
Eucalyptus Growth Responses to Soil Water Storage Capacity in Arenosols and Acrisols Soils: Wood and Biomass Stock Modelling


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. The 3-PG Model
2.3. Experimental Design and Sampling
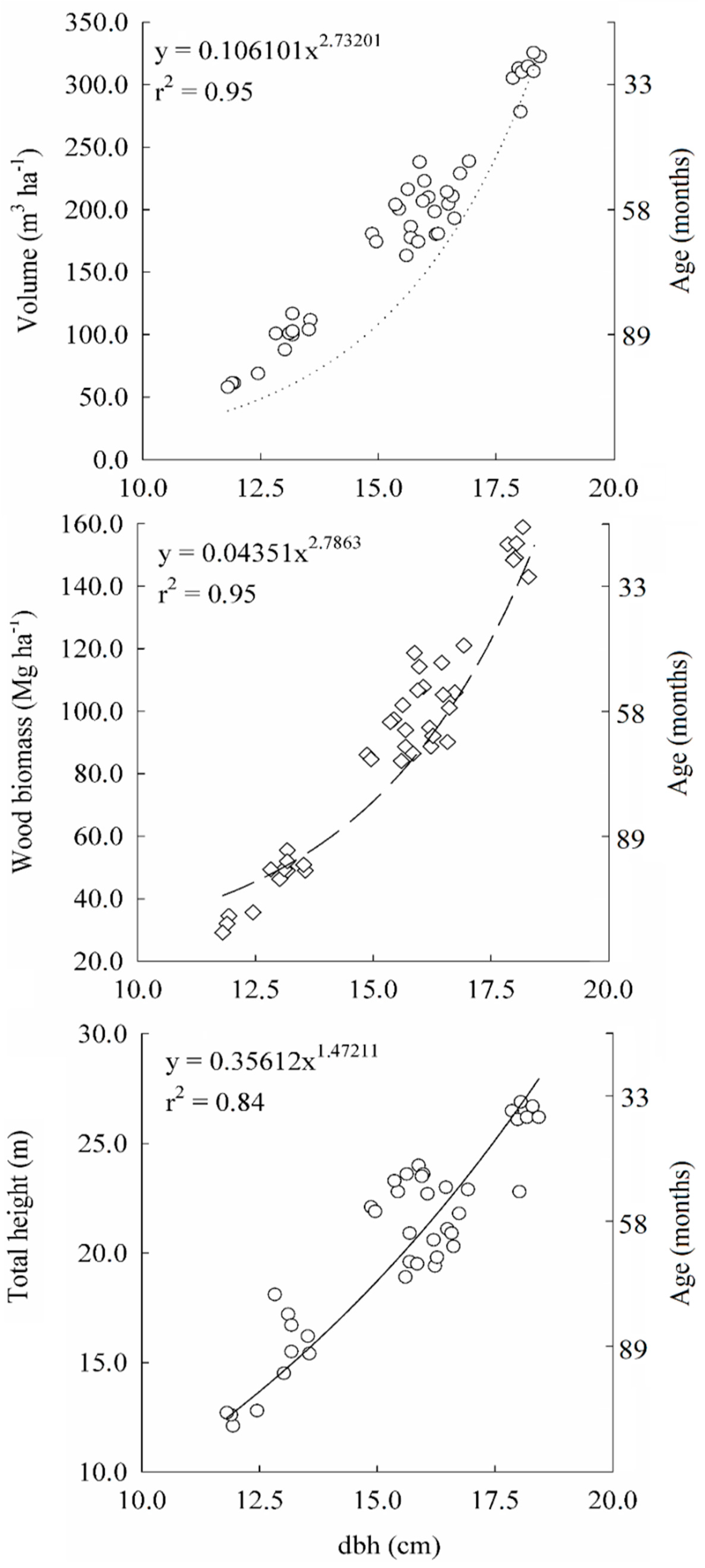
2.4. Growth, Dendrometric Characterization and Aboveground Biomass Stock
2.5. Edaphic Properties
2.6. Parameterization, Calibration and Validation of the 3-PG Model
3. Results
3.1. Parameterization of the 3-PG Model
3.2. Efficiency and Validation of the Model
3.3. Growth, Dendrometric Characterization and Aboveground Biomass Stock
4. Discussion
4.1. Model Parameterization
4.2. Eucalyptus Productivity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Razakamanarivo, R.H.; Razakavololona, A.; Razafindrakoto, M.; Vieilledent, G.; Albrecht, A. Below-ground biomass production and allometric relationships of eucalyptus coppice plantation in the central highlands of Madagascar. Biomass Bioenerg. 2012, 45, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, D.; Song, M. Biomass and carbon storage of Eucalyptus and Acacia plantations in the Pearl River Delta, South China. For. Ecol. Manag. 2012, 277, 90–97. [Google Scholar] [CrossRef]
- Djomo, S.N.; Ac, A.; Zenone, T.; De Groote, T.; Bergante, S.; Facciotto, G.; Sixto, H.; Ciria, P.; Weger, J.; Ceulemans, R. Energy performances of intensive and extensive short rotation cropping systems for woody biomass production in the EU. Renew. Sustain. Energy Rev. 2015, 41, 845–854. [Google Scholar] [CrossRef]
- IBÁ—Indústria Brasileira de Árvores. Relatório 2015. Ano Base 2014. Brasília. 2015. Available online: https://iba.org/images/shared/iba_2015.pdf (accessed on 21 July 2022).
