska. Can. Geotech. J. 1979, 16, 19–33. [Google Scholar] [CrossRef]
- Schmidt, K.M.; Roering, J.J.; Stock, J.D.; Dietrich, W.E.; Montgomery, D.R.; Schaub, T. The variability of root cohesion as an influence on shallow landslide susceptibility in the Oregon Coast Range. Can. Geotech. J. 2001, 38, 995–1024. [Google Scholar] [CrossRef]
- Sidle, R.C.; Pearce, A.; O’Loughlin, J.C.L. Hillslope Stability and Land Use; American Geophysical Union: Washington, DC, USA, 1985. [Google Scholar]
- Diaz, J. Control de la Erosión en Zonas Tropicales; Universidad Industrial de Santander, Libreria UIS: Colombia, Bucaramanga, 2001. [Google Scholar]
- Zeh, H. Soil Bioengineering Construction Type Manual; European Federation for Soil Bioengineering: Zürich, Switzerland, 2007. [Google Scholar]
- Lachat, B. Guide de Protection en Techniques Végétales; Ministere de L’amenagement du Territoire et de L’environnement: Paris, France, 1994. [Google Scholar]
- Stokes, A. Selecting tree species for use in rockfall-protection forests. For. Snow Landsc. Res. 2006, 80, 77–86. [Google Scholar]
- De Baets, S.; Poesen, J.; Reubens, B.; Muys, B.; De Baerdemaeker, J.; Meersmans, J. Methodological framework to select plant species for controlling rill and gully erosion: Application to a Mediterranean ecosystem. Earth Surf. Process. Landforms 2009, 34, 1374–1392. [Google Scholar] [CrossRef]
- Ghestem, M.; Cao, K.; Ma, W.; Rowe, N.; Leclerc, R.; Gadenne, C.; Stokes, A. A framework for identifying plant species to be used as ‘ecological engineers’ for fixing soil on unstable slopes. PLoS ONE 2014, 9, e95876. [Google Scholar] [CrossRef]
- Krautzer, E.; Hacker, B. Soil-Bioengineering: Ecological Restoration with Native Plant and Seed Material. In Proceedings of the Conference HBLFA, Raumberg-Gumpenstein, Gumpenstein, Austria, 9 September 2006. HBLFA Raumberg-Gumpenstein. [Google Scholar]
- Li, X.; Zhang, L.; Zhang, Z. Soil bioengineering and the ecological restoration of riverbanks at the Airport Town, Shanghai, China. Ecol. Eng. 2006, 26, 304–314. [Google Scholar] [CrossRef]
- Cavaillé, P.; Ducasse, L.; Breton, V.; Dommanget, F.; Tabacchi, E.; Evette, A. Functional and taxonomic plant diversity for riverbank protection works: Bioengineering techniques close to natural banks and beyond hard engineering. J. Environ. Manag. 2015, 151, 65–75. [Google Scholar] [CrossRef]
- Janssen, P.; Cavaillé, P.; Bray, F.; Evette, A. Soil bioengineering techniques enhance riparian habitat quality and multi-taxonomic diversity in the foothills of the Alps and Jura Mountains. Ecol. Eng. 2019, 133, 1–9. [Google Scholar] [CrossRef]
- Polster, D.F.; Bio, R.P. Soil bioengineering for riparian restoration. In Proceedings of the Canadian Land Reclamation Association 2006 Conference, Ottawa, ON, Canada, 27–31 August 2006. [Google Scholar]
- Tisserant, M.; Janssen, P.; Evette, A.; González, E.; Cavaillé, P.; Poulin, M. Diversity and succession of riparian plant communities along riverbanks bioengineered for erosion control: A case study in the foothills of the Alps and the Jura Mountains. Ecol. Eng. 2020, 152, 105880. [Google Scholar] [CrossRef]
- Lennox, M.S.; Lewis, D.J.; Jackson, R.D.; Harper, J.; Larson, S.; Tate, K.W. Development of vegetation and aquatic habitat in restored riparian sites of California’s north coast rangelands. Restor. Ecol. 2011, 19, 225–233. [Google Scholar] [CrossRef]
- McClain, C.D.; Holl, K.D.; Wood, D.M. Successional models as guides for restoration of riparian forest understory. Restor. Ecol. 2011, 19, 280–289. [Google Scholar] [CrossRef]
- Martin, F.M.; Janssen, P.; Bergès, L.; Dupont, B.; Evette, A. Higher structural connectivity and resistance against invasions of soil bioengineering over hard-engineering for riverbank stabilisation. Wetl. Ecol. Manag. 2021, 29, 27–39. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Meyer, J.L. Effects of Bioengineered Streambank Stabilization on Bank Habitat and Macroinvertebrates in Urban Streams. Environ. Manag. 2006, 38, 218–226. [Google Scholar] [CrossRef]
- Symmank, L.; Natho, S.; Scholz, M.; Schröder, U.; Raupach, K.; Schulz-Zunkel, C. The impact of bioengineering techniques for riverbank protection on ecosystem services of riparian zones. Ecol. Eng. 2020, 158, 106040. [Google Scholar] [CrossRef]
