
Geometric and Topological Bases of a New Classification of Wood Vascular Tissues, Part 2: Classification of Vessels According to Their Grouping



Abstract
:1. Introduction
State of the Arts
2. Methods and Materials
2.1. Classification of Vessels into Groups According to Their Grouping
- Vessel distribution coefficient (A), which relates the diameter and distance of the examined cells to the density of the vessels.
- Coefficient of influence of the site (B), which connects the diameter and distance of the studied cells with the total area of the vessels.
- Coefficient of concentration (C), which represents the ratio of the distance between the two cells tested (µ1) to the average diameter of these cells ().
- Coefficient of diffusion (D), which represents the ratio of the average diameter between two measured cells () to the distance between them (µ1).
2.2. Solitary Arrangement of Vessels
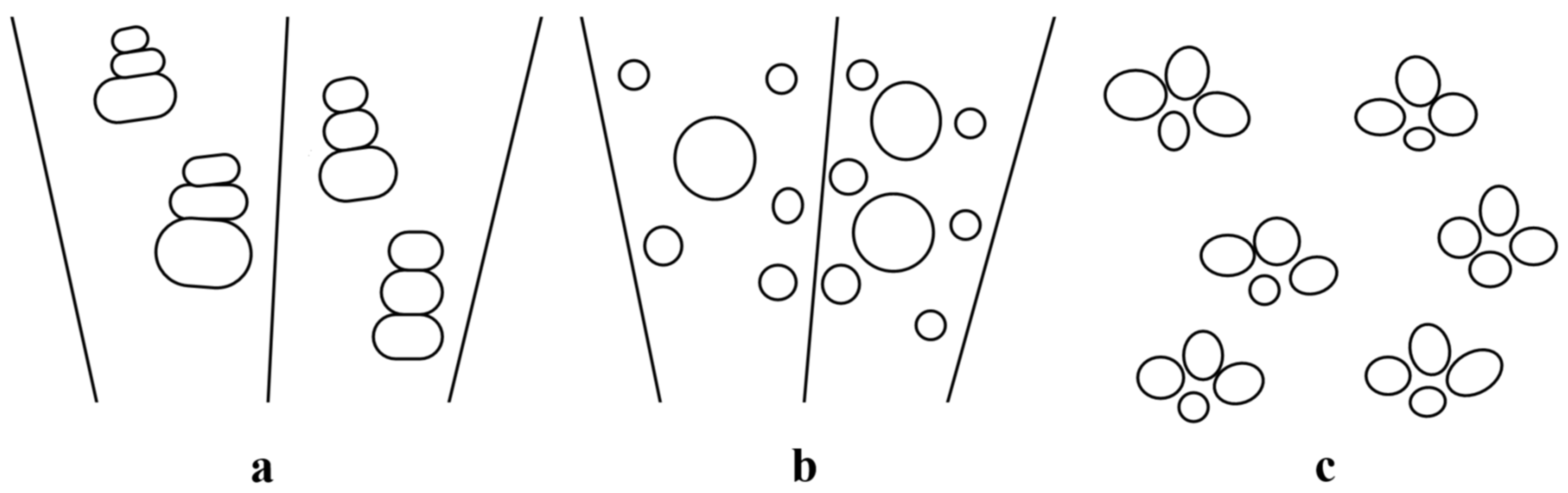
2.3. Arrangement of Vessels in Clusters and Tangential Groups
2.4. Radial and Diagonal Grouping of Vessels
2.5. Heterogeneous Groups (Mixed Arrangement of Large and Small Vessels)
3. Results and Discussion
3.1. Quantitative Definition of Homogeneous and Heterogeneous Anatomical Structures
3.2. Classification of Species with Solitary Vessels
3.3. Classification of Species with Clusters and Tangential Groups
3.4. Classification of Species with Radial and Diagonal Grouping of Vessels
3.5. Classification of Species with Heterogeneous Groups of Vessels
3.6. Classification of Species with Diffuse Groups of Vessels
3.7. Classification of Species with Marginal Groups of Vessels
4. Classification of Groups by Quantitative Indicators
4.1. Classification by Degrees of Homogeneity/Regularity
4.2. Usage of Quantitative Indicators to Identify Wood
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wagenführ, R. Anatomy of Wood; VEB Fachbuchverlag: Leipzig, Germany, 1984; pp. 78–173. [Google Scholar]
- Ugolev, B.N. Wood Science; ACADEMIA: Moscow, Russia, 2006; pp. 65–83. [Google Scholar]
- Enchev, E. Wood Science, Textbook; Publishing House of Zemizdat: Sofia, Bulgaria, 1984; pp. 28–54. [Google Scholar]
- Baas, P. Terminology of imperforate tracheary elements in defence of ubriform fibres with minutely bordered pits. IAWA Bull. 1986, 7, 82–86. [Google Scholar] [CrossRef]
- Wagenführ, R.; Scheiber, C. Atlas of Wood; Springer: Berlin, Germany, 1996; pp. 42–56. [Google Scholar]
