
The Ecological Status of Juniperus foetidissima Forest Stands in the Mt. Oiti-Natura 2000 Site in Greece

 ,
, 
 and
and
Abstract
1. Introduction
2. Study Site and Methods
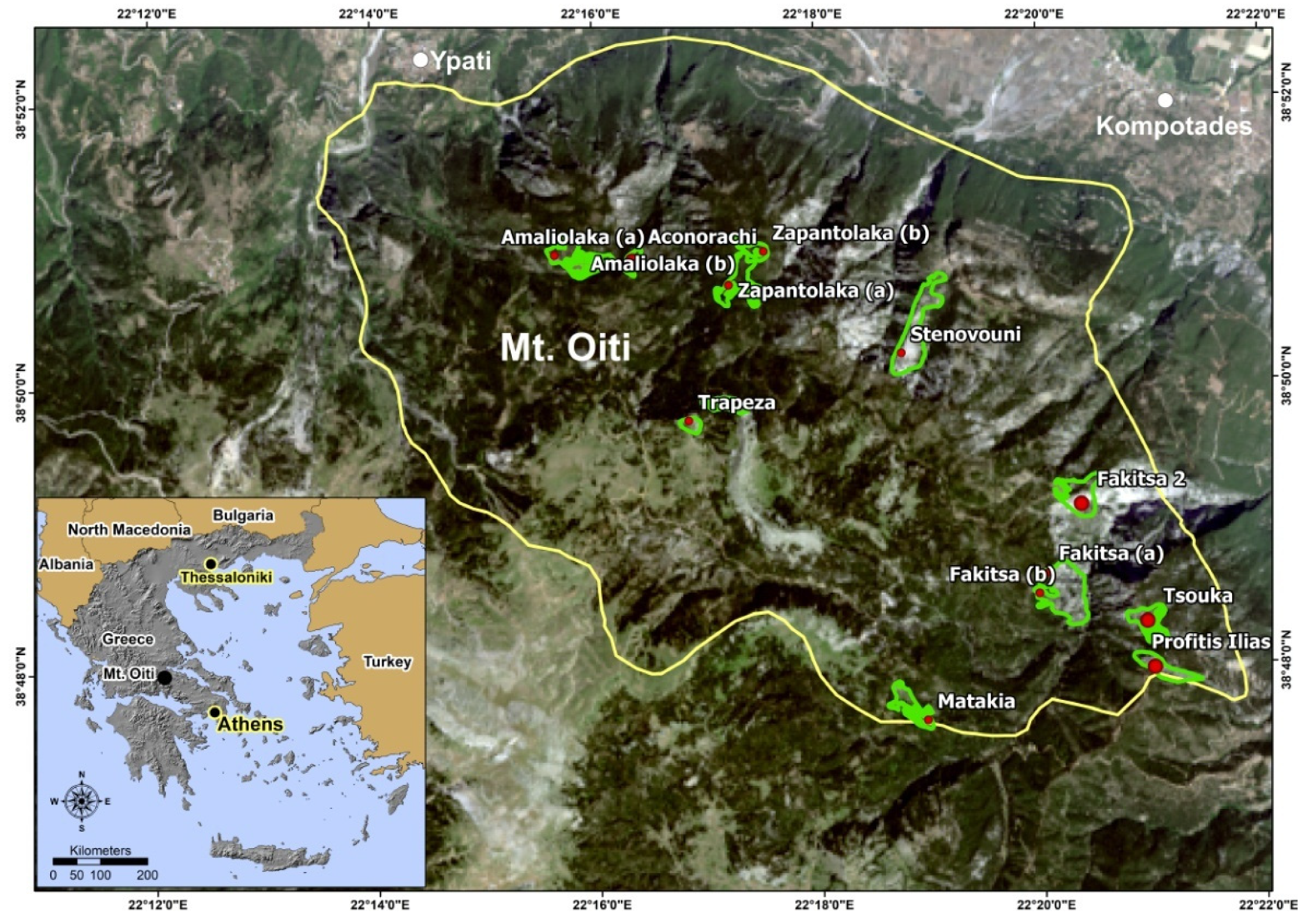
2.1. Study Site
2.2. Plot Design and Data Collection
2.3. Data Analysis
3. Results and Discussion
3.1. Climate Change Trends
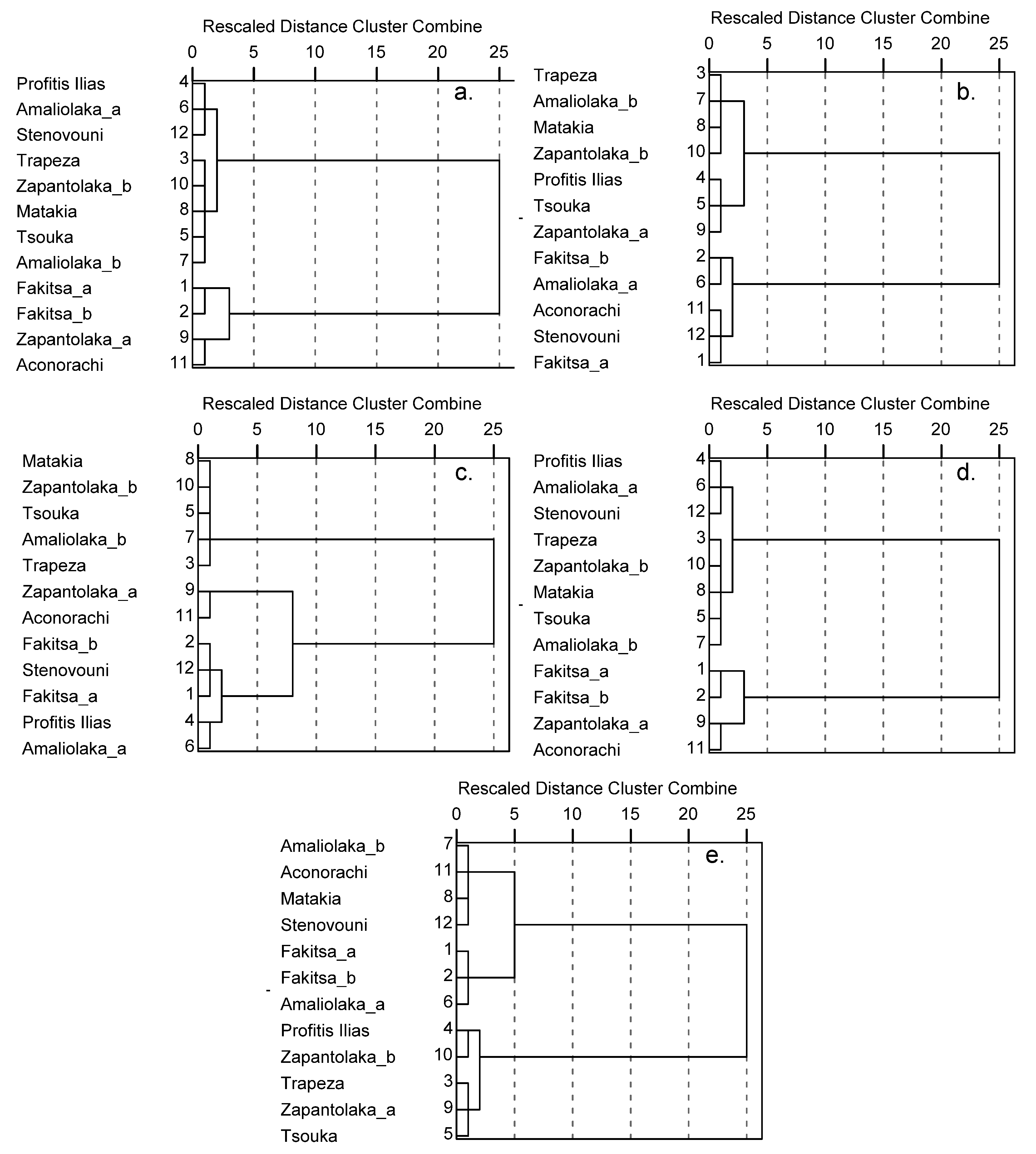
3.2. Subpopulations Distribution and Density
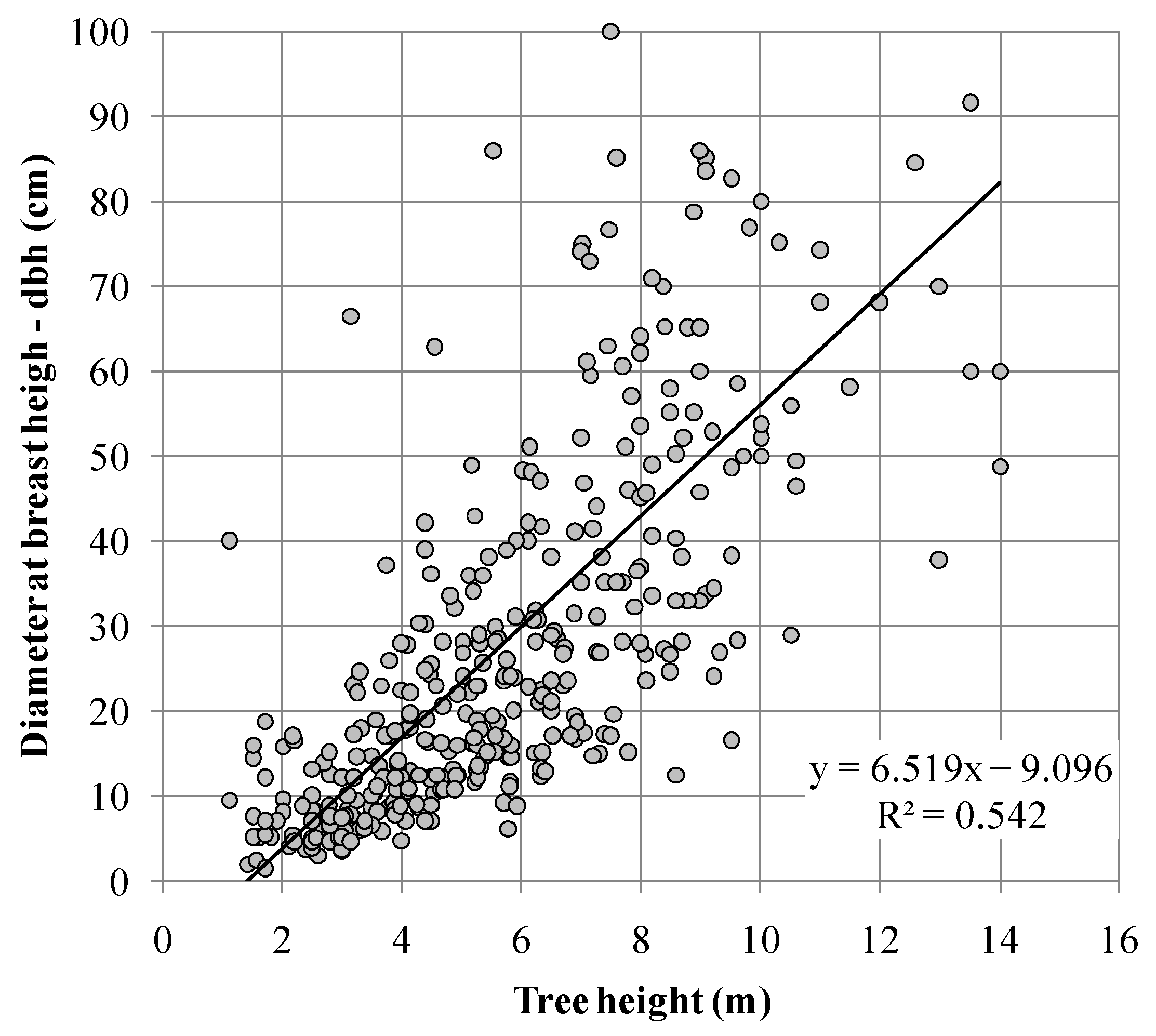
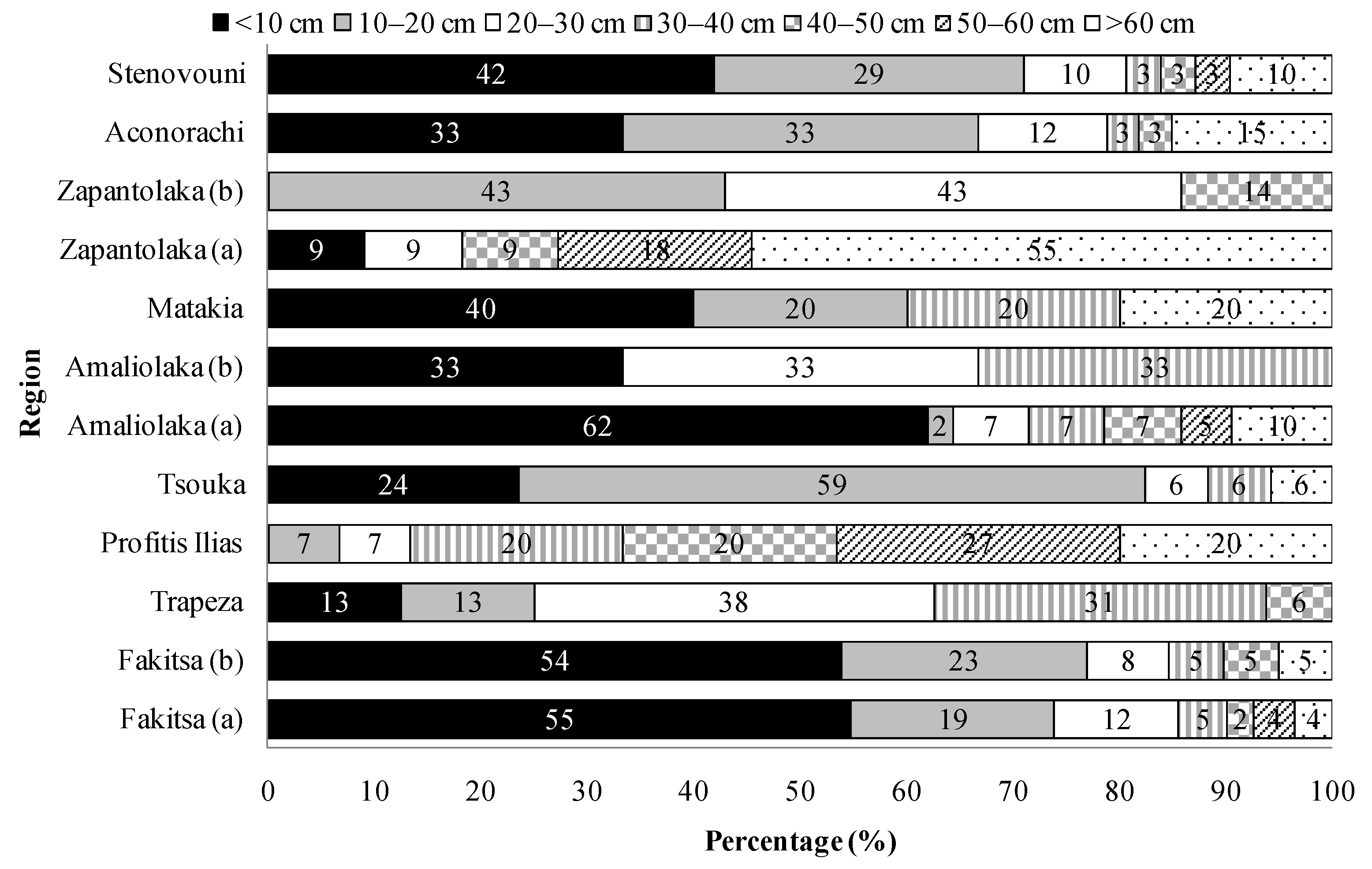
3.3. Trees Size and Structure
3.4. Tree Sex and Reproductive Dynamics
4. Evaluation of J. foetidissima Ecological Status in Mt. Oiti
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | Latitude Coordinates | Longitude Coordinates |
|---|---|---|
| Aconorachi | 38°50′52.88″ N | 22°16′21.25″ E |
