

Arbuscular Mycorrhizal Fungi in Sustainable Agriculture


 ,
,  ,
,  ,
, 
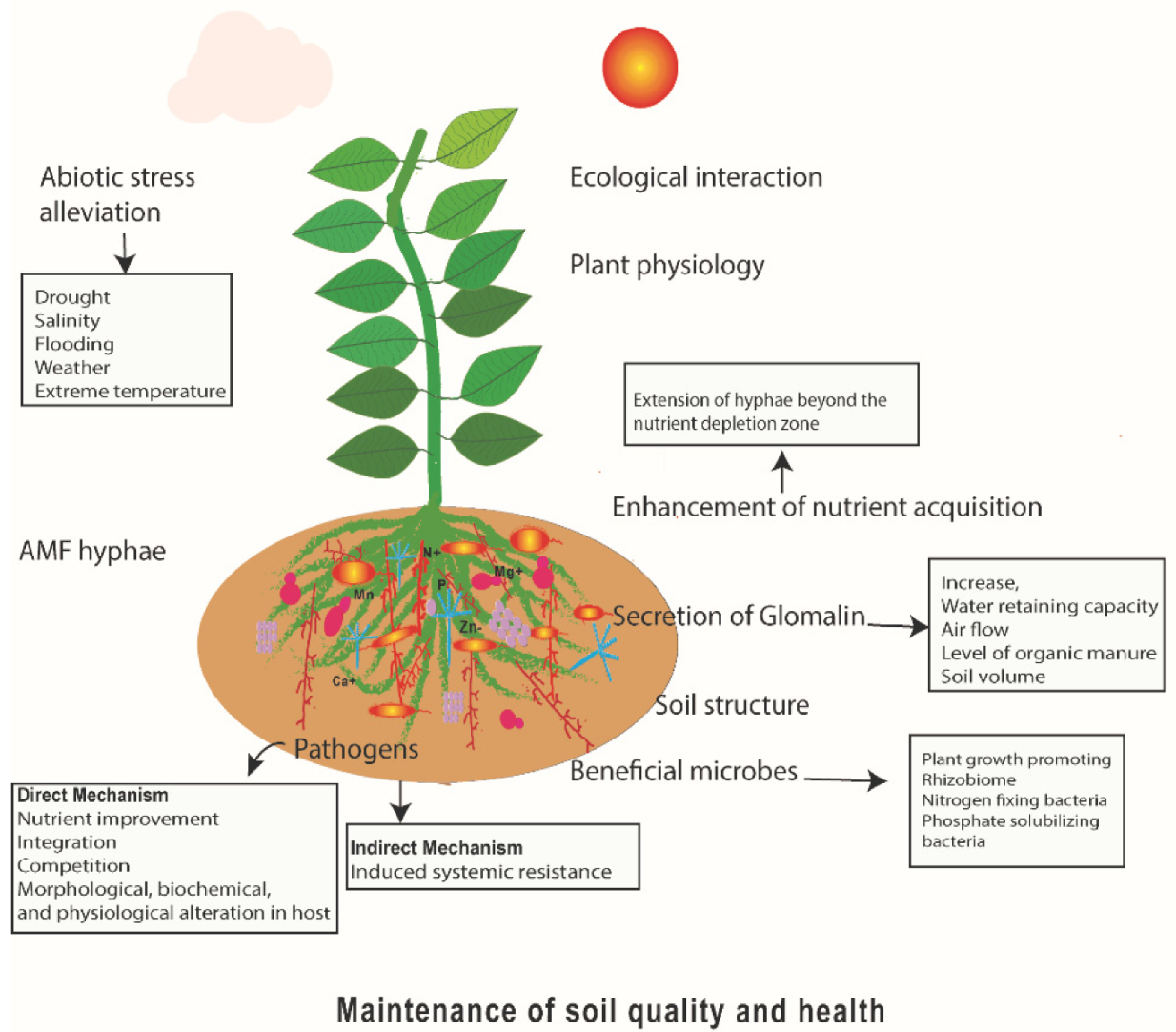{kind=link}
{kind=link}
Abstract
1. Introduction
2. Background of Mycorrhizal Fungi Symbiosis
3. Factors Affecting the Functional Effect of AMF in the Agricultural Field
4. Improvement of Soil Quality and Health
4.1. Maintenance of Plant Physiology
4.1.1. Stimulation of Soil Fertility by Enhancing Nutrient Acquisition
4.1.2. Abiotic Stress Alleviation
4.2. Ecological Interactions
4.2.1. Beneficial Ecological Interactions
4.2.2. Pathogen Biocontrol
5. Drought Resistance
6. Land Restoration
7. Mycorrhizae Improve Seed Germination and Seedling Establishment
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velten, S.; Leventon, J.; Jager, N.; Newig, J. What is sustainable agriculture? A systematic review. Sustainability 2015, 7, 7833–7865. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Charters, M.D.; Elliott, A.J.; Sait, S.M.; Field, K.J. Are mycorrhizal fungi our sustainable saviours? considerations for achieving food security. J. Ecol. 2017, 105, 921–929. [Google Scholar] [CrossRef]
- Singh, J.S.; Pandey, V.C.; Singh, D.P. Efficient soil microorganisms: A new dimension for sustainable agriculture and environmental development. Agric. Ecosyst. Environ. 2011, 140, 339–353. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Swaminathan, M.S. Can science and technology feed the world in 2025? Field Crop Res. 2007, 104, 3–9. [Google Scholar] [CrossRef]
- Wezel, A.; Bellon, S.; Doré, T.; Francis, C.; Vallod, D.; David, C. Agroecology as a science, a movement or a practice. A review. Agron. Sustain. Dev. 2009, 29, 503–515. [Google Scholar] [CrossRef]
- Gianinazzi-pearson, V. Plant cell responses to arbuscular mycorrhizal fungi: Getting to the roots of the symbiosis. Plant Cell 1996, 8, 1871–1883. [Google Scholar] [CrossRef]
- Erik, V.; Van der Heijden, M.G.A.; Rilling, M.C.; Toby, K.E. Mycorrhizal fungal establishment in agricultural soils: Factors determining inoculation success. New Phytol. 2012, 197, 1104–1109. [Google Scholar]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Rosendahl, S. Communities, populations and individuals of arbuscular mycorrhizal fungi. New Phytol. 2008, 178, 253–266. [Google Scholar] [CrossRef]
- Caruso, T.; Hempel, S.; Powell, J.R.; Barto, E.K.; Rillig, M.C. Compositional divergence and convergence in arbuscular mycorrhizal fungal communities. Ecology 2012, 93, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Kodama, K.; Rich, M.K.; Kyozuka, J. An ancestral function of strigolactones as symbiotic rhizosphere signals. Nat. Commun. 2022, 13, 3974. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science. 2020, 367, 6480. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Sato, H.; Yamamoto, S.; Kadowaki, K.; Tanabe, A.S.; Yazawa, S. How are plant and fungal communities linked to each other in belowground ecosystems?A massively parallel pyrosequencing analysis of the association specificity of root-associated fungi and their host plants. Ecol. Evol. 2013, 3, 3112–3124. [Google Scholar] [CrossRef] [PubMed]
