
Plant Nutrition under Climate Change and Soil Carbon Sequestration

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Plant Nutrition under Climate Change
2.1. The Effect of CO2 Elevation
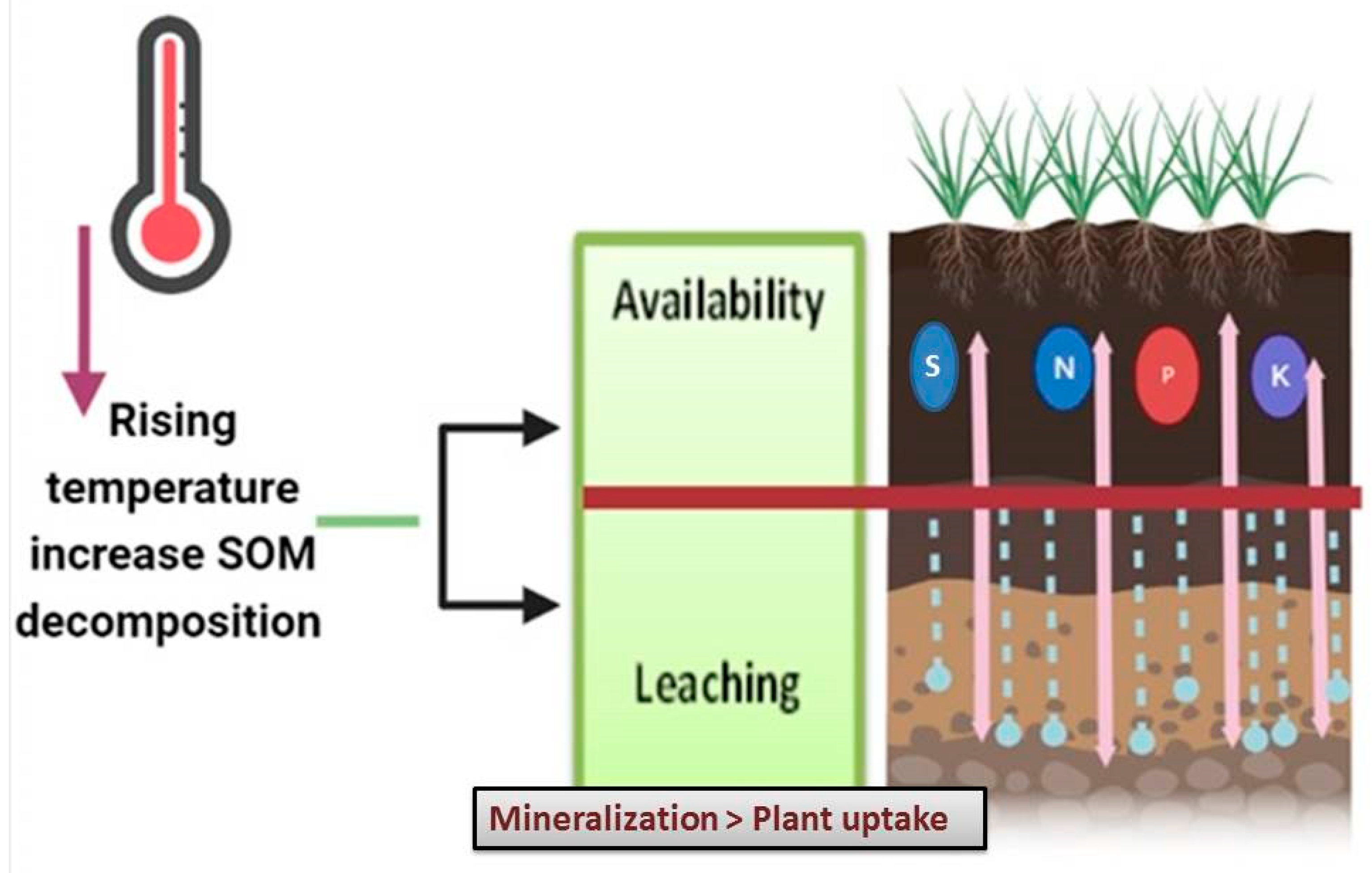
2.2. The Effect of Temperature
2.3. The Effect of Precipitation
3. Soil Carbon Sequestration (SCseq) and Plant Nutrition
3.1. Carbon Storage in the Soil
3.2. Soil Carbon Sequestration
3.3. Relationship of Soil Organic Carbon Sequestration with Other Soil Essential Elements
3.4. Soil C Accumulation or Depletion
4. Conclusions
- Since most studies focused only on the climatic variables, a better understanding of the combination of effects on the availability and uptake of plant nutrients is critically needed.
