
Soil Bacteria and Fungi Respond Differently to Organisms Covering on Leshan Giant Buddha Body


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection
2.3. Determination of Chemical Properties
2.4. DNA Extraction and High-Throughput Sequencing
2.5. Data Processing and Statistical Analyses
3. Results
3.1. Chemical Properties of Samples
3.2. Analysis of Microbial Communities
3.2.1. Alpha Diversity of Bacterial and Fungal Communities
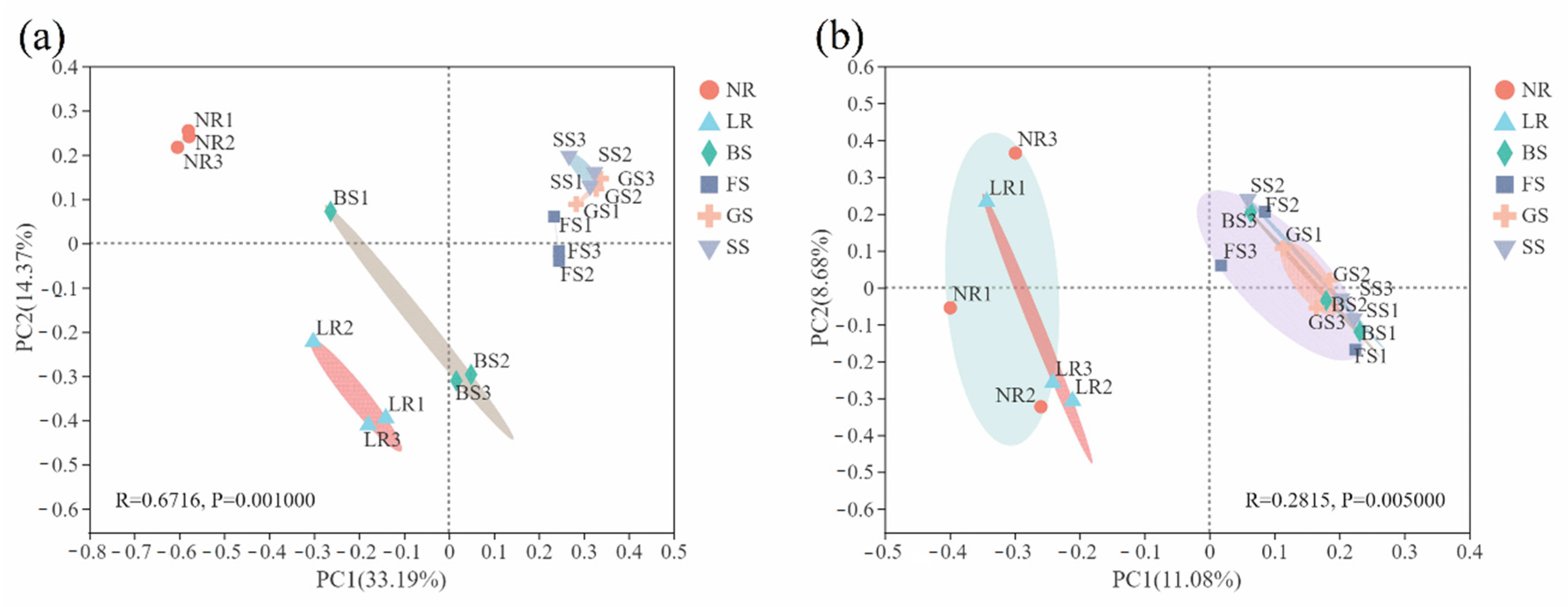
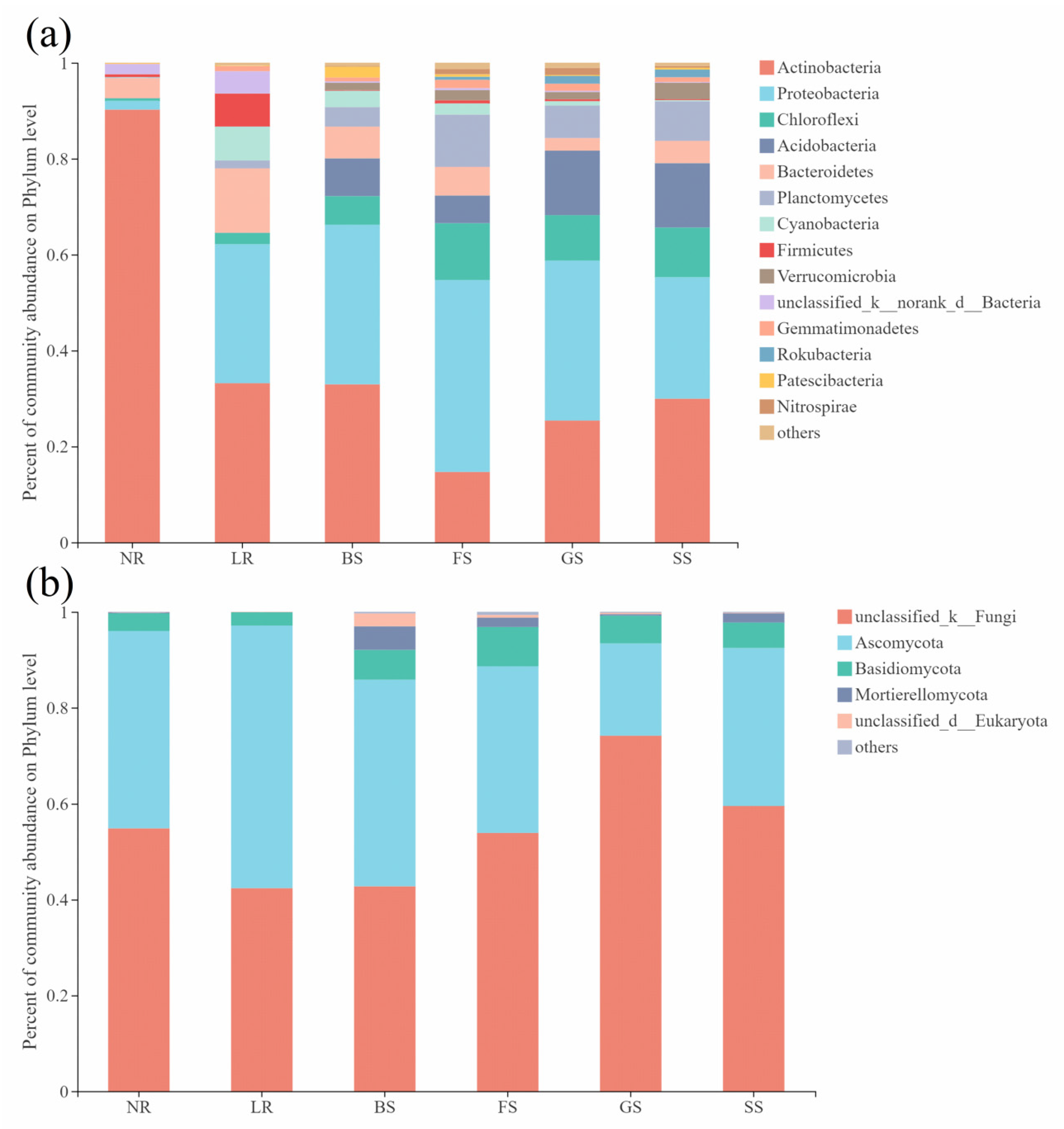
3.2.2. Structure and Composition of Bacterial and Fungal Communities
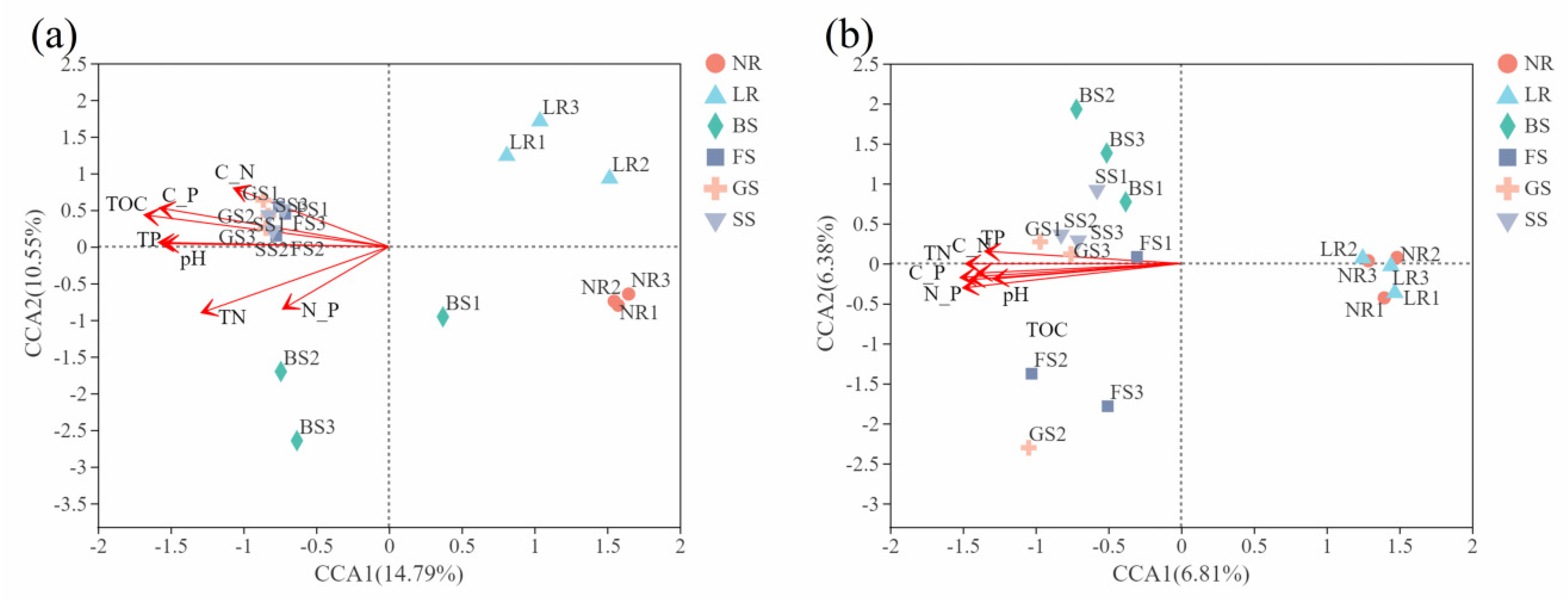
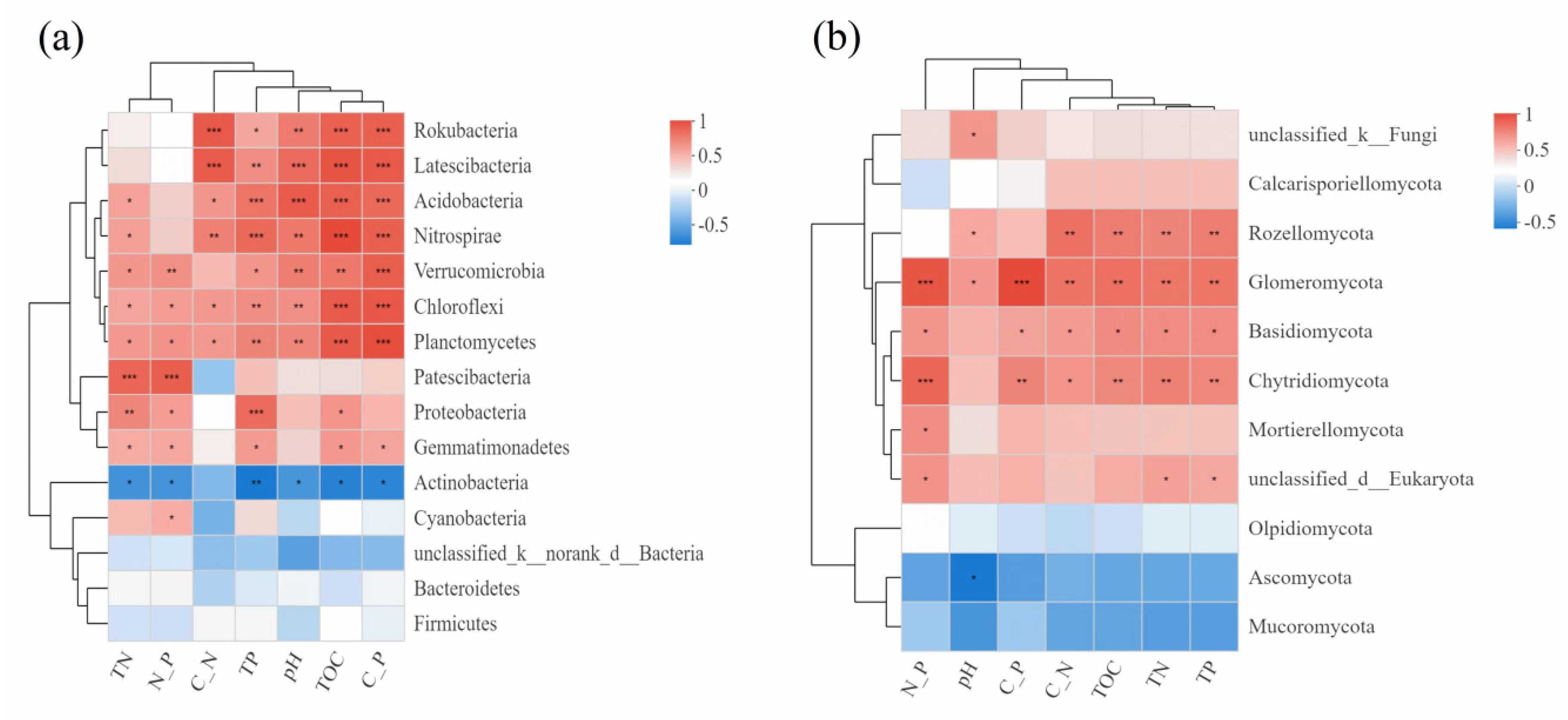
3.2.3. Correlation Analysis between Microbial Diversity and Ecological Factors
4. Discussion
4.1. Responses of Environmental Properties to the Organism Types
4.2. Responses of Microbial Community Diversity and Richness to the Organism Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Freeman, C.; Ostle, N.J. Microbial contributions to climate change through carbon cycle feedbacks. ISME J. 2008, 2, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ding, J.; Peng, Y.; Li, F.; Yang, G.; Liu, L.; Qin, S.; Fang, K.; Yang, Y. Patterns and drivers of soil microbial communities in Tibetan alpine and global terrestrial ecosystems. J. Biogeogr. 2016, 43, 2027–2039. [Google Scholar] [CrossRef]
- Burns, J.H.; Anacker, B.L.; Strauss, S.Y.; Burke, D.J. Soil microbial community variation correlates most strongly with plant species identity, followed by soil chemistry, spatial location and plant genus. Aob Plants 2015, 7, v30. [Google Scholar] [CrossRef]
- Maharning, A.R.; Mills, A.A.S.; Adl, S.M. Soil community changes during secondary succession to naturalized grasslands. Appl. Soil Ecol. 2009, 41, 137–147. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef]
- Singh, B.K.; Millard, P.; Whiteley, A.S.; Murrell, J.C. Unravelling rhizosphere–microbial interactions: Opportunities and limitations. Trends Microbiol. 2004, 12, 386–393. [Google Scholar] [CrossRef]
