


Pelvetia canaliculata as an Aquafeed Supplement for Gilthead Seabream Sparus aurata: A Biorefinery Approach for Seaweed Biomass Valorisation

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Algae Collection and Processing
2.3. Experimental Diets
2.4. Fish and Rearing Conditions
2.5. Growth Performance, Hepatosomatic and Viscerosomatic Index
2.6. Haematological Parameters
2.7. Oxidative Stress Analysis
2.8. Metabolic and Immune Parameters
2.9. Intestine Histology
2.10. Statistical Analysis
3. Results
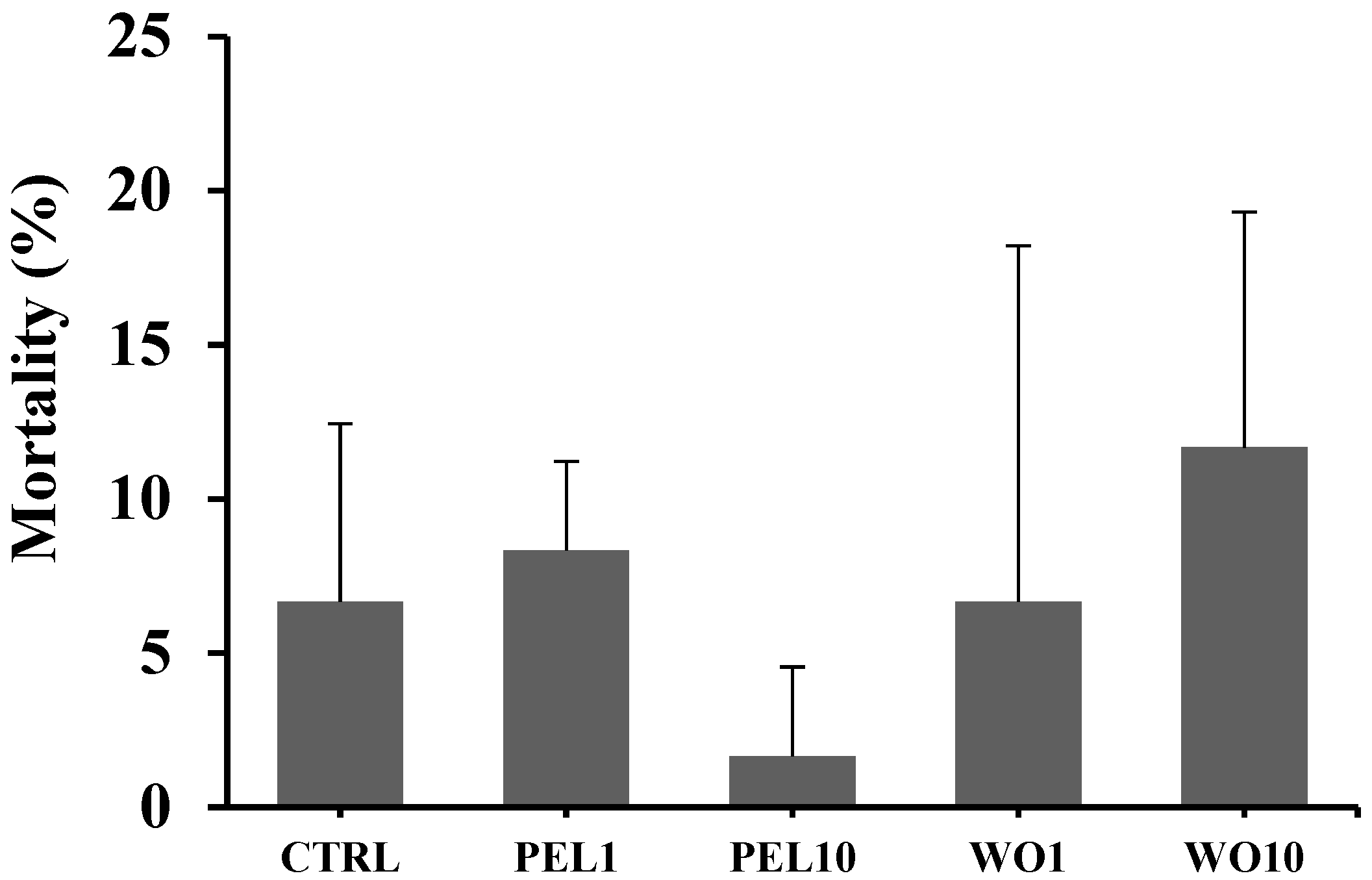
3.1. Growth Performance, Somatic Indexes, and Mortality