- In addition, studies on specific site conditions and the combined effects of soil properties with climatic factors are needed in the future.
- The effects of climatic factors on the biofortification of some important elements in plants that are needed to treat deficiencies in humans should be considered future studies.
- Moreover, studies on the relationship between plant nutrient availability and food security under sever climatic conditions should be undertaken.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lotze-Campe, H. Climate Change, Population Growth, and Crop Production: An Overview. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Hossain, M.S.; Arshad, M.; Qian, L.; Zhao, M.; Mehmood, Y.; Kächele, H. Economic impact of climate change on crop farming in Bangladesh: An application of Ricardian method. Ecol. Econ. 2019, 164, 106354. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Elbanna, B.A.; Al-Najoli, E.; Alsherief, A.; Negm, S.; El-Nour, E.A.; Nofal, A.; Sharabash, S. Agricultural Waste Management for Climate Change Mitigation: Some Implications to Egypt. In Waste Management in MENA Regions, Springer Water Book Series; Negm, A.M., Shareef, N., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Elbehiry, F.; Mahmoud, M.A.; Negm, A. Land Use in Egypt’s Coastal Lakes: Opportunities and Challenges. In Egyptian Coastal Lakes and Wetlands: Part I—Characteristics and Hydrodynamics; Negm, A.M., Bek, M.A., Abdel-Fattah, S., Eds.; Springer International Publishing AG: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Shukla, S.K.; Shee, S.; Maity, S.K.; Solomon, S.; Awasthi, S.K.; Gaur, A.; Pathak, A.D.; Jaiswal, V.P. Soil Carbon Sequestration and Crop Yields in Rice–Wheat and Sugarcane–Ratoon–Wheat Cropping Systems Through Crop Residue Management and Inoculation of Trichoderma viride in Subtropical India. Sugar Tech 2016, 19, 347–358. [Google Scholar] [CrossRef]
- Egli, M.; Bösiger, M.; Lamorski, K.; Sławiński, C.; Plötze, M.; Wiesenberg, G.L.; Tikhomirov, D.; Musso, A.; Hsu, S.-Y.; Raimondi, S. Pedogenesis and carbon sequestration in transformed agricultural soils of Sicily. Geoderma 2021, 402, 115355. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Elbehiry, F. Soil Carbon Sequestration for Climate Change Mitigation: Some Implications to Egypt. In Climate Change Impacts on Agriculture and Food Security in Egypt; Springer: Cham, Switzerland, 2020; pp. 151–181. [Google Scholar] [CrossRef]
- Rajput, V.D.; Chen, Y.; Ayup, M. Effects of high salinity on physiological and anatomical indices in the early stages of Populus euphratica growth. Russ. J. Plant Physiol. 2015, 62, 229–236. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Elbehiry, F. Rice Production in Egypt: The Challenges of Climate Change and Water Deficiency. In Climate Change Impacts on Agriculture and Food Security in Egypt; Springer: Cham, Switzerland, 2020; pp. 295–319. [Google Scholar] [CrossRef]
- Huang, Y.; Ren, W.; Grove, J.; Poffenbarger, H.; Jacobsen, K.; Tao, B.; Zhu, X.; McNear, D. Assessing synergistic effects of no-tillage and cover crops on soil carbon dynamics in a long-term maize cropping system under climate change. Agric. For. Meteorol. 2020, 291, 108090. [Google Scholar] [CrossRef]
- Li, X.; Zhu, W.; Xu, F.; Du, J.; Tian, X.; Shi, J.; Wei, G. Organic amendments affect soil organic carbon sequestration and fractions in fields with long-term contrasting nitrogen applications. Agric. Ecosyst. Environ. 2021, 322, 107643. [Google Scholar] [CrossRef]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019, 443, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Dotaniya, M.L.; Rajendiran, S.; Meena, B.P.; Meena, A.L.; Dotaniya, C.K.; Jat, R.L.; Saha, J.K. Elevated Carbon Dioxide (CO2) and Temperature vis-a-vis Carbon Sequestration Potential of Global Terrestrial Ecosystem. In Conservation Agriculture; Bisht, J.K., Ed.; Springer: Singapore, 2016. [Google Scholar] [CrossRef]