- He, X.; Wang, K.; Zhang, W.; Chen, Z.; Zhu, Y.; Chen, H. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst. Plant Soil 2008, 307, 123–134. [Google Scholar] [CrossRef]
- Kielak, A.; Pijl, A.S.; Van Veen, J.A.; Kowalchuk, G.A. Differences in vegetation composition and plant species identity lead to only minor changes in soil-borne microbial communities in a former arable field. FEMS Microbiol. Ecol. 2008, 63, 372–382. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, M.; Zhang, X.; Sun, Q.; Liu, R.; Lian, B. Shift of the microbial communities from exposed sandstone rocks to forest soils during pedogenesis. Int. Biodeterior. Biodegrad. 2019, 140, 21–28. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhang, X.; Wang, X.; Fu, S.; Wu, S.; Lu, X.; Ren, C.; Han, X.; Yang, G. Soil bacteria and fungi respond differently to plant diversity and plant family composition during the secondary succession of abandoned farmland on the Loess Plateau, China. Plant Soil 2020, 448, 183–200. [Google Scholar] [CrossRef]
- Xu, M.; Gao, D.; Fu, S.; Lu, X.; Wu, S.; Han, X.; Yang, G.; Feng, Y. Long-term effects of vegetation and soil on the microbial communities following afforestation of farmland with Robinia pseudoacacia plantations. Geoderma 2020, 367, 114263. [Google Scholar] [CrossRef]
- Lazzaro, A.; Hilfiker, D.; Zeyer, J. Structures of microbial communities in alpine soils: Seasonal and elevational effects. Front. Microbiol. 2015, 6, 1330. [Google Scholar] [CrossRef] [PubMed]
- Hazard, C.; Gosling, P.; van der Gast, C.J.; Mitchell, D.T.; Doohan, F.M.; Bending, G.D. The role of local environment and geographical distance in determining community composition of arbuscular mycorrhizal fungi at the landscape scale. ISME J. 2013, 7, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Knelman, J.E.; Gasarch, E.; Wang, D.; Nemergut, D.R.; Seastedt, T.R. Plant community and soil chemistry responses to long-term nitrogen inputs drive changes in alpine bacterial communities. Ecology 2016, 97, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Zinger, L.; Lejon, D.P.; Baptist, F.; Bouasria, A.; Aubert, S.; Geremia, R.A.; Choler, P. Contrasting diversity patterns of crenarchaeal, bacterial and fungal soil communities in an alpine landscape. PLoS ONE 2011, 6, e19950. [Google Scholar] [CrossRef]
