

Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress

 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Morphological Observations
2.3. Total Soluble Solids and pH
2.4. Heavy Metal Concentration in Carrots
2.5. Chlorophyll Contents Determination
2.6. Estimation of Antioxidants (TPC, TFC, DPPH)
2.7. Total Soluble Proteins
2.8. Statistical Analysis
3. Results
3.1. Effect of Gibberellic Acid on Carrot Growth under Pb Stress
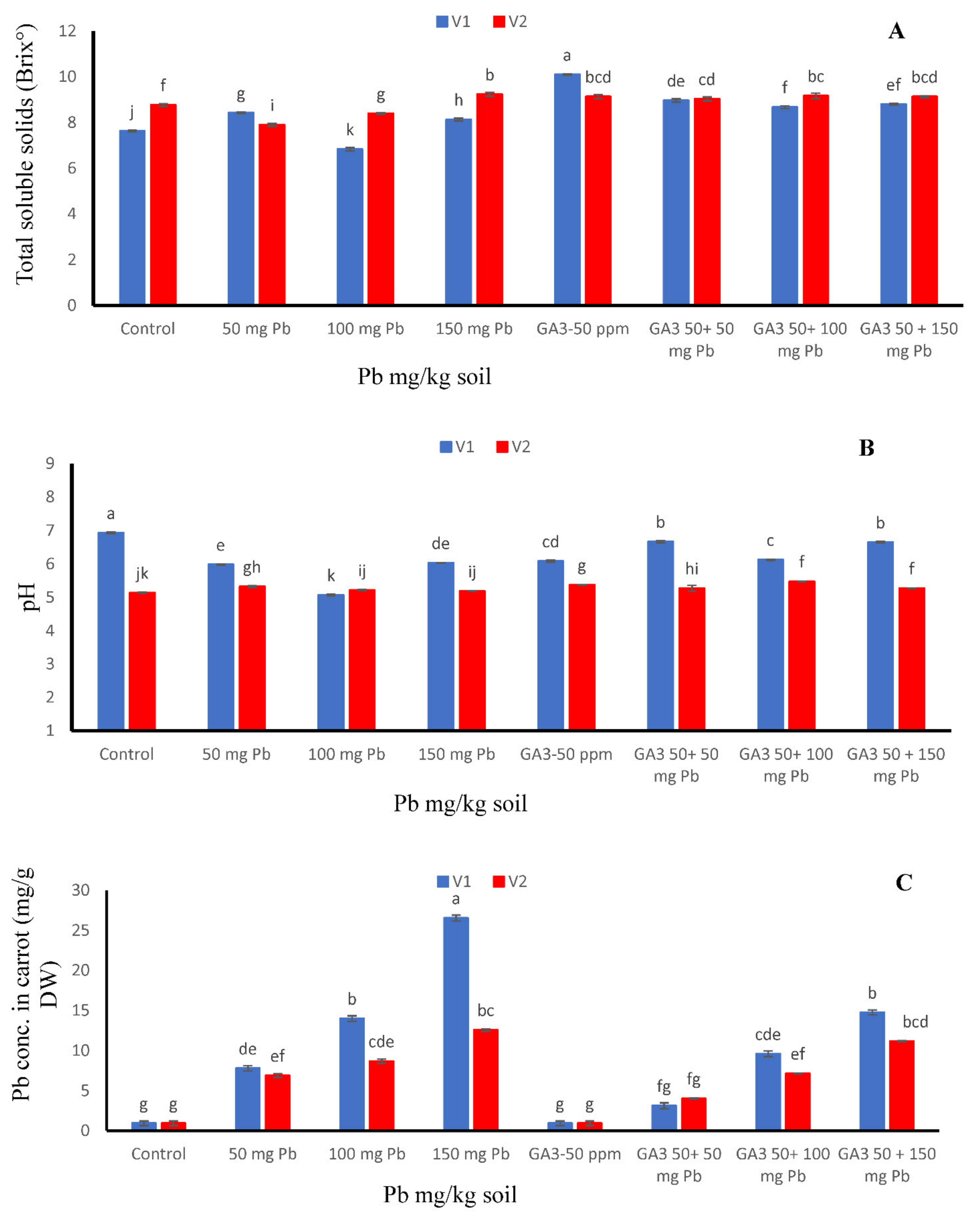
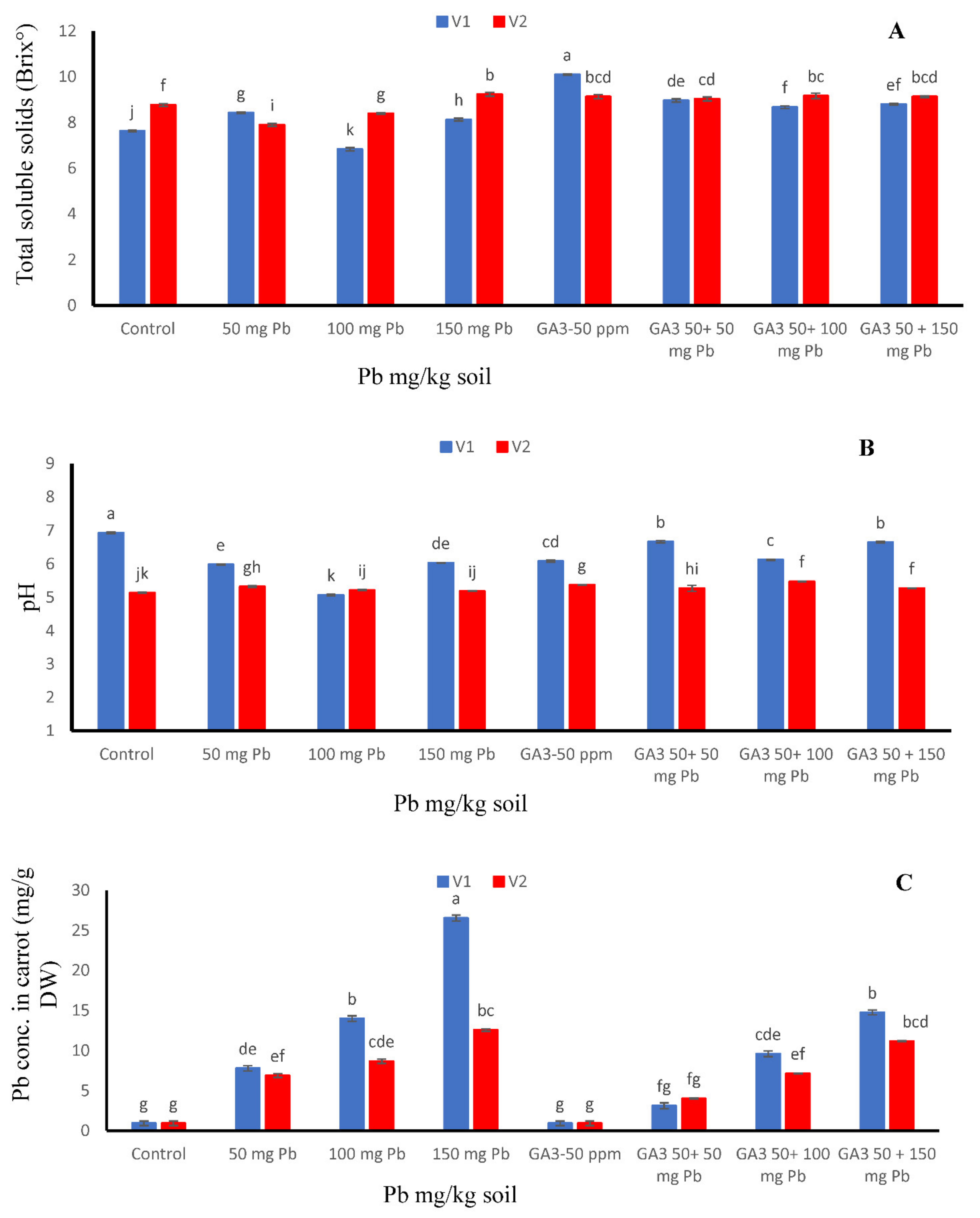
3.2. Total Soluble Solids and pH
3.3. Pb Concentration in Carrot Seedlings
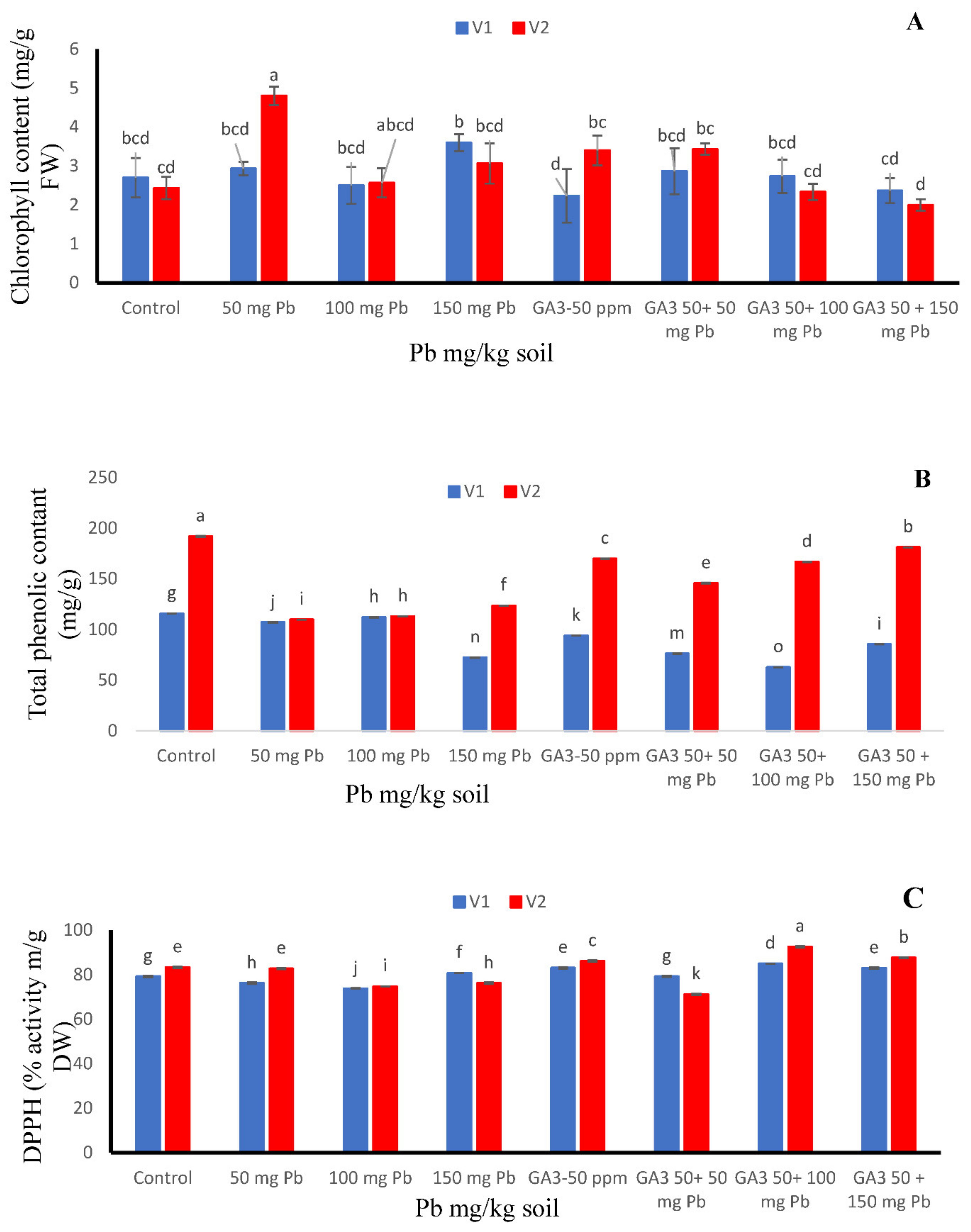
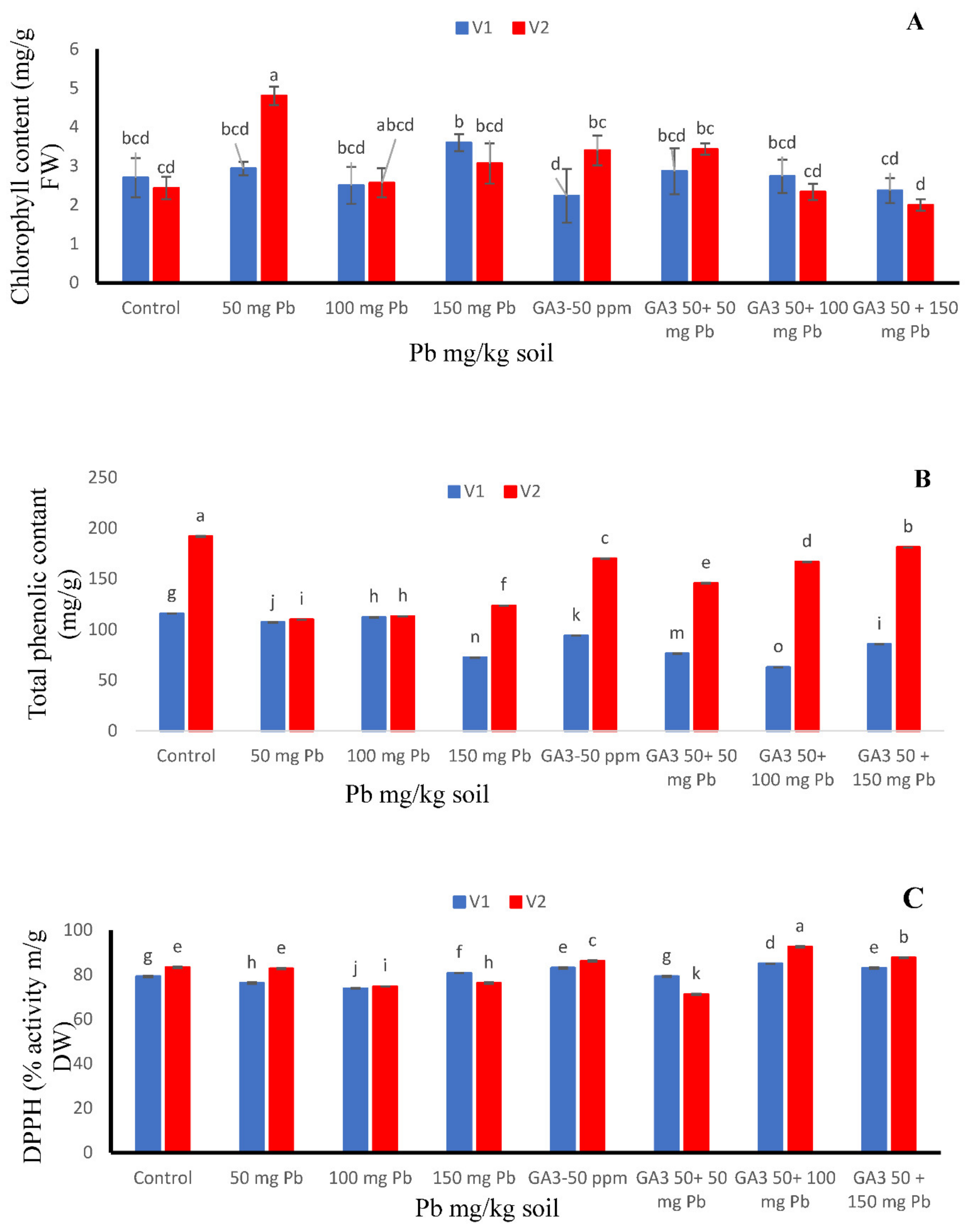
3.4. Chlorophyll Contents
3.5. Total Phenolic Contents
3.6. DPPH Radical Scavenging Activity
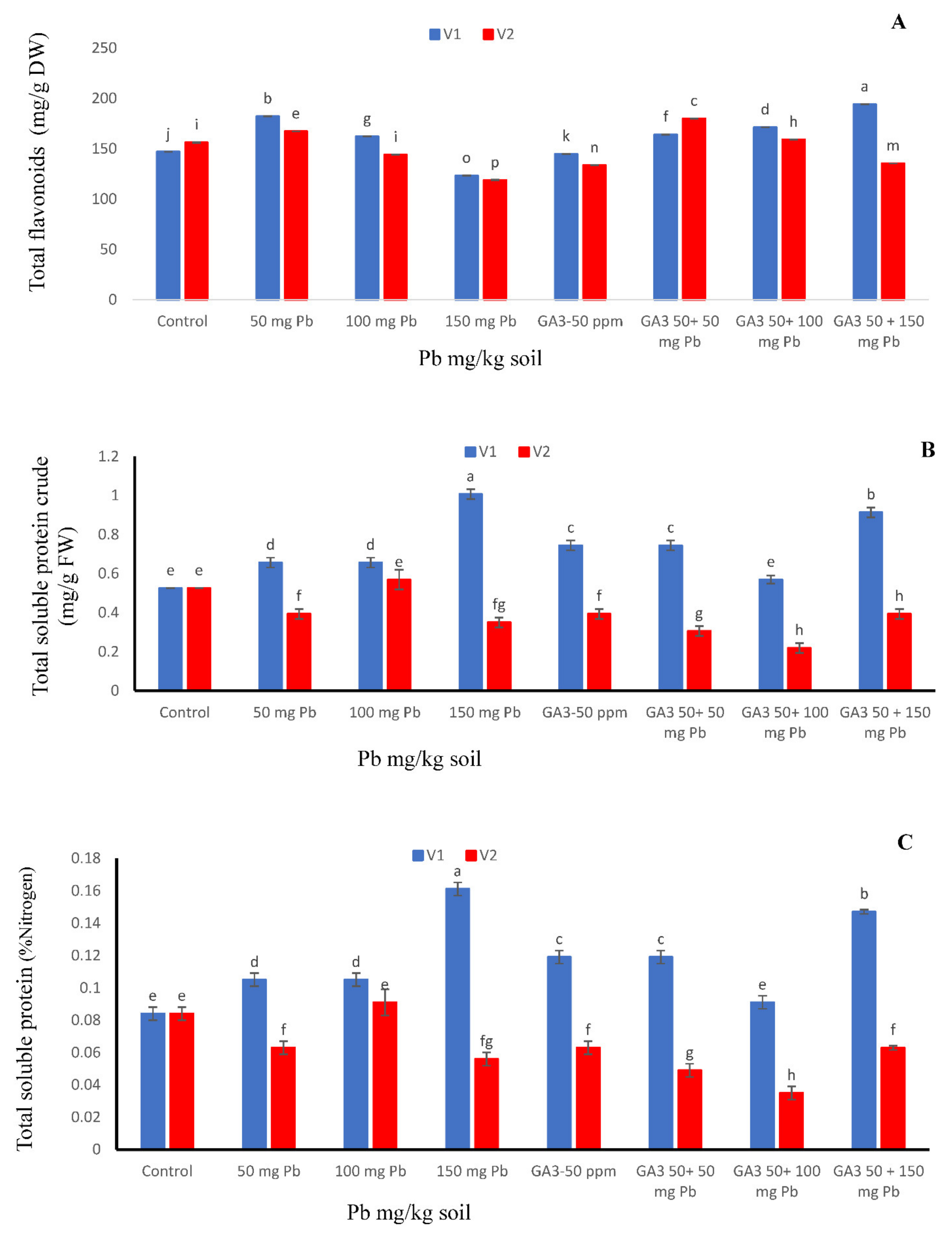
3.7. Total Flavonoid Contents
3.8. Crude Protein Percentage
3.9. Nitrogen Contents (%)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Tariq, F.; Wang, X.; Saleem, M.H.; Khan, Z.I.; Ahmad, K.; Saleem Malik, I.; Munir, M.; Mahpara, S.; Mehmood, N.; Ahmad, T.; et al. Risk Assessment of heavy metals in basmati rice: Implications for public health. Sustainability 2021, 13, 8513. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Noor, I.; El-Esawi, M.A.; Hayat, K.; Rizwan, M.; Abbas, Z.; El-Sheikh, M.A.; Alyemeni, M.N. Iron–lysine mediated alleviation of chromium toxicity in spinach (Spinacia oleracea L.) plants in relation to morpho-physiological traits and iron uptake when irrigated with tannery wastewater. Sustainability 2020, 12, 6690. [Google Scholar] [CrossRef]
- Wierzbicka, M.H.; Przedpeska, E.; Ruzik, R.; Ouerdane, L.; Poeć-Pawlak, K.; Jarosz, M.; Szpunar, J.; Szakiel, A. Comparison of the toxicity and distribution of cadmium and lead in plant cells. Protoplasma 2007, 231, 99–111. [Google Scholar] [CrossRef]
- Lăcătuşu, R.; Lăcătuşu, A.R. Vegetable and fruits quality within heavy metals polluted areas in Romania. Carpathian J. Earth Environ. Sci. 2008, 3, 115–129. [Google Scholar]