| Amaliolaka (a) | 38°50′55.49″ N | 22°15′38.62″ E |
| Amaliolaka (b) | 38°50′51.96″ N | 22°15′54.28″ E |
| Fakitsa (a) | 38°48′38.66″ N | 22°20′3.87″ E |
| Fakitsa (b) | 38°48′29.67″ N | 22°19′58.68″ E |
| Matakia | 38°47′36.96″ N | 22°18′57.22″ E |
| Profitis Ilias | 38°48′0.53″ N | 22°21′0.76″ E |
| Stenovouni | 38°50′12.28″ N | 22°18′45.76″ E |
| Trapeza | 38°49′44.65″ N | 22°16′50.47″ E |
| Tsouka | 38°48′19.24″ N | 22°20′56.60″ E |
| Zapantolaka (a) | 38°50′41.73″ N | 22°17′12.58″ E |
| Zapantolaka (b) | 38°50′55.85″ N | 22°17′31.62″ E |
References
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Arroyo, M.K.; Arianoutsou, M. Plant diversity in Mediterranean-climate regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Davis, G.; Richardson, D. Mediterranean-Type Ecosystems. The Function of Biodiversity; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar] [CrossRef]
- Perrino, E.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a Mediterranean biodiversity hot spot (Gargano–Italy). Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Englera; Botanischer Garten und Botanisches Museum Berlin-Dahlem, Freie Universität Berlin: Berlin, Germany; Hellenic Botanical Society: Athens, Greece, 2013; Volume 31, pp. 1–370. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Supplement. Willdenowia 2016, 46, 301–347. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant Endemism Centres and Biodiversity Hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef]
- Solomou, A.D.; Sfougaris, A. Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape. Agronomy 2021, 11, 161. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T. Propagation of Juniper Species by Plant Tissue Culture: A Mini-Review. Forests 2019, 10, 1028. [Google Scholar] [CrossRef]
- Kasaian, J.; Behravan, J.; Hassany, M.; Emami, S.; Shahriari, F.; Khayyat, M. Molecular characterization and RAPD analysis of Juniperus species from Iran. Genet. Mol. Res. GMR 2011, 10, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Ghasemnezhad, A.; Ghorbanzadeh, A.; Sarmast, M.K.; Ghorbanpour, M. A Review on Botanical, Phytochemical, and Pharmacological Characteristics of Iranian Junipers (Juniperus spp.). In Plant-Derived Bioactives; Springer: Berlin/Heidelberg, Germany, 2020; pp. 493–508. [Google Scholar] [CrossRef]
- Farjon, A. Juniperus foetidissima. The IUCN Red List of Threatened Species 2013: E.T42234A2965043. Available online: https://www.iucnredlist.org/species/42234/2965043 (accessed on 13 March 2021).