- Gong, H.; Gao, J. Soil and climatic drivers of plant SLA (specific leaf area). Glob. Ecol. Conserv. 2019, 20, e00696. [Google Scholar] [CrossRef]
- Koller, E.K.; Phoenix, G.K. Seasonal dynamics of soil and plant nutrients at three environmentally contrasting sites along a sub-Arctic catchment sequence. Polar Biol. 2017, 40, 1821–1834. [Google Scholar] [CrossRef]
- Matias, L.; Castro, J.; Zamora, R. Soil-nutrient availability under a global-change scenario in a Mediterranean mountain ecosystem. Glob. Chang. Biol. 2010, 17, 1646–1657. [Google Scholar] [CrossRef]
- Arndal, M.F.; Merrild, M.P.; Michelsen, A.; Schmidt, I.K.; Mikkelsen, T.N.; Beier, C. Net root growth and nutrient acquisition in response to predicted climate change in two contrasting heathland species. Plant Soil 2013, 369, 615–629. [Google Scholar] [CrossRef]
- Pilbeam, D.J. Breeding crops for improved mineral nutrition under climate change conditions. J. Exp. Bot. 2015, 66, 3511–3521. [Google Scholar] [CrossRef] [Green Version]
- Alshaal, T.; El-Ramady, H.; Al-Saeedi, A.H.; Shalaby, T.; Elsakhawy, T.; Omara, A.E.-D.; Gad, A.; Hamad, E.; El-Ghamry, A.; Mosa, A.; et al. The Rhizosphere and Plant Nutrition Under Climate Change. In Essential Plant Nutrients; Naeem, M., Ed.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Agroecology: Implications for Plant Response to Climate Change. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Fekih, I.B.; Ditta, A.; Xing, S. Role of biochar and plant growth promoting rhizobacteria to enhance soil carbon sequestration—A review. Environ. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef] [PubMed]
- Brouder, S.M.; Volenec, J.J. Impact of climate change on crop nutrient and water use efficiencies. Physiol. Plant. 2008, 133, 705–724. [Google Scholar] [CrossRef]
- Bocchiola, D.; Brunetti, L.; Soncini, A.; Polinelli, F.; Gianinetto, M. Impact of climate change on agricultural productivity and food security in the Himalayas: A case study in Nepal. Agric. Syst. 2019, 171, 113–125. [Google Scholar] [CrossRef]
- Amouzou, K.A.; Lamers, J.P.; Naab, J.B.; Borgemeister, C.; Vlek, P.L.; Becker, M. Climate change impact on water- and nitrogen-use efficiencies and yields of maize and sorghum in the northern Benin dry savanna, West Africa. Field Crop. Res. 2019, 235, 104–117. [Google Scholar] [CrossRef]
- Seneweera, S.; Norton, R.M. Plant Responses to Increased Carbon Dioxide. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.M.; Lobell, D.B. Reduction of transpiration and altered nutrient allocation contribute to nutrient decline of crops grown in elevated CO2 concentrations. Plant Cell Environ. 2013, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Li, L.; Cheng, K.; Zheng, J.; Zheng, J.; Shen, M.; Liu, X.; Pan, G. Changes in micronutrient availability and plant uptake under simulated climate change in winter wheat field. J. Soils Sediments 2016, 16, 2666–2675. [Google Scholar] [CrossRef]
- Shahzad, K.; Sintim, H.; Ahmad, F.; Abid, M.; Nasim, W. Importance of Carbon Sequestration in the Context of Climate Change. In Building Climate Resilience in Agriculture; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Adavi, Z.; Moradi, R.; Saeidnejad, A.H.; Tadayon, M.R.; Mansouri, H. Assessment of potato response to climate change and adaptation strategies. Sci. Hortic. 2018, 228, 91–102. [Google Scholar] [CrossRef]
- Cookson, W.; Osman, M.; Marschner, P.; Abaye, D.; Clark, I.; Murphy, D.; Stockdale, E.; Watson, C. Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol. Biochem. 2007, 39, 744–756. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M.; Gessler, A.; Kreuzwieser, J.; Simon, J.; Papen, H. Nitrogen balance in forest soils: Nutritional limitation of plants under climate change stresses. Plant Biol. 2009, 11 (Suppl. 1), 4–23. [Google Scholar] [CrossRef] [PubMed]