- Ram Singh, S.; Singh, U.; Chaubey, A.; Bhat, M.I. Mycorrhizal fungi for sustainable agriculture a review. Agric. Rev. 2010, 31, 93–104. [Google Scholar]
- Lekberg, Y.; Rosendahl, S.; Olsson, P.A. The fungal perspective of arbuscular mycorrhizal colonization in ‘nonmycorrhizal’ plants. New Phytol. 2014, 4, 1399–1403. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.C.; Negi, S. AMF: The future prospect for sustainable agriculture. Physiol. Mol. Plant Pathol. 2018, 102, 36–45. [Google Scholar] [CrossRef]
- Bonfante, P. The future has roots in the past: The ideas and scientists that shaped mycorrhizal research. New Phytol. 2018, 220, 982–995. [Google Scholar] [CrossRef]
- Salmeron-Santiago, I.A.S.; Martinez-Trujillo, M.; Valdez-Alarcon, J.J.; Pedraza-Santos, M.E.; Santoyo, G.; Pozo, M.J.; Chavez-Barcenas, A.T. An updated review on the modulation of carbon partitioning and allocation in arbuscular mycorrhizal plants. Microorganisms 2022, 10, 75. [Google Scholar] [CrossRef]
- Manoharachary, C.; Kunwar, I.K.; Mukerji, K.G. Some Aspects of Monotropoid Mycorrhizae. In Techniques in Mycorrhizal Studies; Mukerji, K.G., Manoharachary, C., Chamola, B.P., Eds.; Springer: Dorgrecht, The Netherlands, 2002; pp. 435–441. [Google Scholar]
- Soka, G. Arbuscular mycorrhizal symbiosis and ecosystem processes: Prospects for future research in tropical soils. Open J. Ecol. 2014, 4, 11–22. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Khan, M.Y.; Waqas, M.R.; Binyamin, R.; Akhtar, S.; Zahir, Z.A. Arbuscular Mycorrhizas: An Overview. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.S., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 1–327. [Google Scholar]
- Bowles, T.M.; Barrios-Masias, F.H.; Carlisle, E.A.; Cavagnaro, T.R.; Jackson, L.E. Effects of arbuscular mycorrhizae on tomato yield, nutrient uptake, water relations, and soil carbon dynamics under deficit irrigation in field conditions. Sci. Total Environ. 2016, 566, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, E. Mycorrhizal symbiosis modulates the rhizosphere microbiota to promote rhizobia-legume symbiosis. Mol. Plant 2021, 14, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Besserer, A.; Puech-Pagès, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.C.; Roux, C.; Bécard, G.; Séjalon-Delmas, N. Strigolactones Stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Suseela, V. Unraveling arbuscular mycorrhiza-induced changes in plant primary and secondary metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Henke, C.; Jung, E.; Kothe, E. Hartig net formation of Tricholoma vaccinum spruce ectomycorrhizal in hydroponic culture. Environ. Sci. Pollut. 2015, 22, 19394–19399. [Google Scholar] [CrossRef] [PubMed]
- Frater, P.N. Factors Affecting Mycorrhizal Colonization in Schizachyrium scoparium. Master’s Thesis, lowa State University, Ames, IA, USA, 2012. [Google Scholar]
- Sulistiona, W. The role of mycorrhizae on seedling and early growth of sugarcane. In Mycorrhizal Fungi—Utilization in Agriculture and Forestry; IntechOpen: London, UK, 2016; pp. 1–10. [Google Scholar] [CrossRef]