- Clifton, S.J.; Ward, L.K.; Ranner, D.S. The status of juniper Juniperus communis L. in northeast England. Biol. Conserv. 1997, 79, 67–77. [Google Scholar] [CrossRef]
- García, D.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Age structure of Juniperus communis L. in the Iberian peninsula: Conservation of remnant populations in Mediterranean mountains. Biol. Conserv. 1999, 87, 215–220. [Google Scholar] [CrossRef]
- Oostermeijer, J.G.B.; De Knegt, B. Genetic population structure of the wind-pollinated, dioecious shrub Juniperus communis in fragmented Dutch heathlands. Plant Spec. Biol. 2004, 19, 175–184. [Google Scholar] [CrossRef]
- García, D. Regeneración natural del enebro Juniperus communisL. In Áreas de Alta Montaña Mediterranea, Conectando la Ecología Reproductiva con el Reclutamiento Poblacional; Universidad de Granada: Granada, Spain, 1998. [Google Scholar]
- Eriksson, O.; Ehrlén, J. Seed and microsite limitation of recruitment in plant populations. Oecologia 1992, 91, 360–364. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Zamora, R.; Gómez, J.M.; Jordano, P.; Hódar, J.A. Geographical variation in seed production, predation and abortion in Juniperus communis throughout its range in Europe. J. Ecol. 2000, 88, 435–446. [Google Scholar] [CrossRef]
- Zamora, R.; Gómez, J.; García, D.; Hódar, J.A. Ecología reproductiva y regeneración del matorral de la alta montaña de Sierra Nevada: Capacidad de respuesta a las perturbaciones. In Sierra Nevada: Conservación y Desarrollo Sostenible; Rosúa, C., Ed.; Universidad de Granada: Granada, Spain, 1996; Volume 2, pp. 407–422. [Google Scholar]
- Doğan, H.H.; Karadelev, M.; Işiloğlu, M. Macrofungal diversity associated with the scale-leaf juniper trees, Juniperus excelsa and J. foetidissima, distributed in Turkey. Turk. J. Bot. 2011, 35, 219–237. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation dynamics and sex structure of the populations of pioneer dioecious woody plants. Vegetatio 1980, 43, 23–38. [Google Scholar] [CrossRef]
- Rosén, E. Periodic droughts and long-term dynamics of alvar grassland vegetation on Öland, Sweden. Folia Geobot. 1995, 30, 131–140. [Google Scholar] [CrossRef]
- Rosén, E.; Bakker, J.P. Effects of agri-environment schemes on scrub clearance, livestock grazing and plant diversity in a low-intensity farming system on Öland, Sweden. Basic Appl. Ecol. 2005, 6, 195–204. [Google Scholar] [CrossRef]
- Arabatzis, T. Shrubs and Trees in Greece; Oikologiki Kinisi Dramas, Technological Educational Institute of Kavala: Drama, Greece, 1998; p. 292. ISBN 960-85951-2-6. (In Greek) [Google Scholar]
- Korakis, G. Forest Botany (e-Book); Association of Greek Academic Libraries: Athens, Greece, 2015. [Google Scholar]
- Barbero, M.; Quézel, P. Les groupements forestiers de Grèce centro-méridionale. Ecol. Mediterr. 1976, 2, 3–86. [Google Scholar] [CrossRef]
- Quézel, P. Contribution a l’ etude de la vegetation du Vardussia. Biol. Gallo Hell. 1973, 5, 139–166. [Google Scholar]
- Dimopoulos, P. Floristic and Phytosociological Research on Mount Kilini: An Ecological Approach; University of Patras: Patras, Greece, 1993. [Google Scholar]
- Karetsos, G. Study of the Ecology and Vegetation of Mount Iti. Ph.D. Thesis, University of Patras, Patras, Greece, 2002. (In Greek with English Summary). [Google Scholar]
- Stara, K.; Tsiakiris, R.; Wong, J.L. Valuing trees in a changing cultural landscape: A case study from northwestern Greece. Hum. Ecol. 2015, 43, 153–167. [Google Scholar] [CrossRef]
- Médail, F.; Monnet, A.-C.; Pavon, D.; Nikolic, T.; Dimopoulos, P.; Bacchetta, G.; Arroyo, J.; Barina, Z.; Albassatneh, M.C.; Domina, G. What is a tree in the Mediterranean Basin hotspot? A critical analysis. For. Ecosyst. 2019, 6, 1–19. [Google Scholar] [CrossRef]
- Carus, S. Increment and Growth in Crimean Juniper (Juniperus excelsa Bieb.) Stands in Isparta—Siitciiler Region of Turkey. J. Biol. Sci. 2004, 4, 173–179. [Google Scholar] [CrossRef][Green Version]
- Davis, P.; Miller, R.; Tan, K. Flora of Turkey and the Aegean Islands. Vol. 1–9; Edinburg Universty Press: Edinburg, TX, USA, 1965. [Google Scholar]
- European Commission. The Interpretation Manual of European Union Habitats–EUR28; Technical Report; European Commission, DG Environment, Nature ENV B.3: Brussels, Belgium, 2013; p. 146. [Google Scholar]
- NATURA 2000-Standard Data Form for Site GR2440004: Ethnikos Drymos Oitis. Database Release Date: 12/06/2020. Available online: https://natura2000.eea.europa.eu/Natura2000/SDF.aspx?site=GR2440004#7 (accessed on 26 February 2021).