- Adriano, D.C. Arsenic. In Trace Elements in Terrestrial Environments; Springer: New York, NY, USA, 2001; pp. 219–261. [Google Scholar]
- Islam, M.S.; Ahmed, M.K.; Habibullah-Al-Mamun, M. Determination of heavy metals in fish and vegetables in Bangladesh and health implications. Human Ecol. Risk Assess. 2015, 21, 986–1006. [Google Scholar] [CrossRef]
- Saidi, I.; Chtourou, Y.; Djebali, W. Selenium alleviates cadmium toxicity by preventing oxidative stress in sunflower (Helianthus annuus) seedlings. J. Plant Physiol. 2014, 171, 85–91. [Google Scholar] [CrossRef]
- Tian, T.; Ali, B.; Qin, Y.; Malik, Z.; Gill, R.A.; Ali, S.; Zhou, W. Alleviation of lead toxicity by 5-aminolevulinic acid is related to elevated growth, photosynthesis, and suppressed ultrastructural damages in oilseed rape. BioMed. Res. Int. 2014, 2014, 530642. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Zhou, W.J. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhao, X.; Sun, X.; Tan, Q.; Tang, Y.; Nie, Z.; Qu, C.; Chen, Z.; Hu, C. Antioxidant enzyme systems and the ascorbate-glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 2015, 138, 526–536. [Google Scholar] [CrossRef]
- Liu, J.; Qian, M.; Cai, G.; Yang, J.; Zhu, Q. Uptake and translocation of Cd in different rice cultivars and the relation with Cd accumulation in rice grain. J. Hazard. Mater. 2007, 143, 443–447. [Google Scholar] [CrossRef]
- Santner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; He, Q.; Wang, M.; Gao, X.; Chen, J.; Shen, C. Exogenous indole acetic acid alleviates Cd toxicity in tea (Camellia sinensis). Ecotoxicol. Environ. Saf. 2020, 190, 110090. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.; Xia, X.; Li, X.; Shi, K.; Yu, J.; Zhou, Y. Role of brassinosteroid in plant adaptation to abiotic stresses and its interplay with other hormones. Curr. Protein Pept. Sci. 2014, 16, 462–473. [Google Scholar] [CrossRef]