- Liu, Z.; Fu, B.; Zheng, X.; Liu, G. Plant biomass, soil water content and soil N:P ratio regulating soil microbial functional diversity in a temperate steppe: A regional scale study. Soil Biol. Biochem. 2010, 42, 445–450. [Google Scholar] [CrossRef]
- Daniel, D.W.; Smith, L.M.; Belden, J.B.; McMurry, S.T.; Swain, S. Effects of land-use change and fungicide application on soil respiration in playa wetlands and adjacent uplands of the U.S. High Plains. Sci. Total Environ. 2015, 514, 290–297. [Google Scholar] [CrossRef]
- Bakker, M.G.; Schlatter, D.C.; Otto-Hanson, L.; Kinkel, L.L. Diffuse symbioses: Roles of plant-plant, plant-microbe and microbe-microbe interactions in structuring the soil microbiome. Mol. Ecol. 2014, 23, 1571–1583. [Google Scholar] [CrossRef] [PubMed]
- Dang, P.; Gao, Y.; Liu, J.; Yu, S.; Zhao, Z. Effects of thinning intensity on understory vegetation and soil microbial communities of a mature Chinese pine plantation in the Loess Plateau. Sci. Total Environ. 2018, 630, 171–180. [Google Scholar] [CrossRef]
- Williams, A.; Börjesson, G.; Hedlund, K. The effects of 55 years of different inorganic fertiliser regimes on soil properties and microbial community composition. Soil Biol. Biochem. 2013, 67, 41–46. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; de Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.; Schlaeppi, K. A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef]
- Lange, M.; Habekost, M.; Eisenhauer, N.; Roscher, C.; Bessler, H.; Engels, C.; Oelmann, Y.; Scheu, S.; Wilcke, W.; Schulze, E.D.; et al. Biotic and abiotic properties mediating plant diversity effects on soil microbial communities in an experimental grassland. PLoS ONE 2014, 9, e96182. [Google Scholar] [CrossRef]