- Adams, R. Junipers of the World: The Genius Juniperus, 4th ed.; Trafford Publishing: Bloomington, IN, USA, 2014; p. 422. ISBN 978-1490-723259. [Google Scholar]
- Lesjak, M.M.; Beara, I.N.; Orčić, D.Z.; Ristić, J.D.; Anačkov, G.T.; Božin, B.N.; Mimica-Dukić, N.M. Chemical characterisation and biological effects of Juniperus foetidissima Willd. 1806. LWT Food Sci. Technol. 2013, 53, 530–539. [Google Scholar] [CrossRef]
- Parvin Salehi, S.; Mirza, M.; Calagari, M. Composition of the essential oils of Junipers (Juniperus foetidissima and J. oblonga) from Arasbaran protected area. J. Essent. Oil Bear. Plants 2016, 19, 1261–1266. [Google Scholar] [CrossRef]
- Sahin Yaglioglu, A.; Eser, F.; Yaglioglu, M.S.; Demirtas, I. The antiproliferative and antioxidant activities of the essential oils of Juniperus species from Turkey. Flavour Fragr. J. 2020, 35, 511–523. [Google Scholar] [CrossRef]
- Tunalier, Z.; Kirimer, N.; Baser, K. The composition of essential oils from various parts of Juniperus foetidissima. Chem. Nat. Compd. 2002, 38, 43–47. [Google Scholar] [CrossRef]
- Junicoast. LIFE+ Nature Project “Actions for the Conservation of Coastal Dunes with Juniperus spp. in Crete adn the South Aegean (Greece)” LIFE07NAT/GR/000296. Available online: http://www.junicoast.gr/ (accessed on 13 March 2021).
- JunEx. LIFE+ Nature Project “Restoration and Conservation of the Priority Habitat Type *9562 Grecian Juniper Woods in Prespa National Park, Greece” LIFE12 NAT/GR/539. Available online: http://www.junex.gr/index.php/en/ (accessed on 13 March 2021).
- ForOpenForests. LIFE+ Nature Project “Conservation of Priority Forests and Forest Openings in “Ethnikos Drymos Oitis” and “Oros Kallidromo” of Sterea Ellada” LIFE11 NAT/GR/1014. Available online: https://www.foropenforests.org/en (accessed on 13 March 2021).
- Tenerife, J. Life+ Nature Project Restauración de los Bosques Endémicos de Juniperus spp. en Tenerife LIFE04/NAT/ES/000064. Available online: https://www.tenerife.es/life/index.htm (accessed on 13 March 2021).
- Junipercy. LIFE+ Nature Project “Improving the Conservation Status of the Priority Habitat Type 9560* (Endemic Forests with Juniperus spp.) in Cyprus” LIFE10 NAT/CY/000717. Available online: https://ec.europa.eu/environment/life/project/Projects/index.cfm?fuseaction=search.dspPage&n_proj_id=4069 (accessed on 13 March 2021).
- Al Haddabi, L.; Victor, R. The ecological status of juniper woodlands in Al Jabal Al Akhdar, northern mountains of Oman. Int. J. Environ. Stud. 2016, 73, 746–759. [Google Scholar] [CrossRef]
- MacLaren, C.A. Climate change drives decline of Juniperus seravschanica in Oman. J. Arid Environ. 2016, 128, 91–100. [Google Scholar] [CrossRef]
- Tsopelas, P.; Barnes, I.; Wingfield, M.; Xenopoulos, S. Seiridium cardinale on Juniperus species in Greece. For. Pathol. 2007, 37, 338–347. [Google Scholar] [CrossRef]
- Eastwood, A.; Lazkov, G.; Newton, A. The Red List of Trees of Central Asia; Fauna and Flora International: Campridge, UK, 2009; p. 31. [Google Scholar]
- Patzelt, A. Photographic Field Guide to the Plants of the Western Hajar Mountains, Sultanate of Oman, with a Complete Checklist of Vascular Plant Species; Sultan Qaboos University, Academic Publication and Outreach Departmment: Muscat, Oman, 2015; p. 339. [Google Scholar]
- Otto, R.; Barone, R.; Delgado, J.-D.; Arévalo, J.-R.; Garzón-Machado, V.; Cabrera-Rodríguez, F.; Fernández-Palacios, J.-M. Diversity and distribution of the last remnants of endemic juniper woodlands on Tenerife, Canary Islands. Biodivers. Conserv. 2012, 21, 1811–1834. [Google Scholar] [CrossRef]