- Dai, H.; Wei, S.; Pogrzeba, M.; Rusinowski, S.; Krzyżak, J.; Jia, G. Exogenous jasmonic acid decreased Cu accumulation by alfalfa and improved its photosynthetic pigments and antioxidant system. Ecotoxicol. Environ. Saf. 2020, 190, 110176. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Kumar, P.P. Regulation of seed germination and abiotic stresses by gibberellins and abscisic acid. Front. Plant Sci. 2018, 20, 838. [Google Scholar] [CrossRef]
- Jung, H.; Jo, S.H.; Jung, W.Y.; Park, H.J.; Lee, A.; Moon, J.S.; Seong, S.Y.; Kim, J.K.; Kim, Y.S.; Cho, H.S. Gibberellin Promotes Bolting and Flowering via the Floral Integrators RsFT and RsSOC1-1 under Marginal Vernalization in Radish. Plants 2020, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Akand, M.H.; Hossain, M.I.; Sarkar, M.D.; Ullah, A. Effect of Nitrogen on the Growth and Yield of Carrot (Daucus carota L.). Agriculturists 2013, 11, 76–81. [Google Scholar] [CrossRef]
- Abbas, E.D. Effect of GA3 on growth and some physiological characterizes in carrot plant (Daucus carota L.). Ibn AL-Haitham J. Pure Appl. Sci. 2011, 24, 33–39. [Google Scholar]
- Uwah, E.I.; Gimba, M.S.; Gwaski, P.A. Determination of Zn, Mn, Fe and Cu in Spinach and Lettuce Cultivated in Potiskum, Yobe State, Nigeria. J. Agric. Econ. Dev. 2012, 1, 69–74. [Google Scholar]
- Liebert, M.; Licht, U.; Böhm, V.; Bitsch, R. Antioxidant properties and total phenolics content of green and black tea under different brewing conditions. Z. Lebensm. Und-Forsch. A 1999, 208, 217–220. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Chatatikun, M.; Chiabchalard, A. Phytochemical screening and free radical scavenging activities of orange baby carrot and carrot (Daucus carota Linn.) root crude extracts. J. Chem. Pharma. Res. 2013, 5, 97–102. [Google Scholar]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar]