- França, J.S.; Reichert, J.M.; Holthusen, D.; Rodrigues, M.F.; Araújo, E.F. Subsoiling and mechanical hole-drilling tillage effects on soil physical properties and initial growth of eucalyptus after eucalyptus on steeplands. Soil Tillage Res. 2021, 207, 104860. [Google Scholar] [CrossRef]
- Reichert, J.M.; Morales, C.A.S.; Lima, E.M.; Bastos, F.; Sampietro, J.A.; Araújo, E.F.; Srinivasan, R. Best tillage practices for early-growth of clonal eucalyptus in soils with distinct granulometry, drainage and profile depth. Soil Tillage Res. 2021, 212, 105038. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Maguire, D.A.; Monserud, R.A.; Johnson, G.P. A hybrid model for intensively managed Douglas-fir plantations in the Pacific Northwest, USA. Eur. J. For. Res. 2009, 129, 325–338. [Google Scholar] [CrossRef]
- Wang, W.; Peng, C.; Zhang, S.Y.; Zhoua, X.; Larocquec, G.R.; Kneeshawa, D.D.; Leia, X. Development of TRIPLEX-Management model for simulating the response of forest growth to pre-commercial thinning. Ecol. Modell. 2011, 222, 2249–2261. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grote, R.; Reineking, B.; Rotzeri, T.H.; Seifert, S.T. Models for forest ecosystem management: A European perspective. Ann. Bot. 2008, 101, 1065–1087. [Google Scholar] [CrossRef]
- Robinson, A.P.; Ek, A.R. Description and validation of a hybrid model of forest growth and stand dynamics for the Great Lakes region. Ecol. Modell. 2003, 170, 73–104. [Google Scholar] [CrossRef]
- Peng, C.; Liu, J.; Dang, Q.; Apps, M.J.; Jiang, H. TRIPLEX: A generic hybrid model for predicting forest growth and carbon and nitrogen dynamics. Ecol. Modell. 2002, 153, 109–130. [Google Scholar] [CrossRef]
- Sands, P.J.; Landsberg, J.J. Parameterisation of 3-PG for plantation grown Eucalyptus grobulus. For. Ecol. Manag. 2002, 163, 273–292. [Google Scholar] [CrossRef]
- Palma, J.H.N.; Hakamada, R.; Moreira, G.G.; Nobre, S.; Rodrigues, L.C.E. Using 3PG to assess climate change impacts on management plan optimization of Eucalyptus plantations. A case study in Southern Brazil. Sci. Rep. 2021, 11, 2708. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.C.; Landsberg, J.J.; Sands, P.J. Parameterisation of 3-PG model for fast-growing Eucalyptus grandis plantations. For. Ecol. Manag. 2004, 193, 179–195. [Google Scholar] [CrossRef]
- Zhao, M.; Xiang, W.; Deng, X.; Tian, D.; Huang, Z.; Zhou, X.; Yu, G.; He, H.; Pen, C. Application of TRIPLEX model for predicting Cinninghamia lanceolata and Pinus massoniana forest stand production in Hunan Province, southern China. Ecol. Modell. 2013, 250, 58–71. [Google Scholar] [CrossRef]
- Landsberg, J.; Sands, P. Physiological Ecology of Forest Production: Principles, Processes and Models; Academic Press: London, UK, 2010. [Google Scholar]
- Lemos, C.C.Z. Aprimoramentos, Teste e Uso do Modelo 3-PG em Plantios Clonais de Eucalyptus no Nordeste do Estado de São Paulo. Ph.D. Thesis, Universidade de São Paulo, Piracibaba, Brasil, 2012. [Google Scholar]
- Nolè, A.; Law, B.E.; Magnani, F.; Matteucci, G.; Ferrara, A.; Ripullone, F.; Borghetti, M. Application of the 3-PGS model to assess carbon accumulation in forest ecosystems at a regional level. Can. J. For. Res. 2009, 39, 1647–1661. [Google Scholar] [CrossRef]
- Gonzalez-Benecke, C.A.; Jokela, E.J.; Cropper, W.P., Jr.; Bracho, R.; Leduc, D.J. Parameterisation of the 3-PG model for Pinus elliottii stands using alternative methods to estimate fertility rating, biomass partitioning and canopy closure. For. Ecol. Manag. 2014, 327, 55–75. [Google Scholar] [CrossRef]