- Roy, J.; Albert, C.H.; Ibanez, S.; Saccone, P.; Zinger, L.; Choler, P.; Clément, J.C.; Lavergne, S.; Geremia, R.A. Microbes on the cliff: Alpine cushion plants structure bacterial and fungal communities. Front. Microbiol. 2013, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Chalmandrier, L.; Münkemüller, T.; Colace, M.; Renaud, J.; Aubert, S.; Carlson, B.Z.; Clément, J.; Legay, N.; Pellet, G.; Saillard, A.; et al. Spatial scale and intraspecific trait variability mediate assembly rules in alpine grasslands. J. Ecol. 2017, 105, 277–287. [Google Scholar] [CrossRef]
- Chase, J.M. Spatial scale resolves the niche versus neutral theory debate. J. Veg. Sci. 2014, 25, 319–322. [Google Scholar] [CrossRef]
- Chalmandrier, L.; Münkemüller, T.; Gallien, L.; de Bello, F.; Mazel, F.; Lavergne, S.; Thuiller, W. A family of null models to distinguish between environmental filtering and biotic interactions in functional diversity patterns. J. Veg. Sci. 2013, 24, 853–864. [Google Scholar] [CrossRef]
- Liang, Z.; Peng, X.; Luan, Z.; Li, W.; Zhao, Y. Reduction of phosphorus release from high phosphorus soil by red mud. Environ. Earth Sci. 2012, 65, 581–588. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Su, S.; Zhao, Y.; Liu, Z.; Liu, G.; Du, M.; Wu, J.; Bai, D.; Li, B.; Bou, G.; Zhang, X.; et al. Characterization and comparison of the bacterial microbiota in different gastrointestinal tract compartments of Mongolian horses. Microbiologyopen 2020. [Google Scholar] [CrossRef]
- Schultz, J.; Wolf, M. ITS2 sequence–structure analysis in phylogenetics: A how-to manual for molecular systematics. Mol. Phylogenet. Evol. 2009, 52, 520–523. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Romero, E.; Cammeraat, E.; Pérez-Cardiel, E.; Lasanta, T. Effects of secondary succession and afforestation practices on soil properties after cropland abandonment in humid Mediterranean mountain areas. Agric. Ecosyst. Environ. 2016, 228, 91–100. [Google Scholar] [CrossRef]
- Cheng, M.; An, S. Responses of soil nitrogen, phosphorous and organic matter to vegetation succession on the Loess Plateau of China. J. Arid. Land 2015, 7, 216–223. [Google Scholar] [CrossRef]