- Yang, C.; Ellouze, W.; Navarro-Borrell, A.; Taheri, A.E.; Klabi, R.; Dai, M.; Kabir, Z.; Hamel, C. Management of the Arbuscular Mycorrhizal Symbiosis in Sustainable Crop Production. In Mycorrhizal Fungi: Use in Sustainable Agriculture and Land Restoration; Solaiman, Z.M., Abbott, L.K., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 89–118. [Google Scholar]
- Varga, S. Effect of arbuscular mycorrhizal fungi and maternal plant sex on seed germination and early plant establishment. Am. J. Bot. 2015, 102, 358–366. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 2019, 10, e499. [Google Scholar] [CrossRef]
- Carrenho, R.; Trufem, S.F.B.; Bononi, V.L.R.; Silva, E.S. The effect of different soil properties on arbuscular mycorrhizal colonization of peanuts, sorghum and maize. Acta Bot. Bras. 2007, 21, 723–730. [Google Scholar] [CrossRef]
- Wilson, H.E. Climate Change Effects on Arbuscular Mycorrhizal Fungi and Prairie Plants along a Mediterranean Climate Gradient; University of Oregon: Eugene, OR, USA, 2012. [Google Scholar]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J. The contribution of arbuscular mycorrhizal fungi in sustainable maintenace of plant health and soil fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Raut, R.R.; Harale, P.; Kurhe, A. Studies on soil quality paramenters in relations to cropping patterns, micronutrients and pH from Goagalgaon are in Ahmednagar district of Maharashta, India. Int. J. Multidiscip. Curr. Res. 2020, 6, 210–219. [Google Scholar]
- Bunemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Gupta, M.M. Arbuscular Mycorrhizal Fungi: Potential Soil Health Indicators. In Soil Health; Giri, B., Varma, A., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 187–195. [Google Scholar]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of arbuscular mycorrhizal fungi on soil fertility: Contribution in the improvement of physical, chemical and biological properties of the soil. Front. Fungal Biol. 2022, 3, 1–11. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Veresoglou, S.D.; Lehmann, A.; Morris, E.K.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—A meta-analysis. Plant Soil. 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Li, Y.; Xu, J.; Hu, J.; Zhang, T.; Wu, X.; Yang, Y. Arbuscular mycorrhizal fungi and glomalin play a crucial role in soil aggregate stability in Pb—Contaminated soil. Int. J. Environ. Res. Public Health 2022, 19, 5029. [Google Scholar] [CrossRef] [PubMed]
- Bone, F.; Rubio, R.; Morales, A. Arbuscular mycorrhizal fungi and soil aggregation. J. Soil Sc. Plant Nutr. 2008, 8, 9–18. [Google Scholar]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef]
- Herman, D.J.; Firestone, M.K.; Nuccio, E.; Hodge, A. Interactions between an arbuscular mycorrhizal fungus and a soil microbial community mediated litter decomposition. FEMS Microbiol. Ecol. 2012, 80, 236–247. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Hardan, A.N. Importance of mycorrhizae in crop productivity. In Mitigating Environmental Stresses for Agricultural Sustainability in Egypt; Awaad, H., Abu-hashim, M., Negm, A., Eds.; Springer Nature: Cham, Switzerland, 2021; pp. 471–484. [Google Scholar]
- Ingraffia, R.; Amato, G.; Frenda, A.S.; Giambalvo, D. Impacts of arbuscular mycorrhizal fungi on nutrient uptake, N2 fixation, N transfer, and growth in a wheat/faba bean intercropping system. PLoS ONE. 2019, 14, e0213672. [Google Scholar] [CrossRef]