- Wrage, N.; Chapuis-Lardy, L.; Isselstein, J. Phosphorus, Plant Biodiversity and Climate Change. In Sociology, Organic Farming, Climate Change and Soil Science; Sustainable Agriculture Reviews 3; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The temperature dependence of organic-matter decomposition—Still a topic of debate. Soil Biol. Biochem. 2006, 38, 2510–2518. [Google Scholar] [CrossRef]
- Howden, S.M.; Crimp, S. Regional impacts: Australia. In Crop Adaptation to Climate Change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 143–155. [Google Scholar]
- Simon, J.; Dannenmann, M.; Pena, R.; Gessler, A.; Rennenberg, H. Nitrogen nutrition of beech forests in a changing climate: Importance of plant-soil-microbe water, carbon, and nitrogen interactions. Plant Soil 2017, 418, 89–114. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Sun, M.; Shafiq, F.; Khalilzadeh, R.; Gao, Z.-Q. Characterizing Differences in Soil Water Content and Wheat Yield in Response to Tillage and Precipitation in the Dry, Normal, and Wet Years at the Loess Plateau. Int. J. Plant Prod. 2021, 15, 655–668. [Google Scholar] [CrossRef]
- Rosenblatt, A.E.; Schmitz, O.J. Climate Change, Nutrition, and Bottom-Up and Top-Down Food Web Processes. Trends Ecol. Evol. 2016, 31, 965–975. [Google Scholar] [CrossRef]
- Jonasson, S.; Michelsen, A.; Schmidt, I.K.; Nielsen, E.V. Responses in microbes and plants to changed temperature, nutrient, and light regimes in the arctic. Ecology 1999, 80, 1828–1843. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Lam, S.K.; Zhang, X.; Liu, X.; Pan, G. Changes in nutrient uptake and utilization by rice under simulated climate change conditions: A 2-year experiment in a paddy field. Agric. For. Meteorol. 2018, 250, 202–208. [Google Scholar] [CrossRef]
- Avila, J.M.; Gallardo, A.; Gómez-Aparicio, L. Pathogen-induced tree mortality interacts with predicted climate change to alter soil respiration and nutrient availability in Mediterranean systems. Biogeochemistry 2019, 142, 53–71. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, P.; Wei, Z.; Liu, J.; Hu, X.; Liu, F. Effects of elevated CO2 and nitrogen supply on leaf gas exchange, plant water relations and nutrient uptake of tomato plants exposed to progressive soil drying. Sci. Hortic. 2021, 292, 110643. [Google Scholar] [CrossRef]
- Luo, Y.; Peng, Q.; Li, K.; Gong, Y.; Liu, Y.; Han, W. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2020, 199, 105100. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Anderson, I.C.; Khachane, A.; Singh, B.P.; Barton, C.V.; Duursma, R.A.; Ellsworth, D.S.; Singh, B.K. Plant productivity is a key driver of soil respiration response to climate change in a nutrient-limited soil. Basic Appl. Ecol. 2020, 50, 155–168. [Google Scholar] [CrossRef]
- Benbi, D.K.; Brar, J.S. A 25-year record of carbon sequestration and soil properties in intensive agriculture. Agron. Sustain. Dev. 2009, 29, 257–265. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Abowaly, M.; Abu Alkheir, A.; Gad, A.-A. Spatial variation of soil carbon and nitrogen pools by using ordinary Kriging method in an area of north Nile Delta, Egypt. Catena 2014, 113, 70–78. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Tsai, H.; Huang, W.-S.; Huang, S.-T. Carbon Storage along with Soil Profile: An Example of Soil Chronosequence from the Fluvial Terraces on the Pakua Tableland, Taiwan. Land 2021, 10, 447. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.; Li, J.; Zhang, S.; Gao, H.; Wang, B.; Shi, X.; Huang, S.; Xu, M. Two-decade long fertilization induced changes in subsurface soil organic carbon stock vary with indigenous site characteristics. Geoderma 2018, 337, 853–862. [Google Scholar] [CrossRef]
- Cao, X.; Long, H.; Lei, Q.; Wu, S. Estimating Soil Carbon Sequestration Potential in Fine Particles of Top Soils in Hebei Province, China. In Digital Soil Mapping Across Paradigms, Scales and Boundaries; Zhang, G.-L., Ed.; Springer: Singapore, 2016. [Google Scholar] [CrossRef]