3.2. Haematological Parameters
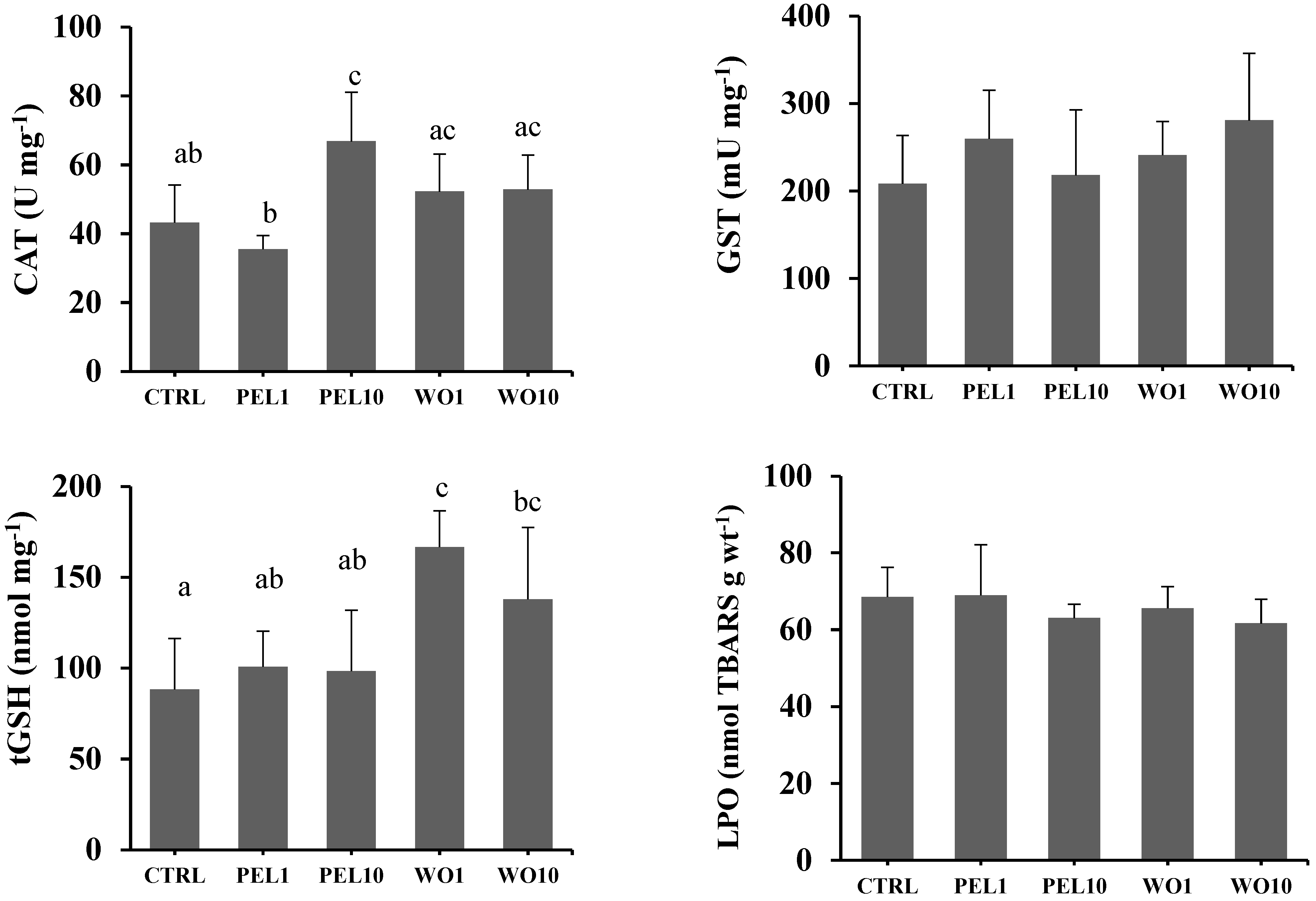
3.3. Oxidative Stress
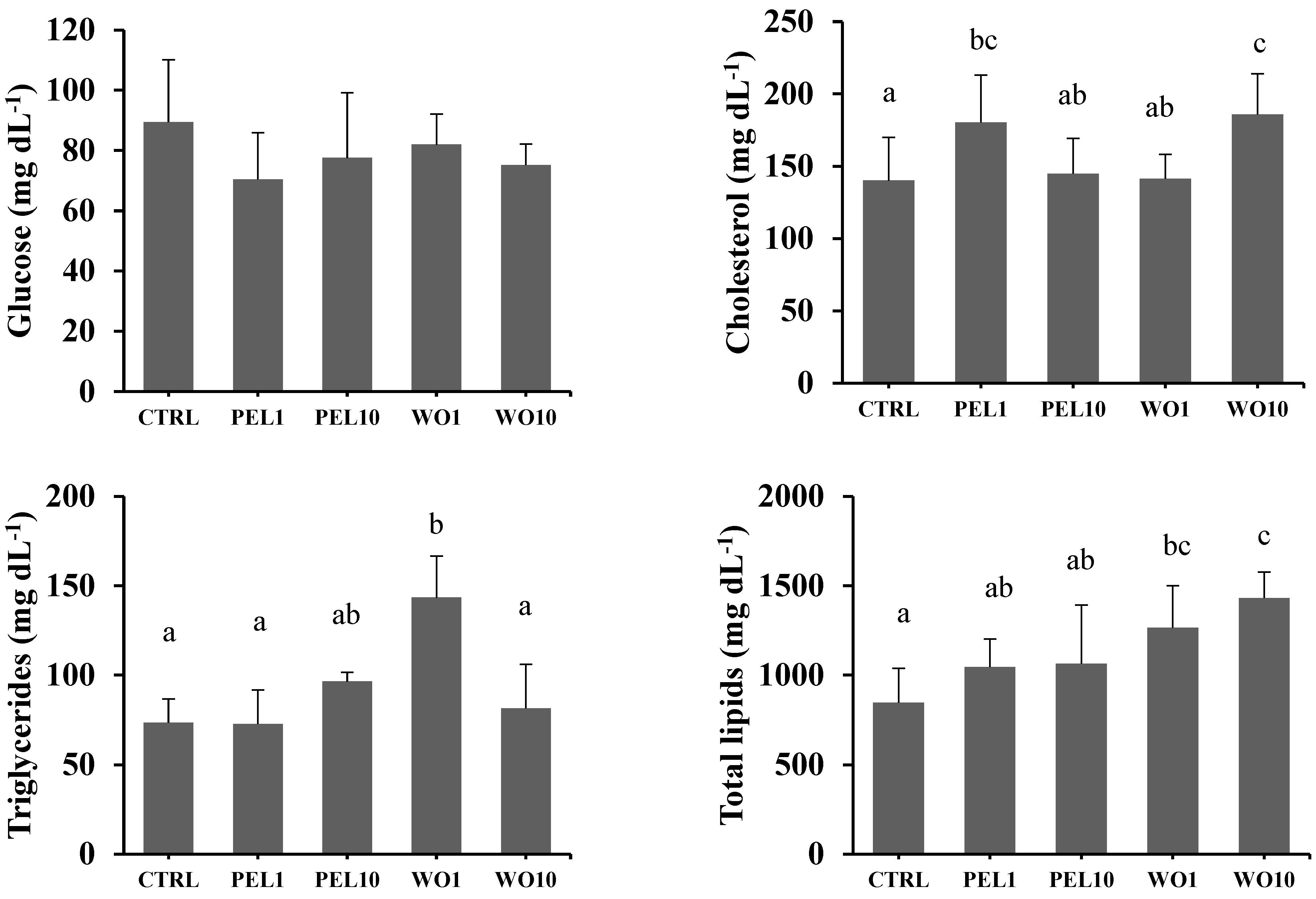
3.4. Metabolic Parameters
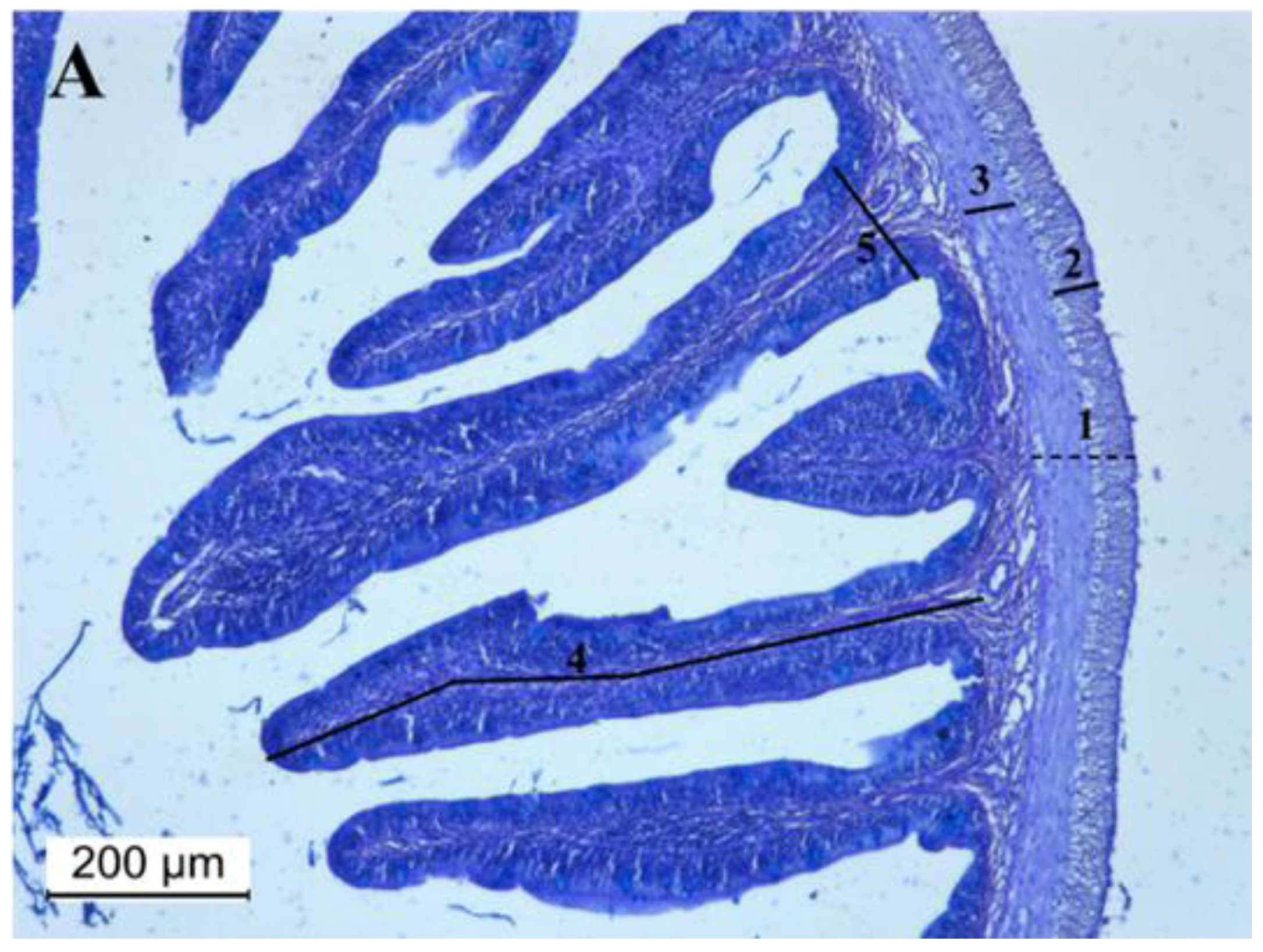
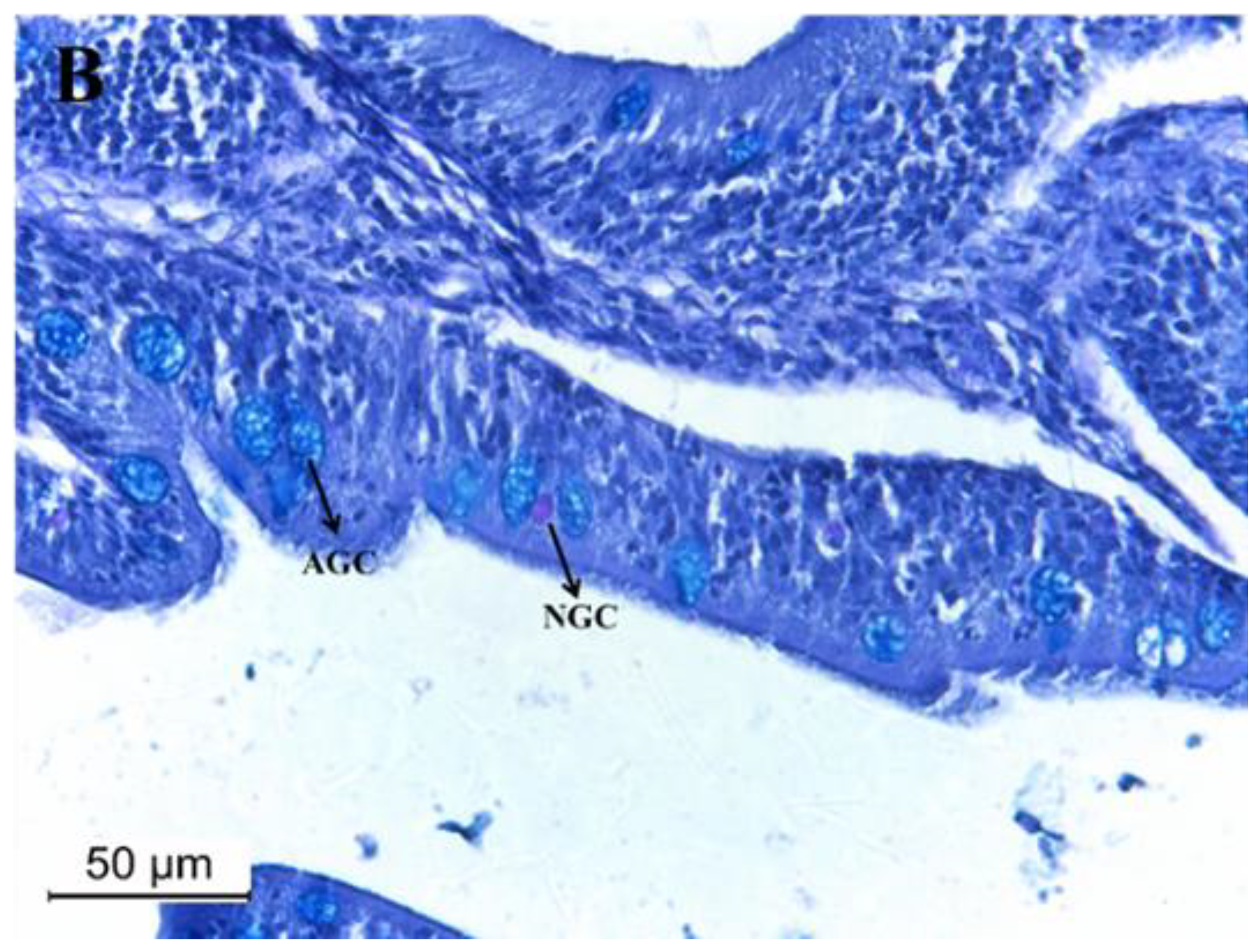
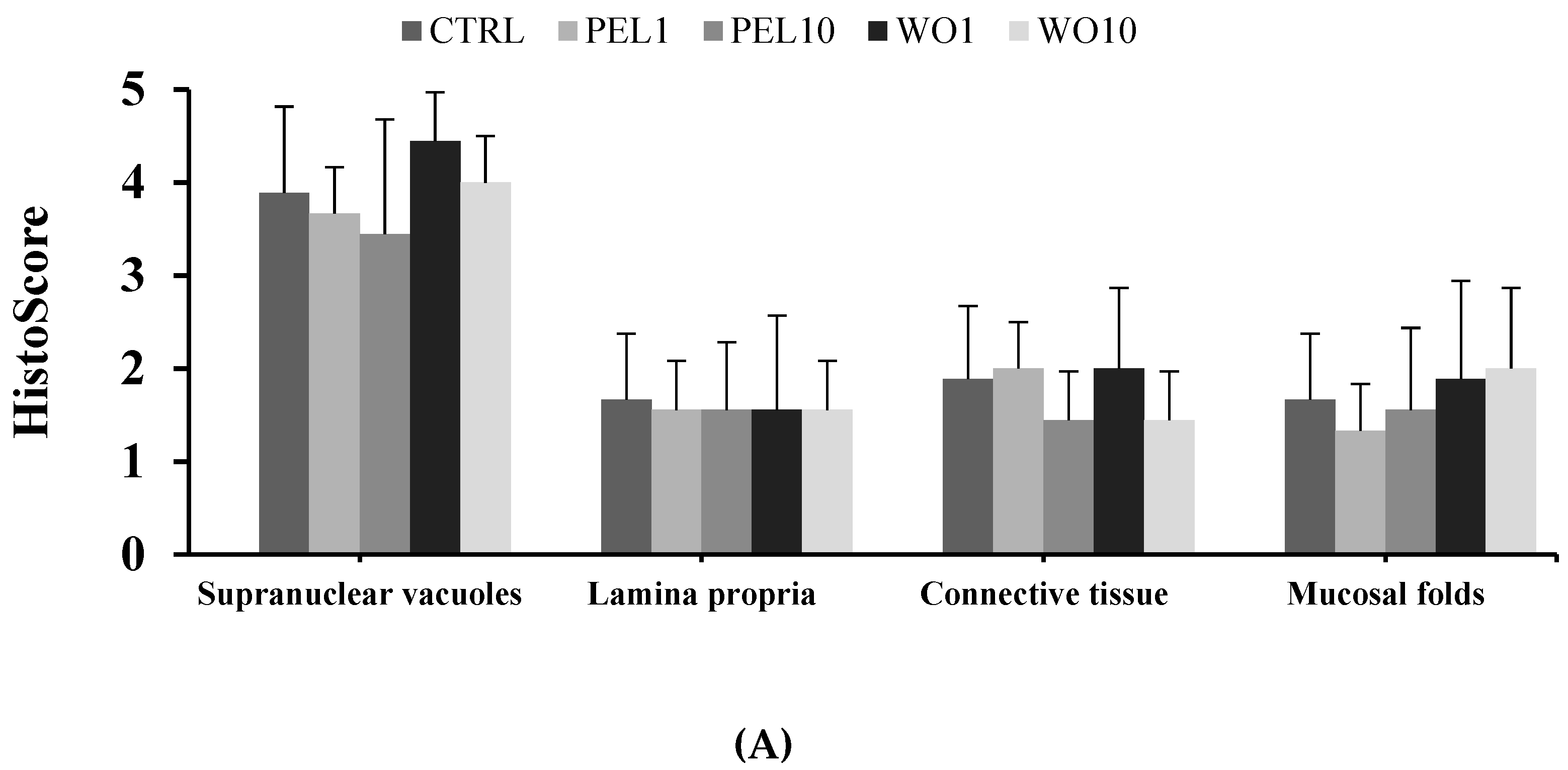