- Zhu, H.; He, X.; Wang, K.; Su, Y.; Wu, J. Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem. Eur. J. Soil Biol. 2012, 51, 1–7. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Ponge, J. Humus forms in terrestrial ecosystems: A framework to biodiversity. Soil Biol. Biochem. 2003, 35, 935–945. [Google Scholar] [CrossRef]
- Yan, M.; Zhang, W.; Zhang, Z.; Wang, L.; Ren, H.; Jiang, Y.; Zhang, X. Responses of soil C stock and soil C loss to land restoration in Ili River Valley, China. Catena 2018, 171, 469–474. [Google Scholar] [CrossRef]
- Sun, C.; Liu, G.; Xue, S. Natural succession of grassland on the Loess Plateau of China affects multifractal characteristics of soil particle-size distribution and soil nutrients. Ecol. Res. 2016, 31, 891–902. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, P.; Xu, G.; Li, Z.; Cheng, S.; Gao, H. Spatial distribution of soil total phosphorus in Yingwugou watershed of the Dan River, China. Catena 2016, 136, 175–181. [Google Scholar] [CrossRef]
- Lane, P.N.J.; Noske, P.J.; Sheridan, G.J. Phosphorus enrichment from point to catchment scale following fire in eucalypt forests. Catena 2011, 87, 157–162. [Google Scholar] [CrossRef]
- Kooijman, A.M.; Jongejans, J.; Sevink, J. Parent material effects on Mediterranean woodland ecosystems in NE Spain. Catena 2005, 59, 55–68. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Yang, L.; Luo, P.; Chen, H.; Xiao, K.; Song, T.; Zhang, W.; He, X.; Chen, H.; et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Reed, S.C.; Cleveland, C.C.; Townsend, A.R. Functional ecology of free-living nitrogen fixation: A contemporary perspective. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 489–512. [Google Scholar] [CrossRef]
- Wu, F.; Yang, W.; Sun, B.; Yang, T.; Chen, X.; Xu, Z.; Song, H. Soil C, N and P stocks and stoichiometry under different vegetation on the surface of the Leshan Giant Buddha. Soil Ecol. Lett. 2020. [Google Scholar] [CrossRef]