- Xie, J.; Hou, M.; Zhou, Y.; Wang, R.; Zhang, S.; Yang, X.; Sun, B. Carbon sequestration and mineralization of aggregate-associated carbon in an intensively cultivated Anthrosol in north China as affected by long term fertilization. Geoderma 2017, 296, 1–9. [Google Scholar] [CrossRef]
- Adhikari, K.R.; Dahal, K.R.; Chen, Z.-S.; Tan, Y.-C.; Lai, J.-S. Rice–wheat cropping system: Tillage, mulch, and nitrogen effects on soil carbon sequestration and crop productivity. Paddy Water Environ. 2015, 15, 699–710. [Google Scholar] [CrossRef]
- Baqir, M.; Shah, A.B.; Kothari, R.; Singh, R.P. Carbon sequestration potential of plantation forestry and improvements in soil nutrient status in a subtropical area of northern India. Environ. Sustain. 2018, 1, 383–392. [Google Scholar] [CrossRef]
- Dad, J.M.; Dand, S.A.; Pala, N.A. The effect of bi-culture cover crops on soil quality, carbon sequestration, and growth characteristics in apple orchards of North Western Himalayas. Agrofor. Syst. 2021, 95, 1745–1758. [Google Scholar] [CrossRef]
- Laamrani, A.; Voroney, P.; Gillespie, A.; Chehbouni, A. Development of a Land Use Carbon Inventory for Agricultural Soils in the Canadian Province of Ontario. Land 2021, 10, 765. [Google Scholar] [CrossRef]
- Yadav, G.S.; Lal, R.; Meena, R.S.; Babu, S.; Das, A.; Bhowmik, S.N.; Datta, M.; Layak, J.; Saha, P. Conservation tillage and nutrient management effects on productivity and soil carbon sequestration under double cropping of rice in north eastern region of India. Ecol. Ind. 2019, 105, 303–315. [Google Scholar] [CrossRef]
- Dignac, M.-F.; Derrien, D.; Barre, P.; Barot, S.; Cécillon, L.; Chenu, C.; Chevallier, T.; Freschet, G.T.; Garnier, P.; Guenet, B.; et al. Increasing soil carbon storage: Mechanisms, effects of agricultural practices and proxies. A review. Agron. Sustain. Dev. 2017, 37, 14. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Manxia, C.; Xiumei, G.; Xiaohua, L.; Hongbo, S.; Zhaopu, L.; Zed, R. Carbon sequestration and Jerusalem artichoke biomass under nitrogen applications in coastal saline zone in the northern region of Jiangsu, China. Sci. Total. Environ. 2016, 568, 885–890. [Google Scholar] [CrossRef]
- Laban, P.; Metternicht, G.; Davies, J. Soil Biodiversity and Soil Organic Carbon: Keeping Drylands Alive; IUCN: Gland, Switzerland, 2018; 24p. [Google Scholar] [CrossRef]
- Goh, K.M. Carbon sequestration and stabilization in soils: Implications for soil productivity and climate change. Soil Sci. Plant Nutr. 2004, 50, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Eswaran, H.; Berg, E.V.D.; Reich, P. Organic Carbon in Soils of the World. Soil Sci. Soc. Am. J. 1993, 57, 192–194. [Google Scholar] [CrossRef] [Green Version]
- Lal, R.; Follett, R.F.; Stewart, B.A.; Kimble, J.M. Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci. 2007, 172, 943–956. [Google Scholar] [CrossRef]
- Stout, B.; Lal, R.; Monger, C. Carbon capture and sequestration: The roles of agriculture and soils. Int. J. Agric. Biol. Eng. 2016, 9, 1–8. [Google Scholar]
- Lal, R. Enhancing crop yields in the developing countries through restoration of the soil organic carbon pool in agricultural lands. Land Degrad. Dev. 2005, 17, 197–209. [Google Scholar] [CrossRef]
- Yadav, G.S.; Das, A.; Babu, S.; Mohapatra, K.P.; Lal, R.; Rajkhowa, D. Potential of conservation tillage and altered land configuration to improve soil properties, carbon sequestration and productivity of maize based cropping system in eastern Himalayas, India. Int. Soil Water Conserv. Res. 2021, 9, 279–290. [Google Scholar] [CrossRef]
- Fan, H.-Z.; Chen, Q.-R.; Qin, Y.-S.; Chen, K.; Tu, S.-H.; Xu, M.-G.; Zhang, W.-J. Soil carbon sequestration under long-term rice-based cropping systems of purple soil in Southwest China. J. Integr. Agric. 2015, 14, 2417–2425. [Google Scholar] [CrossRef]