- Hafemann, L.G.; Oliveira, L.S.; Cavalin, P. Forest species recognition using deep convolutional neural networks. In Proceedings of the 2014 22nd International Conference on Pattern Recognition, Stockholm, Sweden, 24–28 August 2014; pp. 1103–1107. [Google Scholar]
- Kwon, O.; Lee, H.G.; Lee, M.-R.; Jang, S.; Yang, S.-Y.; Park, S.-Y.; Choi, I.-G.; Yeo, H. Automatic Wood Species Identification of Korean Softwood Based on Convolutional Neural Networks. J. Korean Wood Sci. Technol. 2017, 45, 797–808. [Google Scholar] [CrossRef]
- Souza, D.V.; Santos, J.X.; Vieira, H.C.; Naide, T.L.; Nisgoski, S.; Oliveira, L.E.S. An automatic recognition system of Brazilian flora species based on textural features of macroscopic images of wood. Wood Sci. Technol. 2020, 54, 1065–1090. [Google Scholar] [CrossRef]
- da Silva, N.R.; De Ridder, M.; Baetens, J.M.; Van den Bulcke, J.; Rousseau, M.; Bruno, O.M.; Beeckman, H.; De Baets, B. Automated classification of wood transverse cross-section micro-imagery from 77 commercial Central-African timber species. Ann. For. Sci. 2017, 74, 30. [Google Scholar] [CrossRef]
- de Andrade, B.G.; Basso, V.M.; de Figueiredo Latorraca, J.V. Machine vision for field-level wood identification. IAWA J. 2020, 41, 681–698. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, C.K. Hardwood Species Classification with Hyperspectral Microscopic Images. J. Spectrosc. 2019, 2019, 2039453. [Google Scholar] [CrossRef]
- Ravindran, P.; Owens, F.C.; Wade, A.C.; Vega, P.; Muro, R.M.; Shmulsky, R.; Wiedenhoeft, A.C. Field-deployable computer vision wood identification of Peruvian timbers. Front. Plant Sci. 2021, 12, 940. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Jiao, L.; Wiedenhoeft, A.C.; Yin, Y. Machine learning approaches outperform distance-and tree-based methods for DNA barcoding of Pterocarpus wood. Planta 2019, 249, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Richter, H.G.; Dallwitz, M.J. Commercial Timbers: Descriptions, Illustrations, Identification, and Information Retrieval. Available online: https://www.delta-intkey.com/wood/index.htm (accessed on 10 December 2021).
- Benkova, V.E.; Schweingruber, F.H. Anatomy of Russian Woods; Haupt Verlag: Bern, Austria, 2004; pp. 17–48. [Google Scholar]
- Schweingruber, F.H.; Crivellaro, A. Stem Anatomical Features of Dicotyledons. Xylem, Phloem, Cortex and Periderm Characteristics for Ecological and Taxonomical Analysis; Dr. Kessel: Remagen, Germany, 2015; p. 37. Available online: https://www.forstbuch.de/wp-content/uploads/2018/04/SchweingruberCrivellaroWEB.pdf (accessed on 10 December 2021).