- Popoff, N.; Jaunatre, R.; Le Bouteiller, C.; Paillet, Y.; Favier, G.; Buisson, M.; Meyer, C.; Dedonder, E.; Evette, A. Optimization of restoration techniques: In-situ transplantation experiment of an endangered clonal plant species (Typha minima Hoppe). Ecol. Eng. 2020, 160, 106130. [Google Scholar] [CrossRef]
- Knoepp, D.; Swank, W.T. Rates of nitrogen mineralization across an elevation and vegetation gradient in the southern Appalachians. Plant Soil 1998, 204, 235–241. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Bradley, K.L.; Wedin, D.A. Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecol. Lett. 2002, 5, 454–466. [Google Scholar] [CrossRef] [Green Version]
- Debell, D.S.; Whitesell, C.D.; Schubert, T.H. Mixed Plantations of Eucalyptus and Leguminous Trees; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1985. [Google Scholar]
- Khanna, P.K. Comparison of growth and nutrition of young monocultures and mixed stands of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 1997, 94, 105–113. [Google Scholar] [CrossRef]
- Binkley, D.; Senock, R.; Bird, S.; Cole, T.G. Twenty years of stand development in pure and mixed stands of Eucalyptus saligna and nitrogen-fixing Facaltaria moluccana. For. Ecol. Manag. 2003, 182, 93–102. [Google Scholar] [CrossRef]
- Fisher, R. Amelioration of degraded rain forest soils by plantations of native trees. Soil Sci. Soc. Am. J. 1995, 59, 544–549. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.A.; De Faria, S.M. The contribution of N2-fixing tree legumes to land reclamation and sustainability in the tropics. Soil Biol. Biochem. 1997, 29, 897–903. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Macedo, M.O.; Resende, A.S.; Garcia, P.C.; Boddey, R.M.; Jantalia, C.P.; Urquiag, S.E.; Campello, F.C.; Franco, A.A. Changes in soil C and N stocks and nutrient dynamics 13 years after recovery of degraded land using leguminous nitrogen-fixing trees. For. Ecol. Manag. 2008, 255, 1516–1524. [Google Scholar] [CrossRef]
- Norris, J.E.; Stokes, A.; Mickovski, S.B.; Cammeraat, E.; Van Beek, R.; Nicoll, B.C.; Achim, A. (Eds.) Slope Stability and Erosion Control: Ecotechnological Solutions; Springer Science & Business Media: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Petrone, F.; Preti, A. Soil Bioengineering Measures in Latin America: Authocthonal Cuttings Suitability. In Landslide Science and Practice; Volume 7: Social and Economic Impact and Policies; Claudio, M.P., Canuti, K.S., Eds.; Springer: Berlin, Germany, 2013. [Google Scholar]
- Hostettler, S.; Jöhr, A.; Montes, C.; D’Acunzi, A. Community-based landslide risk reduction: A review of a Red Cross soil bioengineering for resilience program in Honduras. Landslides 2019, 16, 1779–1791. [Google Scholar] [CrossRef] [Green Version]
- Maxwald, M.; Crocetti, C.; Ferrari, R.; Petrone, A.; Rauch, H.P.; Preti, F. Soil and Water Bioengineering Applications in Central and South America: A Transferability Analysis. Sustainability 2020, 12, 10505. [Google Scholar] [CrossRef]
- Petrone, A.; Preti, F. Suitability of soil bioengineering techniques in Central America: A case study in Nicaragua. Hydrol. Earth Syst. Sci. 2008, 12, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Mira, E.; Evette, A.; Labbouz, L.; Robert, M.; Rousteau, A.; Tournebize, R. Investigation of the asexual reproductive characteristics of native species for soil bioengineering in the west indies. J. Trop. For. Sci. 2021, 33, 333–342. [Google Scholar] [CrossRef]
- Saur, E.; Carcelle, S.; Guezennec, S.; Rousteau, A. Nodulation of legume species in wetlands of Guadeloupe (Lesser antilles). Wetlands 2000, 20, 730–734. [Google Scholar] [CrossRef]
- Fournet, J. Flore Illustrée des Phanérogames de Guadeloupe et de Martinique, Nouvelle Édition Revue et Augmentée; INRA: Paris, France, 1978. [Google Scholar]
- Acevedo, P.; Strong, M. Catalogue of Seed Plants of the West Indies. Smithson. Contrib. Bot. 2012, 98, 1–1192. [Google Scholar] [CrossRef]
- Tropicos v3. Missouri Botanical Garden. Available online: https://tropicos.org (accessed on 13 January 2022).