- Song, M.; Peng, W.; Du, H.; Xu, Q. Responses of soil and microbial C:N:P stoichiometry to vegetation succession in a karst region of southwest China. Forests 2019, 10, 755. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Fang, Y.; Ma, R.; Lal, R.; An, S.; Huang, Y. Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yunwu Mountain Reserve of China. Ecol. Eng. 2017, 109, 92–100. [Google Scholar] [CrossRef]
- Wang, B.; Liu, G.B.; Xue, S.; Zhu, B. Changes in soil physico-chemical and microbiological properties during natural succession on abandoned farmland in the Loess Plateau. Environ. Earth Sci. 2011, 62, 915–925. [Google Scholar] [CrossRef]
- Xu, C.; Xiang, W.; Gou, M.; Chen, L.; Lei, P.; Fang, X.; Deng, X.; Ouyang, S. Effects of forest restoration on soil carbon, nitrogen, phosphorus, and their stoichiometry in Hunan, southern China. Sustainability 2018, 10, 1874. [Google Scholar] [CrossRef]
- Lei, Y.; Zhou, J.; Xiao, H.; Duan, B.; Wu, Y.; Korpelainen, H.; Li, C. Soil nematode assemblages as bioindicators of primary succession along a 120-year-old chronosequence on the Hailuogou Glacier forefield, SW China. Soil Biol. Biochem. 2015, 88, 362–371. [Google Scholar] [CrossRef]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Hayes, P.; Turner, B.L.; Lambers, H.; Laliberté, E. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 2014, 102, 396–410. [Google Scholar] [CrossRef]
- Kim, M.; Jung, J.Y.; Laffly, D.; Kwon, H.Y.; Lee, Y.K. Shifts in bacterial community structure during succession in a glacier foreland of the High Arctic. FEMS Microbiol. Ecol. 2017, 93, w213. [Google Scholar] [CrossRef]
- Jia, G.; Cao, J.; Wang, C.; Wang, G. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China. Forest Ecol. Manag. 2005, 217, 117–125. [Google Scholar] [CrossRef]
- Juottonen, H.; Männistö, M.; Tiirola, M.; Kytöviita, M.M. Cryptogams signify key transitions of bacteria and fungi in Arctic sand dune succession. New Phytol. 2020, 226, 1836–1849. [Google Scholar] [CrossRef]
- Alfaro, F.D.; Manzano, M.; Marquet, P.A.; Gaxiola, A. Microbial communities in soil chronosequences with distinct parent material: The effect of soil pH and litter quality. J. Ecol. 2017, 105, 1709–1722. [Google Scholar] [CrossRef]