- Schweingruber, F.H.; Börner, A. The Plant Stem. A Microscopic Aspect; Springer Nature: Cham, Switzerland, 2018; pp. 23–65. [Google Scholar]
- Alfonso, V.A.; Baas, P.; Carlquist, S.; Chimelo, J.P.; Rauber Coradin, V.T.; Détienne, P.; Gasson, P.; Grosser, D.; Ilic, J.; Kuroda, K.; et al. IAWA list of microscopic features for hardwood identification, with an Appendix on non-anatomical information. IAWA Bull. 1989, 10, 219–332. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Database | Features Extraction | Classification |
|---|---|---|---|
| Hafemann et al. [6] | 2 Brazilian forest species datasets | No preliminary feature extraction. | CNN |
| Kwon et al. [7] | 5 Korean softwood species | No preliminary feature extraction. | CNN |
| da Silva [9] | 77 commercial timber species from the Democratic Republic of Congo | LPQ and LBP | LDA and PCA |
| Souza et al. [8] | 1901 macroscopic images of wood from 46 Brazilian species | Concatenation of rotation-invariant LBP histograms | SVM |
| de Andrade [10] | 21 species, 2000 macroscopic images | Gray level co-occurrence matrix | SVM |
| Zhao and Wang [11] | 6 hardwood species were studied, where 1440 wood samples are prepared | Mathematical morphological operation and K-L divergence | PCA, KPCA, and MDS, and the three classifiers BP neural network, SVM, and MD are combined for the classification task |
| 1 | European ash Fraxinus excelsior L. | 39 | Makore Tieghemella heckelii (A.Chev.) |
| 2 | White mulberry Morus alba L. | 40 | Ako Antiaris africana Engl. |
| 3 | Black locust Robinia pseudoacacia L. | 41 | Iroko Chlorophora excelsa (Welw.) C.C.Berg |
| 4 | Honey locust Gleditsia triacanthos L. | 42 | Dibetu Lovoa; trichilioides Harms. |
| 5 | Field elm Ulmus campestris Mill. | 43 | Koto Pterygota macrocarpa K.Schum. |
| 6 | Common (European) oak Quercus robur L. | 44 | Eyong Sterculia oblonga Mast. |
| 7 | Sweet chestnut Castanea sativa Mill. | 45 | Avodire Turraeanthus africana Welw. ex C. DC. |
| 8 | Common beech Fagus sylvatica L. | 46 | Emien Alstonia congensis De Wild. |
| 9 | Sycamore Acer pseudoplatanus L. | 47 | Ramin Gonystylus bancanus (Miq.) Kurz. |
| 10 | Black alder Alnus glutinosa (L.) Britton | 48 | Balsa Ochroma lagopus Cav. ex Lam. |
| 11 | Common hornbeam Carpinus betulus L. | 49 | Ilomba Pycnanthus angolensis (Welw.) Warb. |
| 12 | Common hazel Corylus avellana L. | 50 | Limba Terminalia superba Engl. & Diels |
| 13 | Common aspen Populus tremula L. | 51 | Obeche Triplochiton scleroxylon K.Schum. |
| 14 | White willow Salix alba L. | 52 | Duse Afzelia bipindensis Harms |
| 15 | Small-leaved linden Tilia cordata Mill. | 53 | Mukulungu Autranella congolensis (De Wild.) A.Chev. |
| 16 | Horse chestnut Aesculus hippocastanum L. | 54 | Pernambouc Caesalpinia echinata Lam |
| 17 | Silver birch Betula pendula (Regel) | 55 | Brazilian rosewood Dalbergia nigra Allem. ex Benth., |
| 18 | Common pear Pyrus communis L. | 56 | Macassar Ebony Diospyros celebica Bakh. |
| 19 | Rowan Sorbus aucuparia L. | 57 | Lignum vitae Guaiacum officinale L. |
| 20 | Wild service tree Sorbus torminalis (L.) Crantz | 58 | Bubinga Guibourtia tessmannii (Harms) J.Leonard |
| 21 | Persian silk tree Albizia julibrissin Durazz. | 59 | Mansonia Mansonia altissima A.Chev. |
| 22 | Pecan (Hickory) Carya pecan (Marshall) Nutt. | 60 | Wenge Millettia laurentii De Wild. |
| 23 | Persian walnut Juglans regia L. | 61 | Wild cashew Anacardium excelsum (Bertero & Balb. ex Kunth) Skeels |