- Válio, I.F.; Scarpa, F.M. Germination of seeds of tropical pioneer species under controlled and natural conditions. Braz. J. Bot. 2001, 24, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Sautu, A.; Baskin, J.M.; Baskin, C.C.; Condit, R. Studies on the seed biology of 100 native species of trees in a seasonal moist tropical forest, Panama, Central America. For. Ecol. Manag. 2006, 234, 245–263. [Google Scholar] [CrossRef]
- Swaine, M.D.; Whitmore, T.C. On the definition of ecological species groups in tropical rain forests. Vegetatio 1988, 75, 81–86. [Google Scholar] [CrossRef]
- Laime, E.M.O.; Alves, E.U.; Guedes, R.S.; Silva, K.B.; de Souza Oliveira, D.C.; da Silva Santos, S. Emergency and initial growth of seedlings of Inga ingoides (Rich.) Willd. in function of the position and depth of sowing. Semina: Ciências Agrárias 2010, 31, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Palomino, J.; Barra, M. Especies forestales nativas con potencial para reforestación en la provincia de oxapampa y fichas técnicas de las especies de mayor prioridad. PRO Nat. 2003, 109. Available online: http://www.infobosques.com/descargas/biblioteca/70.pdf (accessed on 13 January 2022).
- Leão, J.R.; Lima, J.P.D.C.; Pinto, S.D.N.; de Paiva, A.V. Germinação de sementes e crescimento inicial de plântulas de ingá-mirim. Revsista Bras. Arborização Urbana 2012, 7, 11–19. [Google Scholar] [CrossRef]
- Mendes-Rodrigues, C.; Ferreira, W.R.; De Lima, J.A.; Dornelles, M.C.; Ranal, M.a.; De Santana, D.G. Germinação de embriões de duas espécies de Inga (Mimosaceae). Rev. Bras. Biociências 2007, 5, 561–563. [Google Scholar]
- Augspurger, C.K. Light Requirements of Neotropical Tree Seedlings: A Comparative Study of Growth and Survival. J. Ecol. 1984, 72, 777. [Google Scholar] [CrossRef]
- Poorter, L.; Markesteijn, L. Seedling Traits Determine Drought Tolerance of Tropical Tree Species. Biotropica 2008, 40, 321–331. [Google Scholar] [CrossRef]
- McLaren, K.P.; McDonald, M.A. The effects of moisture and shade on seed germination and seedling survival in a tropical dry forest in Jamaica. For. Ecol. Manag. 2003, 183, 61–75. [Google Scholar] [CrossRef]
- Queenborough, S.A.; Burslem, D.F.R.P.; Garwood, N.C.; Valencia, R. Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival. Ecology 2007, 88, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, B.M.; Kursar, T.A.; Tyree, M.T. Drought effects on seedling survival in a tropical moist forest. Trees 2005, 19, 312–321. [Google Scholar] [CrossRef]
- Schemske, D.W.; Mittelbach, G.G.; Cornell, H.V.; Sobel, J.M.; Roy, K. Is There a Latitudinal Gradient in the Importance of Biotic Interactions? Annu. Rev. Ecol. Evol. Syst. 2009, 40, 245–269. [Google Scholar] [CrossRef] [Green Version]
- Coley, P.D.; Barone, J.A. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 1996, 27, 305–335. [Google Scholar] [CrossRef]
- Coste, S.; Roggy, J.C.; Imbert, P.; Born, C.; Bonal, D.; Dreyer, E. Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf nitrogen concentration and shade tolerance. Tree Physiol. 2005, 25, 1127–1137. [Google Scholar] [CrossRef] [Green Version]
- Michalet, S.; Rohr, J.; Warshan, D.; Bardon, C.; Roggy, J.-C.; Domenach, A.M.; Czarnes, S.; Pommier, T.; Combourieu, B.; Guillaumaud, N.; et al. Phytochemical analysis of mature tree root exudates in situ and their role in shaping soil microbial communities in relation to tree N-acquisition strategy. Plant Physiol. Biochem. 2013, 72, 169–177. [Google Scholar] [CrossRef]
- Janzen, D.H. The defenses of legumes against herbivores. In Advances in Legume Systematics; Polhill, R.M., Raven, P.R., Eds.; Academic Press: London, UK, 1981; Available online: http://hdl.handle.net/11606/1296. (accessed on 13 January 2022).