- Ding, C.F.; Zhang, T.L.; Wang, X.X.; Zhou, F.; Yang, Y.R.; Yin, Y. Effects of soil type and genotype on lead concentration in rootstalk vegetables and the selection of cultivars for food safety. J. Environ. Manag. 2013, 122, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.; Nunez, J.J.; Davis, R.M. Influence of gibberellic acid on carrot growth and severity of Alternaria leaf blight. Plant Dis. 2000, 84, 555–558. [Google Scholar] [CrossRef]
- Ali, B.; Mwamba, T.M.; Gill, R.A.; Yang, C.; Ali, S.; Daud, M.K.; Wu, Y.; Zhou, W.J. Improvement of element uptake and antioxidative defense in Brassica napus under lead stress by application of hydrogen sulfide. Plant Growth Regul. 2014, 74, 261–273. [Google Scholar] [CrossRef]
- Skrebsky, E.C.; Tabald, L.A.; Pereira, B.; Rauber, R.; Maldaner, J.; Cargnelutti, D.; Gonçalves, J.F.; Castro, G.Y.; Shetinger, M.R.C.; Nicoloso, F.T. Effect of cadmium on growth, micronutrient concentration, and d-aminolevulinic acid dehydratase and acid phosphatase activities in plants of Pfaffia glomerata. Braz. J. Plant Physiol. 2008, 20, 285–294. [Google Scholar] [CrossRef]
- Iqbal, N.; Nazar, R.; Khan, M.I.R.; Masood, A.; Khan, N.A. Role of gibberellins in regulation of source–sink relations under optimal and limiting environmental conditions. Curr. Sci. 2011, 7, 998–1007. [Google Scholar]
- Hadi, F.; Bano, A.; Fuller, M.P. The improved phytoextraction of lead (Pb) and the growth of maize (Zea mays L.): The role of plant growth regulators (GA3 and IAA) and EDTA alone and in combinations. Chemosphere 2010, 80, 457–462. [Google Scholar] [CrossRef]
- Lima, F.D.S.; do Nascimento, C.W.; da Silva, F.B.; de Carvalho, V.G.; Ribeiro Filho, M.R. Lead concentration and allocation in vegetable crops grown in a soil contaminated by battery residues. Hortic. Bras. 2009, 27, 362–365. [Google Scholar] [CrossRef]
- Ghani, M.A.; Abbas, M.M.; Amjad, M.; Ziaf, K.; Ali, B.; Shaheen, T.; Awan, F.S.; Khan, A.N. Production and characterization of tomato derived from Interspecific hybridization between cultivated tomato and its wild relatives. J. Hortic. Sci. Biotechnol. 2020, 95, 506–520. [Google Scholar] [CrossRef]
