
Increased Likelihood of High Nitrous Oxide (N2O) Exchange in Soils at Reduced Microbial Diversity


Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Preparation
2.2. Experimental Setup
2.3. CO2 and N2O Measurements
2.4. Statistics
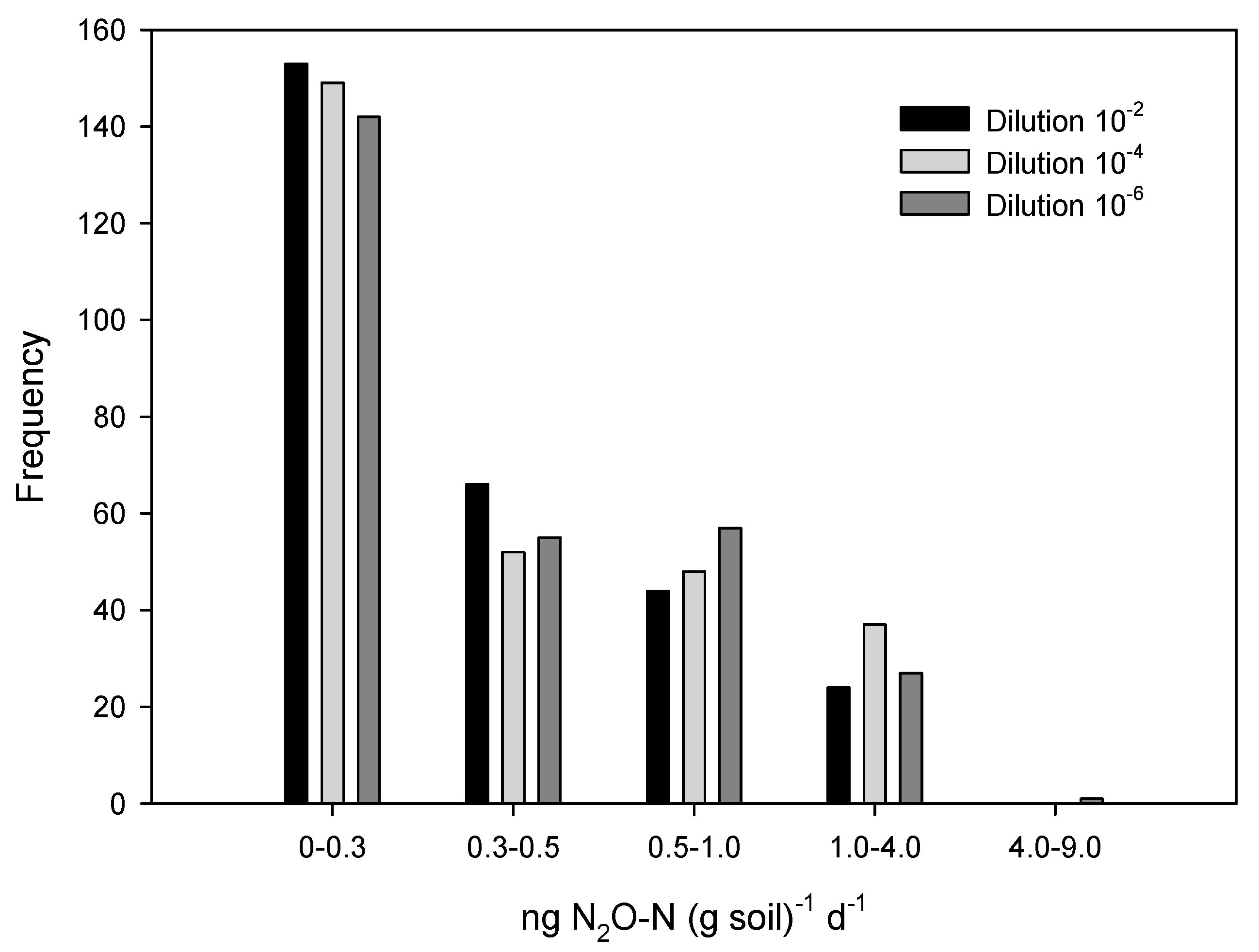
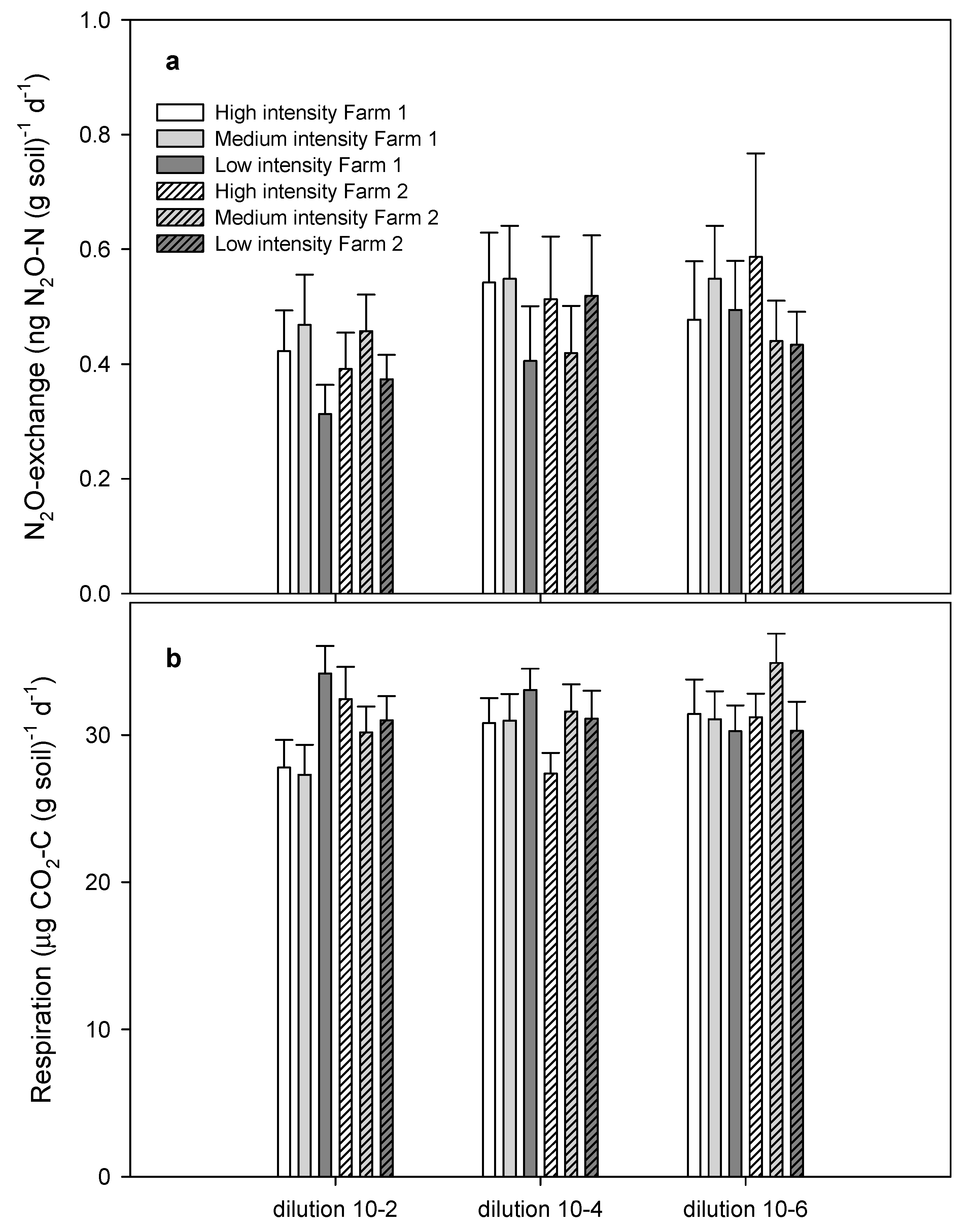
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mi, L.; Wang, G.; Jin, J.; Sui, Y.; Liu, J.; Liu, X. Comparison of microbial community structures in four Black soils along a climatic gradient in northeast China. Can. J. Soil Sci. 2012, 92, 543–549. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Hol, W.H.G.; de Boer, W.; Termorshuizen, A.J.; Meyer, K.M.; Schneider, J.H.M.; van Dam, N.M.; van Veen, J.A.; van der Putten, W.H. Reduction of rare soil microbes modifies plant-herbivore interactions. Ecol. Lett. 2010, 13, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Kusel, K.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef]
- Tiedje, J.M. Ecology of denitrification and dissimilatory nitrate reduction to ammonium. In Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John Wiley and Sons: New York, NY, USA, 1988; pp. 179–244. [Google Scholar]
- Smith, M.S.; Tiedje, J.M. Phases of denitrification following oxygen depletion in soil. Soil Biol. Biochem. 1979, 11, 261–267. [Google Scholar] [CrossRef]
- Aboobakar, A.; Cartmell, E.; Stephenson, T.; Jones, M.; Vale, P.; Dotro, G. Nitrous oxide emissions and dissolved oxygen profiling in a full-scale nitrifying activated sludge treatment plant. Water Res. 2013, 47, 524–534. [Google Scholar] [CrossRef]
- Goreau, T.J.; Kaplan, W.A.; Wofsy, S.C.; McElroy, M.B.; Valois, F.W.; Watson, S.W. Production of NO2- and N2O by nitrifying bacteria at reduced concentrations of oxygen. Appl. Environ. Microbiol. 1980, 40, 526–532. [Google Scholar] [CrossRef]