- Macedo, M.L.R.; Garcia, V.A.; Maria das Graças, M.F.; Richardson, M. Characterization of a Kunitz trypsin inhibitor with a single disulfide bridge from seeds of Inga laurina (SW.) Willd. Phytochemistry 2007, 68, 1104–1111. [Google Scholar] [CrossRef]
- Janzen, D.H.; Lynn, D.G.; Fellowst, L.E.; Hallwachs, W. The indole alkaloid, hypaphorine and Pterocarpus seed protection. Phytochemistry 1982, 21, 1035–1037. [Google Scholar] [CrossRef]
- Nasto, M.K.; Winter, K.; Turner, B.L.; Cleveland, C.C. Nutrient acquisition strategies augment growth in tropical N2-fixing trees in nutrient-poor soil and under elevated CO2. Ecology 2019, 100, e02646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought-and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Boonman, C.C.; van Langevelde, F.; Oliveras, I.; Couédon, J.; Luijken, N.; Martini, D.; Veenendaal, E.M. On the importance of root traits in seedlings of tropical tree species. New Phytol. 2020, 227, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Veneklaas, E.L.; Poorter, L. Growth and carbon partitioning of tropical tree seedlings in contrasting light environments. In Inherent Variation in Plant Growth. Physiological Mechanisms and Ecological Consequences; Van Vuuren, M.M.I., Ed.; Leiden, The Netherlands, 1998; pp. 337–361. Available online: https://agris.fao.org/agris-search/search.do?recordID=NL2012018952 (accessed on 13 January 2022).
- Smith-Martin, C.M.; Gei, M.G.; Bergstrom, E.; Becklund, K.K.; Becknell, J.M.; Waring, B.G.; Werden, L.K.; Powers, J.S. Effects of soil type and light on height growth, biomass partitioning, and nitrogen dynamics on 22 species of tropical dry forest tree seedlings: Comparisons between legumes and nonlegumes. Am. J. Bot. 2017, 104, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunel, C.; Stahl, C.; Heuret, P.; Nicolini, E.; Baraloto, C. Disentangling the effects of environment and ontogeny on tree functional dimensions for congeneric species in tropical forests. New Phytol. 2020, 226, 385–395. [Google Scholar] [CrossRef]
- McConnaughay, K.D.M.; Coleman, J.S. Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients. Ecology 1999, 80, 2581–2593. [Google Scholar] [CrossRef]
- Imbert, D.; Dulormne, M. Restauration du Couvert Forestier dans les ESPACES inondables D’arrière-Mangrove: Comment Faciliter le Retour de la Forêt à Pterocarpus Officinalis; UAG: Pointe à Pitre, France, 2013. [Google Scholar]
- Green, P.T.; Juniper, P.A. Seed–seedling allometry in tropical rain forest trees: Seed mass-related patterns of resource allocation and the ‘reserve effect’. J. Ecol. 2004, 92, 397–408. [Google Scholar] [CrossRef]
- Acevedo-Rodríguez, P.; Strong, M.T. Floristic Richness and Affinities in the West Indies. Bot. Rev. 2008, 74, 5–36. [Google Scholar] [CrossRef]
- Silva, C.C. Conservação de Sementes de Inga laurina (Sw.) Willd. 2016. Available online: https://repositorio.ufpb.br/jspui/handle/123456789/12085 (accessed on 13 January 2022).