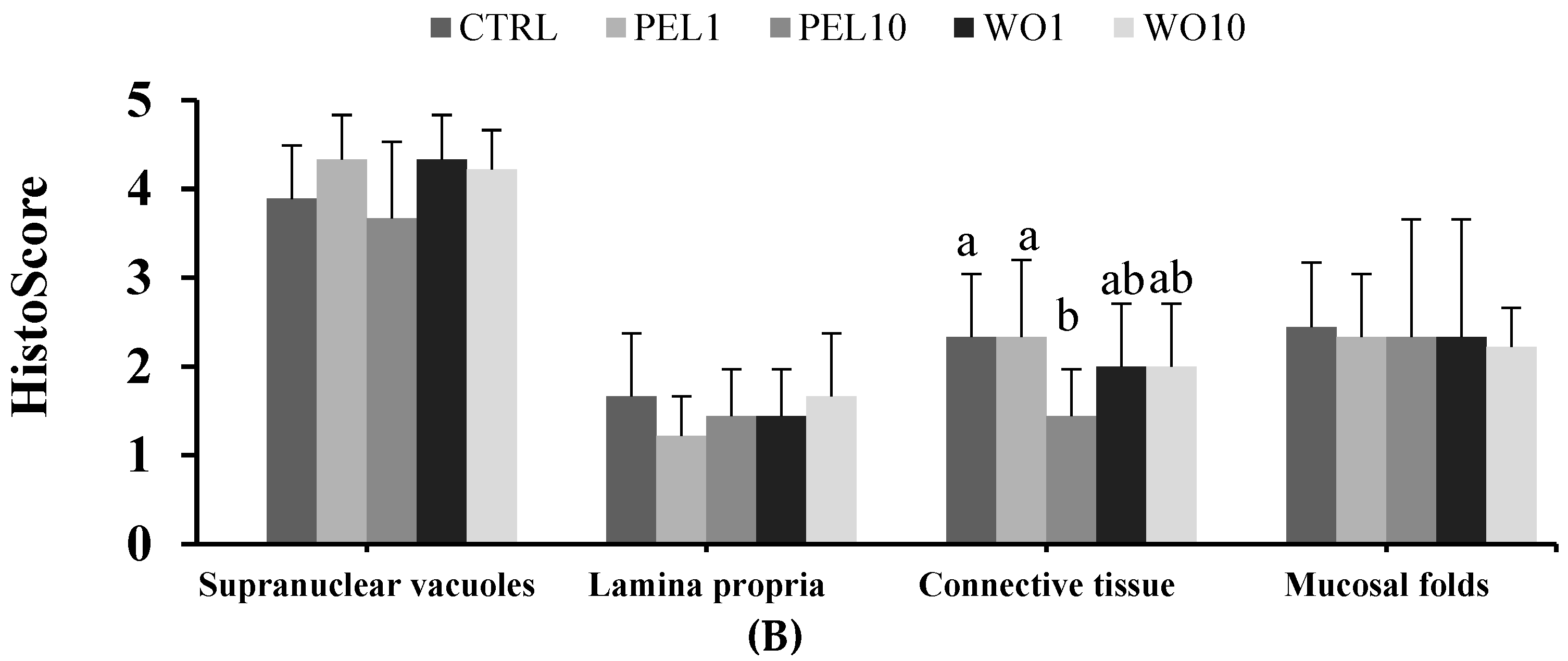
3.5. Intestine Histomorphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Fishery and Aquaculture Statistics. Glob. Aquac. Prod. 2021. Available online: www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 30 November 2021).