- Groffman, P.M.; Butterbach-Bahl, K.; Fulweiler, R.W.; Gold, A.J.; Morse, J.L.; Stander, E.K.; Tague, C.; Tonitto, C.; Vidon, P. Challenges to incorporating spatially and temporally explicit phenomena (hotspots and hot moments) in denitrification models. Biogeochemistry 2009, 93, 49–77. [Google Scholar] [CrossRef]
- Sexstone, A.J.; Revsbech, N.P.; Parkin, T.B.; Tiedje, J.M. Direct measurement of oxygen profiles and denitrification rates in soil aggregates. Soil Sci. Soc. Am. J. 1985, 49, 645–651. [Google Scholar] [CrossRef]
- Graf, D.R.H.; Zhao, M.; Jones, C.M.; Hallin, S. Soil type overrides plant effect on genetic and enzymatic N2O production potential in arable soils. Soil Biol. Biochem. 2016, 100, 125–128. [Google Scholar] [CrossRef]
- Christensen, S.; Simkins, S.; Tiedje, J.M. Temporal patterns of soil denitrification: Their stability and causes. Soil Sci. Soc. Am. J. 1990, 54, 1614–1618. [Google Scholar] [CrossRef]
- Becker, J.; Eisenhauer, N.; Scheu, S.; Jousset, A. Increasing antagonistic interactions cause bacterial communities to collapse at high diversity. Ecol. Lett. 2012, 15, 468–474. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Bakker, M.G.; Bradeen, J.M.; Kinkel, L.L. Plant community richness and microbial interactions structure bacterial communities in soil. Ecology 2015, 96, 134–142. [Google Scholar] [CrossRef] [PubMed]
- van Elsas, J.D.; Hill, P.; Chroňáková, A.; Grekova, M.; Topalova, Y.; Elhottová, D.; Krištůfek, V. Survival of genetically marked Escherichia coli O157: H7 in soil as affected by soil microbial community shifts. ISME J. 2007, 1, 204–214. [Google Scholar] [CrossRef]