- Due, L.; Noll, M.; Meier, B.E. High Diversity of Diazotrophs in the Forefield of a Receding Alpine Glacier. Microb. Ecol. 2009, 1, 179–190. [Google Scholar]
- McCarthy, A.J.; Williams, S.T. Actinomycetes as agents of biodegradation in the environment—A review. Gene 1992, 115, 189–192. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Zhou, H.; Ganjurjav, H.; Wang, X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef]
- Pan, C.; Bao, Y.; Guo, A.; Ma, J. Environmentally Relevant-Level CeO2 NP with Ferrous Amendment Alters Soil Bacterial Community Compositions and Metabolite Profiles in Rice-Planted Soils. J. Agric. Food Chem. 2020, 68, 8172–8184. [Google Scholar] [CrossRef] [PubMed]
- Sait, M.; Davis, K.E.R.; Janssen, P.H. Effect of pH on isolation and distribution of members of subdivision 1 of the phylum Acidobacteria occurring in soil. Appl. Environ. Microb. 2006, 72, 1852–1857. [Google Scholar] [CrossRef]
- Chen, X.; Su, Y.; He, X.; Hu, L.; Liang, Y.; Feng, S.; Ge, Y.; Xiao, W. Basidiomycetous laccase gene diversity in two subtropical forest soils. Ying Yong Sheng Tai Xue Bao 2011, 22, 2699–2704. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Groups | NR | LR | BS | FS | GS | SS |
|---|---|---|---|---|---|---|
| TOC (g·kg−1) | 2.45 ± 0.48 d | 6.48 ± 1.15 d | 26.71 ± 4.97 c | 48.51 ± 4.48 b | 64.03 ± 4.28 a | 68.86 ± 8.69 a |
| TN(g·kg−1) | 0.71 ± 0.12 e | 1.66 ± 0.26 d | 4.02 ± 0.66 c | 5.2 ± 0.6 b | 6.28 ± 0.31 a | 6.49 ± 0.71 a |
| TP(g·kg−1) | 0.19 ± 0.03 d | 0.34 ± 0.07 c | 0.51 ± 0.06 b | 0.58 ± 0.05 b | 0.74 ± 0.09 a | 0.82 ± 0.1 a |
| C:N | 3.45 ± 0.22 d | 3.91 ± 0.11 d | 6.63 ± 0.5 c | 9.34 ± 0.32 b | 10.19 ± 0.22 a | 10.6 ± 0.26 a |
| C:P | 12.72 ± 0.2 d | 18.95 ± 0.77 c | 52.35 ± 4.21 b | 84.33 ± 3.18 a | 86.99 ± 5.37 a | 83.68 ± 0.47 a |
| N:P | 3.7 ± 0.26 d | 4.85 ± 0.28 c | 7.9 ± 0.47 b | 9.03 ± 0.34 a | 8.55 ± 0.68 a b | 7.9 ± 0.2 b |
| pH | 6.68 ± 0.1 c | 6.65 ± 0.23 c | 6.91 ± 0.07 b c | 6.91 ± 0.23 b c | 7.28 ± 0.1 a | 7.15 ± 0.03 a b |