- Johnson, N.C.; Gehring, C.A. Mycorrhizas: Symbiotic Mediators of Rhizosphere and Ecosystem Processes. In The Rhizosphere; Cardon, Z.G., Whitbeck, J.L., Eds.; Elsevier Academic Press: Burlington, VT, USA, 2007; pp. 73–100. [Google Scholar]
- Fan, L.; Dalpe, Y.; Fang, C.; Dube, C.; Khanizadeh, S. Influence of arbuscular mycorrhizae on biomass and root morphology of selected strawberry cultivars under salt stress. Botany 2011, 89, 397–403. [Google Scholar] [CrossRef]
- Bücking, H.; Liepold, E.; Ambilwade, P. The role of the mycorrhizal symbiosis in nutrient uptake of plants and the regulatory mechanisms underlying these transport processes. In Plant Science; Dhal, N.K., Sahu, S.C., Eds.; IntechOpen: London, UK, 2012; Volume 4, pp. 108–132. [Google Scholar]
- Ortas, I.; Rafique, M. The Mechanisms of Nutrient Uptake by Arbuscular Mycorrhizae. In Mycorrhiza—Nutrient Uptake, Biocontrol, Ecorestoration, 4th ed.; Varma, A., Prasad, R., Tuteja, N., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 1–19. [Google Scholar]
- Liu, J.; Guo, C.; Chen, Z.; He, J.; Zou, Y. Mycorrhizal inoculation modulates root morphology and root phytohormone responses in trifoliate orange under drought stress. Emir. J. Food Agric. 2016, 28, 251–256. [Google Scholar] [CrossRef]
- Issam, A.; Ghadraoui, O.E.; Serbouti, S.; Ahamad, H.; Mansouri, I.; Kamari, F.E.; Taroq, A.; Ousaaid, D.; Squalli, W.; Farah, A. The mechanisms of absorption and nutrients transport in plants: A review. Trop. J. Nat Prod. Res. 2022, 6, 8–14. [Google Scholar]
- Smith, S.E.; Gianinazzi-pearson, V.; Koide, R.; Cairney, J.W.G. Nutrient transporters in mycorrhizas: Structure, physiology and consequences for efficiency of the symbiosis. Plant Soil 1994, 159, 103–113. [Google Scholar] [CrossRef]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Kobae, Y. Dynamic phosphate uptake in arbuscular mycorrhizal roots under field conditions. Front. Environ. Sci. 2019, 6, 159. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bucking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient exchange and regulation in arbuscular mycorrhizal symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef]
- Chandrasekarana, M. Meta-analytical approach on arbuscular mycorrhizal fungi inoculation efficiency on plant growth and nutrient uptake. Agriculture 2020, 10, 370. [Google Scholar] [CrossRef]
- Nowicka, B.; Ciura, J.; Szymanska, R.; Kruk, J. Improving photosynthesis, plant productivity and abiotic stress tolerance—Current trends and future perspectives. J. Plant Physiol. 2018, 231, 415–433. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–58. [Google Scholar] [CrossRef]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant. Sci. 2019, 10, 470. [Google Scholar] [CrossRef]