- Valente, M.L.; Reichert, J.M.; Cavalcante, R.B.L.; Minella, J.P.G.; Evrard, O.; Srinivasan, R. Afforestation of degraded grasslands reduces sediment transport and may contribute to streamflow regulation in small catchments in the short-run. Catena 2021, 204, 105371. [Google Scholar] [CrossRef]
- Ebling, E.D.; Reichert, J.M.; Pelaez, J.J.Z.; Rodrigues, M.F.; Valente, M.L.; Cavalcante, R.B.L.; Reggiani, P.; Srinivasan, R. Event-based hydrology and sedimentation in paired watersheds under commercial eucalyptus and grasslands in the Brazilian Pampa biome. Int. Soil Water Conserv. Res. 2021, 9, 180–194. [Google Scholar] [CrossRef]
- Stape, J.L.; Binkley, D.; Ryan, M.G. Eucalyptus production and the supply use and the efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For. Ecol. Manag. 2004, 193, 17–31. [Google Scholar] [CrossRef]
- Almeida, A.C.; Siggins, A.; Batista, T.R.; Beadle, C.; Fonseca, S.; Loos, R. Mapping the effect of spatial and temporal variation in climate and soils on Eucalyptus plantation production with 3-PG, a process-based growth model. For. Ecol. Manag. 2010, 259, 1730–1740. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Goncalves, J.L.M.; Sparovek, G. Koppen’s climate classification map for Brazil. Meteorol. Zeitschrif 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Moreno, J.A. Clima do Rio Grande do Sul; Secretaria da Agricultura: Porto Alegre, Brazil, 1961.
- IUSS Working Group WRB. World Reference Base for Soil Resources 2015; FAO: Rome, Italy, 2015. [Google Scholar]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Cunha, T.J.F.; Oliveira, J.B. Sistema Brasileiro de Classificação de Solos; Embrapa: Brasília, Brazil, 2013. [Google Scholar]
- Stape, J.L.; Binkley, D.; Ryan, M.G. Production and carbon allocation in a clonal Eucalyptus plantation with water and nutrient manipulations. For. Ecol. Manag. 2008, 255, 920–930. [Google Scholar] [CrossRef]
- Leite, F.P.; Silva, I.R.; Novais, R.F.; Barros, N.F.; Neves, J.C.L.; Villani, E.M.A. Nutrient relations during a eucalyptus cycle at different population densities. Rev. Bras. Cienc. Solo 2011, 35, 949–959. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.B.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Laclau, J.P.; Arnaud, M.; Bouillet, J.P.; Ranger, J. Spatial distribution of Eucalyptus roots in a deep sandy soil in the Congo: Relationships with the ability of the stand to take up water and nutrients. Tree Physiol. 2001, 21, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Christina, M.; Laclau, J.P.; Goncalves, J.L.M.; Jourdan, C.; Nouvellon, Y.; Bouillet, J.P. Almost symmetrical vertical growth rates above and below ground in one of the world’s most productive forest. Ecosphere 2011, 2, 27–30. [Google Scholar] [CrossRef]
- Stape, J.L.; Binkley, D.; Ryan, M.G.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.A.; et al. The Brazilian Eucalyptus Potential Productivity Project: Influence of water, nutrients and stand uniformity on wood production. For. Ecol. Manag. 2010, 259, 1684–1694. [Google Scholar] [CrossRef]
- Baesso, R.C.E. Modelagem do Comportamento Ecofisiológico de Plantios de Eucalipto Submetidos a Aumentos na Concentração de CO2. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2011. [Google Scholar]
- Klippel, V.H. Modelagem Ecofisiológica de Cultivos de Eucalipto em Regiões Subtropicais do Brasil. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2015. [Google Scholar]
- Suzuki, L.E.A.S.; Reichert, J.M.; Albuquerque, J.A.; Reinert, D.J.; Kaiser, D.R. Dispersion and flocculation of Vertisols, Alfisols and Oxisols in Southern Brazil. Geoderma Reg. 2015, 5, 64–70. [Google Scholar] [CrossRef]