| Sample Groups | α-Diversity Indices | |||||
|---|---|---|---|---|---|---|
| Sobs | Shannon | Ace | Chao | Coverage | ||
| NR | 156 ± 112 d | 0.71 ± 0.41 d | 516 ± 401 c | 380 ± 323 c | 0.9977 | |
| LR | 346 ± 187 d | 3.49 ± 0.27 c | 538 ± 337 c | 511 ± 315 c | 0.9968 | |
| Bacteria | BS | 1088 ± 176 c | 5.4 ± 0.97 b | 1225 ± 219 b | 1285 ± 245 b | 0.994 |
| FS | 1801 ± 52 a | 6.53 ± 0.13 a | 2054 ± 97 a | 2093 ± 79 a | 0.9891 | |
| GS | 1518 ± 123 a b | 6.29 ± 0.18 a | 1725 ± 113 a b | 1761 ± 119 a b | 0.991 | |
| SS | 1323 ± 68 b c | 6.17 ± 0.1 a b | 1475 ± 101 a b | 1524 ± 103 b | 0.9929 | |
| NR | 1131 ± 415 a | 5.05 ± 0.95 a | 1162 ± 407 a | 1180 ± 396 a | 0.9989 | |
| LR | 2434 ± 1929 a | 4.61 ± 2.26 a | 2465 ± 1941 a | 2482 ± 1960 a | 0.9979 | |
| Fungi | BS | 2927 ± 1839 a | 6.3 ± 1.37 a | 2992 ± 1850 a | 3025 ± 1875 a | 0.9963 |
| FS | 4143 ± 3798 a | 6.91 ± 1.98 a | 4244 ± 3890 a | 4275 ± 3898 a | 0.9944 | |
| GS | 3896 ± 206 a | 7.69 ± 0.34 a | 3945 ± 217 a | 3998 ± 222 a | 0.9966 | |
| SS | 2805 ± 1720 a | 6.67 ± 1.75 a | 2855 ± 1699 a | 2889 ± 1725 a | 0.9974 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Wang, M.; Wu, F.; Sun, B.; Yang, T.; Song, H. Soil Bacteria and Fungi Respond Differently to Organisms Covering on Leshan Giant Buddha Body. Sustainability 2021, 13, 3897. https://doi.org/10.3390/su13073897
Chen X, Wang M, Wu F, Sun B, Yang T, Song H. Soil Bacteria and Fungi Respond Differently to Organisms Covering on Leshan Giant Buddha Body. Sustainability. 2021; 13(7):3897. https://doi.org/10.3390/su13073897
Chicago/Turabian StyleChen, Xuli, Manfei Wang, Fujia Wu, Bo Sun, Tianyu Yang, and Huixing Song. 2021. "Soil Bacteria and Fungi Respond Differently to Organisms Covering on Leshan Giant Buddha Body" Sustainability 13, no. 7: 3897. https://doi.org/10.3390/su13073897
APA StyleChen, X., Wang, M., Wu, F., Sun, B., Yang, T., & Song, H. (2021). Soil Bacteria and Fungi Respond Differently to Organisms Covering on Leshan Giant Buddha Body. Sustainability, 13(7), 3897. https://doi.org/10.3390/su13073897