- Sarangzai, A.M.; Ahmed, M.; Ahmed, A.; Tareen, L.; Jan, S.U. The ecology and dynamics of Juniperus excelsa forest in Balochistan-Pakistan. Pak. J. Bot. 2012, 44, 1617–1625. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The Threat of the Combined Effect of Biotic and Abiotic Stress Factors in Forestry Under a Changing Climate. Front. Plant Sci. 2020, 11, 1874. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.g.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- de Vries, S.M.; Alan, M.; Bozzano, M.; Burianek, V.; Collin, E.; Cottrell, J.; Ivankovic, M.; Kelleher, C.; Koskela, J.; Rotach, P. Pan-European strategy for genetic conservation of forest trees and establishment of a core network of dynamic conservation units. Eur. For. Genet. Resour. Programme 2015, 7, 3. [Google Scholar]
- Fisher, M.; Gardner, A.S. The status and ecology of a Juniperus excelsa subsp. polycarpos woodland in the northern mountains of Oman. Vegetatio 1995, 119, 33–51. [Google Scholar] [CrossRef]
- Hanna, S. Field Guide to the Geology of Oman. 1. Western Hajar Mountains and Musandam; Historical Association: Muscat, Oman, 1995; p. 178. [Google Scholar]
- Solomou, A.; Proutsos, N.; Karetsos, G.; Tsagari, C. Effects of climate change on vegetation in Mediterranean forests: A review. Int. J. Environ. Agric. Biotechnol. 2017, 2, 240–247. [Google Scholar] [CrossRef]
- Gruwez, R.; De Frenne, P.; De Schrijver, A.; Vangansbeke, P.; Verheyen, K. Climate warming and atmospheric deposition affect seed viability of common juniper (Juniperus communis) via their impact on the nutrient status of the plant. Ecol. Res. 2017, 32, 135–144. [Google Scholar] [CrossRef]
- MacLaren, C.A. The Juniper & Olive Survey Report, Vegetation Assessment: Part 1. In The Juniper and Olive Woodlands Project; The National Field Research Centre for Environmental Conservation, Oman Earthwatch Programme: Muscat, Oman, 2015; p. 41. [Google Scholar]
- Karetsos, G.; Solomou, A.D.; Trigas, P.; Tsagari, K. The vascular flora of Mt. Oiti National Park and the surrounding area in Greece. J. For. Sci. 2018, 64, 435–454. [Google Scholar] [CrossRef]
- Brofas, G.B.L.; Galanos, F.; Albanis, K. Management Plan for the National Park of Parnassus; Forest Research Institute of Athens: Athens, Greece, 1996. (In Greek) [Google Scholar]
- Fisher, M. Decline in the juniper woodlands of Raydah Reserve in southwestern Saudi Arabia: A response to climate changes? Glob. Ecol. Biogeogr. Lett. 1997, 379–386. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows; Version 23.0; IBM Corp.: Armonk, NY, USA, 2015; Available online: https://www.ibm.com/analytics/spss-statistics-software (accessed on 25 February 2021).
- Kendall, M.G. Rank Correlation Methods; Charles Griffin and Co. Ltd.: London, UK, 1948; p. 160. [Google Scholar]
- Mann, H.B. Nonparametric tests against trend. Econom. J. Econom. Soc. 1945, 245–259. [Google Scholar] [CrossRef]
- Helsel, D.R.; Hirsch, R.M. Statistical Methods in Water Resources; Elsevier: Amsterdam, The Netherlands, 1992; Volume 49. [Google Scholar]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Tsiros, I.X.; Nastos, P.; Proutsos, N.D.; Tsaousidis, A. Variability of the aridity index and related drought parameters in Greece using climatological data over the last century (1900–1997). Atmos. Res. 2020, 240, 104914. [Google Scholar] [CrossRef]
- Emaminasab, M.; Oladi, R.; Pourtahmasi, K.; Shirvany, A. The potential of Juniperus foetidissima Willd. tree and Juniperus oblonga MB shrub for dendroclimatology in Arasbaran forests. For. Wood Prod. 2020, 73, 353–363. [Google Scholar] [CrossRef]
- Proutsos, N.; Tigkas, D. Growth Response of Endemic Black Pine Trees to Meteorological Variations and Drought Episodes in a Mediterranean Region. Atmosphere 2020, 11, 554. [Google Scholar] [CrossRef]
- Thanos, C.; Kaltsis, A.; Koutsovoulou, K.; Skourti, E.; Sarris, D. Composition and Structure of Juniperus macrocarpa Subpopulations. Final Report. Junicoast LIFE+ Nature Project: “Actions for the Conservation of Coastal Dunes with Juniperus spp. in Crete and the South Aegean (LIFE07NAT_GR_000296)”-Action A3. 2010, p. 81. Available online: http://www.junicoast.gr/documents/0000/0048/LIFE_07_NAT_GR_000296_Deliverable_A.3rev1.pdf (accessed on 22 March 2021). (In Greek with English Executive Summary).