- Nagappan, S.; Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Mahata, C.; Al-Jabri, H.; Vatland, A.K.; Kumar, G. Potential of microalgae as a sustainable feed ingredient for aquaculture. J. Biotechnol. 2021, 341, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Teodósio, R.; Aragão, C.; Colen, R.; Carrilho, R.; Dias, J.; Engrola, S. A nutritional strategy to promote gilthead seabream performance under low temperatures. Aquaculture 2021, 537, 736494. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Batista, S.; Ribeiro, C.; Pereira, R.; Oliveira, B.; Garrido, I.; Baião, L.F.; Tulli, F.; Messina, M.; Pierre, R.; et al. Physical processing or supplementation of feeds with phytogenic compounds, alginate oligosaccharide or nucleotides as methods to improve the utilization of Gracilaria gracilis by juvenile European seabass (Dicentrarchus labrax). Aquaculture 2021, 530, 735914. [Google Scholar] [CrossRef]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed Potential in the Animal Feed: A Review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- Pereira, V.; Marques, A.; Gaivão, I.; Rego, A.; Abreu, H.; Pereira, R.; Santos, M.A.; Guilherme, S.; Pacheco, M. Marine macroalgae as a dietary source of genoprotection in gilthead seabream (Sparus aurata) against endogenous and exogenous challenges. Comp. Biochem. Physiol. C 2019, 219, 12–24. [Google Scholar] [CrossRef]
- Araújo, M.; Rema, P.; Sousa-Pinto, I.; Cunha, L.M.; Peixoto, M.J.; Pires, M.A.; Seixas, F.; Brotas, V.; Beltran, C.; Valente, L.M.P. Dietary inclusion of IMTA-cultivated Gracilaria vermiculophylla in rainbow trout (Oncorhynchus mykiss) diets: Effects on growth, intestinal morphology, tissue pigmentation, and immunological response. J. Appl. Phycol. 2016, 28, 679–689. [Google Scholar] [CrossRef]
- Passos, R.; Correia, A.P.; Ferreira, I.; Pires, P.; Pires, D.; Gomes, E.; do Carmo, B.; Santos, P.; Simões, M.; Afonso, C.; et al. Effect on health status and pathogen resistance of gilthead seabream (Sparus aurata) fed with diets supplemented with Gracilaria gracilis. Aquaculture 2021, 531, 735888. [Google Scholar] [CrossRef]
- Passos, R.; Correia, A.P.; Pires, D.; Pires, P.; Ferreira, I.; Simões, M.; do Carmo, B.; Santos, P.; Pombo, A.; Afonso, C.; et al. Potential use of macroalgae Gracilaria gracilis in diets for European seabass (Dicentrarchus labrax): Health benefits from a sustainable source. Fish Shellfish Immunol. 2021, 119, 105–113. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Salas-Leitón, E.; Pereira, L.F.; Queiroz, A.; Magalhães, F.; Pereira, R.; Abreu, H.; Reis, P.A.; Gonçalves, J.F.M.; Ozório, R.O.A. Role of dietary seaweed supplementation on growth performance, digestive capacity and immune and stress responsiveness in European seabass (Dicentrarchus labrax). Aquac. Rep. 2016, 3, 189–197. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Ferraz, R.; Magnoni, L.J.; Pereira, R.; Gonçalves, J.F.; Calduch-Giner, J.; Pérez-Sánchez, J.; Ozório, R.O.A. Protective effects of seaweed supplemented diet on antioxidant and immune responses in European seabass (Dicentrarchus labrax) subjected to bacterial infection. Sci. Rep. 2019, 9, 16134. [Google Scholar] [CrossRef] [Green Version]
- Vizcaíno, A.J.; Fumanal, M.; Sáez, M.I.; Martínez, T.F.; Moriñigo, M.A.; Fernández-Díaz, C.; Anguis, V.; Balebona, M.C.; Alarcón, F.J. Evaluation of Ulva ohnoi as functional dietary ingredient in juvenile Senegalese sole (Solea senegalensis): Effects on the structure and functionality of the intestinal mucosa. Algal Res. 2019, 42, 101608. [Google Scholar] [CrossRef]
- Leandro, A.; Pereira, L.; Gonçalves, A.M.M. Diverse Applications of Marine Macroalgae. Mar. Drugs 2020, 18, 17. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vaquero, M.; Ravindran, R.; Walsh, O.; O’Doherty, J.; Jaiswal, A.K.; Tiwari, B.K.; Rajauria, G. Evaluation of Ultrasound, Microwave, Ultrasound–Microwave, Hydrothermal and High Pressure Assisted Extraction Technologies for the Recovery of Phytochemicals and Antioxidants from Brown Macroalgae. Mar. Drugs 2021, 19, 309. [Google Scholar] [CrossRef]
- Gomez-Zavaglia, A.; Lage, M.A.P.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The Potential of Seaweeds as a Source of Functional Ingredients of Prebiotic and Antioxidant Value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yu, M. Role of Goblet Cells in Intestinal Barrier and Mucosal Immunity. J. Inflamm. Res. 2021, 14, 3171–3183. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Maneesh, A.; Makkar, F. Antioxidant Activity of Brown Seaweeds. J. Aquat. Food Prod, Technol. 2017, 26, 406–419. [Google Scholar] [CrossRef]