- Gubiani, P.I.; Reichert, J.M.; Campbell, C.; Reinert, D.J.; Gelain, N.S. Assessing errors and accuracy in dew-point potentiometer and pressure plate extractor measurements. Soil Sci. Soc. Am. J. 2013, 77, 19–24. [Google Scholar] [CrossRef]
- Sands, P.J. Adaptation of 3-PG to Novel Species: Guidelines for Data Collection and Parameter Assignment; Cooperative Research Center for Sustainable Production Forestry: Hobart, Australia, 2004. [Google Scholar]
- Moriasi, D.N.; Arnold, J.G.; Van Liew, M.W.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans. Am. Soc. Agric. Biol. Eng. 2007, 50, 885–900. [Google Scholar] [CrossRef]
- Willmott, C.J. On the validation of models. Phys. Geogr. 1981, 2, 184–194. [Google Scholar] [CrossRef]
- Londero, E.K.; Schumacher, M.V.; Szymczak, D.A.; Araújo, E.F. Calibração do modelo 3-PG para Eucalyptus saligna Smith na Região de Guaíba—RS. Cienc. Florest. 2015, 25, 293–305. [Google Scholar] [CrossRef]
- Inkotte, J.; Mafra, A.L.; Rios, P.D.A. Deposição de serapilheira em reflorestamentos de eucalipto e florestas nativas nas regiões Planalto e Oeste do Estado de Santa Catarina. Sci. For. 2015, 43, 261–270. [Google Scholar]
- Eyles, A.; Worledge, D.; Sands, P.; Ottenschlaeger, M.L.; Paterson, S.C.; Mendham, D.; O’Grady, A.P. Ecophysiological responses of a young blue gum (Eucalyptus globulus) plantation to weed control. Tree Physiol. 2012, 32, 1008–1020. [Google Scholar] [CrossRef]
- England, J.R.; Attiwill, P.M. Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Muell. Trees Struct. Funct. 2006, 20, 79–90. [Google Scholar] [CrossRef]
- Raich, J.W.; Clark, D.A.; Schwendenmann, L.; Wood, T.E. Aboveground tree growth varies with belowground carbon allocation in a tropical rainforest environment. PLoS ONE 2014, 9, e100275. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zheng, X.; Ren, Y. Parameter optimization of the 3PG model based on sensitivity analysis and a Bayesian method. Forests 2020, 11, 1369. [Google Scholar] [CrossRef]
- Landsberg, J.J.; Johnsen, K.H.; Albaugh, T.J.; Allen, H.L.; McKeand, S.E. Applying 3-PG, a simple process-based model designed to produce practical results, to data from loblolly pine experiments. For. Sci. 2001, 47, 43–51. [Google Scholar] [CrossRef]
- Vaz, C.M.P.; Freitas Iossi, M.; Mendonça Naime, J.; Macedo, A.; Reichert, J.M.; Reinert, D.J.; Cooper, M. Validation of the Arya and Paris water retention model for Brazilian soils. Soil Sci. Soc. Am. J. 2005, 69, 577–583. [Google Scholar] [CrossRef]
- Reichert, J.M.; Albuquerque, J.A.; Kaiser, D.R.; Reinert, D.J.; Urach, F.L.; Carlesso, R. Estimation of water retention and availability for Rio Grande do Sul soils. Rev. Bras. Cienc. Solo 2009, 33, 1547–1560. [Google Scholar] [CrossRef]
- Reichert, J.M.; Albuquerque, J.A.; Solano Peraza, J.E.; Costa, A. Estimating water retention and availability in cultivated soils of southern Brazil. Geoderma Reg. 2020, 21, e00277. [Google Scholar] [CrossRef]
- Guimarães, D.P.; Guenther, G.; Silva, C.; Sans, L.M.A.; Leite, F.P. Uso do modelo de crescimento 3-PG para o zoneamento do potencial produtivo do eucalipto no estado de Minas Gerais. Rev. Bras. Agrometeorol. 2006, 15, 192–197. [Google Scholar]
- Rascon, N.J.L. Modelagem Ecofisiológica do Desenvolvimento do Eucalipto na Amazônia. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2012. [Google Scholar]
- Almeida, A.C. Needs and opportunities for using a process-based productivity model as a practical tool in Eucalyptus plantations. For. Ecol. Manag. 2004, 193, 167–177. [Google Scholar] [CrossRef]
- Ferreira, D.H.A.A.; Leles, P.S.S.; Machado, E.C.; Abreu, A.H.M.; Abilio, F.M. Crescimento de clone de Eucalyptus urophylla x E. grandis em diferentes espaçamentos. Floresta 2014, 44, 431–440. [Google Scholar] [CrossRef]