| 24 | Empress tree Paulownia tomentosa (Thunb.) Steud. | 62 | Verawood Bulnesia arborea (Jacq.) Engl. |
| 25 | Teac Tectona grandis L.f. | 63 | Guanandi Calophyllum brasiliense Cambess. |
| 26 | Wild cherry Prunus avium L. | 64 | Lemonwood Calycophyllum multiflorum Griseb. |
| 27 | European buckthorn Rhamnus cathartica L. | 65 | Faro Daniellia ogea (Harms) Rolfe ex Holland |
| 28 | Okoumé Aucoumea klaineana Pierre | 66 | Indian laurel Terminalia elliptica Willd. |
| 29 | Sapelli Entandrophragma cylindricum Sprague | 67 | Angelim Hymenolobium petraeum Ducke |
| 30 | Sipo Entandrophragma utile Sprague | 68 | Silky Oak Grevillea robusta A.Cunn. ex R.Br. |
| 31 | Karri Eucalyptus diversicolor F.Muell. | 69 | Boa Gambeya beguei Aubrév. & Pellegr. |
| 32 | Kotibe Nesogordonia papaverifera (A.Chev.) Capuron ex N.Hallé | 70 | Slender Tea-tree Brevia leptospermum L. |
| 33 | African mahogany Khaya ivorensis A.Chev. | 71 | Alcornoque (Sapupira) Bowdichia sp (Tul.) Ducke |
| 34 | Honduras mahogany Swietenia macrophylla King | 72 | Afzelia Afzelia africana Sm. ex Pers. |
| 35 | Krabac (Mersava) Anisoptera cochinchinensis Pierre | 73 | Honey Bee Tree Koompassia excelsa (Becc.) Taub. |
| 36 | Dabema Piptadeniastrum africanum (Hook.f.) Brenan | 74 | Framire Terminalia ivorensis A.Chev. |
| 37 | African padauk Pterocarpus soyauxii Taub. | 75 | Red lauan (Nemesu) Shorea pauciflora King |
| 38 | Dark-red meranti Shorea polysperma (Blanco) Merr. | 76 | Ocotea Ocotea puberula (Rich.) Nees |
| Degree of Homogeneity/Regularity | Indicator (Value Classes; Var. Coefficient; Number of Species; Share) | |||||
|---|---|---|---|---|---|---|
| Diameter d, µm | Distances v, µm | Indicator A | Indicator B | Indicator C | Indicator D | |
| Homogeneous and regular | 2; <15%; 34; 44.7% | <10; <30%; 22; 28.9% | <3; <60%; 28; 36.8% | <3; <30%; 37; 48.7% | <5; <40%; 18; 23.7% | <7; <30%; 13; 17.1% |
| Moderately inhomogeneous and irregular | 3–4; 16–24%; 29; 38.2% | 10–16; 31–49%; 33; 43.4% | 3–6; 61–79%; 23; 30.3% | 3–6; 31–39%; 23; 30.3% | 5–6; 41–49%; 31; 40.8% | 7–9; 31–69%; 39; 51.3% |
| Highly inhomogeneous and irregular | >4; >25% 13; 17.1% | >16; >50% 21; 27.6% | >6; >80% 25; 32.9% | >6; >40% 16; 21.1% | >6; >50% 27; 35.5% | >9; >70% 24; 31.6% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardarov, N.; Christoff, N.; Todorov, V. Geometric and Topological Bases of a New Classification of Wood Vascular Tissues, Part 2: Classification of Vessels According to Their Grouping. Sustainability 2022, 14, 2031. https://doi.org/10.3390/su14042031
Bardarov N, Christoff N, Todorov V. Geometric and Topological Bases of a New Classification of Wood Vascular Tissues, Part 2: Classification of Vessels According to Their Grouping. Sustainability. 2022; 14(4):2031. https://doi.org/10.3390/su14042031
Chicago/Turabian StyleBardarov, Nikolai, Nicole Christoff, and Vladislav Todorov. 2022. "Geometric and Topological Bases of a New Classification of Wood Vascular Tissues, Part 2: Classification of Vessels According to Their Grouping" Sustainability 14, no. 4: 2031. https://doi.org/10.3390/su14042031
APA StyleBardarov, N., Christoff, N., & Todorov, V. (2022). Geometric and Topological Bases of a New Classification of Wood Vascular Tissues, Part 2: Classification of Vessels According to Their Grouping. Sustainability, 14(4), 2031. https://doi.org/10.3390/su14042031