- Shahid, M.; Nayak, A.K.; Puree, C.; Tripathi, R.; Lal, B.; Gautam, P.; Bhattacharyya, P.; Mohanty, S.; Kumar, A.; Panda, B.B.; et al. Carbon and nitrogen fractions and stocks under 41 years of chemical and organic fertilization in a sub-humid tropical rice soil. Soil Tillage Res. 2017, 170, 136–146. [Google Scholar] [CrossRef]
- Meena, R.S.; Sandeep, K.S.; Yadav, G.S. Soil Carbon Sequestration in Crop Production. In Nutrient Dynamics for Sustainable Crop Production; Meena, R.S., Ed.; Springer: Singapore, 2020; pp. 1–39. [Google Scholar] [CrossRef]
- Liang, C.; Zhu, X. The soil Microbial Carbon Pump as a new concept for terrestrial carbon sequestration. Sci. China Earth Sci. 2021, 64, 545–558. [Google Scholar] [CrossRef]
- Feng, Q.; An, C.; Chen, Z.; Wang, Z. Can deep tillage enhance carbon sequestration in soils? A meta-analysis towards GHG mitigation and sustainable agricultural management. Renew. Sustain. Energy Rev. 2020, 133, 110293. [Google Scholar] [CrossRef]
- Crystal-Ornelas, R.; Thapa, R.; Tully, K.L. Soil organic carbon is affected by organic amendments, conservation tillage, and cover cropping in organic farming systems: A meta-analysis. Agric. Ecosyst. Environ. 2021, 312, 107356. [Google Scholar] [CrossRef]
- Kumar, N.; Nath, C.; Hazra, K.; Das, K.; Venkatesh, M.; Singh, M.; Singh, S.; Praharaj, C.; Singh, N. Impact of zero-till residue management and crop diversification with legumes on soil aggregation and carbon sequestration. Soil Tillage Res. 2019, 189, 158–167. [Google Scholar] [CrossRef]
- Nath, C.; Kumar, N.; Das, K.; Hazra, K.; Praharaj, C.; Singh, N. Impact of variable tillage based residue management and legume based cropping for seven years on enzymes activity, soil quality index and crop productivity in rice ecology. Environ. Sustain. Indic. 2021, 10, 100107. [Google Scholar] [CrossRef]
- Kumari, R.; Kundu, M.; Das, A.; Rakshit, R.; Sahay, S.; Sengupta, S.; Ahmad, F. Long-Term Integrated Nutrient Management Improves Carbon Stock and Fruit Yield in a Subtropical Mango (Mangifera indica L.) Orchard. J. Soil Sci. Plant Nutr. 2019, 20, 725–737. [Google Scholar] [CrossRef]
- Hazra, K.K.; Nath, C.P.; Singh, U.; Praharaj, C.S.; Kumar, N.; Singh, S.S.; Singh, N.P. Diversification of maize-wheat cropping system with legumes and integrated nutrient management increases soil aggregation and carbon sequestration. Geoderma 2019, 353, 308–319. [Google Scholar] [CrossRef]
- Chandel, S.; Datta, A.; Yadav, R.K.; Dheri, G.S. Does Saline Water Irrigation Influence Soil Carbon Pools and Nutrient Distribution in Soil under Seed Spices? J. Soil Sci. Plant Nutr. 2021, 21, 949–966. [Google Scholar] [CrossRef]
- Hübner, R.; Kühnel, A.; Lu, J.; Dettmann, H.; Wang, W.; Wiesmeier, M. Soil carbon sequestration by agroforestry systems in China: A meta-analysis. Agric. Ecosyst. Environ. 2021, 315, 107437. [Google Scholar] [CrossRef]
- Mayer, S.; Wiesmeier, M.; Sakamoto, E.; Hübner, R.; Cardinael, R.; Kühnel, A.; Kögel-Knabner, I. Soil organic carbon sequestration in temperate agroforestry systems—A meta-analysis. Agric. Ecosyst. Environ. 2021, 323, 107689. [Google Scholar] [CrossRef]
- Madigan, A.P.; Zimmermann, J.; Krol, D.J.; Williams, M.; Jones, M.B. Full Inversion Tillage (FIT) during pasture renewal as a potential management strategy for enhanced carbon sequestration and storage in Irish grassland soils. Sci. Total Environ. 2021, 805, 150342. [Google Scholar] [CrossRef]
- Shi, S.; Peng, C.; Wang, M.; Zhu, Q.; Yang, G.; Yang, Y.; Xi, T.; Zhang, T. A global meta-analysis of changes in soil carbon, nitrogen, phosphorus and sulfur, and stoichiometric shifts after forestation. Plant Soil 2016, 407, 323–340. [Google Scholar] [CrossRef]
- Pimentel, D.; Burgess, M. Maintaining sustainable and environmentally friendly fresh produce production in the context of climate change. In Global Safety of Fresh Produce; Woodhead Publishing: Sawston, UK, 2014; pp. 133–139. [Google Scholar] [CrossRef]
- Rice, C. Carbon Cycle in Soils | Dynamics and Management. Encycl. Soils Environ. 2005, 164–170. [Google Scholar] [CrossRef]