- Hupel, M.; Lecointre, C.; Meudec, A.; Poupart, N.; Gall, E.A. Comparison of photoprotective responses to UV radiation in the brown seaweed Pelvetia canaliculata and the marine angiosperm Salicornia ramosissima. J. Exp. Mar. Biol. Ecol. 2011, 401, 36–47. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Silva, A.; Rodrigues, C.; Garcia-Oliveira, P.; Lourenço-Lopes, C.; Silva, S.A.; Garcia-Perez, P.; Carvalho, A.P.; Domingues, V.F.; Barroso, M.F.; Delerue-Matos, C.; et al. Screening of Bioactive Properties in Brown Algae from the Northwest Iberian Peninsula. Foods 2021, 10, 1915. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Rai, D.K.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Enrichment of polyphenol contents and antioxidant activities of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.P.; Smyth, T.J. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2014, 10, 524–535. [Google Scholar] [CrossRef]
- Chojnacka, K.; Skrzypczak, D.; Mikula, K.; Witek-Krowiak, A.; Izydorczyk, G.; Kuligowski, K.; Bandrów, P.; Kułażýnski, M. Progress in sustainable technologies of leather wastes valorization as solutions for the circular economy. J. Clean. Prod. 2021, 313, 127902. [Google Scholar] [CrossRef]
- Hakim, M.M.; Patel, I.C. A review on phytoconstituents of marine brown algae. Future J. Pharm. Sic. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- Afonso, N.C.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Brown Macroalgae as Valuable Food Ingredients. Antioxidants 2019, 8, 365. [Google Scholar] [CrossRef]
- Mirabella, N.; Castellani, V.; Sala, S. Current options for the valorization of food manufacturing waste: A review. J. Clean. Prod. 2014, 65, 28–41. [Google Scholar] [CrossRef]
- Sousa, G.; Trifunovska, M.; Antunes, M.; Miranda, I.; Moldão, M.; Alves, V.; Vidrih, R.; Lopes, P.A.; Aparicio, L.; Neves, M.; et al. Optimization of Ultrasound-Assisted Extraction of Bioactive Compounds from Pelvetia canaliculata to Sunflower Oil. Foods 2021, 10, 1732. [Google Scholar] [CrossRef]
- Martins, M.; Soares, C.; Figueiredo, I.; Sousa, B.; Torres, A.C.; Sousa-Pinto, I.; Veiga, P.; Rubal, M.; Fidalgo, F. Fucoid macroalgae have distinct physiological mechanisms to face emersion and submersion periods in their southern limit of distribution. Plants 2021, 10, 1892. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef]
- Bird, R.P.; Draper, A.H. Comparative studies on different methods of malondihaldehyde determination. Methods Enzymol. 1984, 90, 105–110. [Google Scholar]
- Clairborne, A. Catalase activity. In Handbook of Methods of Oxygen Radical Research; Grenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Almeida, J.R.; Oliveira, C.; Gravato, C.; Guilhermino, L. Linking behavioural alterations with biomarkers responses in the European seabass Dicentrarchus labrax L. exposed to the organophosphate pesticide fenitrothion. Ecotoxicology 2010, 19, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Frasco, M.F.; Guilhermino, L. Effects of dimethoate and beta-naphthoflavone on selected biomarkers of Poecilia reticulata. Fish Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione-S-transferases, the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Baker, M.; Cemiglia, G.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]
- Rodrigues, A.C.M.; Gravato, C.; Quintaneiro, C.; Bordalo, M.D.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Energetic costs and biochemical biomarkers associated with esfenvalerate exposure in Sericostoma vittatum. Chemosphere 2017, 189, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Batista, S.; Medina, A.; Pires, M.A.; Moriñigo, M.A.; Sansuwan, K.; Fernandes, J.M.O.; Valente, L.M.P.; Ozório, R.O.A. Innate immune response, intestinal morphology and microbiota changes in Senegalese sole fed plant protein diets with probiotics or autolyzed yeast. Appl. Microbiol. Biotechnol. 2016, 100, 7223–7238. [Google Scholar] [CrossRef] [PubMed]
- Batista, S.; Pereira, R.; Oliveira, B.; Baião, L.F.; Flemming, J.; Tulli, F.; Messina, M.; Silva, J.L.; Abreu, H.; Valente, L.M.P. Exploring the potential of seaweed Gracilaria gracilis and microalga Nannochloropsis oceanica, single or blended, as natural dietary ingredients for European seabass Dicentrarchus labrax. J. Appl. Phycol. 2020, 32, 2041–2059. [Google Scholar] [CrossRef]
- Knudsen, D.; Uran, P.; Arnous, A.; Koppe, W.; Frøkiær, H. Saponin-containing subfractions of soybean molasses induce enteritis in the distal intestine of Atlantic salmon. J. Agric. Food Chem. 2007, 55, 2261–2267. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2018, 500, 237–242. [Google Scholar] [CrossRef]
- Fazio, F.; Saoca, C.; Casella, S.; Fortino, G.; Piccione, G. Relationship between blood parameters and biometric indices of Sparus aurata and Dicentrarchus labrax cultured in onshore tanks. Mar. Freshw. Behav. Physiol. 2015, 48, 289–296. [Google Scholar] [CrossRef]
- Jawad, L.A.; Al-Mukhtar, M.A.; Ahmed, H.K. The relationship between haematocrit and some biological parameters of the Indian shad, Tenualosa ilisha (family Clupeidae). Anim. Biodivers. Conserv. 2004, 27, 47–52. [Google Scholar]