- Yadav, R.S.; Mahatma, M.K.; Thirumalaisamy, P.P.; Meena, H.N.; Bhaduri, D.; Arora, S.; Panwar, J. Arbuscular Mycorrhizal Fungi (AMF) for Sustainable Soil and Plant Health in Salt-Affected Soils. In Bioremediation of Salt Affected Soils: An Indian Perspective; Arora, S., Singh, A.K., Singh, Y.P., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 133–156. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Upreti, K.K.; Bhatt, R.M.; Panneerselvam, P.; Varalakshmi, L.R. Morpho-physiological responses of grape rootstock “Dogridge” to arbuscular mycorrhizal fungi inoculation under salinity stress. Int. J. Fruit Sci. 2016, 16, 191–209. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: A meta-analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Garg, N. High Effectiveness of Rhizophagus irregularis is linked to superior modulation of antioxidant defence mechanisms in Cajanus cajan (L.) Millsp. genotypes grown under salinity stress. Mycorrhiza 2017, 27, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Sagar, A.; Rathore, P.; Ramteke, P.W.; Ramakrishna, W.; Reddy, M.S.; Pecoraro, L. Plant growth promoting rhizobacteria, arbuscular mycorrhizal fungi and their synergistic interactions to counteract the negative effects of saline soil on agriculture: Key macromolecules and mechanisms. Microorganisms 2021, 9, 1491. [Google Scholar] [CrossRef]
- Cano-tamayo, A.I.; Zuleta-Patino, D.M.; Rios-osorio, L.A. Role of mycorrhiza-forming fungi in the bioremediation of agricultural soils contaminated with heavy metals: A systematic review. Rev. Hechos. Microbiol. 2016, 7, 61–75. [Google Scholar]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, L.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi—From ecology to application. Front. Plant Sci. 2018, 9, e1270. [Google Scholar] [CrossRef]
- Shan, B.Q.; Qu, M.M.; Li, J.; Li, C. Bioremediation of petroleum contaminated soil by plants and arbuscular mycorrhizal fungi in northern Shaanxi. Acta Prataculturae Sin. 2016, 25, 87–94. [Google Scholar]
- De Andrade, S.A.L.; De Silveira, A.P.D.; Jorge, R.A.; De Abreu, M.F. Cadmium accumulation in sunflower plants influenced by mycorrhiza. Int. J. Phytoremediat. 2008, 10, 1–13. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.I.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, e1106. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Behl, R.K.; Singh, K.P.; Jain, P.; Narula, N. Performance and gene effects for wheat yield under inoculation of arbuscular mycorrhiza fungi and Azotobacter chroococcum. Plant Soil Environ. 2004, 50, 409–415. [Google Scholar] [CrossRef]
- Abdurrahman, M.; Somenahally, A.; Gentry, T. Interactions of arbuscular mycorrhizal fungi with hyphosphere microbial communities in a saline soil: Impacts on phosphorus availability and alkaline phosphatase gene abundance. Soil Syst. 2020, 4, 63. [Google Scholar]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, e1089. [Google Scholar] [CrossRef] [PubMed]
- Nacoon, S.; Jogloy, S.; Riddech, N.; Mongkolthanaruk, W.; Kuyper, T.W.; Boonlue, S. Interaction between phosphate solubilizing bacteria and arbuscular mycorrhizal fungi on growth promotion and Tuber inulin content of Helianthus tuberosus L. Sci. Rep. 2020, 10, 4916. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.D.J.; Miranda, J.P.; Ferrari, J.; Hartley, S.E.; Hodge, A. Aphids influence soil fungal communities in conventional agricultural systems. Front. Plant Sci. 2019, 10, e895. [Google Scholar] [CrossRef] [PubMed]