- Barrozo, L.M.; Ursulino, E.A.; de Araujo, L.R.; Sena, D.D.A.; de Medeiros, D.S.; dos Santos, J.C. Quality seeds Inga physiological function of drying. Biosci. J. 2014, 30, 645–654. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier: New York, NY, USA, 1998. [Google Scholar]
- Cruz, E.D.; Martins, F.D.O.; Carvalho, J.E.U.D. Biometria de frutos e sementes e germinação de jatobá-curuba (Hymenaea intermedia Ducke, Leguminosae—Caesalpinioideae). Rev. Bras. Botânica 2001, 24, 161–165. [Google Scholar] [CrossRef]
- Muller., F. Diversité Génétique, Adaptation de Pterocarpus Officinalis Jacq. et de ses Symbiotes dans des Forêts Marécageuses de la Région Caraïbe. Ph.D. Thesis, Université des Antilles la Guyane, Pointe à Pitre, France, 2006. [Google Scholar]
- Gao, J.; Wang, F.; Gao, Y.; Stangl, R. Root architecture characteristics of plant inlay in live slope grating. For. Stud. China 2007, 9, 177–181. [Google Scholar] [CrossRef]
- Shah, B.H. Field Guide on Soil Bioengineering for Slope Stabilization in Timor-Leste; Exch. South–South Exp. TF011068, Trust Fund; World Bank Office: Dili, Timor-Leste, 2012. [Google Scholar]
- Stangl, R. Hedge brush layers and live crib walls—stand development and benefits. In Eco-and Ground Bio-Engineering: The Use of Vegetation to Improve Slope Stability. Developments in Plant and Soil Sciences; Stokes, A., Spanos, I., Norris, J.E., Cammeraat, E., Eds.; Springer: Dordrecht, The Netherland, 2007; Volume 103. [Google Scholar] [CrossRef]
- Dhital, Y.P.; Kayastha, R.B.; Shi, J. Soil bioengineering application and practices in Nepal. Environ. Manag. 2013, 51, 354–364. [Google Scholar] [CrossRef]
- IGN Diagnostic des Forêts de la Guadeloupe. CD971-; IGN-ONF Deal Guadeloupe: Basse-Terre, Guadeloupe, 2015. [Google Scholar]
- Chaer, G.M.; Resende, A.S.; Campello, E.F.C.; de Faria, S.M.; Boddey, R.M. Nitrogen-fixing legume tree species for the reclamation of severely degraded lands in Brazil. Tree Physiol. 2011, 31, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.M.; Curcio, G.R.; Bonnet, A.; Uhlmann, A.; Palma, V.H. Initial growth of native tree species in a degraded soil with presence of plinthite in Biome Cerrado, Brasília-DF. Nativa: Pesquisas Agrárias e Ambientais 2018, 6, 787–794. [Google Scholar] [CrossRef]