- Rokaya, P.R.; Baral, D.R.; Gautam, D.M.; Shrestha, A.K.; Paudyal, K.P. Effect of Pre-harvest application of gibberellic acid on fruit quality and shelf life of mandarin. J. Plant Sci. 2016, 7, 1033–1039. [Google Scholar] [CrossRef]
- Fairbrother, A.; Wenstel, R.; Sappington, K.; Wood, W. Framework for metals risk assessment. Ecotoxicol. Environ. Saf. 2007, 68, 145–227. [Google Scholar] [CrossRef]
- Ali, M.H.; Al-Qahtani, K.M. Assessment of some heavy metals in vegetables, cereals and fruits in Saudi Arabian markets. Egy. J. Aquat. Res. 2012, 38, 31–37. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M.; Słaba, M.; Mazur, J. Effect of nickel on antioxidative enzyme activities, praline and chlorophyll contents in wheat shoots. Biol. Plant 2006, 50, 653–659. [Google Scholar] [CrossRef]
- Singh, N.; Ma, L.Q.; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pterisensi formis L. Plant Sci. 2006, 170, 274–282. [Google Scholar] [CrossRef]
- Zhu, X.F.; Jiang, T.; Wang, Z.W.; Lei, G.J.; Shi, Y.Z.; Li, G.X.; Zheng, S.J. Gibberellic acid alleviates cadmium toxicity by reducing nitric oxide accumulation and expression of IRT1 in Arabidopsis thaliana. J. Hazard. Mater. 2012, 239, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Wang, Z.W.; Dong, F.; Lei, G.J.; Shi, Y.Z.; Li, G.X.; Zheng, S.J. Exogenous auxin alleviates cadmium toxicity in Arabidopsis thaliana by stimulating synthesis of hemicellulose 1 and increasing the cadmium fixation capacity of root cell walls. J. Hazard. Mater. 2013, 263, 98–403. [Google Scholar] [CrossRef]
- Al-Hakimi, A.M.A. Modification of cadmium toxicity in pea seedlings by kinetin. Plant Soil Environ. 2007, 53, 129. [Google Scholar] [CrossRef]
- El-Monem, A.; Sharaf, M.M.; Farghal, I.I.; Sofy, M. Role of gibberellic acid in abolishing the detrimental effects of Cd and Pb on broad bean and lupin plants. J. Agric. Biol. Sci. 2009, 5, 6–13. [Google Scholar]