- Schouteden, N.; Waele, D.D.; Panis, B.; Vos, C.M. Arbuscular mycorrhizal fungi for the biocontrol of plant-parasitic nematodes: A review of mechanisms involved. Front. Microbiol. 2015, 6, e1280. [Google Scholar] [CrossRef]
- Tahat, M.M.; Sijim, K.; Othman, R. Mycorrhizal fungi as a biocontrol agent. Plant Pathol. J. 2010, 9, 198–207. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef]
- Young, T.; Cameron, D.D.; Phoenix, G.K. Using AMF inoculation to improve the nutritional status of Prunella vulgaris plants in green roof substrate during establishment. Urban For. Urban Green. 2015, 14, 959–967. [Google Scholar] [CrossRef]
- del Mar Alguacil, M.; Torrecillas, E.; Lozano, Z.; Roldán, A. Evidence of differences between the communities of arbuscular mycorrhizal fungi colonizing galls and roots of Prunus persica infected by the root-knot nematode Meloidogyne incognita. Appl. Environ. Microbiol. 2011, 77, 8656–8661. [Google Scholar] [CrossRef] [PubMed]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 419. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, N. Mycorrhizal Fungi as Bioprotectors of Crops Against Verticillium wilt—A hypothetical scenario under changing environmental conditions. Plants 2020, 9, 1468. [Google Scholar] [CrossRef] [PubMed]
- Folli-Pereira, M.d.S.; Garlet, J.; Bertolazi, A.A. Arbuscular mycorrhizal fungi and their potential applications for sustainable agriculture, Agriculturally Important Fungi for Sustainable Agriculture. In Perspective for Diversity and Crop Productivity; Yadav, A.N., Mishra, S., Kour, D., Yadav, N., Kumar, A., Eds.; Springer: Cham, Switzerland, 2020; Volume 1, pp. 109–119. [Google Scholar]
- Begum, H.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved drought tolerance by AMF inoculation in maize (Zea mays) involves physiological and biochemical implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, Y.; Shi, Z.; Feng, G. Arbuscular mycorrhizal fungi can accelerate the restoration of degraded spring grassland in central Asia. Rangel. Ecol. Manag. 2012, 65, 426–432. [Google Scholar] [CrossRef]
- Shuab, R.; Lone, R.; Ahmad, J.; Reshi, Z.A. Arbuscular Mycorrhizal Fungi: A Potential Tool for Restoration of Degraded Land. In Mycorrhiza-Nutrient Uptake, Biocontrol, Ecorestoration; Varma, A., Prasad, R., Tuteja, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 415–434. [Google Scholar]
- De Moura, M.A.; Oki, Y.; Arantes-Garcia, L.; Cornelissen, T.; Nunes, Y.R.F.; Fernandes, G.W. Mycorrhiza fungi application as a successful tool for worldwide mine land restoration: Current state of knowledge and the way forward. Ecol. Eng. 2022, 178, e106580. [Google Scholar] [CrossRef]
- Prayudyningsih, R.; Sari, R.; Mangopang, A.D. Isolation of indigenous arbuscular mycorrhizal fungi (AMF) to support revegetation on the nickel post-mining land. Earth. Environ. Sci. 2019, 308, 012038. [Google Scholar]
- Asmelash, F.; Bekele, T.; Birhane, E. The potential role of arbuscular mycorrhizal fungi in the restoration of degraded lands. Front. Microbiol. 2016, 7, e1095. [Google Scholar] [CrossRef]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Jaunatre, R.; Fonvielle, N.; Spiegelberger, T.; Buisson, E.; Dutoit, T. Recovery of arbuscular mycorrhizal fungi root colonization after severe anthropogenic disturbance: Four species assessed in old-growth Mediterranean grassland. Folia Geobot. 2016, 51, 319–332. [Google Scholar] [CrossRef]