- Rocha, J.H.T. Reflexos do Manejo de Resíduos Florestais na Produtividade, Nutrição e Fertilidade do Solo em Plantações de Eucalyptus grandis. Ph.D. Thesis, Universidade de São Paulo, Piracicaba, Brazil, 2014. [Google Scholar]
- Fontan, I.C.I.; Reis, G.G.; Reis, M.G.F.; Leite, H.G.; Monte, M.A.; Ramos, D.C.; Souza, F.C. Growth of pruned eucalypt clone in an agroforestry system in southeastern Brazil. Agrofor. Syst. 2011, 83, 121–131. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; Gonçalves, J.L.M.; Gava, J.L.; Godinho, T.O.; Melo, E.A.S.C.; Bazani, J.H.; Hubner, A.; Junior, J.C.A.; Wichert, M.P. Forest residue maintenance increased the wood productivity of a Eucalyptus plantation over two short rotations. For. Ecol. Manag. 2016, 379, 1–10. [Google Scholar] [CrossRef]
- Machado, R.R.; Conceição, S.V.; Leite, H.G.; Souza, A.L.; Wolff, E. Evaluation of forest growth and carbon stock in forestry projects by system dynamics. J. Clean. Prod. 2015, 96, 520–530. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Baker, T.G. Interactive effects of simultaneously applied thinning, pruning and fertilizer application treatments on growth, biomass production and crown architecture in a young Eucalyptus nitens plantation. For. Ecol. Manag. 2012, 267, 104–116. [Google Scholar] [CrossRef]
- Schumacher, M.V.; Witschoreck, R.; Calil, F.N. Biomassa em povoamentos de Eucalyptus spp. de pequenas propriedades rurais de Vera Cruz, RS. Cienc. Florest. 2011, 21, 17–22. [Google Scholar] [CrossRef]
- Reichert, J.M.; Prevedello, J.; Gubiani, P.I.; Vogelmann, E.S.; Reinert, D.J.; Consensa, C.O.B.; Soares, J.C.W.; Srinivasan, R. Eucalyptus tree stockings effect on water balance and use efficiency in subtropical sandy soil. For. Ecol. Manag. 2021, 497, 119473. [Google Scholar] [CrossRef]
- Gonçalves, J.L.D.M.; Alvares, C.A.; Gonçalves, T.D. Mapeamento de solos e da produtividade de plantações de Eucalyptus grandis, com uso de sistema de informação geográfica. Sci. For. 2012, 40, 187–201. [Google Scholar]
- Gonçalves, J.L.D.M.; Alvares, C.A.; Higa, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; Ferraz, S.F.B.; Lima, W.P.; Brancalion, P.H.S.; Hubnera, A.; et al. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Stape, J.L.; Laclau, J.P.; Bouillet, J.P.; Ranger, J. Assessing the effects of early silvicultural management on long-term site productivity of fast-growing eucalypt plantations: The Brazilian experience. South. For. 2008, 70, 105–118. [Google Scholar] [CrossRef]
- Laclau, J.P.; Ranger, J.; Gonçalves, J.L.M.; Maquère, V.; Krusche, A.V.; M’Bouf, A.T.; Nouvellon, Y.; Saint-Andre, L.; Bouillet, J.P.; Piccolo, M.C.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations. Main features shown by intensive monitoring in Congo and Brazil. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Alaoui, A.; Lipiec, J.; Gerke, H.H. A review of the changes in the soil pore system due to soil deformation. Soil Tillage Res. 2011, 115–116, 1–15. [Google Scholar] [CrossRef]
- Balieiro, F.C.; Oliveira, W.C.; Pereira, M.G.; Anjos, L.H.C.; Piccolo, M.C.; Jaccoud, C.F. Fertilidade e carbono do solo e uso da água pelo eucalipto numa topossequência em Seropédica, RJ. Rev. Árvore 2008, 32, 153–162. [Google Scholar] [CrossRef]
- Reichert, J.M.; Corcini, A.L.; Awe, G.O.; Reinert, D.J.; Albuquerque, J.A.; García Gallarreta, C.C.; Docampo, R. Onion-forage cropping systems on a Vertic Argiudoll in Uruguay: Onion yield and soil organic matter, aggregation, porosity and permeability. Soil Tillage Res. 2022, 216, 105229. [Google Scholar] [CrossRef]