- Masood, A.; Khan, M.I.R.; Fatma, M.; Asgher, M.; Khan, N.A. Involvement of ethylene in gibberellic acid-induced sulfur assimilation, photosynthetic responses, and alleviation of cadmium stress in mustard. Plant Physiol. Biochem. 2016, 104, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fargašová, A. Phytotoxic effects of Cd, Zn, Pb, Cu and Fe on Sinapis alba L. seedlings and their accumulation in roots and shoots. Biol. Plant. 2001, 44, 471–473. [Google Scholar] [CrossRef]
- Ali, B.; Song, W.J.; Hu, W.Z.; Luo, X.N.; Gill, R.A.; Wang, J.; Zhou, W.J. Hydrogen sulfide alleviates lead-induced photosynthetic and ultrastructural changes in oilseed rape. Ecotoxicol. Environ. Saf. 2014, 102, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Kanwal, S.; Uddin, F.; Azmat, R. Phytotoxicity of Pb II. Changes in chlorophyll absorption spectrum due to toxic metal Pb stress on Phaseolus mungo and Lens culinaris. Pak. J. Biol. Sci. 2006, 9, 2062–2068. [Google Scholar] [CrossRef]
- Kaimoyo, E.; Farag, M.A.; Sumner, L.W.; Wasmann, C.; Cuello, J.L.; VanEtten, H. Sublethal levels of electric current elicit the biosynthesis of plant secondary metabolites. J. Biotechnol. Progr. 2008, 24, 377–384. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Govarthanan, M.; Kamala-Kannan, S.; Kim, S.A.; Seo, Y.S.; Park, J.H.; Oh, B.T. Synergistic effect of chelators and Herbaspirillum sp. GW103 on lead phytoextraction and its induced oxidative stress in Zea mays. Arch. Microbiol. 2016, 198, 737–742. [Google Scholar] [CrossRef]
- Li, Y.; Qin, C.X.; Gao, B.; Hu, Y.; Xu, H. Lead-resistant strain KQBT-3 inoculants of Tricholoma lobayensis Heim that enhance remediation of lead-contaminated soil. Environ. Technol. 2015, 36, 2451–2458. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Roles of root and shoot tissues in transport and accumulation of cadmium, lead, nickel, and strontium. Russ. J. Plant Physiol. 2008, 55, 1–22. [Google Scholar] [CrossRef]