- Powlson, D.; Cai, Z.; Lemanceau, P. 7 Soil Carbon Dynamics and Nutrient Cycling. Soil Carbon Sci. Manag. Policy Mult. Benefits 2014, 71, 98. [Google Scholar]
- Grover, M.; Maheswari, M.; Desai, S.; Gopinath, K.; Venkateswarlu, B. Elevated CO2: Plant associated microorganisms and carbon sequestration. Appl. Soil Ecol. 2015, 95, 73–85. [Google Scholar] [CrossRef]
- Sánchez-Navarro, V.; Zornoza, R.; Faz, A.; Fernández, J.A. Comparing legumes for use in multiple cropping to enhance soil organic carbon, soil fertility, aggregates stability and vegetables yields under semiarid conditions. Sci. Hortic. 2018, 246, 835–841. [Google Scholar] [CrossRef]
- Zhang, D.; Yao, P.; Zhao, N.; Cao, W.; Zhang, S.; Li, Y.; Huang, D.; Zhai, B.; Wang, Z.; Gao, Y. Building up the soil carbon pool via the cultivation of green manure crops in the Loess Plateau of China. Geoderma 2018, 337, 425–433. [Google Scholar] [CrossRef]
- Meena, A.L.; Jha, P.; Dotaniya, M.L.; Kumar, B.; Meena, B.P.; Jat, R.L. Carbon, Nitrogen and Phosphorus Mineralization as Influenced by Type of Organic Residues and Soil Contact Variation in Vertisol of Central India. Agric. Res. 2019, 9, 232–240. [Google Scholar] [CrossRef]
- Nieder, R.; Benbi, D.K. Carbon and Nitrogen in the Terrestrial Environment; Springer Science & Business Media: Dordrecht, The Netherlands, 2008; 432p. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Abowaly, M.; Gad, A.; Abu_Alkheir, A.; Elbehiry, F. Restoration and sequestration of carbon and nitrogen in the degraded northern coastal area in Nile Delta, Egypt for climate change mitigation. J. Coast. Conserv. 2016, 21, 105–114. [Google Scholar] [CrossRef]
- Elbasiouny, H.; Elbehiry, F. Soil carbon and nitrogen stocks and fractions for improving soil quality and mitigating climate change. Egypt. J. Soil Sci. 2019, 59, 131–144. [Google Scholar] [CrossRef]
- Singh, G.; Sangwan, S.; Narwal, E.; Annapurna, K. Microbial Proteins and Soil Carbon Sequestration. In Adaptive Soil Management: From Theory to Practices; Rakshit, A., Ed.; Springer: Singapore, 2017; pp. 131–139. [Google Scholar] [CrossRef]
- Hines, F.L. Nitrogen, sulfur, and phosphorus and the sequestering of carbon. In Soil Processes and the Carbon Cycle; Lal, R., Kimble, J.M., Follett, R.F., Stewart, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 315–319. [Google Scholar]
- E Richardson, A.; A Kirkby, C.; Banerjee, S.; Kirkegaard, J. The inorganic nutrient cost of building soil carbon. Carbon Manag. 2014, 5, 265–268. [Google Scholar] [CrossRef]
- Knops, J.M.; Tilman, D. Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology 2000, 81, 88–98. [Google Scholar] [CrossRef]
- de Oca, P.C.M. Effect of Land Use, Climate and Soil Structure on Soil Organic Carbon in Costa Rican Ecoregions. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2009. [Google Scholar]
- Bouajila, A.; Gallali, T. Land use effect on soil and particulate organic carbon, and aggregate stability in some soils in Tunisia. Afr. J. Agric. Res. 2010, 5, 764–774. [Google Scholar]
- Cambardella, C.A.; Elliott, E.T. Particulate Soil Organic-Matter Changes across a Grassland Cultivation Sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Su, Y.Z.; Liu, W.J.; Yang, R.; Chang, X.X. Changes in Soil Aggregate, Carbon, and Nitrogen Storages Following the Conversion of Cropland to Alfalfa Forage Land in the Marginal Oasis of Northwest China. Environ. Manag. 2009, 43, 1061–1070. [Google Scholar] [CrossRef]
- Wu, H.; Tang, J.; Huang, H.; Chen, W.; Meng, Y. Net Carbon Sequestration Performance of Cropland Use in China’s Principal Grain-Producing Area: An Evaluation and Spatiotemporal Divergence. Land 2021, 10, 714. [Google Scholar] [CrossRef]
- Don, A.; Schumacher, J.; Scherer-Lorenzen, M.; Scholten, T.; Schulze, E.-D. Spatial and vertical variation of soil carbon at two grassland sites—Implications for measuring soil carbon stocks. Geoderma 2007, 141, 272–282. [Google Scholar] [CrossRef]