- Ferreto, D.O.C.; Reichert, J.M.; Lopes Cavalcante, R.B.; Srinivasan, R. Water budget fluxes in catchments under grassland and Eucalyptus plantations of different ages. Can. J. For. Res. 2021, 51, 513–523. [Google Scholar] [CrossRef]
- Horn, D.; Ernani, P.R.; Sangoi, L.; Schweitzer, C.; Cassol, P.C. Parâmetros cinéticos e morfológicos da absorção de nutrientes em cultivares de milho com variabilidade genética contrastante. Rev. Bras. Cienc. Solo 2006, 30, 77–85. [Google Scholar] [CrossRef]
- Chmura, D.J.; Rahman, M.S.; Tjoelker, M.G. Crown structure and biomass allocation patterns modulate aboveground productivity in young loblolly pine and slash pine. For. Ecol. Manag. 2007, 243, 219–230. [Google Scholar] [CrossRef]
- Reis, G.G.; Reis, M.D.G.F.; Fontan, I.D.C.I.; Monte, M.A.; Gomes, A.N.; Oliveira, C.H.R. Crescimento de raízes e da parte aérea de clones de híbridos de Eucalyptus grandis × Eucalyptus urophylla e de Eucalyptus camaldulensis × Eucalyptus spp submetidos a dois regimes de irrigação no campo. Rev. Árvore 2006, 30, 921–931. [Google Scholar] [CrossRef]
- Epron, D.; Laclau, J.P.; Almeida, J.C.R.; Gonçalves, J.L.M.; Ponton, S.; Sette Jr, C.R.; Delgado-Rojas, J.S.; Bouillet, J.P.; Nouvellon, Y. Do changes in carbon allocation account for the growth response to potassium and sodium applications in tropical Eucalyptus plantations? Tree Physiol. 2011, 32, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Zewdie, M.; Olsson, M.; Verwijst, T. Above-ground biomass production and allometric relations of Eucalyptus globulus Labill. coppice plantations along a chronosequence in the central highlands of Ethiopia. Biomass Bioenerg. 2009, 33, 421–428. [Google Scholar] [CrossRef]
- Bouillet, J.P.; Laclau, J.P.; Gonçalves, J.L.M.; Voigtlaender, M.; Gavad, J.L.; Leite, F.P.; Hakamadaf, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Santos, F.M.; Balieiro, F.C.; Ataíde, D.H.S.; Diniz, A.R.; Chaer, G.M. Dynamics of aboveground biomass accumulation in monospecific and mixed-species plantations of Eucalyptus and Acacia on a Brazilian sandy soil. For. Ecol. Manag. 2016, 363, 86–97. [Google Scholar] [CrossRef]
- Paixão, F.A.; Soares, C.P.B.; Jacovine, L.A.G.; Silva, M.L. ; Leite, H.G.; Silva, G.F. Quantificação do estoque de carbono e avaliação econômica de diferentes alternativas de manejo em um plantio de eucalipto. Rev. Árvore 2006, 30, 411–420. [Google Scholar] [CrossRef]
- Muñoz, F.; Rubilar, R.; Espinosa, M.; Cancino, J.; Toro, J.; Herrera, M. The effect of pruning and thinning on above ground aerial biomass of Eucalyptus nitens (Deane & Maiden) Maiden. For. Ecol. Manag. 2008, 255, 365–373. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Soares, C.P.B.; Fehrmann, L.; Jacovine, L.A.G.; Von Gadow, K. Aboveground and belowground biomass and carbon estimates for clonal eucalyptus trees in southeast Brazil. Rev. Árvore 2015, 39, 353–363. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuri, C.; Van Noordwijk, M.; Neufeldt, H. Allometry and partitioning of above- and below-ground biomass in farmed eucalyptus species dominant in Western Kenyan agricultural landscapes. Biomass Bioenerg. 2013, 55, 276–284. [Google Scholar] [CrossRef]
- Pinkard, E.; Neilsen, W. Crown and stand characteristics of Eucalyptus nitens in response to initial spacing: Implications for thinning. For. Ecol. Manag. 2003, 172, 215–227. [Google Scholar] [CrossRef]
- Reichert, J.M.; Bervald, C.M.P.; Rodrigues, M.F.; Kato, O.R.; Reinert, D.J. Mechanized land preparation in eastern Amazon in fire-free forest-based fallow systems as alternatives to slash-and-burn practices: Hydraulic and mechanical soil properties. Agric. Ecosyst. Environ. 2014, 192, 47–60. [Google Scholar] [CrossRef]