- Gardner, A.S.; Fisher, M. The distribution and status of the montane juniper woodlands of Oman. J. Biogeogr. 1996, 23, 791–803. [Google Scholar] [CrossRef]
- Ahmed, M.; Buzdar, A.; Shafiq, M. Pattern of change in seed characteristics in Juniperus excelsa in Baluchistan [Pakistan]. Pak. J. Agric. Res. 1989. [Google Scholar] [CrossRef]
- Chambers, J.C.; Vander Wall, S.B.; Schupp, E.W. Seed and seedling ecology of pinon and juniper species in the pygmy woodlands of western North America. Bot. Rev. 1999, 65, 1–38. [Google Scholar] [CrossRef]
- Gruwez, R.; De Frenne, P.; De Schrijver, A.; Leroux, O.; Vangansbeke, P.; Verheyen, K. Negative effects of temperature and atmospheric depositions on the seed viability of common juniper (Juniperus communis). Ann. Bot. 2014, 113, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, K.; Adriaenssens, S.; Gruwez, R.; Michalczyk, I.M.; Ward, L.K.; Rosseel, Y.; Van den Broeck, A.; Garcia, D. Juniperus communis: Victim of the combined action of climate warming and nitrogen deposition? Plant Biol. 2009, 11, 49–59. [Google Scholar] [CrossRef]
- Verheyen, K.; Schreurs, K.; Vanholen, B.; Hermy, M. Intensive management fails to promote recruitment in the last large population of Juniperus communis (L.) in Flanders (Belgium). Biol. Conserv. 2005, 124, 113–121. [Google Scholar] [CrossRef]
- García, D. Interaction between juniper Juniperus communis L. and its fruit pest insects: Pest abundance, fruit characteristics and seed viability. Acta Oecol. 1998, 19, 517–525. [Google Scholar] [CrossRef]
- Houle, G.; Babeux, P. Variations in rooting ability of cuttings and in seed characteristics of five populations of Juniperus communis var. depressa from subarctic Quebec. Can. J. Bot. 1994, 72, 493–498. [Google Scholar] [CrossRef]
- Juan, R.; Pastor, J.; Fernandez, I.; Diosdado, J.C. Seedling emergence in the endangered Juniperus oxycedrus subsp. macrocarpa (Sm.) Ball in southwest Spain. Acta Biol. Cracov. Bot 2006, 48, 49–58. [Google Scholar]
- Ward, L.K. The conservation of juniper: Longevity and old age. J. Appl. Ecol. 1982, 917–928. [Google Scholar] [CrossRef]
- Milios, E.; Pipinis, E.; Petrou, P.; Akritidou, S.; Smiris, P.; Aslanidou, M. Structure and regeneration patterns of the Juniperus excelsa Bieb. stands in the central part of the Nestos valley in the northeast of Greece, in the context of anthropogenic disturbances and nurse plant facilitation. Ecol. Res. 2007, 22, 713–723. [Google Scholar] [CrossRef]
- Quézel, P. Végétation des hautes montagnes de la Grèce méridionale. Vegetatio 1964, 289–385. [Google Scholar] [CrossRef]
- Eriksson, G.; Namkoong, G.; Roberds, J.H. Dynamic gene conservation for uncertain futures. For. Ecol. Manag. 1993, 62, 15–37. [Google Scholar] [CrossRef]
- Perrino, E.V.; Musarella, C.M.; Magazzini, P. Management of grazing Italian river buffalo to preserve habitats defined by Directive 92/43/EEC in a protected wetland area on the Mediterranean coast: Palude Frattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Calaciura, B.; Spinelli, O. Management of Natura 2000 Habitats. 5210 Arborescent Matorral with Juniperus spp. Technical Report 2008 10/24; European Commission (DG ENV B2): Brussels, Belgium, 2008; p. 25. Available online: https://ec.europa.eu/environment/nature/natura2000/management/habitats/pdf/5210_Arborescent_matorral_Juniperus.pdf (accessed on 22 March 2021).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proutsos, N.; Solomou, A.; Karetsos, G.; Tsagari, K.; Mantakas, G.; Kaoukis, K.; Bourletsikas, A.; Lyrintzis, G. The Ecological Status of Juniperus foetidissima Forest Stands in the Mt. Oiti-Natura 2000 Site in Greece. Sustainability 2021, 13, 3544. https://doi.org/10.3390/su13063544
Proutsos N, Solomou A, Karetsos G, Tsagari K, Mantakas G, Kaoukis K, Bourletsikas A, Lyrintzis G. The Ecological Status of Juniperus foetidissima Forest Stands in the Mt. Oiti-Natura 2000 Site in Greece. Sustainability. 2021; 13(6):3544. https://doi.org/10.3390/su13063544
Chicago/Turabian StyleProutsos, Nikolaos, Alexandra Solomou, George Karetsos, Konstantinia Tsagari, George Mantakas, Konstantinos Kaoukis, Athanassios Bourletsikas, and George Lyrintzis. 2021. "The Ecological Status of Juniperus foetidissima Forest Stands in the Mt. Oiti-Natura 2000 Site in Greece" Sustainability 13, no. 6: 3544. https://doi.org/10.3390/su13063544
APA StyleProutsos, N., Solomou, A., Karetsos, G., Tsagari, K., Mantakas, G., Kaoukis, K., Bourletsikas, A., & Lyrintzis, G. (2021). The Ecological Status of Juniperus foetidissima Forest Stands in the Mt. Oiti-Natura 2000 Site in Greece. Sustainability, 13(6), 3544. https://doi.org/10.3390/su13063544