- Cruz-Neto, O.; Machado, I.C.; Duarte, J.A.; Lopes, A.V. Synchronous phenology of hawkmoths (Sphingidae) and Inga species (Fabaceae–Mimosoideae): Implications for the restoration of the Atlantic forest of northeastern Brazil. Biodiv. Conserv. 2011, 20, 751–765. [Google Scholar] [CrossRef]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 1866–1878. [Google Scholar] [CrossRef]
- Campbell, J.D.; Taylor, M.A.; Stephenson, T.S.; Watson, R.A.; Whyte, F.S. Future climate of the Caribbean from a regional climate model. Int. J. Climatol. 2011, 31, 1866–1878. [Google Scholar] [CrossRef]
- Lowry, J.B. Effect of drought on Mount Kinabalu. Malayan Nat. J. 1973, 26, 178–179. [Google Scholar]
- Werner, W.L. Canopy dieback in the upper montane rain forests of Sri Lanka. GeoJournal 1988, 17, 245–248. [Google Scholar] [CrossRef]
- Pounds, J.A.; Fogden, M.P.L.; Campbell, J.H. Biological response to climate change on a tropical mountain. Nature 1999, 398, 611–615. [Google Scholar] [CrossRef]
- Joseph, P. La végétation des Petites Antilles: Principaux traits floristiques et effets plausibles du changement climatique. VertigO-la Rev. Électronique en Sci. L’environnement 2011, 11, 1. [Google Scholar] [CrossRef]
- Ste-Marie, C.; Nelson, E.A.; Dabros, A.; Bonneau, M.E. Assisted migration: Introduction to a multifaceted concept. For. Chron. 2011, 87, 724–730. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, J.S.; Hellmann, J.J.; Schwartz, M.W. A framework for debate of assisted migration in an era of climate change. Conserv. Biol. 2007, 21, 297–302. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Months | Description | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | J | F | M | A | M | J | J | A | S | O | N | D | Ecosystem | Biogéography |
| Inga ingoides (Rich.) Willd. | Seasonal evergreen forest/rainforest | Native to the Lesser Antilles (Dominica, Guadeloupe, Marie Galante, Martinique, St. Lucia, St. Vincent), Margarita, Trinidad, and South America (Bolivia, Brazil, Colombia, French Guiana, Guyana, Peru, Suriname, Venezuela). | ||||||||||||
| Inga laurina (Sw.) Willd. | Seasonal evergreen forest/rainforest | Native to Cuba, Hispaniola, Puerto Rico, Virgin Islands (St. Croix, St. John, St. Thomas, Tortola, Virgin Gorda), the Lesser Antilles (Antigua, Barbados, Dominica, Grenada, Guadeloupe, Marie Galante, Martinique, Montserrat, Saba, St. Kitts, St. Lucia, St. Vincent), Trinidad, Mexico, Central America and South America (Argentina, Belize, Bolivia, Brazil, Colombia, Costa Rica, Ecuador, El Salvador, French Guiana, Guatemala, Honduras, Mexico, Nicaragua, Panama, Paraguay, Peru, Suriname, Venezuela). | ||||||||||||