- Reichert, J.M.; Rodrigues, M.F.; Bervald, C.M.P.; Kato, O.R. Fire-free fallow management by mechanized chopping of biomass for sustainable agriculture in eastern Amazon: Effects on soil compactness, porosity, and water retention and availability. Land Degrad. Dev. 2016, 27, 1403–1412. [Google Scholar] [CrossRef]
- Reichert, J.M.; Amado, T.J.C.; Reinert, D.J.; Rodrigues, M.F.; Suzuki, L.E.A.S. Land use effects on subtropical, sandy soil under sandyzation/desertification processes. Agric. Ecosyst. Environ. 2016, 233, 370–380. [Google Scholar] [CrossRef]
- Viera, M.; Schumacher, M.V.; Araújo, E.F.; Corrêa, R.S.; Caldeira, M.V.W. Deposição de serapilheira e nutrientes em plantio de Eucalyptus urophylla × E. globulus. Flor. Amb. 2014, 21, 327–338. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Name | Units | Value |
|---|---|---|---|
| Foliage: stem partitioning ratio @ D = 2 cm | pFS2 | - | 0.81 |
| Foliage: stem partitioning ratio @ D = 20 cm | pFS20 | - | 0.22 |
| Constant in the stem mass vs. diameter relationship | aS | - | 0.04351 |
| Power in the stem mass vs. diameter relationship | nS | - | 2.7863 |
| Maximum fraction of NPP to roots | pRx | - | 0.25 |
| Minimum fraction of NPP to roots | pRn | - | 0.14 |
| Maximum litterfall rate | ℽFx | 1/month | 0.25 |
| Litterfall rate at t = 0 | ℽF0 | 1/month | 0.001 |
| Age at which litterfall rate has median value | tℽF | months | 12 |
| Average monthly root turnover rate | ℽR | 1/month | 0.025 |
| Temperature modifier (fT) | |||
| Minimum temperature for growth | Tmin | deg. C | −2 |
| Optimum temperature for growth | Topt | deg. C | 25 |
| Maximum temperature for growth | Tmax | deg. C | 34 |
| Frost modifier (fFRost) | |||
| Days production lost per frost day | kF | days | 2 |
| Soil water modifier (fSW) | |||
| Moisture ratio deficit for fq = 0.5 | SWconst | - | 0.5 |
| Power of moisture ratio deficit | SWpower | - | 5 |
| Atmospheric CO2 modifier (fCO2) | |||
| Assimilation enhancement factor at 700 ppm | fCalpha700 | - | 1.4 |
| Canopy conductance enhancement factor at 700 ppm | fCg700 | - | 0.7 |
| Fertility effects | |||
| Value of ‘m’ when FR = 0 | m0 | - | 0 |
| Value of ‘fNutr’ when FR = 0 | fN0 | - | 0.5 |
| Power of (1-FR) in ‘fNutr’ | fNn | - | 1 |
| Maximum stand age used in age modifier | MaxAge | years | 9 |
| Power of relative age in function for fAge | nAge | - | 4 |
| Relative age to give fAge = 0.5 | rAge | - | 0.95 |
| Mortality rate for large t | ℽNx | %/year | 0 |
| Seedling mortality rate (t = 0) | ℽN0 | %/year | 0 |
| Age at which mortality rate has median value | tℽN | years | 2 |
| Shape of mortality response | nℽN | - | 1 |
| Max. stem mass per tree @ 1000 trees/hectare | wSx1000 | kg/tree | 300 |
| Power in self-thinning rule | thinPower | - | 1.5 |
| Fraction mean single-tree foliage biomass lost per dead tree | mF | - | 0 |
| Fraction mean single-tree root biomass lost per dead tree | mR | - | 0.2 |
| Fraction mean single-tree stem biomass lost per dead tree | mS | - | 0.2 |
| Specific leaf area at age 0 | SLA0 | m2/kg | 5.6 |
| Specific leaf area for mature leaves | SLA1 | m2/kg | 7.6 |
| Age at which specific leaf area = (SLA0 + SLA1)/2 | tSLA | years | 4.5 |
| Extinction coefficient for absorption of PAR by canopy | k | - | 0.5 |
| Age at canopy cover | fullCanAge | years | 5 |
| Maximum proportion rainfall evaporated canopy | MaxIntcptn | - | 0.15 |
| LAI for maximum rainfall interception | LAImaxIntcptn | - | 3.33 |
| Canopy quantum efficiency | alpha | molC/molPAR | 0.068 |
| Ratio NPP/GPP | Y | - | 0.50 |