- Lamhamdi, M.; Bakrim, A.; Aarab, A.; Lafont, R.; Sayah, F. Effects of lead phytotoxicity on wheat seed germination and seedling growth. Comptes Rendus Biol. 2011, 334, 118–126. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Treatments | Varieties | Root Color | Hardness | Sweetness | Bitterness | Crunchiness |
|---|---|---|---|---|---|---|
| 50 mg Pb | T-29 | Red | Hard | Less | Yes | Yes |
| Maverick | Orange | Hard | Less | Yes | Yes | |
| 100 mg Pb | T-29 | Deep Red | Soft | Sweet | No | No |
| Maverick | Orange | Soft | Sweet | No | No | |
| 150 mg Pb | T-29 | Pale Red | Soft | Sweet | No | No |
| Maverick | Orange | Soft | Sweet | No | No | |
| 50 ppm GA3 | T-29 | Pale Red | Hard | Less | Yes | Yes |
| Maverick | Pale Orange | Soft | Less | No | Fewer | |
| 50 mg Pb + 50 ppm GA3 | T-29 | Red | Soft | Less | Yes | No |
| Maverick | Orange | Soft | Less | No | No | |
| 100 mg Pb + 50 ppm GA3 | T-29 | Red | Soft | Sweet | No | No |
| Maverick | Deep Orange | Soft | Sweet | No | No | |
| 150 mg Pb + 50 ppm GA3 | T-29 | Pale Red | Hard | Less | Yes | Yes |
| Maverick | Orange | Hard | Less | No | Yes | |
| Control | T-29 | Deep Red | Soft | Sweet | No | Fewer |
| Maverick | Deep Orange | Soft | Sweet | No | Fewer |
| Treatments | Varieties | Root Length (cm) | Root Diameter (cm) | Plant Height (cm) | Plant Fresh Weight (g) |
|---|---|---|---|---|---|
| 50 mg Pb | T-29 | 17.44 ± 1.36 cdef | 27.03 ± 1.15 ab | 45.00 ± 1.73 c | 150 ± 23.03 a |
| Maverick | 20.28 ± 0.46 abcd | 21.10 ± 1.10 defg | 19.39 ± 1.02 h | 80.53 ± 11.81 def | |
| 100 mg Pb | T-29 | 16.00 ± 0.67 ef | 25.34 ± 0.31 abc | 36.90 ±2.33 d | 86.93 ± 8.67 bcdef |
| Maverick | 15.40 ± 0.768 f | 19.18 ± 0.54 fgh | 20.44 ± 0.52 h | 55.6 ± 8.22 ef | |
| 150 mg Pb | T-29 | 17.72 ± 1.77 cdef | 22.90 ± 1.26 cde | 46.00 ±1.78 c | 109.2 ±12.83 abcd |
| Maverick | 18.00 ± 0.78 bcdef | 18.40 ± 0.75 ghi | 21.61 ± 0.86 gh | 49.53 ± 4.03 ef | |
| 50 ppm GA3 | T-29 | 21.00 ± 0.83 abc | 23.46 ± 2.05 cde | 46.22 ± 0.61 c | 134.93 ± 10.04 ab |
| Maverick | 20.05 ± 0.73 abcd | 22.30 ± 1.11 cde | 28.44 ± 1.02 ef | 85.6 ± 13.31 cdef | |