- Magnoni, L.J.; Martos-Sitcha, J.A.; Queiroz, A.; Calduch-Giner, J.A.; Gonçalves, J.F.M.; Rocha, C.M.R.; Abreu, H.T.; Schrama, J.W.; Ozório, R.O.A.; Pérez-Sánchez, J. Dietary supplementation of heat-treated Gracilaria and Ulva seaweeds enhanced acute hypoxia tolerance in gilthead sea bream (Sparus aurata). Biol. Open 2017, 6, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leggatt, R.A.; Scheer, K.W.; Afonso, L.O.B.; Iwama, G.K. Triploid and diploid rainbow trout do not differ in their stress response to transportation. N. Am. J. Aquac. 2006, 68, 1–8. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Mendes, S.I.; Varela, J.L.; Ruiz-Jarabo, I.; Rico, R.; Figueroa, F.L.; Abdala, R.; Moriñigo, M.A.; Mancera, J.M.; Alarcón, F.J. Growth, tissue metabolites and digestive functionality in Sparus aurata juveniles fed different levels of macroalgae, Gracilaria cornea and Ulva rigida. Aquac. Res. 2016, 47, 3224–3238. [Google Scholar] [CrossRef]
- Guerreiro, I.; Magalhães, R.; Coutinho, F.; Couto, A.; Sousa, A.; Delerue-Matos, C.; Domingues, V.F.; Oliva-Teles, A.; Peres, H. Evaluation of the seaweeds Chondrus crispus and Ulva lactuca as functional ingredients in gilthead seabream (Sparus aurata). J. Appl. Phycol. 2019, 31, 2115–2124. [Google Scholar] [CrossRef]
- Basto-Silva, C.; Enes, P.; Oliva-Teles, A.; Balbuena-Pecino, S.; Navarro, I.; Capilla, E.; Guerreiro, I. Dietary protein source and protein/carbohydrate ratio affects appetite regulation-related genes expression in gilthead seabream (Sparus aurata). Aquaculture 2021, 533, 736142. [Google Scholar] [CrossRef]
- Silva-Brito, F.; Alexandrino, D.A.M.; Jia, Z.; Mo, Y.; Kijjoa, A.; Abreu, H.; Carvalho, M.F.; Ozório, R.; Magnoni, L. Fish performance, intestinal bacterial community, digestive function and skin and fillet attributes during cold storage of gilthead seabream (Sparus aurata) fed diets supplemented with Gracilaria by-products. Aquaculture 2021, 541, 736808. [Google Scholar] [CrossRef]
- Batista, S.; Ramos, M.A.; Cunha, S.; Barros, R.; Cristóvão, B.; Rema, P.; Pires, M.A.; Valente, L.M.P.; Ozório, R.O.A. Immune responses and gut morphology of Senegalese sole (Solea senegalensis, Kaup 1858) fed monospecies and multispecies probiotics. Aquac. Nutr. 2014, 21, 625–634. [Google Scholar] [CrossRef]
- Silva, D.M.; Valente, L.M.P.; Sousa-Pinto, I.; Pereira, R.; Pires, M.A.; Seixas, F.; Rema, P. Evaluation of IMTA-produced seaweeds (Gracilaria, Porphyra, and Ulva) as dietary ingredients in Nile tilapia, Oreochromis niloticus L., juveniles. Effects on growth performance and gut histology. J. Appl. Phycol. 2015, 27, 1671–1680. [Google Scholar] [CrossRef]
- Sotoudeh, E.; Mardani, F. Antioxidant-related parameters, digestive enzyme activity and intestinal morphology in rainbow trout (Oncorhynchus mykiss) fry fed graded levels of red seaweed, Gracilaria pygmaea. Aquac. Nutr. 2018, 24, 777–785. [Google Scholar] [CrossRef]
- Khojasteh, S.M.B. The morphology of the post-gastric alimentary canal in teleost fishes: A brief review. Int J. Aquat. Sci. 2012, 3, 71–88. [Google Scholar]
- Torrecillas, S.; Terova, G.; Makol, A.; Serradell, A.; Valdenegro, V.; Gini, E.; Izquierdo, M.; Acosta, F.; Montero, D. Dietary phytogenics and galactomannan oligosaccharides in low fish meal and fish oil-based diets for European sea bass (Dicentrarchus labrax) juveniles: Effects on gut health and implications on in vivo gut bacterial translocation. PLoS ONE 2019, 14, e0222063. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Couto, A.; Diógenes, A.F.; Corraze, G.; Panserat, S.; Serra, C.R.; Oliva-Teles, A. Vegetable oil and carbohydrate-rich diets marginally affected intestine histomorphology, digestive enzymes activities, and gut microbiota of gilthead sea bream juveniles. Fish Physiol. Biochem. 2019, 45, 681–695. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Composition | |
|---|---|
| Proteins (%) | 7.72 ± 0.13 |
| Lipids (%) | 5.12 ± 0.41 |
| Ash (%) | 21.40 ± 0.04 |
| Carbohydrates (%) | 65.76 ± 0.43 |
| Pigments | |
| Chlorophylls (pheophytin a /Kg) | 602 ± 30 |
| Carotenoids (mg β-carotene/Kg) | 236 ± 12 |
| Proteins (%) | 5.26 ± 0.09 |
| Lipids (%) | 35.27 ± 1.13 |
| Ash (%) | 14.60 ± 0.03 |
| Carbohydrates (%) | 44.86 ± 0.29 |
| CTRL | PEL1 | PEL10 | WO1 | WO10 | |
|---|---|---|---|---|---|
| Ingredients (%) | |||||
| Fishmeal | 20 | 20 | 20 | 20 | 20 |
| Fish protein hydrolysate | 5 | 5 | 5 | 5 | 5 |
| Fish gelatine | 2 | 2 | 2 | 2 | 2 |
| Poultry meal | 10 | 10 | 10 | 10 | 10 |
| Pea protein concentrate | 5 | 5 | 5 | 5 | 5 |
| Wheat gluten | 9.6 | 9.7 | 10.4 | 9.7 | 10.1 |
| Corn gluten meal | 6 | 6 | 6 | 6 | 6 |
| Soybean meal | 15 | 15 | 15 | 15 | 15 |
| Wheat meal | 15.5 | 14.4 | 5.1 | 14.7 | 8.4 |
| Vitamin and mineral premix | 1 | 1 | 1 | 1 | 1 |
| Monocalcium phosphate | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 |
| Fish oil | 6 | 6 | 6 | 6 | 6 |
| Soybean oil | 6 | 6 | 5.6 | 5.7 | 2.6 |
| Pelvetia powder | 1 | 10 | |||
| Pelvetia waste | 1 | 10 | |||
| Proximate composition (%) | |||||
| Protein | 48.9 ± 0.8 | 47.2 ± 0.7 | 48.6 ± 0.6 | 47.1 ± 2.0 | 47.9 ± 1.4 |