- Rosmim, A.T.; Íris, V.; Sónia, V.G.; Rui, O.; Célia, M.M.; Orlando, A.Q. Arbuscular mycorrhizal fungi (AMF) promote the growth of the pioneer dune plant of coastal areas. African J. Microbiol. Res. 2020, 14, 579–586. [Google Scholar] [CrossRef]
- Requena, N.; Perez-Solis, E.; Azcón-Aguilar, C.; Jeffries, P.; Barea, J.M. Management of indigenous plant-microbe symbioses aids restoration of desertified ecosystems. Appl. Environ. Microbiol. 2001, 67, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, S.; Lendzemo, V.; Langer, I.; Schweiger, P.; Khaosaad, T.; Toussaint, J.; Vierheilig, H. Flavonoids and strigolactones in root exudates as signals in symbiotic and pathogenic plant fungus interaction. Molecules 2007, 12, 1290–1306. [Google Scholar] [CrossRef] [PubMed]
- Ellouze, W.; Hamel, C.; Cruz, A.F.; Ishii, T.; Gan, Y.; Bouzid, S.; St-Arnaud, M. Phytochemicals and spore germination: At the root of AMF host preference? Appl. Soil Ecol. 2012, 60, 98–104. [Google Scholar] [CrossRef]
- Xiuxiu, S.; Yansu, L.; Xianchang, Y.; Chaoxing, H. Effects of arbuscular mycorrhizal fungi (AMF) inoculums on cucumber seedlings. Adv. Plants Agric. 2019, 9, 127–130. [Google Scholar]
- Van Der Heijden, M.G.A. Arbuscular mycorrhizal fungi as support system for seedling establishment in grassland. Ecol. Lett. 2004, 7, 293–303. [Google Scholar] [CrossRef]
- Shao, S.C.; Luo, Y.; Jacquemyn, H. Co-cultures of mycorrhizal fungi do not increase germination and seedling development in the epiphytic orchid Dendrobium nobile. Front. Plant Sci. 2020, 11, e571426. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S.A. Influence of mycorrhizal fungi on seed germination and growth in terrestrial and epiphytic orchids. Saudi J. Biol. Sci. 2019, 26, 495–502. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalamulla, R.; Karunarathna, S.C.; Tibpromma, S.; Galappaththi, M.C.A.; Suwannarach, N.; Stephenson, S.L.; Asad, S.; Salem, Z.S.; Yapa, N. Arbuscular Mycorrhizal Fungi in Sustainable Agriculture. Sustainability 2022, 14, 12250. https://doi.org/10.3390/su141912250
Kalamulla R, Karunarathna SC, Tibpromma S, Galappaththi MCA, Suwannarach N, Stephenson SL, Asad S, Salem ZS, Yapa N. Arbuscular Mycorrhizal Fungi in Sustainable Agriculture. Sustainability. 2022; 14(19):12250. https://doi.org/10.3390/su141912250
Chicago/Turabian StyleKalamulla, Ruwanthika, Samantha C. Karunarathna, Saowaluck Tibpromma, Mahesh C. A. Galappaththi, Nakarin Suwannarach, Steven L. Stephenson, Suhail Asad, Ziad Salman Salem, and Neelamanie Yapa. 2022. "Arbuscular Mycorrhizal Fungi in Sustainable Agriculture" Sustainability 14, no. 19: 12250. https://doi.org/10.3390/su141912250
APA StyleKalamulla, R., Karunarathna, S. C., Tibpromma, S., Galappaththi, M. C. A., Suwannarach, N., Stephenson, S. L., Asad, S., Salem, Z. S., & Yapa, N. (2022). Arbuscular Mycorrhizal Fungi in Sustainable Agriculture. Sustainability, 14(19), 12250. https://doi.org/10.3390/su141912250