- Liu, M.; Bjørnlund, L.; Rønn, R.; Christensen, S.; Ekelund, F. Disturbance promotes non-indigenous bacterial invasion in soil microcosms: Analysis of the roles of resource availability and community structure. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B. 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Salonius, P.O. Metabolic capabilities of forest soil microbial-populations with reduced species-diversity. Soil Biol. Biochem. 1981, 13, 1–10. [Google Scholar] [CrossRef]
- Hol, W.H.G.; de Boer, W.; de Hollander, M.; Kuramae, E.E.; Meisner, A.; van der Putten, W.H. Context dependency and saturating effects of loss of rare soil microbes on plant productivity. Front. Plant Sci. 2015, 6, 485. [Google Scholar] [CrossRef]
- Carvalho, S.; van der Putten, W.H.; Hol, W.H.G. The potential of hyperspectral patterns of winter wheat to detect changes in soil microbial community composition. Front. Plant Sci. 2016, 7, 759. [Google Scholar] [CrossRef]
- de Vries, F.T.; Thebault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Jørgensen, H.B.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef]
- Philippot, L.; Spor, A.; Henault, C.; Bru, D.; Bizouard, F.; Jones, C.M.; Sarr, A.; Maron, P.A. Loss in microbial diversity affects nitrogen cycling in soil. ISME J. 2013, 7, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Kelsic, E.D.; Zhao, J.; Vetsigian, K.; Kishony, R. Counteraction of antibiotic production and degradation stabilizes microbial communities. Nature 2015, 521, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition interrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbies, K.E.; Massheder, A.R. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2018, 69, 10–20. [Google Scholar] [CrossRef]