| Lonchocarpus heptaphyllus (Poir.) DC. | Seasonal evergreen forest | Native to Cuba, Hispaniola, Jamaica, Puerto Rico, the Lesser Antilles (Guadeloupe, Martinique, St. Kitts, St. Lucia), Margarita, Trinidad (cultivated), Central America and South America (Costa Rica, Ecuador, Guyana, French Guiana, Mexico, Panama, Suriname, Venezuela). | ||||||||||||
| Lonchocarpus roseus (Mill.) DC. | Riparian seasonal evergreen forest | Native to the Lesser Antilles (Guadeloupe, Martinique), Hispaniola and Puerto Rico. | ||||||||||||
| Pterocarpus officinalis (Jacq.) | Swamp forest | Native to Cuba, Hispaniola, Jamaica, Puerto Rico, the Lesser Antilles (Dominica, Grenada, Guadeloupe, Marie Galante, Martinique, St. Lucia, St. Vincent), Trinidad, Mexico, Central America and South America (Belize, Brazil, Colombia, Costa Rica, Ecuador, French Guiana, Guatemala, Guyana, Honduras, Mexico, Nicaragua, Panama, Peru, Suriname, Venezuela). | ||||||||||||
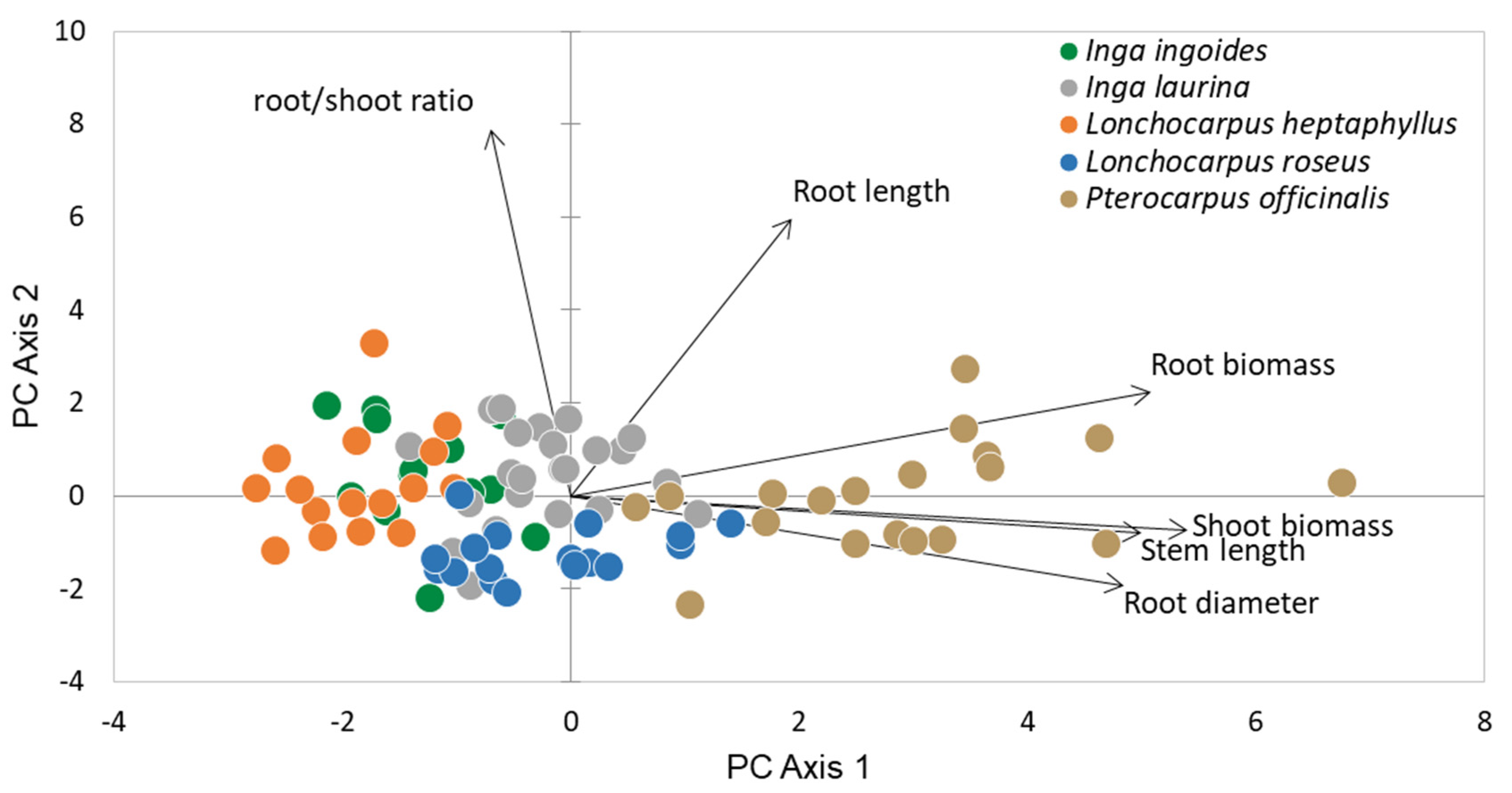
| PCA1 (57%) | PCA2 (22%) | PCA3 (12%) | |
|---|---|---|---|
| Shoot biomass | 0.521 | −0.071 | −0.085 |
| Root biomass | 0.490 | 0.214 | −0.312 |
| Stem length | 0.482 | −0.076 | 0.197 |
| Root diameter | 0.466 | −0.186 | −0.159 |
| Root length | 0.187 | 0.574 | 0.756 |
| Root/Shoot | −0.068 | 0.761 | −0.509 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mira, E.; Rousteau, A.; Tournebize, R.; Labbouz, L.; Robert, M.; Evette, A. The Conservation and Restoration of Riparian Forests along Caribbean Riverbanks Using Legume Trees. Sustainability 2022, 14, 3709. https://doi.org/10.3390/su14073709
Mira E, Rousteau A, Tournebize R, Labbouz L, Robert M, Evette A. The Conservation and Restoration of Riparian Forests along Caribbean Riverbanks Using Legume Trees. Sustainability. 2022; 14(7):3709. https://doi.org/10.3390/su14073709
Chicago/Turabian StyleMira, Eléonore, Alain Rousteau, Régis Tournebize, Lucie Labbouz, Marie Robert, and André Evette. 2022. "The Conservation and Restoration of Riparian Forests along Caribbean Riverbanks Using Legume Trees" Sustainability 14, no. 7: 3709. https://doi.org/10.3390/su14073709
APA StyleMira, E., Rousteau, A., Tournebize, R., Labbouz, L., Robert, M., & Evette, A. (2022). The Conservation and Restoration of Riparian Forests along Caribbean Riverbanks Using Legume Trees. Sustainability, 14(7), 3709. https://doi.org/10.3390/su14073709