| Minimum canopy conductance | MinCond | m/s | 0 |
| Maximum canopy conductance | MaxCond | m/s | 0.032 |
| LAI for maximum canopy conductance | LAIgcx | - | 2.96 |
| Defines stomatal response to VPD | CoeffCond | 1/mBar | 0.0324 |
| Canopy boundary layer conductance | BLcond | m/s | 0.2 |
| Branch and bark fraction at age 0 | fracBB0 | - | 0.29 |
| Branch and bark fraction for mature stands | fracBB1 | - | 0.15 |
| Age at which fracBB = (fracBB0 + fracBB1)/2 | tBB | years | 3.5 |
| Minimum basic density—for young trees | rhoMin | t/m3 | 0.421 |
| Maximum basic density—for older trees | rhoMax | t/m3 | 0.458 |
| Age at which rho = (rhoMin + rhoMax)/2 | tRho | years | 4 |
| Constant in the stem height relationship | aH | - | 0.3561 |
| Power of DBH in the stem height relationship | nHB | - | 1.44211 |
| Power of stocking in the stem height relationship | nHN | - | 0 |
| Constant in the stem volume relationship | aV | - | 0.1061 |
| Power of DBH in the stem volume relationship | nVB | - | 2.752012 |
| Power of stocking in stem volume relationship | nVN | - | 0 |
| Intercept of net v. solar radiation relationship | Qa | W/m2 | −90 |
| Slope of net v. solar radiation relationship | Qb | - | 0.8 |
| Molecular weight of dry matter | gDM_mol | gDM/mol | 24 |
| Conversion of solar radiation to PAR | molPAR_MJ | mol/MJ | 2.3 |
| Statistical Indicators | ACRI | ARE | ACRI | ARE | ACRI | ARE | ACRI | ARE |
|---|---|---|---|---|---|---|---|---|
| h | dbh | V | WB | |||||
| NSE | 0.90 | 0.91 | 0.80 | 0.71 | 0.82 | 0.81 | 0.83 | 0.83 |
| PBIAS (%) | −2.25 | 2.96 | −1.50 | 0.86 | −11.56 | −9.00 | −11.46 | −8.77 |
| Willmott | 0.92 | 0.89 | 0.88 | 0.75 | 0.85 | 0.80 | 0.85 | 0.83 |
| r | 0.97 | 0.98 | 0.95 | 0.87 | 0.97 | 0.95 | 0.96 | 0.96 |
| c | 0.89 | 0.87 | 0.84 | 0.65 | 0.82 | 0.77 | 0.82 | 0.80 |
| Statistical Indicator | h | dbh | V | WB |
|---|---|---|---|---|
| NSE | 0.94 | −0.93 | 0.90 | 0.96 |
| PBIAS (%) | 2.00 | 7.20 | 5.00 | 4.00 |
| Willmott | 0.97 | 0.67 | 0.96 | 0.96 |
| r | 0.99 | 0.96 | 0.99 | 0.99 |
| Area | H * (m) | Dbh * (m) | VWB * (m3 ha−1) | WBT * (Mg ha−1) |
|---|---|---|---|---|
| ACRI33 | 14.2 b (9%) | 0.131 a (16%) | 80.6 a (42%) | 27.5 a (32%) |
| ARE33 | 13.5 a (17%) | 0.130 a (18%) | 70.8 a (57%) | 27.4 a (42%) |
| ACRI58 | 20.9 b (19%) | 0.160 a (12%) | 193.8 a (40%) | 78.2 a (41%) |
| ARE58 | 19.6 a (15%) | 0.160 a (10%) | 195.9 a (32%) | 79.6 a (35%) |
| ACRI89 | 27.3 b (10%) | 0.189 b (17%) | 373.9 b (32%) | 177.2 b (35%) |
| ARE89 | 23.5 a (21%) | 0.165 a (28%) | 272.3 a (55%) | 130.0 a (56%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalli, J.P.; Frank de Araújo, E.; Reichert, J.M. Eucalyptus Growth Responses to Soil Water Storage Capacity in Arenosols and Acrisols Soils: Wood and Biomass Stock Modelling. Sustainability 2022, 14, 12215. https://doi.org/10.3390/su141912215
Cavalli JP, Frank de Araújo E, Reichert JM. Eucalyptus Growth Responses to Soil Water Storage Capacity in Arenosols and Acrisols Soils: Wood and Biomass Stock Modelling. Sustainability. 2022; 14(19):12215. https://doi.org/10.3390/su141912215
Chicago/Turabian StyleCavalli, Jean Pierre, Elias Frank de Araújo, and José Miguel Reichert. 2022. "Eucalyptus Growth Responses to Soil Water Storage Capacity in Arenosols and Acrisols Soils: Wood and Biomass Stock Modelling" Sustainability 14, no. 19: 12215. https://doi.org/10.3390/su141912215
APA StyleCavalli, J. P., Frank de Araújo, E., & Reichert, J. M. (2022). Eucalyptus Growth Responses to Soil Water Storage Capacity in Arenosols and Acrisols Soils: Wood and Biomass Stock Modelling. Sustainability, 14(19), 12215. https://doi.org/10.3390/su141912215