| Fat | 17.4 ± 0.2 | 17.0 ± 1.1 | 16.9 ± 0.6 | 17.6 ± 0.3 | 16.8 ± 0.4 |
| Ash | 6.33 ± 0.07 | 6.37 ± 0.08 | 8.22 ± 0.04 | 6.58 ± 0.05 | 7.73 ± 0.04 |
| Moisture | 7.08 ± 0.15 | 9.08 ± 0.11 | 7.97 ± 0.13 | 6.12 ± 0.05 | 6.49 ± 0.07 |
| Carbohydrates | 19.9 ± 0.6 | 19.9 ± 1.4 | 17.8 ± 0.3 | 22.1 ± 2.3 | 20.6 ± 1.1 |
| CTRL | PEL1 | PEL10 | WO1 | WO10 | |
|---|---|---|---|---|---|
| Growth | |||||
| Initial body weight (g) | 15.4 ± 3.5 | 15.6 ± 3.7 | 15.4 ± 3.4 | 15.5 ± 3.8 | 15.0 ± 3.3 |
| Final body weight (g) | 33 ± 8 | 34 ± 9 | 33 ±11 | 37 ±11 | 33 ± 9 |
| Weight gain (%) | 111 ± 3 | 119 ± 20 | 112 ± 5 | 144 ± 14 | 119 ± 14 |
| Specific growth rate (% day−1) | 0.69 ± 0.01 | 0.72 ± 0.08 | 0.70 ± 0.02 | 0.82 ± 0.05 | 0.72 ± 0.06 |
| Voluntary feed intake (% BW day −1) | 2.26 ± 0.16 | 2.05 ± 0.19 | 2.17 ± 0.09 | 2.16 ± 0.13 | 2.18 ± 0.02 |
| Daily growth index (BW day−1) | 1.60 ± 0.05 | 1.70 ± 0.23 | 1.62 ± 0.06 | 1.96 ± 0.15 | 1.66 ± 0.15 |
| Feed conversion rate | 1.40 ± 0.11 | 1.22 ± 0.02 | 1.33 ± 0.09 | 1.14 ± 0.08 | 1.30 ± 0.12 |
| Somatic indexes | |||||
| Hepatosomatic index | 1.24 ± 0.07 | 1.30 ± 0.16 | 0.98 ± 0.26 | 1.15 ± 0.19 | 1.24 ± 0.04 |
| Viscerosomatic index | 8.67 ± 0.70 | 8.72 ± 1.62 | 8.28 ± 1.38 | 7.81 ± 0.54 | 8.74 ± 0.59 |
| CTRL | PEL1 | PEL10 | WO1 | WO10 | |
|---|---|---|---|---|---|
| WBC (×104 µL−1) | 9.5 ± 2.2 a | 7.8 ± 1.5 ab | 6.8 ± 1.3 b | 6.9 ± 1.8 b | 7.6 ± 1.9 ab |
| RBC (×106 µL−1) | 2.58 ± 0.42 | 2.71 ± 0.70 | 2.23 ± 0.61 | 2.46 ± 0.48 | 2.35 ± 0.60 |
| Ht (%) | 29.6 ± 5.2 ab | 26.0 ± 4.0 a | 25.5 ± 6.1 a | 33.2 ± 6.7 b | 29.0 ± 1.5 ab |
| Hb (g dL−1) | 1.39 ± 0.43 | 1.10 ± 0.11 | 1.04 ± 0.18 | 1.27 ± 0.31 | 1.04 ± 0.38 |
| MCV (µm3) | 104 ± 47 | 101 ± 26 | 111 ± 18 | 129 ± 18 | 121 ± 25 |
| MCH (pg cell−1) | 5.7 ± 2.4 | 4.2 ± 1.4 | 4.9 ± 1.2 | 4.9 ± 0.2 | 4.3 ± 1.8 |
| MCHC (g 100 mL−1) | 4.4 ± 0.3 | 4.1 ± 0.8 | 4.0 ± 0.4 | 3.9 ± 0.8 | 3.5 ± 1.0 |
| CTRL | PEL1 | PEL10 | WO1 | WO10 | |
|---|---|---|---|---|---|
| Anterior intestine | |||||
| Muscularis externa (µm) | 112 ± 16 | 117 ± 17 | 119 ± 29 | 116 ± 17 | 113 ± 17 |
| Outer longitudinal layer (µm) | 44 ± 13 a | 44 ± 14 a | 47 ± 18 a | 42 ± 12 ab | 41 ± 13 b |
| Inner circular layer (µm) | 47 ± 12 a | 43 ± 12 b | 51 ± 21 a | 44 ± 12 b | 42 ± 12 b |
| Villus length (µm) | 829 ± 242 ab | 865 ± 203 bd | 955 ± 283 c | 878 ± 168 cd | 827 ± 281 a |
| Villus width (µm) | 183 ± 60 a | 181 ± 64 a | 153 ± 49 b | 147 ± 46 bc | 138 ± 41 c |
| Goblet cells (no. GC fold−1) | 38 ± 18 a | 44 ± 22 a | 57 ± 29 b | 42 ± 21 a | 40 ± 22 a |
| Neutral GC (no. GC fold−1) | 0.61 ± 0.88 a | 0.14 ± 0.47 b | 1.8 ± 1.9 c | 1.4 ± 1.6 c | 1.0 ± 1.3 d |
| Acid GC (no. GC fold−1) | 36 ± 16 a | 44 ± 22 b | 56 ± 30 c | 45 ± 15 b | 39 ± 22 a |
| Posterior intestine | |||||
| Muscularis externa (µm) | 133 ± 42 ab | 119 ± 29 a | 152 ± 37 b | 120 ± 14 a | 144 ± 28 b |
| Outer longitudinal layer (µm) | 52 ± 21 a | 53 ± 18 a | 60 ± 20 b | 60 ± 19 b | 62 ± 24 b |
| Inner circular layer (µm) | 44 ± 14 a | 40 ± 12 b | 48 ± 17 c | 45 ± 13 ac | 47 ± 14 c |
| Villus length (µm) | 728 ± 192 a | 760 ± 187 ab | 810 ± 218 b | 762 ± 290 a | 697 ± 174 a |
| Villus width (µm) | 168 ± 60 ab | 158 ± 54 a | 182 ± 65 b | 165 ± 55 a | 169 ± 48 b |
| Goblet cells (no. GC fold−1) | 37 ± 18 ab | 34 ± 17 ac | 37 ± 18 ab | 41 ± 22 b | 30 ± 13 c |
| Neutral GC (no. GC fold−1) | 0.63 ± 0.89 a | 0.00 ± 0.00 b | 0.11 ± 0.36 c | 0.17 ± 0.48 c | 0.20 ± 0.56 c |
| Acid GC (no. GC fold−1) | 35 ± 17 a | 34 ± 17 a | 37 ± 18 ab | 42 ± 22 b | 31 ± 12 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pires, D.; Passos, R.; do Carmo, B.; Tchobanov, C.F.; Forte, S.; Vaz, M.; Antunes, M.; Neves, M.; Tecelão, C.; Baptista, T. Pelvetia canaliculata as an Aquafeed Supplement for Gilthead Seabream Sparus aurata: A Biorefinery Approach for Seaweed Biomass Valorisation. Sustainability 2022, 14, 11469. https://doi.org/10.3390/su141811469
Pires D, Passos R, do Carmo B, Tchobanov CF, Forte S, Vaz M, Antunes M, Neves M, Tecelão C, Baptista T. Pelvetia canaliculata as an Aquafeed Supplement for Gilthead Seabream Sparus aurata: A Biorefinery Approach for Seaweed Biomass Valorisation. Sustainability. 2022; 14(18):11469. https://doi.org/10.3390/su141811469
Chicago/Turabian StylePires, Damiana, Ricardo Passos, Beatriz do Carmo, Carolina F. Tchobanov, Sara Forte, Mariana Vaz, Madalena Antunes, Marta Neves, Carla Tecelão, and Teresa Baptista. 2022. "Pelvetia canaliculata as an Aquafeed Supplement for Gilthead Seabream Sparus aurata: A Biorefinery Approach for Seaweed Biomass Valorisation" Sustainability 14, no. 18: 11469. https://doi.org/10.3390/su141811469
APA StylePires, D., Passos, R., do Carmo, B., Tchobanov, C. F., Forte, S., Vaz, M., Antunes, M., Neves, M., Tecelão, C., & Baptista, T. (2022). Pelvetia canaliculata as an Aquafeed Supplement for Gilthead Seabream Sparus aurata: A Biorefinery Approach for Seaweed Biomass Valorisation. Sustainability, 14(18), 11469. https://doi.org/10.3390/su141811469