- van Groeningen, J.W.; Velthof, G.L.; Oenema, O.; van Groeningen, K.J.; van Kessel, C. Towards an agronomic assessment of N2O emissions: A case study for arable crops. Eur. J. Soil Sci. 2010, 61, 903–913. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Dilution | 10−2 | 10−4 | 10−6 |
|---|---|---|---|
| n | 287 | 286 | 282 |
| Mean | 0.40 | 0.49 | 0.49 |
| Skewness | 2.44 | 3.30 | 6.14 |
| Effect | F-Value | p-Value |
|---|---|---|
| Dilution | 3.06 | 0.08 |
| Farm | 0.21 | 0.64 |
| Cultivation intensity | 0.54 | 0.58 |
| Dilution × farm | 0.59 | 0.44 |
| Dilution × cultivation intensity | 0.23 | 0.79 |
| Farm × cultivation intensity | 0.50 | 0.60 |
| Dilution × farm × cultivation intensity | 0.70 | 0.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christensen, S.; Hol, W.H.G.; Kurm, V.; Vestergård, M. Increased Likelihood of High Nitrous Oxide (N2O) Exchange in Soils at Reduced Microbial Diversity. Sustainability 2021, 13, 1685. https://doi.org/10.3390/su13041685
Christensen S, Hol WHG, Kurm V, Vestergård M. Increased Likelihood of High Nitrous Oxide (N2O) Exchange in Soils at Reduced Microbial Diversity. Sustainability. 2021; 13(4):1685. https://doi.org/10.3390/su13041685
Chicago/Turabian StyleChristensen, Søren, Wilhelmina H. Gera Hol, Viola Kurm, and Mette Vestergård. 2021. "Increased Likelihood of High Nitrous Oxide (N2O) Exchange in Soils at Reduced Microbial Diversity" Sustainability 13, no. 4: 1685. https://doi.org/10.3390/su13041685
APA StyleChristensen, S., Hol, W. H. G., Kurm, V., & Vestergård, M. (2021). Increased Likelihood of High Nitrous Oxide (N2O) Exchange in Soils at Reduced Microbial Diversity. Sustainability, 13(4), 1685. https://doi.org/10.3390/su13041685