- Zapata, F. Introduction to nitrogen management in agricultural systems. In Guidelines on Nitrogen Management in Agricultural Systems; International Atomic Energy Agency: Seibersdorf, Austria, 2008; p. 1. [Google Scholar]
- Qadir, M.; Noble, A.D.; Schubert, S.; Thomas, R.T.; Arslan, A. Sodicity-induced land degradation and its sustainable management: Problems and prospects. Land Degrad. Dev. 2006, 17, 661–676. [Google Scholar] [CrossRef]
- Choudhury, S.G.; Yaduvanshi, N.P.S.; Chaudhari, S.K.; Sharma, D.R.; Nayak, D.C.; Singh, S.K. Effect of nutrient management on soil organic carbon sequestration, fertility, and productivity under rice-wheat cropping system in semi-reclaimed sodic soils of North India. Environ. Monit. Assess. 2018, 190, 117. [Google Scholar] [CrossRef] [PubMed]
- Srinivasarao, C.; Lal, R.; Kundu, S.; Thakur, P.B. Conservation Agriculture and Soil Carbon Sequestration. In Conservation Agriculture; Farooq, M., Siddique, K.H.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Lal, R. Residue management, conservation tillage and soil restoration for mitigating greenhouse effect by CO2 enrichment. Soil Tillage Res. 1997, 43, 81–107. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P. Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 2014, 17, 288–293. [Google Scholar] [CrossRef]
- Mahmud, A.A.; Upadhyay, S.K.; Srivastava, A.K.; Bhojiya, A.A. Biofertilizers: A Nexus between soil fertility and crop productivity under abiotic stress. Curr. Res. Environ. Sustain. 2021, 3, 100063. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Saxena, A.K.; Singh, J.S.; Singh, D.P. Impact of Native ST-PGPR (Bacillus pumilus; EU927414) on PGP Traits, Antioxidants Activities, Wheat Plant Growth and Yield under Salinity. Clim. Chang. Environ. Sustain. 2019, 7, 157. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Edrisi, S.A. Developing sustainable measures to restore fly ash contaminated lands: Current challenges and future prospects. Land Degrad. Dev. 2021, 32, 4817–4831. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Climate Variable | Study Objective | The Effect on Plant | The Ecosystem | Ref. |
|---|---|---|---|---|
|
|
| Two plants community predominated by Cassiope tetragona (L.) | [39] |
|
|
| Mediterranean forest, shrubland, and open areas | [16] |
| Study the root nutrient uptake under predicted climate change |
| Dry heathland/grassland | [17] |
| Investigate the effect of CO2 enrichment and warming on the availability and plant uptake of soil micronutrients |
|
| [28] |
| Investigate the effect of environmental conditions on nutrient availability |
|
| [15] |
| Studying the effect of CO2 enrichment and canopy warming on nutrient concentration and accumulation (among other aims) |
|
| [40] |
| Examining the effect of climatic variables and pathogens on C, N, and P |
| Mediterranean forest soil | [41] |
|
| [42] | ||
|
|
| Desert soil | [43] |
|
|
| Eucalyptus globulus plants | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbasiouny, H.; El-Ramady, H.; Elbehiry, F.; Rajput, V.D.; Minkina, T.; Mandzhieva, S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability 2022, 14, 914. https://doi.org/10.3390/su14020914
Elbasiouny H, El-Ramady H, Elbehiry F, Rajput VD, Minkina T, Mandzhieva S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability. 2022; 14(2):914. https://doi.org/10.3390/su14020914
Chicago/Turabian StyleElbasiouny, Heba, Hassan El-Ramady, Fathy Elbehiry, Vishnu D. Rajput, Tatiana Minkina, and Saglara Mandzhieva. 2022. "Plant Nutrition under Climate Change and Soil Carbon Sequestration" Sustainability 14, no. 2: 914. https://doi.org/10.3390/su14020914
APA StyleElbasiouny, H., El-Ramady, H., Elbehiry, F., Rajput, V. D., Minkina, T., & Mandzhieva, S. (2022). Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability, 14(2), 914. https://doi.org/10.3390/su14020914