| 50 mg Pb + 50 ppm GA3 | T-29 | 19.40 ± 2.51 abcde | 24.10 ± 1.3 bcd | 45.60 ± 1.16 c | 87.07 ± 21.00 bcdef |
| Maverick | 16.89 ± 2.19 def | 20.57 ± 1.12 efg | 25.90 ± 0.77 efg | 48.73 ± 7.28 ef | |
| 100 mg Pb + 50 ppm GA3 | T-29 | 21.80 ± 1.44 ab | 23.70 ± 0.25 cd | 47.60 ±2.56 bc | 90.93 ± 9.86 bcde |
| Maverick | 15.33 ± 0.76 f | 15.80 ± 0.42 i | 29.90 ± 0.86 e | 43.77 ± 9.08 ef | |
| 150 mg Pb + 50 ppm GA3 | T-29 | 22.90 ± 2.12 a | 21.53 ± 1.51 def | 54.90 3.69 a | 129.2 ± 17.8 abc |
| Maverick | 16.66 ± 1.15 def | 16.9 ± 0.64 hi | 27.44 ± 0.97 ef | 41.6 ± 6.05 f | |
| Control | T-29 | 17.80 ± 0.48 cdef | 27.87 ± 0.96 a | 52.70 ±4.16 ab | 149.57 ± 47.75 a |
| Maverick | 17.44 ± 1.81 cdef | 16.84 ± 0.93 hi | 24.11 ± 1.41 fgh | 39.67 ± 3.71 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghani, M.A.; Abbas, M.M.; Ali, B.; Aziz, R.; Qadri, R.W.K.; Noor, A.; Azam, M.; Bahzad, S.; Saleem, M.H.; Abualreesh, M.H.; et al. Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress. Sustainability 2021, 13, 12329. https://doi.org/10.3390/su132112329
Ghani MA, Abbas MM, Ali B, Aziz R, Qadri RWK, Noor A, Azam M, Bahzad S, Saleem MH, Abualreesh MH, et al. Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress. Sustainability. 2021; 13(21):12329. https://doi.org/10.3390/su132112329
Chicago/Turabian StyleGhani, Muhammad Awais, Muhammad Mehran Abbas, Basharat Ali, Rukhsanda Aziz, Rashad Waseem Khan Qadri, Anam Noor, Muhammad Azam, Saleem Bahzad, Muhammad Hamzah Saleem, Muyassar H. Abualreesh, and et al. 2021. "Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress" Sustainability 13, no. 21: 12329. https://doi.org/10.3390/su132112329
APA StyleGhani, M. A., Abbas, M. M., Ali, B., Aziz, R., Qadri, R. W. K., Noor, A., Azam, M., Bahzad, S., Saleem, M. H., Abualreesh, M. H., Alatawi, A., & Ali, S. (2021). Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress. Sustainability, 13(21), 12329. https://doi.org/10.3390/su132112329